
Abstract
Regarding both the neural crest origin and neuronal potential of stem cells from human exfoliated deciduous teeth (SHED), here, we assessed their potential in addition to neural induced SHED (iSHED) for functional recovery when transplanted in a rat model for acute contused spinal cord injury (SCI). Following transplantation, a significant functional recovery was observed in both groups relative to the vehicle and control groups as determined by the open field locomotor functional test. We also observed that animals that received iSHED were in a better state as compared with the SHED group. Immunohistofluorescence evaluation 5 weeks after transplantation showed neuronal and glial differentiation and limited proliferation in both groups. However, myelin basic protein and chondroitin sulfate proteoglycan NG2—oligodendrocyte markers—were increased and glial fibrillary acidic protein—astrocyte marker—was decreased in the iSHED group in comparison with the SHED group. These findings have demonstrated that transplantation of SHED or its derivatives could be a suitable candidate for the treatment of SCI as well as other neuronal degenerative diseases.
Introduction
R
Dental stem cells are expanded in vitro and display multidifferentiation potential, with the capacity to give rise to osteogenic, odontogenic, chondrogenic, adipogenic, and neurogenic cell lineages and endothelial cells [9 –16]. However, the precise relationship among these different human dental tissue–derived MSC-like populations remains unclear [9,10] and they even have different differentiation potentials under the same cell culture conditions [17]. For example, in a standard cell culture medium, SHED expressed the neural stem cell marker Pax6, but DFPCs did not express it [17].
It is interesting to note that SHED cells naturally express neural cell markers which perhaps are related to their neural crest origin [18,19]. In our previous study, we have proposed a new approach for the neural induction of SHED [14]. This approach resulted in the formation of a homogenous population of neural-like cells with an extensive and notable degree of neurite extension, increased expression of neural markers such as NESTIN, neural cell adhesion molecule (NCAM), TUJ1, TH, and TAU at both the mRNA and protein levels, and which were maintained in culture for over 21 days. In this approach, initially SHED cells were exposed to basic fibroblast growth factor (bFGF) for 7 days in the absence of serum to induce cell differentiation and then were further induced to neural-like cells in the presence of bFGF and Shh [14].
Considering both the neural crest origin and neuronal potential of SHED cells, in the current study we examined the potential effect of undifferentiated and induced SHED (iSHED) cells on promoting in vivo neuronal differentiation fate and functional recovery, in contused injured spinal cords of adult rats.
Materials and Methods
SHED used for this study were prepared as previously reported [14]. Briefly, separated pulps from human exfoliated deciduous incisors were digested by collagenase type I containing 5% fetal calf serum (FCS) for 1 h at 37°C to achieve dissociated cells. After washing in medium, the cell suspension was cultured in α-MEM supplemented with 10% FCS for 3 days. The cells were subcultured by trypsin/EDTA and replated in the same medium. The cells were passaged every 3 days.
Neural induction
For neural induction, SHED were plated with a density of 5,000 cells/cm2 on poly-L-ornithine (PLO)/fibronectin (Fib)-coated dishes and cultured for 7 days in neurobasal medium (Invitrogen, 21103-049) containing 1% B27 (Invitrogen, 17504-044), 1% ITS (Invitrogen, 41400-045), and 100 ng/mL bFGF (Royan Institute) as a cytokine. The half of the medium was renewed every 3 days.
To prepare PLO/Fib-coated plates, the dishes were incubated with PLO (15 mg/mL; Sigma-Aldrich, P4957) in phosphate-buffered saline (PBS; Invitrogen, 21600-010) overnight at 37°C and, after gently washing the plates with PBS, they were incubated with Fib (50 μg/mL; Invitrogen, 33016-015), in PBS for 6 h.
Immunocytochemistry and flow cytometry analysis
For immunocytochemistry, cells were fixed with 4% paraformaldehyde (PFA; Sigma-Aldrich). After permeabilization with 0.2% Triton X-100, cells were incubated with a primary antibody (Supplementary Table S1; Supplementary Data are available online at
RNA isolation, reverse transcription, and quantitative RT-polymerase chain reaction
Total RNA of SHED and iSHED was isolated using RNeasy mini kit (Qiagen) according to the manufacturer's protocol. To remove genomic DNA contamination, all RNA samples were subjected to DNaseI (EN0521, Fermentas) treatment.
cDNA synthesis was performed using the RevertAid™ H Minus First Strand cDNA Synthesis Kit (K1632, Fermentas), 0.2 μg random hexamer primer, and 1 μg total RNA per reaction, according to the manufacturer's instructions.
Gene expression was assessed by quantitative RT-polymerase chain reaction (PCR) for candidate genes in a Rotor-Gene 6000 (Corbett Life Science) using the following program, stage 1: 95°C for 4 min, stage 2: 95°C for 10 s, 60°C for 30 s, and 72°C for 30 s, for 40 cycles. At the end of the run, melting profile was determined to demonstrate the synthesis of a single PCR product. The primers used were designed using the primer design software, Beacon Designer7 [20]. Primer sequences, expected fragment size, and Gene Bank accession numbers are listed in Supplementary Table S2.
