
Abstract
The ability to stop producing or replacing cells at the appropriate time is essential, as uncontrolled growth can lead to loss of function and even cancer. Tightly regulated mechanisms coordinate the growth of stem cell progeny with the patterning needs of the host organism. Despite the importance of proper termination during regeneration, cell turnover, and embryonic development, very little is known about how tissues determine when patterning is complete during these processes. Using planarian flatworms, we show that the planar cell polarity (PCP) pathway is required to stop the growth of neural tissue. Although traditionally studied as regulators of tissue polarity, we found that loss of the PCP genes Vangl2, DAAM1, and ROCK by RNA interference (individually or together) resulted in supernumerary eyes and excess optical neurons in intact planarians, while regenerating planarians had continued hyperplasia throughout the nervous system long after controls ceased new growth. This failure to terminate growth suggests that neural tissues use PCP as a readout of patterning, highlighting a potential role for intact PCP as a signal to stem and progenitor cells to halt neuronal growth when patterning is finished. Moreover, we found this mechanism to be conserved in vertebrates. Loss of Vangl2 during normal development, as well as during Xenopus tadpole tail regeneration, also leads to the production of excess neural tissue. This evolutionarily conserved function of PCP represents a tractable new approach for controlling the growth of nerves.
Introduction
T
We used the planarian model system, popular for regeneration studies due to a robust ability to regenerate any body part [1], to begin elucidating the sensor that terminates growth. An accessible population of adult stem cells [2] makes planaria an excellent in vivo system for studying the mechanisms by which adult stem cells direct large-scale morphologies. During tissue growth, stem and progenitor cells must be integrated with surrounding tissues, suggesting there exists a mechanism that directs stem cells to proliferate only the necessary structures—stem cells and/or new tissues must sense when that growth is complete. We decided to investigate whether the versatile planar cell polarity (PCP) pathway (Fig. 1A) might function as part of this termination sensor. Originally identified as a determinant of epithelial tissue polarity from the asymmetrical localization of pathway proteins within single cells [3], PCP also regulates cell size, proliferation, and migration [4 –6], as well as neuronal morphogenesis and the migration of both neurons and neural progenitor cells during development [7 –10]. In planarian flatworms, the conserved PCP pathway is known to regulate epithelial polarity as in other organisms [11]. We hypothesized that PCP, which coordinates individual cell behaviors into tissue-wide organization, could be involved in neuronal growth regulation signals on an organism-wide scale.
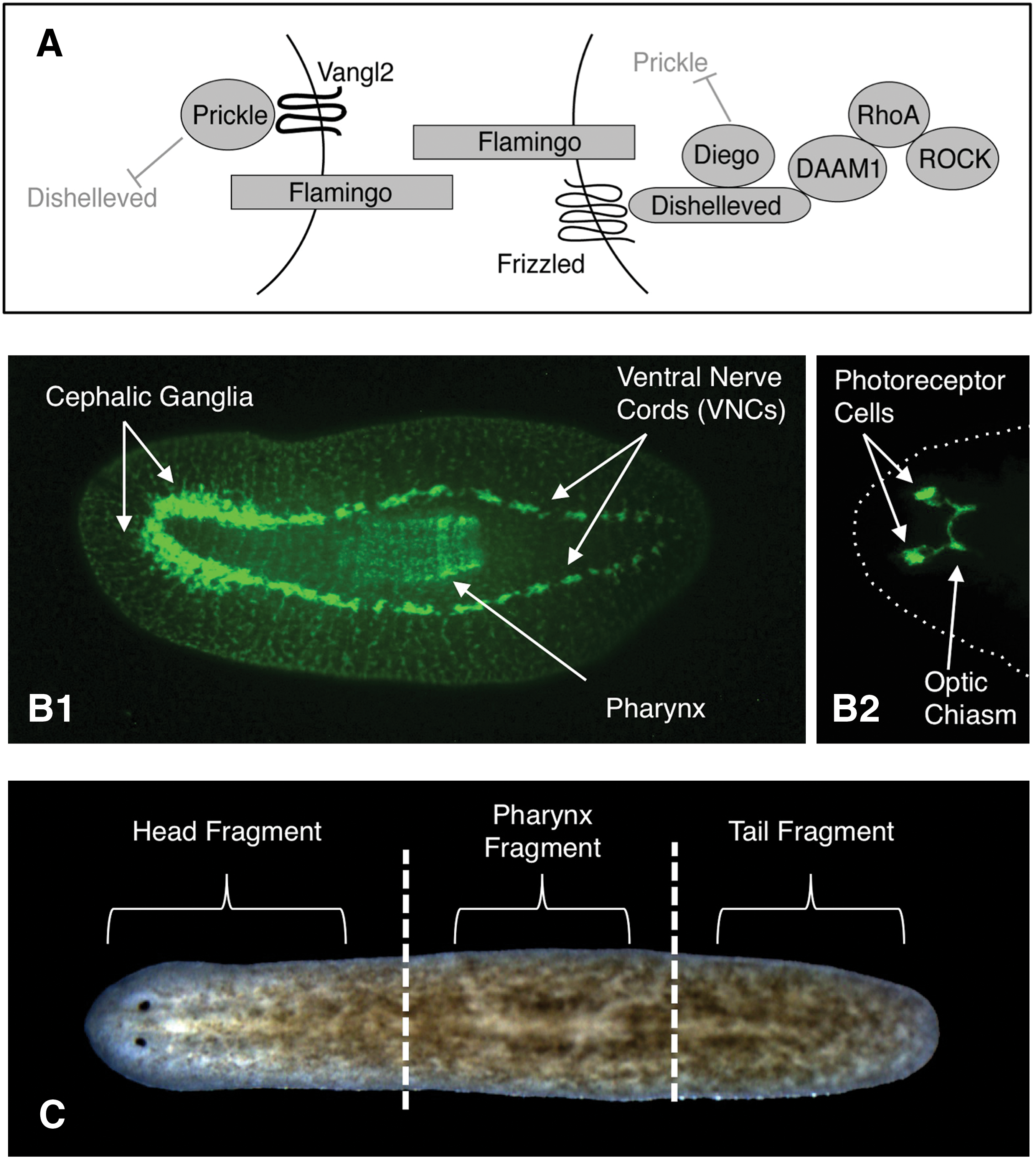
