
Abstract
Embryonic stem cells (ESCs) can differentiate to endothelial progenitor cells and vascular endothelial cells (VECs), but the mechanism is largely unknown. In this study, we synthesized 2 chiral compounds (R-ABO and S-ABO) and identified R-ABO as an effective inducer of ESC differentiation into VECs. Furthermore, we found that R-ABO induced ESC differentiation into VECs via homeobox containing 1 (Hmbox1) that acted upstream of fibroblast growth factor 2 (FGF-2). The data suggest that R-ABO is a novel tool for ESC differentiation into VECs, and Hmbox1 is a key regulator in this differentiation process. These findings provide information on a novel target and a new platform for further investigating the gene control of ESC differentiation to VECs.
Introduction
V
Embryonic stem cells (ESCs) have been proposed as a preferable source of endothelial cells due to their low immunogenic profile and high proliferative potential [3]. It has been shown that ESCs can be induced to differentiate into VECs in vitro in a leukemia inhibitory factor (LIF)-free growth medium combined with some growth factors [4 –6]. However, the differentiation rate is very low. It is crucial to develop a productive method of VECs from ESCs for therapeutic and tissue-engineering applications. Moreover, the underlying mechanism of ESC differentiation into VECs is not well understood.
In our previous studies, we synthesized a series of 2,3-dihydro-3-hydroxymethyl-1,4-benzoxazine derivatives by a tandem reduction–oxirane opening of 2-nitroaroxymethyloxiranes. Among the 7 compounds (3a–g), only 6-amino-2, 3-dihydro-3-hydroxymethyl-1, 4-benzoxazine (ABO) (3g) markedly inhibited the apoptosis of human umbilical vein endothelial cells [7]. Furthermore, we found that ABO induced angiogenesis both in vitro and in vivo [8]. Otherwise, our recent study showed that ABO could induce bone marrow stromal cell (BMSC) differentiation to VECs by elevating Hmbox1 expression [9]. ESCs are different from BMSCs in proliferation potential and pluripotency, so we are interested in whether ABO can induce ESC differentiation into VECs or not.
In this article, ESCs were treated with 2 chiral compounds (R-ABO and S-ABO). The results showed that R-ABO could effectively induce ESC differentiation into VECs. ESCs treated with R-ABO for 10 days could significantly express mature VEC markers, such as CD31, VE-cadherin, antihemophiliac factor (AHF, also known as F VIII), endothelial nitric oxide synthase (eNOS), and vascular endothelial growth factor (VEGF) receptor 2 (Flk-1). ESCs induced by R-ABO for 10 days formatted the capillary-like structure on Matrigel and facilitated acetylated low-density lipoprotein uptake. In the differentiation system of ESCs induced by R-ABO, the level of Hmbox1 was increased. Knockdown of Hmbox1 in ESCs by its short-hairpin RNA (shRNA) inhibited the differentiation. Furthermore, we found that Hmbox1 acted upstream of fibroblast growth factor 2 (FGF-2) to induce ESC differentiation to VECs. The data suggested that R-ABO induced ESC differentiation into VECs via upregulating Hmbox1 that was in the upstream of an FGF-2 classical proangiogenic pathway in ESCs.
Materials and Methods
Synthesis of R-ABO and S-ABO compounds
According to our previous article [7], enantioselective ABOs were synthesized from commercially available chiral glycidyl tosylates (R)- and (S)- as starting materials [10]. Briefly, 2,4-dinitrophenol reacted with (S)-glycidyl tosylate in the presence of NaH in DMF to afford (S)-2-((2,4-dinitrophenoxy)methyl)oxirane. Then, the tandem reduction–oxirane opening of (S)-2-((2,4-dinitrophenoxy)methyl)oxirane in the presence of iron powder and acid gave (R)-(6-amino-3,4-dihydro-2H-benzo[b][1,4]oxazin-3-yl)methanol, (R)-ABO [
Cell Culture
The mouse ESC line (Strain 129/SVEV) and primary mouse embryonic fibroblasts were purchased from Millipore. Undifferentiated ESCs were grown on a feeder layer of mouse embryonic fibroblasts. Strain 129/SVEV was maintained in a high-glucose DMEM (Gibco) supplemented with 15% fetal bovine serum (FBS) (Gibco), 2 mM
ESC differentiation
Differentiated ESCs were cultured on 0.1% gelatin-coated dishes as monolayer and maintained in a high-glucose DMEM (Gibco) supplemented with 15% FBS (Gibco), 2 mM
In vitro capillary-like tube formation assay
The formation of capillary-like structures in ESCs that were induced by 10 μM R-ABO for 10 days was assessed on Matrigel (BD Biosciences) as previously described [11]. Cells were seeded on 24-well plates coated with Matrigel at 5×104 cells per well in a basal high-glucose DMEM and incubated at 37°C for 60 min. Cells were treated with or without R-ABO for 10 days in the absence of serum and then incubated at 37°C for 48 h. Tube formation was observed under an inverted phase-contrast microscope (Nikon) at 12 and 24 h. The length of tubes was measured in random fields from each well by the National Institutes of Health Image program.