The PCR mix in each well included 5 μL of SYBR®Premix Ex Taq™ II (RR081Q, Takara Bio., Inc.), 3 μL dH2O, 0.5 μL each of the forward and reverse primers (5 pmol/μL), and 1 μL of single strand cDNA (16 ng/μL) in a final reaction volume of 10 μL.
For relative quantification, data were analyzed using the comparative Ct method [20]. The output data from Rotor-Gene 6000 Analysis software (version 1.7, Corbett Life Science) were transferred to Microsoft Excel for analysis. For each unknown sample, the relative amount was calculated by normalization of each target gene to the reference gene beta actin (delta Ct), and then 2−ΔCt was calculated.
Surgical procedure, experimental grouping, and transplantation
SCI model
A total of 35 male Wistar rats (Pasteur Institute) with weights of ∼250–300 g were used for this study. All animal care and experimental procedures were approved according to the Institutional Review Board and the Institutional Ethical Committee. The minimum suitable number of animals was used for statistical analysis. Two rats were housed per cage with free access to food and water throughout the study. Anesthesia was performed by intraperitoneal injection with a combination of ketamine (90 mg/kg) and xylazine (9.9 mg/kg) [21].
After anesthesia, the thoracic back area was shaved and washed by povidone iodine. Following an incision through the skin, the paravertebral muscle was separated from overlying attachments at thoracic levels 8–11 (T8-T11); laminectomy was carried out at T10. After laminectomy, the vertebral column was stabilized by clamping the spinal processes, both cranial and caudal to the laminectomy area. The contusion injury was carried out by dropping a 10 g weight rod from a 25-mm height onto the dorsal surface of the spinal cord (NYU impactor). All rats, postoperatively, were hydrated with Ringer's solution (5 mL, s.c.) and kept in warmed cages until recovery. Manual bladder expression was regularly administered after surgery, 2–3 times per day for 2 weeks until bladder reflex returned to normal.
One week after injury, 2 injured spinal cords were dissected and fixed in 4% PFA and prepared for hematoxylin and eosin (H&E) staining at 10-μm thicknesses for observation of cavity formation after SCI.
Cell transplantation
Seven days after SCI, rats were randomly assigned to the following 4 groups: control group with no treatment, vehicle-transplanted group that received only the vehicle injection (5 μL PBS), SHED and iSHED groups that received 5 μL PBS of suspension containing 0.5×106 cells per animal. The respective cells were harvested by trypsin treatment and, after washing with PBS, cell suspensions were injected into the spinal cord lesions with a 25 μL sterile Hamilton syringe coupled to a 27-gauge needle through a glass capillary (inner diameter ∼50 μm) with the use of the A Kopf microstereotaxic injection system (David Kopf Instruments) at a 60 mL per hour infusion rate. After grafting, the syringe was raised 1 mm and left in place for 5 min to minimize cell diffusion up the needle track. In addition, a sample of the resulting single-cell suspension was stained with trypan blue dye exclusion for viability assessment. Finally, injured animals were given gentamycin (15 mg/kg, s.c.) and lactated Ringer's solution (5 mL, s.c.), kept on a heating pad until awake, and housed in standard rat cages at temperatures of 27°C. Starting 2 days before transplantation, all rats received daily s.c. injections of cyclosporine-A (Sandimmune; Novartis Pharmaceuticals; 10 mg/kg) to prevent cell rejection.
Behavioral assessment
Behavioral recovery in rats was assessed using an open field locomotor scale, described by Basso, Beattie and Bresnahan (BBB) [22] by 2 blinded observers for comparison of the functional outcome of all groups over time. For this assessment, 8 rats from each group were placed in an open field (75×125 cm) and observed for 5 min. Hind limb function was scored from 0 to 21 (flight paralysis to normal gait). Performance according to the behavioral task was performed 3 days before injury, prior to transplantation, and continued once a week for an additional 5 weeks after transplantation.
Sacrifice and cryosection
After behavioral analysis, all rats were given a lethal dose of anesthesia, fixed by transcardial perfusion with 400 mL of ice-cold 4% PFA (pH 7.4), followed by 0.1 M cold PBS.
Spinal cords containing injured parts from each animal were dissected and again fixed with 4% PFA for 24 h. For cryoprotection, samples were kept in 30% sucrose for 96 h at 4°C and finally, longitudinal sections of 10-μm thicknesses were prepared and placed on tissue-adhering slides. These slides were placed at −80°C until further analysis.