Our investigations looked specifically at growth termination of the nervous system, uncovering a role for PCP in terminating the growth of neural tissues during planarian regeneration and homeostatic cell turnover. The central nervous system (CNS) of freshwater planaria consists of a bi-lobed cephalic ganglia (brain) and 2 ventral nerve cords (VNCs) that run the length of the animal (Fig. 1B1), an optic chiasm connected to photoreceptor cells (Fig. 1B2), as well as an intricate network of commissural and sensory neurons throughout the periphery [12,13]. In addition, planaria possess most of the same neurotransmitters found in vertebrates [14 –16], making them an excellent model system for understanding neural patterning [17]. Our data show that loss of PCP through gene silencing in planarians resulted in continued neural hyperplasia for at least 6 weeks after the normal 2-week regeneration period, as well as causing the formation of excess neurons in intact planarians during normal cell turnover. Further, PCP inhibition in Xenopus tadpoles revealed that PCP-mediated regulation of neural growth during regeneration is conserved in vertebrates, while demonstrating that PCP also limits neuronal growth during embryogenesis. Together, our results suggest that the mechanism of PCP signaling acts to restrict neural growth during regeneration, homeostasis, and development.
Methods and Materials
Cloning
Homologs to Vang-like 2 (Vangl2/Van Gogh/Strabismus), Dishevelled-associated activator of morphogenesis 1 (DAAM1), and Rho-associated kinase (ROCK) were used to identify the Schmidtea mediterranea counterparts in the Genome Database [18], and the planarian genes were generated by common protocols. For details, see Supplementary Materials and Methods (Supplementary Data available online at
Colony care and amputations
The CIW4 clonal line of asexual S. mediterranea was used and maintained as previously described [19]. Worms 4–7 mm long were starved for ≥1 week prior to use. Details on care and amputations are provided in Supplementary Materials and Methods.
In situ hybridization
Worms for whole-mount in situ hybridization were formalin fixed, and in situs were performed as described by Pearson et al. [20]. Use of this protocol may explain the differences between our Smed-vang-1 staining and that previously reported [11]—which used a different protocol and did not observe intestinal upregulation (see Supplementary Fig. S3 for probe controls). Probes: Piwi-1 [21]; NB.21.11.e [22]; sFRP-1 and WntP-2/Wnt11-5, kind gifts of P. Reddien [23]; and Cintillo [24].
Immunofluorescence
Worms were fixed in Carnoy's fixative and antibody stained by common protocols. For details, see Supplementary Materials and Methods. Primary antibodies: anti-phosphorylated histone H3, 1:250; anti-arrestin, 1:10,000 (kind gift from K. Watanabe); and anti-synapsin, 1:50 (Developmental Studies Hybridoma Bank). Secondary antibodies: goat anti-mouse Alexa488, 1:400 (Sigma); or for H3P, a horseradish peroxidase (HRP)-conjugated anti-Rabbit with TSA-Alexa568 anti-HRP (Molecular Probes). For excess photoreceptor counts, discrete areas of ectopic anti-arrestin staining were scored (eg, see yellow arrows in arrestin staining images).
RNA interference
For RNA interference (RNAi) experiments, dsRNA was generated and injected as described by Oviedo et al. [25]. For details, see Supplementary Materials and Methods. Combination PCP-Cocktail-RNAi was made by injecting with equal parts of DAAM1, ROCK, and Vang-1 RNAi. Control injections were with either PC2-RNAi or VENUS-GFP RNAi.