Immunofluorescence assay
Immunofluorescence assay was performed as previously described [12]. In brief, after treatment, cells were fixed with 4% paraformaldehyde for 15 min and blocked with 3% normal goat serum for 20 min at room temperature (RT). Cells were incubated with CD31, Flk1, F VIII, VE-cadherin, or eNOS primary antibody (1/100) at 4°C overnight. Then, cells were treated with a corresponding FITC-conjugated secondary antibody (1/200) at 37°C for 1 h. Cells were rinsed 3 times with 0.1 M PBS to eliminate the uncombined secondary antibody. A laser-scanning confocal microscope (LSCM; Leica) was used for fluorescence detection. Images are representatives of 3 independent experiments.
Acetylated low-density lipoprotein uptake assay
ESCs induced by 10 μM R-ABO for 10 days were seeded on 24-well culture plates. Then, cells were incubated in 10 μg/mL Alexa Fluor 488-acetylated low-density lipoprotein (Alexa 488-Ac-LDL) or 1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine perchlorate-acetylated low-density lipoprotein (Dil-Ac-LDL; Invitrogen) at 37°C for 4 to 6 h. After incubation, the cells were fixed with 4% paraformaldehyde for 15 min and were incubated with propidium iodide (PI) for 15 min at RT. The cells were observed under an LSCM (Leica).
Quantitative real-time PCR
RNA was isolated, and real-time PCR was performed as previously described [9]. The list of primers is summarized in Supplementary Table S1.
shRNA interference
shRNA against Hmbox1 was purchased from Origene. A standard protocol for transfection was used. Constructs were transfected using TurboFectin 8.0 (Origene) in a high-glucose DMEM followed by replacement of serum post-transfection. Twenty-four hours post-transfection, the medium was replaced. Pick up the green fluorescent protein (GFP)-positive clone into 96-well plate until there was no GFP-negative cell in the clone. shRNA sequences are shown in Supplementary Table S2.
Western blotting
Western blotting was performed as described previously [13]. Briefly, whole-cell lysates were applied to a 12% or 9% SDS-PAGE gel and subsequently electroblotted onto polyvinylidene difluoride membranes. After blocking the membrane at RT for 1 h, the membrane was incubated with various primary antibodies (anti-Hmbox1, anti-CD133, anti-CD31, anti-VEGF, anti-FGF-2, platelet-derived growth factor-BB (PDGF-BB), or anti-GAPDH) at 4 °C overnight. After a washing in PBST and PBS, the membrane was incubated with peroxidase-conjugated secondary antibodies for 1 h at RT, and the signals were chromogenously developed by use of 3,3′- diaminobenzidine. GAPDH was used as a loading control. The relative quantity of the protein level was analyzed by Quantity One software (Bio-Rad).
Flow cytometry
ESCs were treated with 10 μM R-ABO for 10 days. Then, the cells were harvested by trypsinization and pipetting to produce a single-cell suspension. To determine the surface CD31 antigen expression, the cells were incubated with the FITC-labeled rat anti-mouse CD31 (BD pharmingen). The cells were analyzed using an FACSCaliber flow cytometer (BD Biosciences). CD31-positive cells were determined as the proportion of the population with higher fluorescence than 95% of the isotype control.