Immunohistofluorescence staining and quantification
For double-label immunohistochemistry (IHC), sections were washed with PBS and permeabilization was carried out by 0.4% Triton X-100 in PBS (except for membranous markers). After nonspecific site blocking by 10% goat serum and 1 mg/mL BSA, sections were incubated for 2 h with primary antibodies that are as follows: Ki67, microtubule associated protein-2 (MAP2), NCAM, glial fibrillary acidic protein (GFAP), myelin basic protein (MBP), and chondroitin sulfate proteoglycan NG2, which, respectively, address grafted cells (human) versus host cells, mitotic activity, as well as neuronal, astrocytic, and oligodendrocytic phenotypes. Surplus antibodies were removed by washing and slides were incubated with the appropriate FITC- and TRITC-conjugated secondary antibodies for 1 h at 37°C and nuclear staining was carried out by DAPI. Normal IgG from species with originating from the host primary antibodies served as negative controls. Antibodies implemented for IHC are shown in Supplementary Table S1. A fluorescent microscope (Olympus BX51) equipped with an Olympus DP70 camera was used to count the number of HNu/DAPI-positive cells that were double labeled with different cell markers from 4 rats (n=4).
Statistical analysis
Quantitative data were expressed as mean±standard deviation. All quantitative RT-PCR (qRT-PCR) experiments were conducted in at least 3 independent cultures. Real-time data were analyzed by t-test. One-way analysis of variance followed by Tukey's post hoc was used to compare the BBB scores of all groups. Independent t-test was applied to determine statistical differences between primed (iSHED) and unprimed (SHED) transplantation groups. All statistical analyses were performed on standard statistics computer software (SPSS 16 for Windows).
Results
Neural induction
SHED presented a mesenchymal-like morphology (Fig. 1A) and well characterized in our previous study [14]. Upon bFGF treatment, the cytoplasm in the flat cells initially retracted toward the nucleus and formed a contracted multipolar cell body that left membranous processes that were similar to peripheral extensions (Fig. 1A). Following the fixation process, cells were immunostained and NESTIN expression, a neural precursor marker, was observed (Fig. 1A). Flow cytometry analysis for NESTIN showed that over 80% of the cells were positive both before and after neural induction (Fig. 1A). Despite this similarity, a shift in the histogram of flow cytometry analysis suggested a higher expression intensity for NESTIN in the induced cells in comparison with the undifferentiated cells (Fig. 1B, P<0.05), which corresponds to the amount of NESTIN protein expressed by the cells.