Image collection and statistics
Images were collected using either a Nikon SMZ1500 microscope with a Retiga 2000R camera (Q-Imaging) and Q-Capture imaging software, or an Olympus BX61 compound microscope with a Hamamatsu Orca AG CCD camera and IP Labs or MetaMorph imaging software. Adobe Photoshop was used to orient and scale images and improve contrast. Data were neither added nor subtracted; original images available upon request. Microsoft Excel was used to calculate Student's t-test (2-tailed distribution, 2 independent samples, unequal variance).
Xenopus experiments
Xenopus laevis larvae were cultured via approved protocols (Institutional Animal Care and Use Committee, #M2008-08). Four-cell-stage embryos were injected into 2 of 4 cells using 1 mM Vangl2 morpholino (Vangl2-MO; Gene Tools). The Vangl2-MO [26,27] is tagged with flourescein, and its presence in stage-40 tail tissues was identified by fluorescence. For tail regeneration, tails with good flourescein expression at stages 40–41 were amputated at the midpoint between the anus and the tip. Tadpoles were cultured in 0.1 × Marc's modified Ringers at 22°C for 10 days. Xenopus embryos were fixed overnight in 100 mM MOPS at pH 7.4, 2 mM EGTA, 1 mM MgSO4, 3.7% v/v formaldehyde [28] and processed for immunohistochemistry using standard protocols. Antiacetylated tubulin (Sigma) at 1:1,000 and goat anti-mouse Alexa555 (Molecular Probes) at 1:500 were used to visualize the nervous system.
Results and Discussion
Cloning planarian PCP genes
Searching the S. mediterranea genome [18] by sequence homology, we identified and cloned full-length planarian homologs for Van Gogh 2 (Vangl2, recently named Smed-vang-1 in 11), Rho-associated kinase (Smed-ROCK, Supplementary Fig. S1), and Dishevelled-associated activator of morphogenesis 1 (Smed-DAAM1, Supplementary Fig. S2). When analyzed by in situ hybridization in untreated, intact planaria, these genes were expressed throughout the worm, with some upregulation in the gastrovascular tract and anterior neural tissues (Fig. 2 and Supplementary Fig. S3). Analyses during regeneration showed that all 3 genes were upregulated in both the anterior and posterior blastemas. In particular, both Vang-1 and DAAM1 appeared to be slightly upregulated in the brain primordia at day 3 of regeneration (Fig. 2, red arrows). This upregulation was gone by 7 days, although all 3 genes continued to be expressed in both blastemas throughout regeneration (Supplementary Fig. S3).
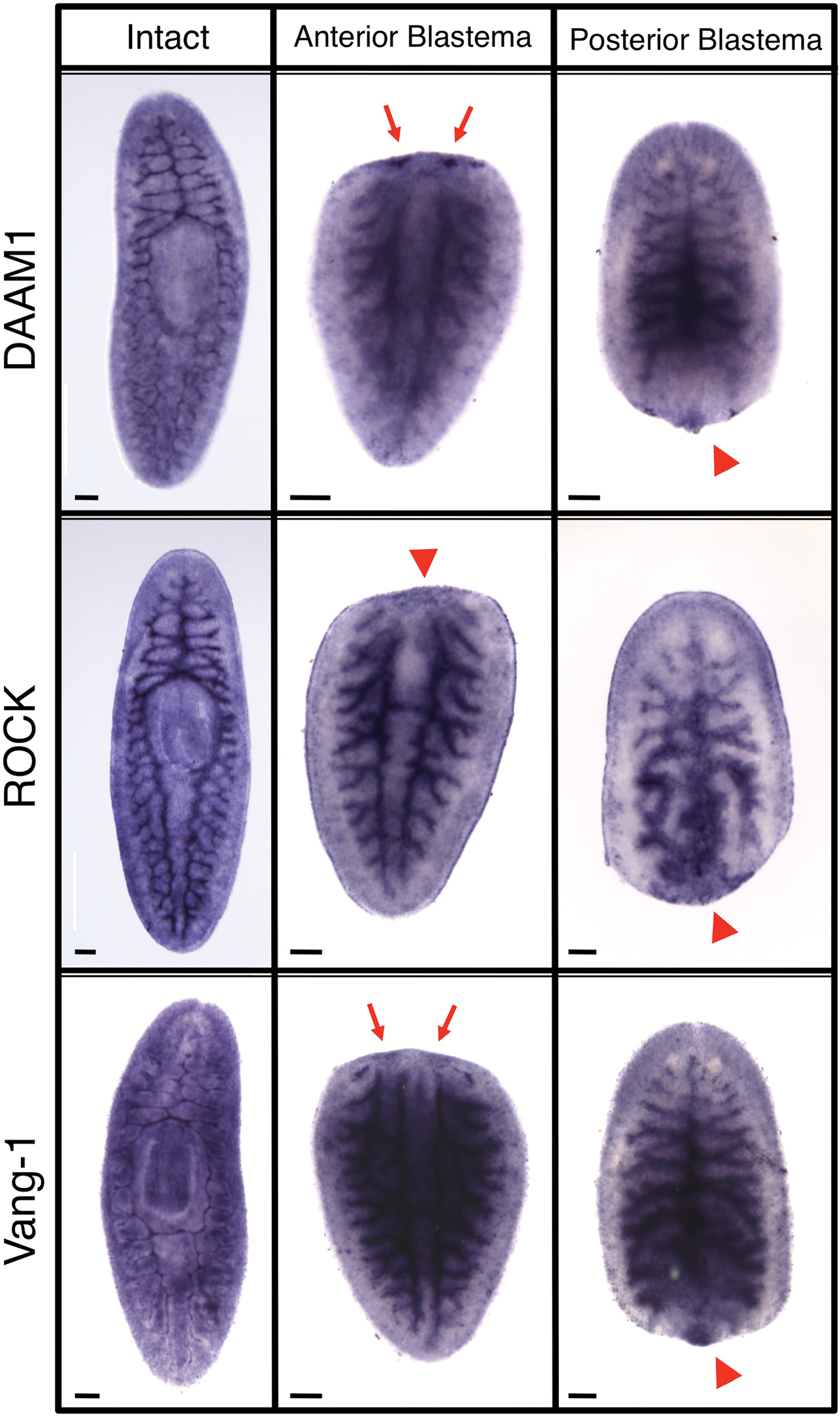