Statistical analyses
Data are presented as means±standard error and analyzed by SPSS software (SPSS, Inc.). Mean values were derived from at least 3 independent experiments. Differences at P<0.05 were considered statistically significant.
Results
R-ABO, but not S-ABO, induced ESC differentiation into endothelial-like cells
In achiral environments, enantiomers have the same properties, but the effects are quite different in chiral environments. The reason is that enzymes and receptors can be chirally selected in biological environments. In some cases, an isomer may cause a negative impact in a racemic mixture. So, the better option is to get the more active enantiomer in a single enantiomer drug [14]. ABO used in our previous study is a mixture of the 2 chiral compounds, R-ABO and its enantiomer S-ABO. To clarify which one is more effective, we synthesized R-ABO and its enantiomer S-ABO, respectively (see Supplementary Data).
To find the most effective small molecule for inducing ESC differentiation into VECs, we treated ESCs with racemic ABO, R-ABO, and S-ABO at 1 to 20 μM for 1 day to 10 days, respectively. As shown in Fig. 1, ESCs treated with racemic ABO or R-ABO, but not S-ABO, effectively differentiated to VEC-like cells compared with the control group in morphology.
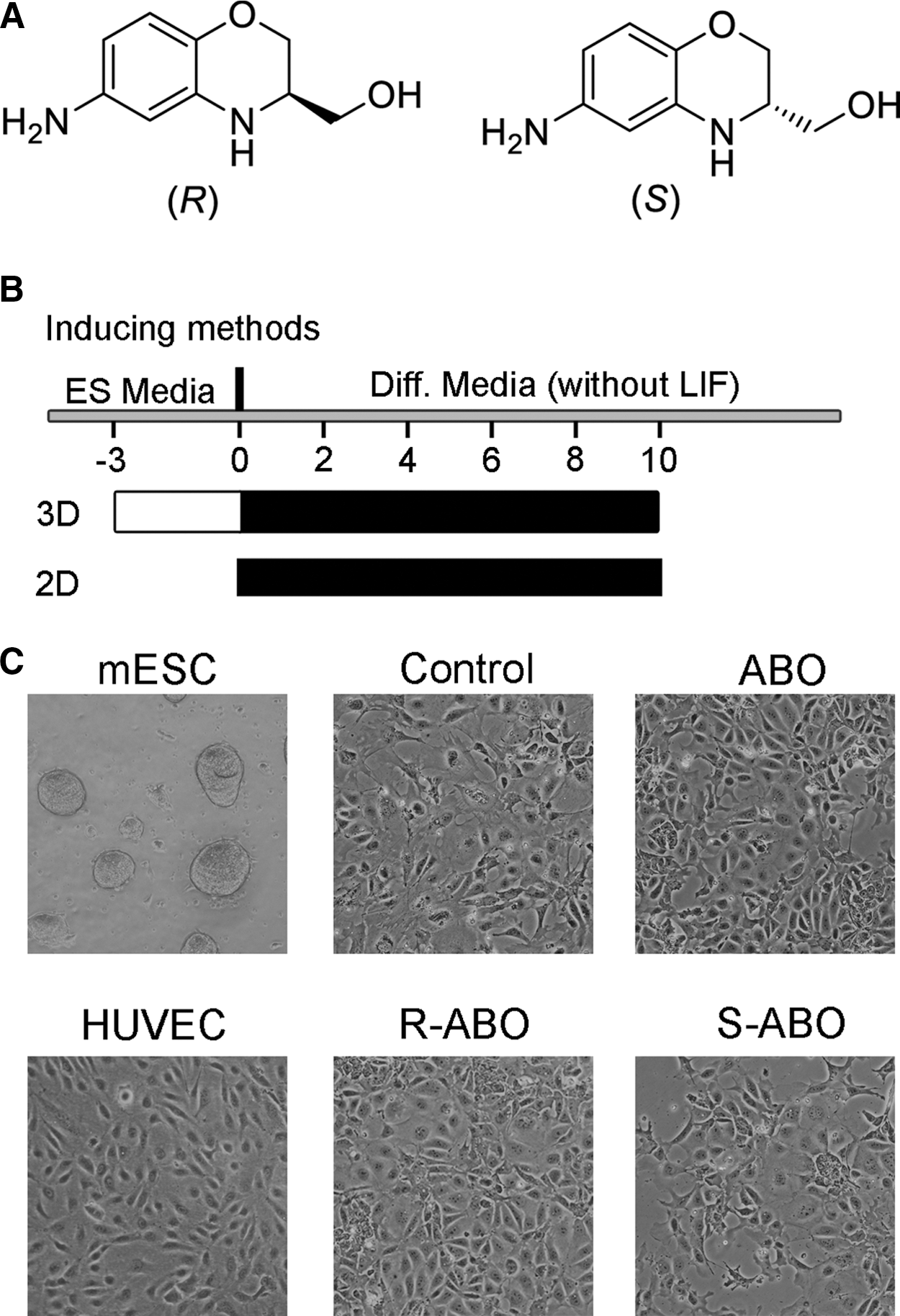
The small molecule ABO, R-ABO, and S-ABO induced ESCs' differentiation into endothelial-like cells.