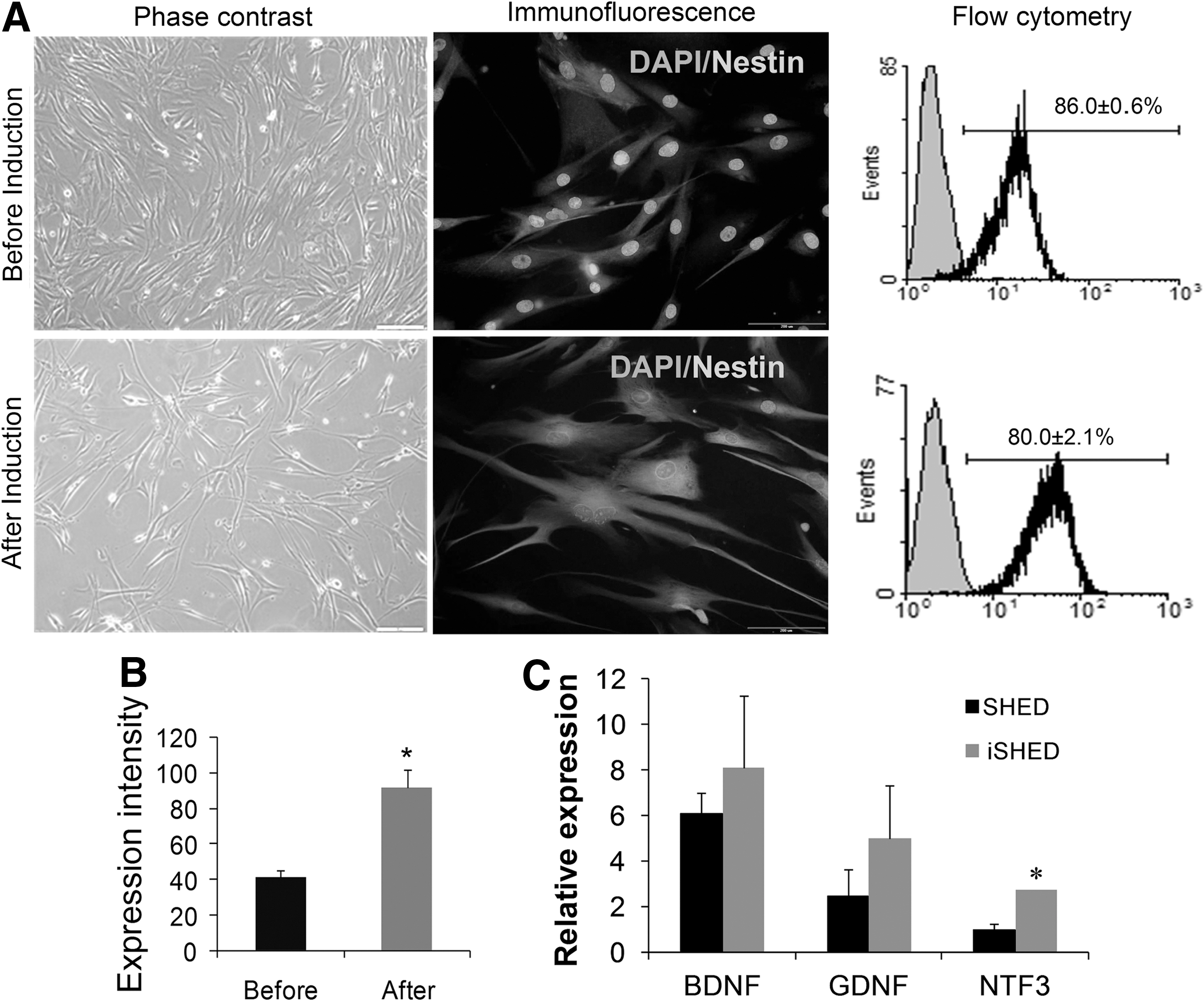
Characteristics of SHED cells before and after basic fibroblast growth factor induction for 7 days. Phase-contrast images, immunofluorescence staining, and flow cytometry of the cells
Real-time RT-PCR analysis of neurotrophins in SHED and iSHED showed the expression of brain-derived neurotrophic factor (BDNF), glial-derived neurotrophic factor (GDNF), neurotrophin 3 (NTF3) in both groups, but they were expressed more in iSHED group (Fig. 1C, NTF3: P<0.05).
Histological assessment of contusion injury
H&E staining for coronal sections showed distinct interruption of myelin homogeneity and cavity formation usually observed in injured spinal cords of rats (Fig. 2A). GFAP immunohistochemical staining of the longitudinal spinal cord sections from injured animals after 7 days revealed glial scar formation around the cavity (Fig. 2B).
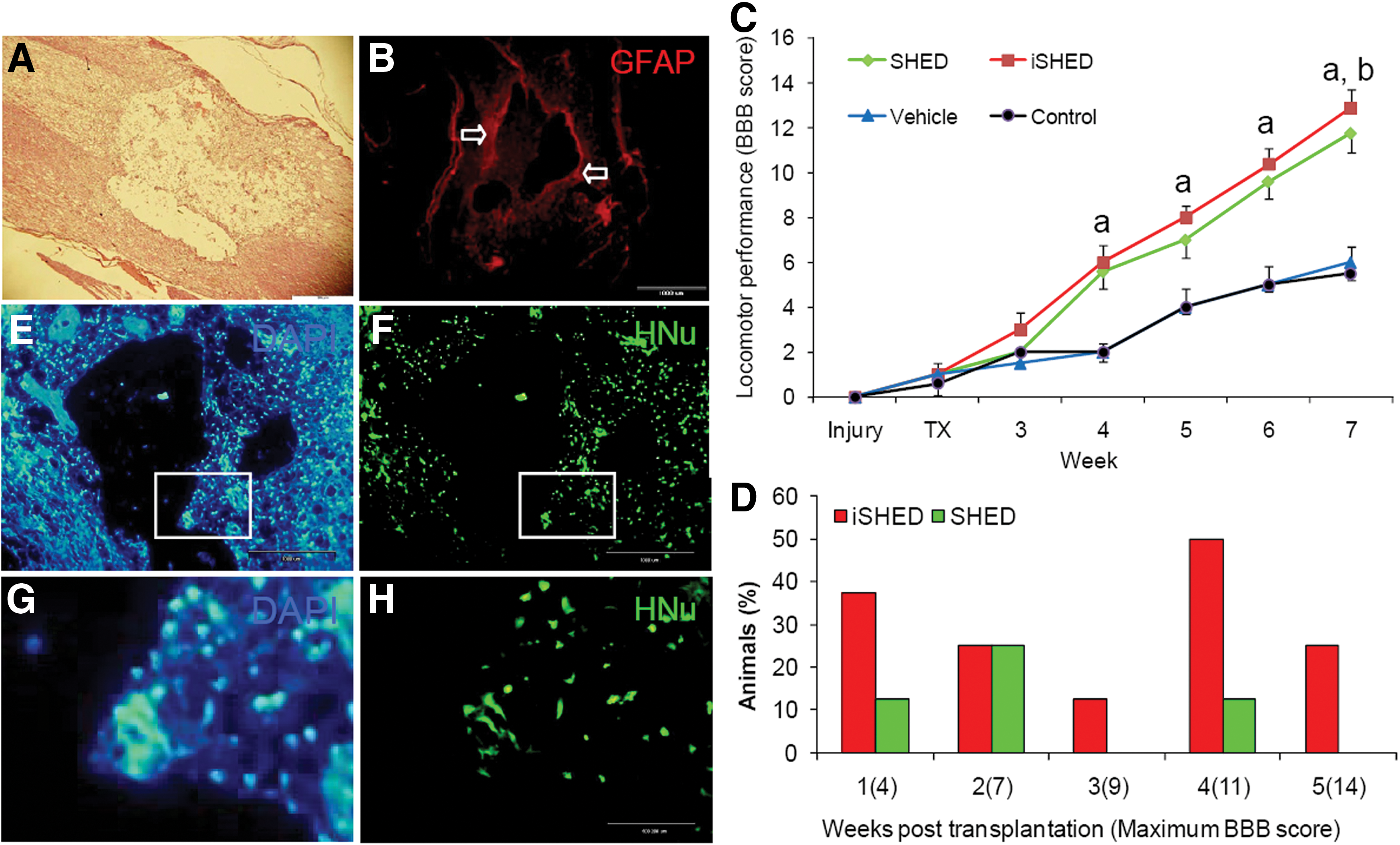
Injury, BBB score, and cell tracing after grafting in a spinal cord injury model. Hematoxylin and eosin–stained photomicrograph of lesion site representative of disintegration and interruption of spinal tissue
Functional recovery
To determine functional recovery after SCI, the BBB open-field walking test was performed before and after injury at regular intervals, up to 5 weeks after transplantation (6 weeks after injury, Fig. 2C). After the contusion injury, all rats displayed complete and flaccid paralysis of both hind limbs. Injured rats continued to recover 1 week before transplantation. To ensure that the initial BBB scores were uniform between groups, BBB was tested at day 6, 1 day prior to transplantation. Rats with BBB scores of one, indicating slight movement of 1 or 2 joints, were randomly included in the experimental groups. Figure 2G shows the mean BBB score of the experiments. In contrast to the vehicle and control groups, rats in the cell transplantation groups showed a significant improvement in function as early as the second week after transplantation and thereafter to the end point of the study (at least P<0.05). Animals in the SHED- and iSHED-transplanted groups showed almost nearly parallel recovery after transplantation for 5 weeks. The BBB recovery scores for SHED- and iSHED-transplanted groups at the end of the study were 12.87±0.82 and 11.75±0.88, respectively, while the corresponding scores in the vehicle and control groups were 6±0.75 and 5.5±0.37. Final motor recovery after 5 weeks was recorded in the iSHED-transplanted group in comparison with the SHED-transplanted group (12.87±0.82 and 11.75±0.88, respectively), which was significantly higher. In addition, 37.5% of the rats in the iSHED group showed frequent to consistent weight-supported plantar steps and frequent forelimb–hindlimb coordination 5 weeks after transplantation.