Planarian planar cell polarity (PCP) genes. In situ hybridization patterns in intact (whole) and regenerating (at day 3 post-amputation) bisected planaria. Vangl2 (Smed-vang-1), Rho-associated kinase (Smed-ROCK), and Dishevelled-associated activator of morphogenesis 1 (Smed-DAAM1). Arrowheads indicate expression in the blastema. Arrows indicate expression in brain primordia. Anterior is up. Scale bars: 100 μm. Color images available online at
Loss of PCP results in excess visual neuron growth during planarian regeneration
Since new tissue growth is perhaps most easily assessed as part of blastema formation, we began by investigating the role of PCP during regeneration. We used RNAi to silence Vang-1, DAAM1, and ROCK in regenerating head, pharynx, and tail fragments (Fig. 1C), and uncovered a role for PCP specifically in restricting the growth of the regenerating nervous system. As regeneration in planarians is typically scored at 2 weeks post-amputation, we assessed animals at 8 weeks post-amputation to identify possible effects from continued regenerative growth. The first phenotype observed was an increase in eye structures following PCP inhibition. During regeneration, eye structures first become visible ∼5–7 days following decapitation. Although it usually occurs normally, eye regeneration in planaria is somewhat labile; even during normal asexual fissioning, a 1% occurrence of supernumerary eye regeneration has been reported [29]. When untreated controls are experimentally amputated, the appearance of excess eye structures (such as pigment cells) during regeneration can be as high as 6.7% (n=30).
We found that regenerates injected with RNAi targeting individual PCP pathway members possessed excess eyes and supernumerary eye structures (such as pigment cells) in an average of 31.1% of animals (Figs. 3A2 and 4), versus only 7.2% when control RNAi was injected (Fig. 3A1). Specific counts of pigment cells in Smed-vang-1(RNAi)-injected animals at 8 weeks of regeneration revealed an average of 1.16 extra pigment cells per animal (red arrows in Fig. 3A), as compared with only 0.1 extra pigment cells in controls (n>18, P<0.001). Inhibition of single PCP genes by RNAi also resulted in several eye patterning defects, including abnormal eye alignment and asymmetric eye size and orientation (Supplementary Fig. S4), consistent with PCP's known role in patterning. To maximize the effects of PCP inhibition, RNAi targeting Vang-1, DAAM1, and ROCK was combined (PCP Cocktail RNAi), which resulted in up to a 2-fold increase in phenotypic effects (Fig. 3B). In particular, this combination increased visual cell regeneration, leading to a “cauliflower” phenotype where numerous excess pigment cells were overlaid (Fig. 3A5 and Supplementary Fig. S4). This demonstrates that the PCP pathway regulates eye regeneration, suggesting a role for PCP in stopping the formation of visual neurons as well as in neural patterning.