R-ABO induced ESC differentiation into endothelial progenitor cells and mature endothelial cells
To demonstrate that R-ABO can induce ESC differentiation into VECs, we examined the levels of the specific markers expressed in vascular endothelial progenitor cells and mature endothelial cells, including CD133, CD31, VE-cadherin, antihemophiliac factor (AHF, also known as F VIII), eNOS, and VEGF receptor 2 (Flk-1) by immunofluorescence and western blotting. As CD31 is a common specific marker of mature VECs, we detected its expression in ESCs treated with 1, 10, or 20 μM R-ABO for 10 days. The results showed that R-ABO could most efficiently elevate the level of CD31 at 10 μM (Fig. 2A). Thus, the other specific markers of VECs were examined after ESCs were treated with 10 μM R-ABO. We found that the level of CD133 protein was increased at the sixth day and the eighth day (Fig. 2B), and the level of VE-cadherin was obviously increased at the tenth day (Fig. 2C, D). Otherwise, the levels of AHF, CD31, eNOS, and Flk-1 were increased in R-ABO-treated ESCs at the tenth day (Supplementary Fig. S1).
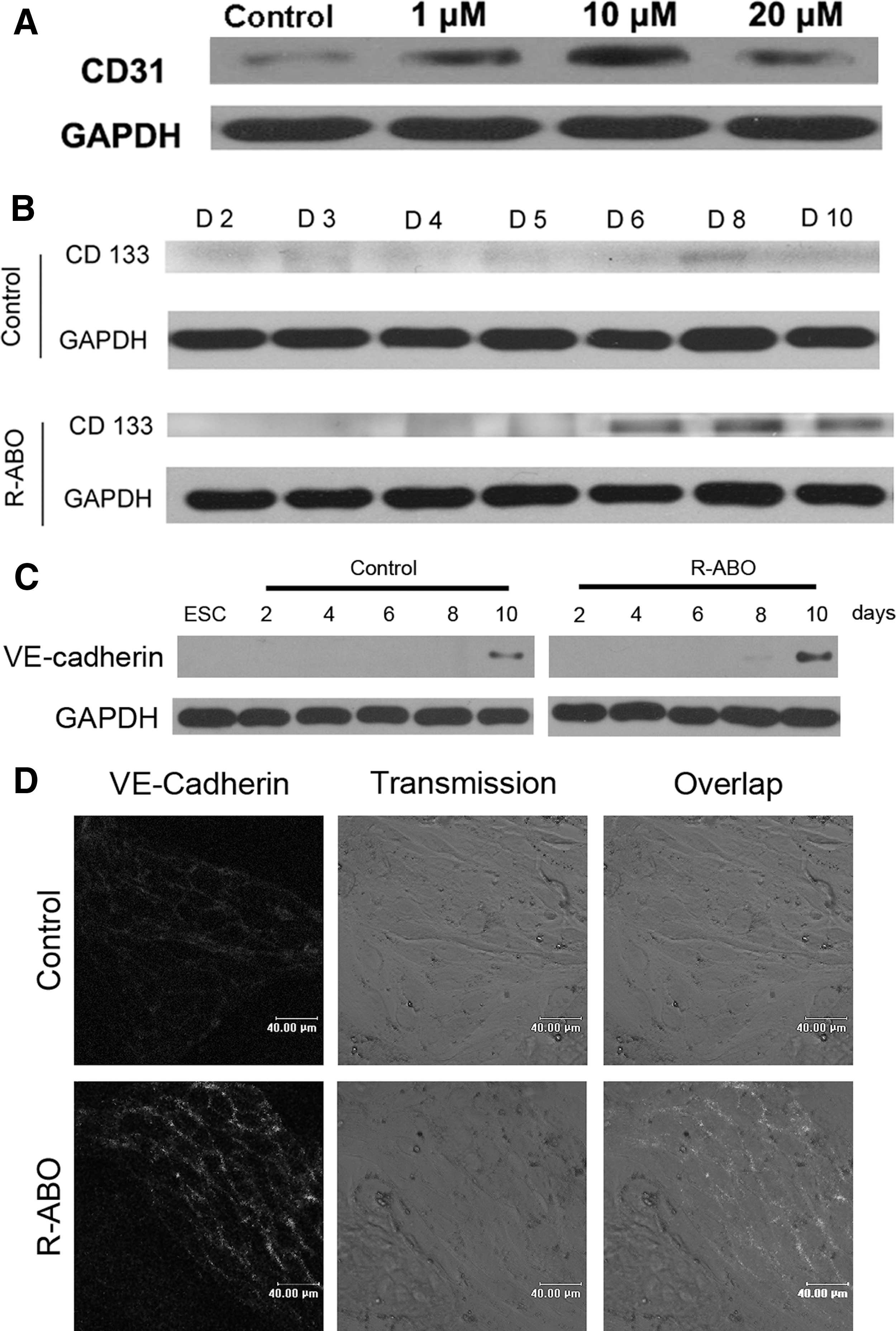
R-ABO-treated ESCs differentiated into endothelial progenitor cells and expressed mature endothelial cell markers.
R-ABO induced ESC differentiation into functional endothelial cells