We found that a larger percentage of animals had higher BBB scores in the iSHED group compared with the SHED group. For example, 25% of the rats that received iSHED scored 14 compared with the SHED group whose scores were less 5 weeks after transplantation (Fig. 2D).
Survival and fate of transplanted cells
Transcardial perfusion was performed after the last locomotor recovery evaluation. Finally 16 mm of the spinal cord, which included the injured site, was dissected for immunohistochemical studies.
To evaluate the survival and migration of transplanted cells, longitudinal spinal cord sections from each of the grafted cell groups were assessed for HNu, a human cell tracer marker. The majority of HNu-positive cells were located around the cavity, which suggested that the grafted cells migrated in the host spinal cord and remained viable (Fig. 2E–H).
The fate of cells grafted into the spinal cord as examined by fluorescent double immunhistochemical staining for HNu and cell-specific markers are shown in Fig. 3 and Supplementary Fig. S1 (MAP2-, NCAM-, GFAP-, MBP-, and NG2-positive cells in SHED-transplanted and iSHED-transplanted groups, respectively). Quantification of positive cells revealed no significant difference in MAP2 and NCAM expression as neuronal markers between the 2 transplanted cell groups (19.6%±1.5% and 27.3%±1.5% in the SHED-transplanted group, and 20.3%±1.2% and 30%±1.6% in the iSHED-transplanted group, respectively). In contrast, significant differences (P<0.05) were seen between the 2 transplantation groups in the relative percentages of astrocyte and oligodendrocyte markers, including GFAP, MBP, and NG2 (26.0%±1.2%, 4.0%±1.0%, and 2.6%±1.0% in the SHED-transplanted group and 21.0%±1.2%, 11.0%±0.8%, and 7.0%±1.2% in the iSHED-transplanted group, respectively). In addition, some HNu-positive grafted cells in both groups remained NESTIN positive (31.0%±1.6% and 29%±2.5% of cells in the SHED-transplanted group and iSHED-transplanted group, respectively; Fig. 4). To examine cell division of grafted cells in vivo, the sections were double stained with HNu and anti-Ki67. Moreover, <3% of the transplanted cells were Ki67 positive in both cell transplanted groups (Fig. 4), which showed low cell division.