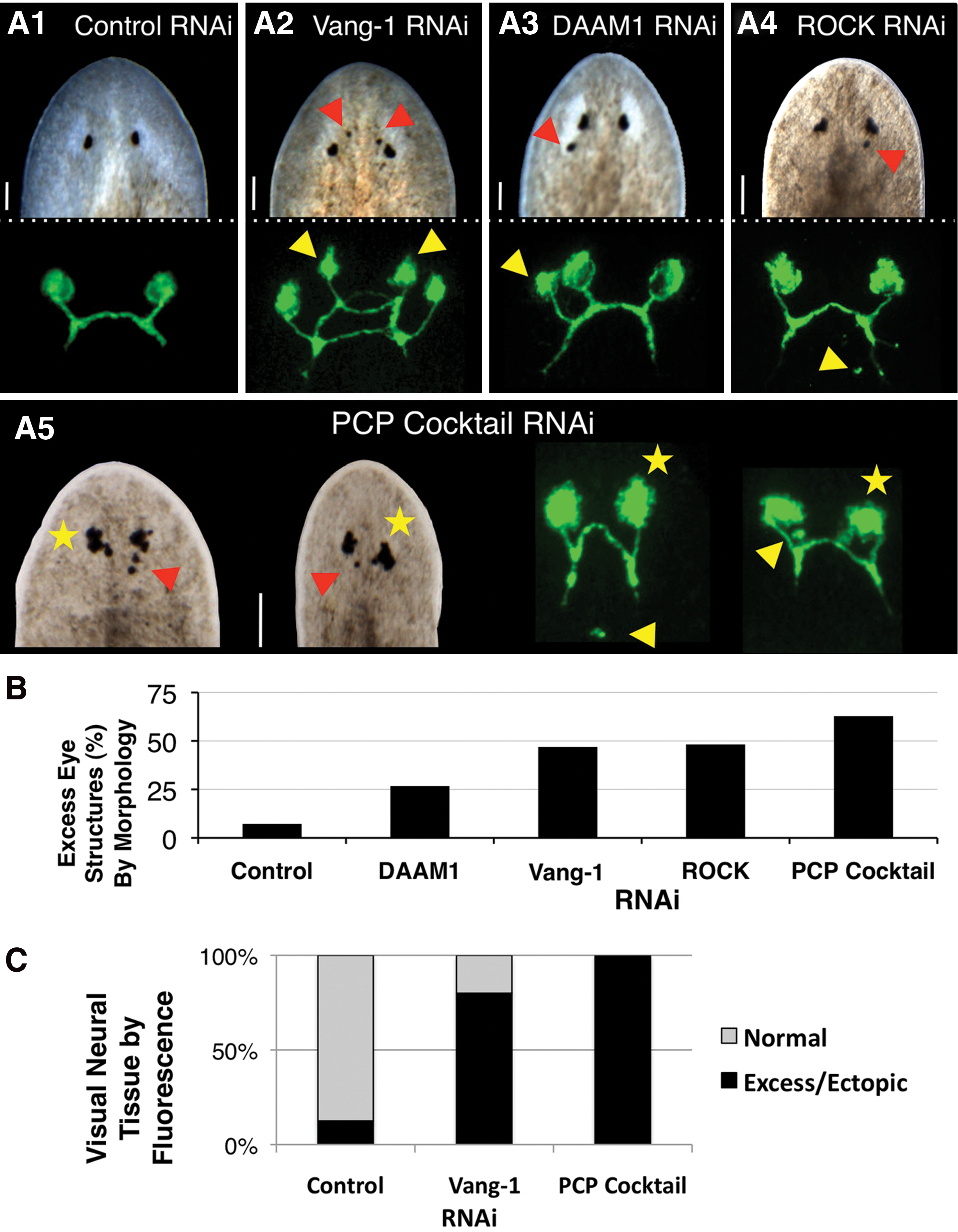
PCP regulates eye innervation and visual neuron formation during regeneration.
To corroborate our morphological observations, we examined the placement of optic nerves using a visual-neuron-specific marker that recognizes planarian photoreceptor cells (Fig. 3A, C). By 8 weeks of regeneration (6 weeks after regeneration is typically considered complete) we observed both extra photoreceptors and optic neuronal projections following RNAi injection for each of the 3 genes in regenerating head and tail fragments. Counts of Smed-vang-1 (RNAi)-injected animals revealed an average of 1.7 discrete areas of excess photoreceptor staining per animal (yellow arrows in Fig. 3A) when Vang-1 was inhibited, as compared with 0.3 in controls (n≥20, P<0.001). The increase in visual neurons detected by immunofluorescence when single PCP genes were inhibited was much more penetrant than the corresponding increase in eye structures detected by morphology alone (81% on average, vs. 31.1%).
Strikingly, injection of the combination PCP Cocktail RNAi, while producing similar phenotypes to individual RNAi injections, increased the incidence of excess optic neural tissue to 100% of tested regenerates (Fig. 3C). Additionally, ectopic photoreceptors (normally anterior to the optic chiasm) were seen atypically in more posterior tissues (yellow arrows in Fig. 3A5), although none was ever observed outside of the tissues anterior to the pharynx. Analyses of anterior and posterior molecular markers confirmed that PCP inhibition had no effect on the anterior-posterior polarity of the fragments (Supplementary Fig. S5). The occurrence of excess photoreceptors following inhibition of PCP genes (both individually and together) is consistent with excess optic neural growth following RNAi to another PCP pathway member, RhoA, in a different planarian species [30]. Our data reveal that multiple visual cell types (both pigment cells and photoreceptors) were affected by PCP inhibition. This implicates PCP signaling in regulating the formation of new neurons in planarians, rather than merely augmenting axon regeneration. Together, these data suggest that PCP regulates optic neuronal replacement during regeneration, controlling both the extent and patterning of visual innervation.