To investigate R-ABO-induced ESC differentiation to functional VECs, we performed a capillary-like tube formation assay on Matrigel, which is a common method for identification of functional VECs. ESCs were treated with 1 to 20 μM R-ABO for 10 days and seeded on Matrigel-coated 24-well plates. The results showed that R-ABO effectively promoted the capillary-like structure formation at 10 μM (Supplementary Fig. S2 and Fig. 3A, B). To clarify which differentiation system is more effective to increase VEC production, we treated ESCs with 10 μM R-ABO at a 2-dimensional (2D) and 3-dimensional (3D) differentiation system for 10 days. The results of capillary-like tube formation assay showed that 2 kinds of differentiation systems have the similar effect (Supplementary Fig. S2). Because VECs are functionally defined by their uptake capacity of acetylated low-density lipoprotein from plasma [15], we investigated a number of Alexa Fluor 488-acetylated low-density lipoprotein-positive VECs in ABO-treated ESCs. As shown in Fig. 3C and Supplementary Fig. S3, the proportion of ESCs that differentiated into functional VECs was 45.4% after treatment with 10 μM R-ABO for 10 days. To further quantify the differentiation rate in our R-ABO-induced differentiation system, we performed FACS to quantify the CD31-positive cells. The FACS results showed that the rate of CD31-positive cells was 32.13% in the R-ABO treated group, which was much more than spontaneous differentiation rate (2.71%) in the control group (Fig. 3D). These data suggest that R-ABO promotes ESC differentiation into functional VECs.