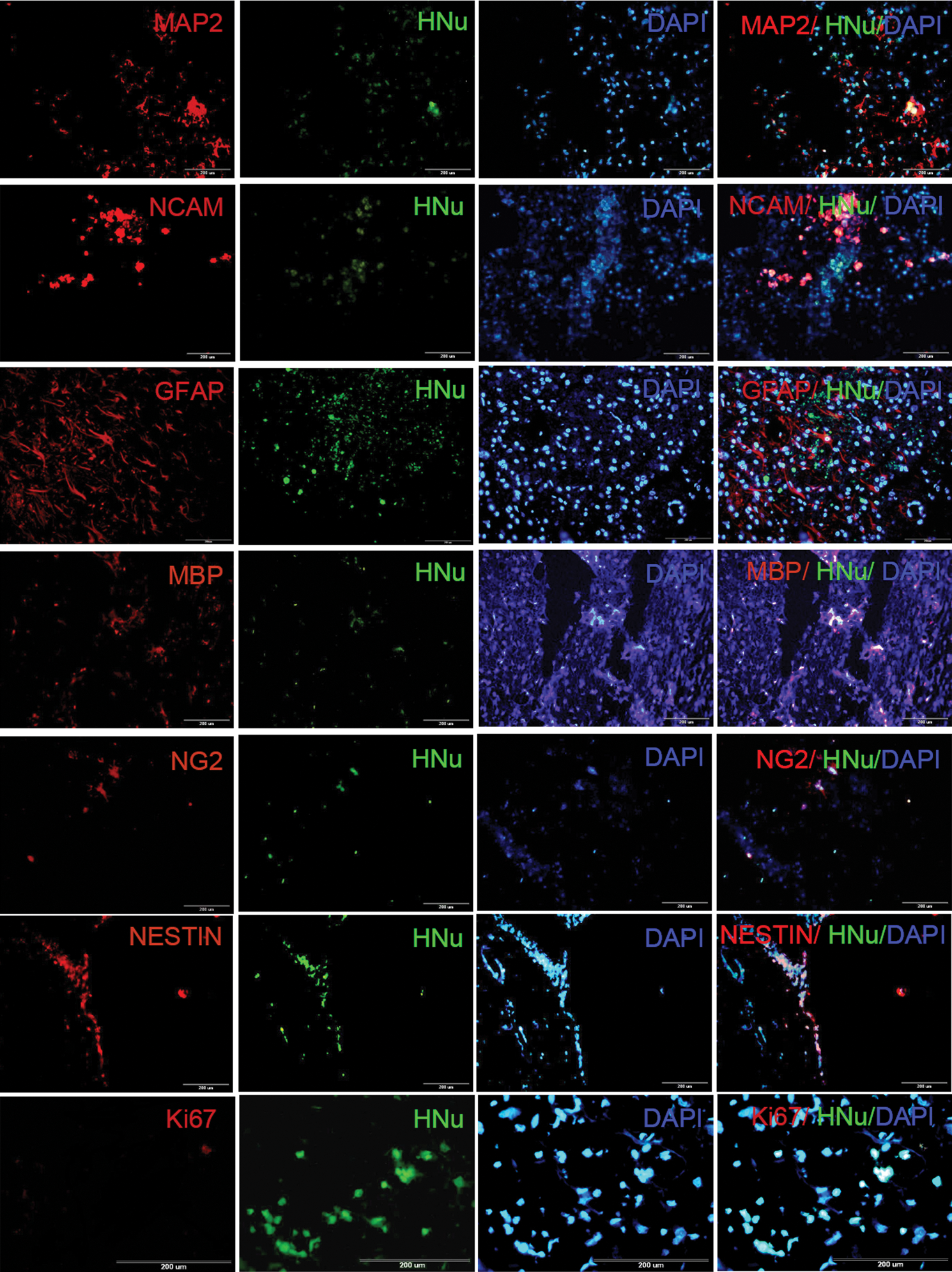
The fate of transplanted iSHED in the spinal cord. Immunohistofluorescence staining was performed for iSHED and SHED (Supplementary Fig. 1), 5 weeks after transplantation. Double immunohistostaining was performed using HNu, a transplanted human cell marker; and NESTIN, a progenitor marker; or MAP2 and NCAM, 2 mature neuron markers; or GFAP, an astrocyte marker; or MBP and NG2, 2 mature oligodendrocyte markers; or Ki67, a mitotic marker. MBP, myelin basic protein; MAP2, microtubule associated protein-2; NCAM, neural cell adhesion molecule. Color images available online at
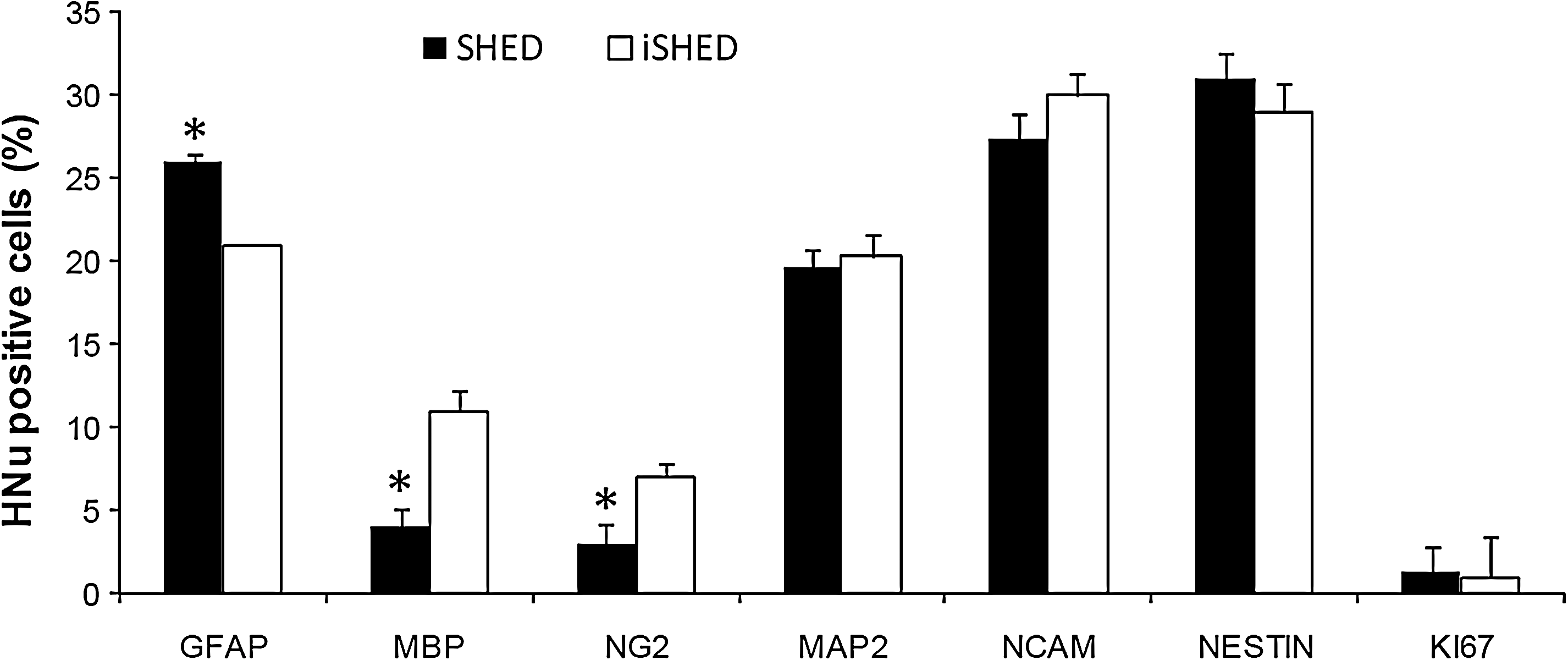
The quantification of immunohistofluorescence stainings to evaluate cell fate acquisition in iSHED group after transplantation in spinal cord. Histological analysis of cells 5 weeks posttransplantation showed that the double staining for HNu/NESTIN and neural markers (HNu/NCAM and HNu/MAP) showed similar frequency in both groups. However, the iSHED group revealed more oligodendrocyte markers, MBP and NG2 expression, and less astrocyte marker, GFAP, in comparison with the SHED group. Therefore, it may be that iSHED cells committed to an oligodendrocyte lineage in vivo. *P<0.05, by independent t-test.
Discussion
Here we have demonstrated the neuroplasticity of SHED monolayer on PLO/Fib-coated dishes in neurobasal medium supplemented with B27, ITS, and bFGF for 7 days in the absence of serum. Previously, we showed that these iSHED cells have potential to differentiate into neuronal-like cells with elongated and elaborated axon or dendrite-like structures expressing PSA-NCAM, NeuN, Tau, or TH, and glial cells with expression of GFAP after treatment with SHH/FGF8 for further 5 days [14,23]. In our study, the SHED cells expressed similar percentage of NESTIN before and after induction; however, the analysis of flow cytometry histograms revealed a shift in its expression, which suggested a prominent intensity for that as indicated before [14] in comparison with SHED cells. The high expression of NESTIN in SHED cells before induction was in agreement with previous studies [24] and maybe related to the neural crest cell origin of the dental pulp [19].