PCP is required to terminate visual neuron growth during planarian homeostasis
Morphostasis of adult bodies requires continual replacement of aging tissues. Because neural growth occurs not just during regeneration, but also during homeostatic cell renewal, we next looked at eye morphology in intact worms (not undergoing regeneration) following PCP inhibition. Although excess eye structures never occurred in control worms (n=9), supernumerary eye structures began to appear 2–4 weeks after injection of Smed-vang-1(RNAi) (Supplementary Fig. S6A). At 8 weeks post-injection, Smed-vang-1(RNAi) in intact worms resulted in 29.2% of animals with supernumerary eye structures (n=26). The delayed timing for the onset of excess neural tissue formation in intact PCP-inhibited worms is consistent with a role for PCP during cell turnover in the planarian nervous system, although it does not rule out cell differentiation effects. These data suggest that the PCP pathway is required to inhibit neural growth during homeostasis, as well as during regeneration.
Loss of PCP leads to neural hyperplasia throughout the planarian nervous system
To determine whether PCP's role in neuron renewal was limited to visual nerves, we examined the entire planarian nervous system following PCP silencing. We continued our analyses using the combination PCP Cocktail RNAi, since the phenotypic penetrance of RNAi to individual genes was suboptimal. We found that PCP Cocktail RNAi in regenerating pharynx fragments resulted in increased nerve induction and disrupted neural patterning throughout the animal when visualized by a pan-neural maker (Fig. 4). To identify regeneration-specific phenotypes (by reducing the effects from PCP regulation of neural homeostasis), we injected RNAi following amputation and scored at 2 weeks postamputation when neural regeneration is first considered complete. Analyses of the commissural neurons that connect the brain and 2 VNCs revealed that following injection of PCP Cocktail RNAi there was a 51.9% increase in the number of transverse commissures, as well as a 32.1% increase in the number of associated lateral processes when compared with controls (Fig. 4A, B). These data demonstrate that the presence of excess neuronal growth following PCP inhibition is not limited to the visual neurons but extends throughout the planarian nervous system. We also observed additional neuronal phenotypes in PCP Cocktail RNAi–injected worms. In the peripheral nervous system, the dorsal submuscular nerve plexus was mispatterned, with dense and irregularly spaced neurons consistent with increased nerve regeneration (Fig. 4C). The CNS exhibited patterning defects as well, with excess and/or ectopic neurons extending from the cephalic ganglia (Fig. 4D) (without an apparent increase in the overall size of the brain region) and along the VNCs (Fig. 4D, E). Together, excess and/or ectopic CNS and peripheral neural tissue occurred in 81.5% of PCP Cocktail RNAi–injected regenerates (vs. 8.3% with control RNAi). Together, these data suggest that PCP regulates replacement of the entire nervous system during regeneration.
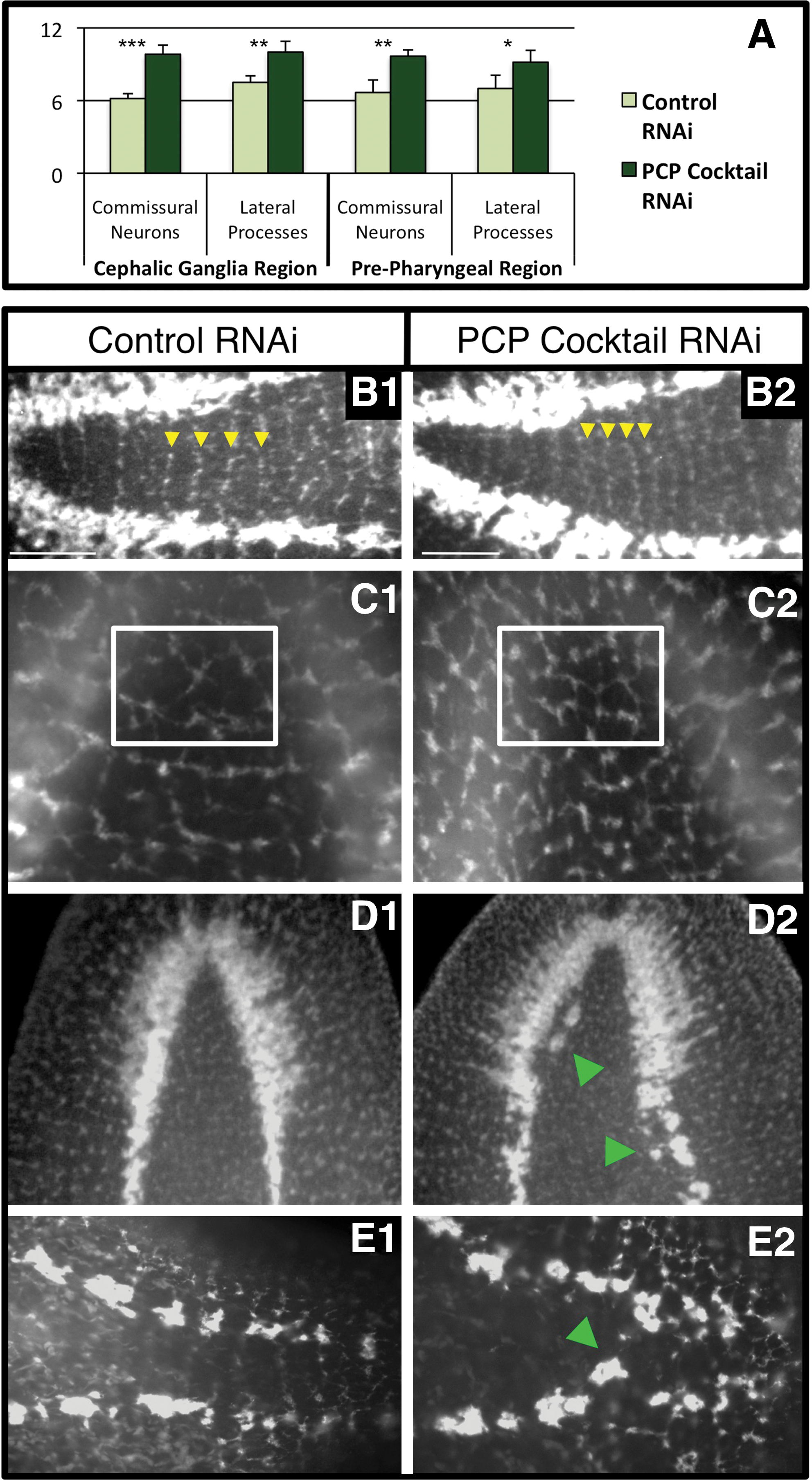
Loss of PCP results in neural hyperplasia. PCP regulates regeneration of both the central and peripheral nervous systems.
Loss of PCP results in increased proliferation and stem cell progeny