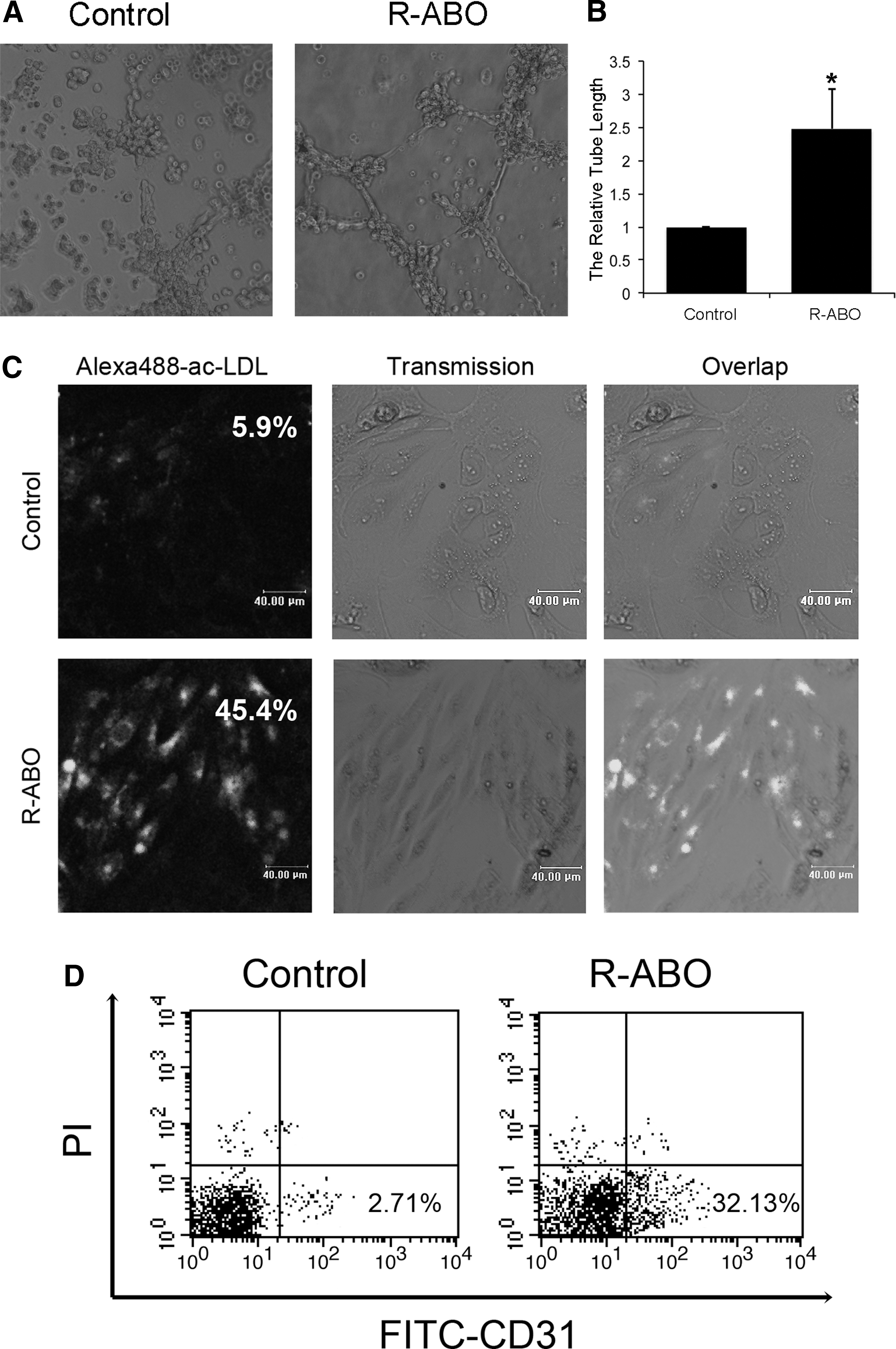
R-ABO induced ESC differentiation into functional endothelial cells.
Hmbox1 regulated ESC differentiation into functional endothelial cells
To understand the mechanism of R-ABO-induced differentiation of ESCs into VECs, we investigated the effect of ABO on the expression of homeobox containing 1 (Hmbox1) gene that was shown to be involved in BMSC differentiation into VECs induced by ABO in our previous study [9]. The results showed that R-ABO instead of S-ABO promoted Hmbox1 expression dramatically after treatment for 2 to 6 days (Fig. 4A–C). At the same time, the level of integrin beta4 (ITGB4), an important pluripotency-related integrin, was decreased significantly [16]. When Hmbox1 was knocked down in ESCs by its shRNA, R-ABO failed to promote CD133 expression (Supplementary Fig. S4 and Fig. 4D), and could not induce ESC differentiation into functional VECs (Fig. 5). It was reported that Hmbox1 acted as a potential transcription repressor [17 –21]. Here, we first report that it is a key regulator in ESC differentiation to VECs.