Additionally, we have investigated the potential of SHED and iSHED in recovery of locomotor function of rats with contusive SCI. Our data revealed that SHED and iSHED transplantation 7 days after contusive injury at the tenth thoracic vertebral (T10) level resulted in significant locomotor functional recovery after SCI compared with the control and vehicle groups. These improvements can be attributed to either their production of diffusible trophic factors or the differentiation potential to neural cells (glials and neurons). Previous studies have revealed that bFGF-induced or bFGF-enhanced proliferation of neuronal precursors has a regulatory role on self-renewal [14,25,26]. Jiang et al. have shown that multipotent adult progenitor cells (MAPC) treated with bFGF induced to neural cells [27]. In this regard, we have recently observed the in vitro differentiation potential of SHED cells [14].
Therefore, in the current study, we evaluated the therapeutic effects of transplanted neural iSHED on the functional recovery of a rat SCI contusion model. Interestingly, we found that transplantation of iSHED improved significant locomotor functional recovery after SCI. This improvement was higher in the iSHED group compared with the SHED group. It was also demonstrated that transplantation of neurally induced bone marrow–derived MSCs resulted in higher locomotor recovery in rat model of SCI than MSC transplantation [28,29].
Histological analysis of cells 5 weeks posttransplantation revealed that HNu-positive cells expressed neural markers (NCAM and MAP2) at the same frequency in both groups. However, the iSHED group exhibited more MBP and NG2 expression—oligodendrocyte markers—and less GFAP—astrocyte marker—in vivo after cell transplantation in comparison with the SHED group. This result maybe related to higher potential of iSHED cells for neural differentiation [14]. Additionally, a previous report has shown that bFGF promotes migration and proliferation of cells during the early stages of oligodendrocyte lineage development [30]. Therefore, it may induce iSHED to commit to an oligodendrocyte lineage following transplantation.
A literature study has suggested that this functional improvement may arise from trophic and neurotrophic secretion and modification of inflammation by grafted cells [5,31], which enhanced plasticity and neuroprotection. Our qRT-PCR analysis showed that iSHED expressed more neurotrophins (BDNF and GDNF) and especially, NTF3, which is important in promotion of axonal bridging beyond cellular grafts in the injured spinal cord [32]. Interestingly, grafting dental pulp tissue into a hemisected spinal cord increased the number of surviving motoneurons [33]. It was demonstrated that this event maybe related to functional bioactivity of the dental pulp–derived neurotrophic factors in vivo by rescuing motoneurons [33]. Therefore, the difference between SHED and iSHED in this study, at 5 weeks, may also be due to higher secretion of these products by the latter group, since neural induction had no negative effect on survival of iSHED cells in the host body. Similar to previous studies for SHED cells [5,31], we also showed survival and integration of iSHED into an injured spinal cord. Additionally, no evidence of tumor formation was observed in the transplantation groups 5 weeks postcell transplantation. However, to confirm exclusion of teratoma and tumor formation, it needs long-term evaluation. Moreover, only <3% of the transplanted cells were Ki67-positive cells in both groups, which indicated low cell division. The Ki67, which is a nucleus marker of proliferating cells and is expressed in all cell cycle phases except for G0- and Ki67-positive cells [34], was found randomly dispersed across the graft area without evidence of clustering in specific sites.
It was demonstrated that human dental tissue–derived stem cells have better neural property than bone marrow–derived MSCs [35]. Obtaining stem cells from SHED is easy and convenient, with little or no trauma; thus, they can be considered as a perfect opportunity for stem cell transplantation, which could be used along with other human dental tissue–derived stem cell populations via cell banking [36,37] to treat future injuries or ailments of wisdom teeth and permanent teeth. Additionally, SHED cells have immunomodulatory properties such as bone marrow–derived MSCs [38] and have very similar proteomic profiles to that of MSC-like cells derived from other tissues [39]. Further, SHED cells provide a viable source for dental pulp tissue engineering [40]. Therefore, it is better to obtain stem cells when a child is young and healthy and the cells have well-proliferative potential [37]. The dental stem cells have capability to provide enough cells for potential clinical applications [41,42].
In conclusion, the results from this study provide evidence that compared with vehicle, SHED and particularly iSHED transplantation has displayed higher functional improvement in rats with contused spinal cords. It may be that iSHED, at least partially, has more potential for differentiation into oligodendrocytes and/or neurotrophic secretion in the lesioned site of the spinal cord. The next question would be whether the transplanted cells integrate into the spinal tissue and replace damaged cells. This study suggests that transplantation of iSHED cells may be a suitable candidate for the treatment of SCI as well as other neuronal degenerative diseases.
Footnotes
Acknowledgment
This study was funded by a grant provided from Royan Institute and Iranian Council for Stem Cell Technology.
Author Disclosure Statement
None of the authors have any conflicts of interest to disclose and all authors support submission to this journal.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.