Our data demonstrate that PCP regulates neuronal growth, but this could be the result of disrupted neuronal patterning rather than a failure to terminate growth. If increased nerve formation was due to initial mispatterning, then we would expect the number of ectopic nerves to stay the same after regeneration was completed. However, if excess neural tissue was the result of a failure to halt regenerative growth, we would expect to see continued nerve formation over time. This last hypothesis was consistent with our observations of eye regrowth in PCP Cocktail RNAi–injected worms, where excess eye structures appeared to gradually accumulate rather than arise immediately (Fig. 5A). To distinguish between the 2 hypotheses, regenerating Vang-1-inhibited worms were followed over the course of 8 weeks. RNAi to Smed-vang-1 alone was used because its incomplete penetrance would highlight any increases in eye growth. When the same worms were examined visually every 2 weeks, we found that the incidence of excess eye structures doubled within 2 months, with the largest increase between 6 and 8 weeks (Fig. 5B1). The initial incidence of excess eye structures at 2 weeks of regeneration was comparable to our previously observed incidence at 8 weeks as a result of homeostasis (29% for each). However, the much higher incidence of excess eye structures by 8 weeks of regeneration (57%), along with the continued formation of new eye tissue over the course of the trial, indicates that these results are regeneration specific. The data suggest that loss of PCP results in more than disrupted neuronal patterning, but that PCP genes play a role in halting neuronal regeneration.

PCP inhibits stem-cell-mediated growth during regeneration.
This continual maintenance of regenerative growth implies that there was similarly extended mitotic activity in PCP-inhibited worms. Planarian regeneration is accompanied by a marked mitotic increase [31], leading to new tissue formation and patterning. When we examined mitotic activity by phosphorylated histone-3 labeling, we observed that increased mitosis was still present at 8 weeks of regeneration in Vang-1-inhibited regenerates (Fig. 5B3), which was not seen in control regenerates (Fig. 5B2). This is consistent with our observations that neural growth continues over time when PCP is inhibited, and suggests that PCP is required for regeneration-mediated proliferation to cease.
In planarians, the only mitotically active cells are a large population of pluripotent adult stem cells (neoblasts) responsible for replacing all lost or damaged cells [32,33]. Increased mitotic activity following PCP inhibition could be the result of increased numbers of neoblasts. However, when visualized by Smed-piwi-1 staining, we did not observe an increase in the neoblast population in PCP Cocktail RNAi–injected regenerates (Fig. 5C). Alternatively, sustained mitosis could be the result of an increase in stem cell progeny produced from the normal neoblast population. Using a marker of early neoblast descendents, Smed-NB.21.11e [22], we observed a modest increase at 8 weeks post-amputation in neoblast progeny after PCP inhibition (Fig. 5D), consistent with increased neural growth throughout the entire planarian nervous system. While our data are indicative of increased neuron numbers due to more proliferative cycles (resulting in additional stem cell progeny), it does not rule out the possibility that PCP inhibition also influences stem cell fate changes. If such fate changes do exist, they likely involve the new stem cell progeny, rather than altering the differentiation of existing stem cells, as our analysis revealed no evidence of defects that would result from diverting neoblasts from making other necessary tissues. However, detailed investigation of other organ systems in PCP-inhibited regenerates will be required to determine whether or not stem cell fates are being switched to neural at the expense of another cell type. It is noteworthy that at 8 weeks of regeneration overall body morphology in PCP-inhibited worms was indistinguishable from controls—PCP inhibition did not result in either larger/overgrown or smaller/undersized worms (Supplementary Fig. S6B). This suggests at a minimum that the master regulator that maintains planarian body shape over time (allometric scaling) is still functional even when PCP is disrupted.
PCP restricts neural growth during vertebrate regeneration and development
The data suggest that PCP is required to limit neural tissue growth during both homeostasis and regeneration in planarians. To test whether the PCP pathway also regulates neural replacement in vertebrates, we examined the effects of a Vangl2 morpholino (XVangl2-MO) [26,27] during Xenopus tadpole tail regeneration (Fig. 6), a well-established regenerative model we have previously used to identify novel regulators of regeneration [34,35]. Xenopus Vangl2 is upregulated in neural tissues during development and is involved in both gastrulation and neural tube closure [36,37]. Following tail amputation, XVangl2-MO-injected tadpoles regenerated with excess neural tissue that extended ectopically toward the tip of the new tail, corresponding to a 32.8% increase in neuronal length (Fig. 6A). This suggests that PCP-mediated termination of neural growth is conserved in Xenopus regeneration. Further, we found that Vangl2 inhibition during normal tadpole development (without amputation) revealed excess neuronal growth along the flank (Fig. 6B), as well as a 20.9% increase in neuronal length toward the tip of the tail (Fig. 6C). This suggests that Vangl2 also acts to restrict neural growth in Xenopus during embryonic development. The data implicate a conserved role for PCP in regulating neural growth during regeneration in both vertebrates and invertebrates.

PCP regulation of neural growth is conserved in vertebrates. Immunofluorescent anti-acetylated-tubulin labeling of Xenopus tadpole neurons.
Following inhibition of PCP genes, our data demonstrate that excess neural tissue is produced and excess proliferation occurs in planarian regenerates, continuing to arise long after controls have ceased regenerative growth. While the data are consistent with loss of PCP resulting in increased neurons due to increased stem cell progeny, subsequent investigations will specifically delineate the individual contributions of proliferation versus axonal patterning in PCP regulation of neural growth. Similarly, our data show that PCP inhibition leads to ectopic neural differentiation, as illustrated by the appearance of excess eye structures when PCP is lost (Fig. 3). This could reflect a failure to spatially restrict neural differentiation (thus causing photoreceptor cells to occur within the endogenous field as well as ectopically). While our data cannot exclude this hypothesis, any contribution to spatial restriction is probably secondary, since ectopic eye structures were never seen outside of the normal cephalic/head region. Instead, our findings suggest that the major role of PCP during neural regeneration is to temporally restrict neural differentiation (thus when lost causing neural growth to continue long after neural regeneration should be complete), as illustrated by the gradual appearance of excess pigment cells after regeneration of the primary eyes in PCP-inhibited worms (Fig. 5A).
The data suggest that PCP genes are required to properly terminate neural growth during regeneration, homeostasis, and development—3 instances in which large-scale pattern depends on growth regulation of individual cells. PCP regulation of neural growth during development suggests that planar polarity may play a role in neural plasticity by inhibiting the number of neurons formed during neurogenesis; this model will also be investigated in subsequent studies. In addition to highlighting a mechanism by which (neural) tissue growth is normally stopped, our results indicate that a conserved mechanism stops growth in diverse biological processes. Our data also provide evidence that limiting the extent of neural tissue is in fact an ancestral role of PCP, as Vangl2 inhibition during Xenopus development and tail regeneration also resulted in increased neural growth. Although invertebrates, planarians' similarity to the common bilaterian ancestor suggests that PCP's role in restricting neural growth is conserved throughout the Bilateria. This raises the interesting speculation that this ancestral role for PCP may have been co-opted in non-regenerative organisms during tumorigenesis. This is supported by growing evidence for the involvement of PCP during cancer, from findings such as the tumor suppressor role of Vangl2 [38,39]. Our data show that disrupted PCP signals the formation of new neural tissue and suggests that reestablishing intact PCP halts further neural replacement. In this way, we propose that PCP acts as a sensor by which neural tissue can determine the status of its fine-level organization, signaling “all is complete” when replacement is no longer needed. Finally, our data show that PCP inhibition promotes regeneration, which suggests that pharmacological PCP inhibitors might be a viable approach toward translational therapies.
Footnotes
Acknowledgments
The authors thank L. Vandenberg for help with Xenopus, C. Stevenson for molecular assistance, and K. Watanabe and P. Reddien for reagents. W.S.B. also thanks J.L.B. for continued support. W.S.B. was supported by NIH Kirschstein-NRSA grant F32 GM08354. M.L. gratefully acknowledges support by the Mathers Foundation, as well as NSF grant IBN# 0347295, NHTSA grant DTNH22-06-G-00001, and NIH grant HD055850.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.