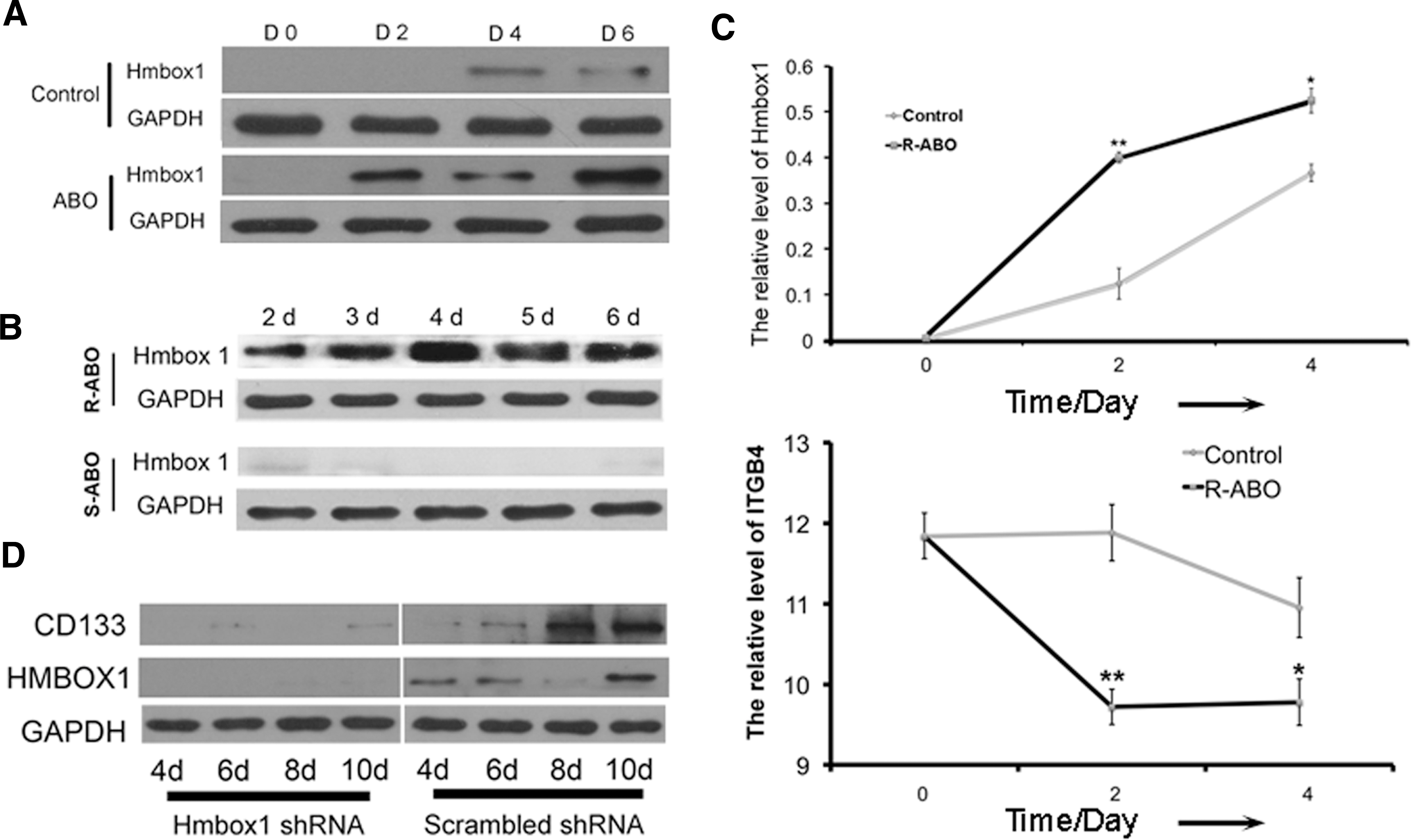
R-ABO promoted ESC differentiation into endothelial progenitor cells via Hmbox1.
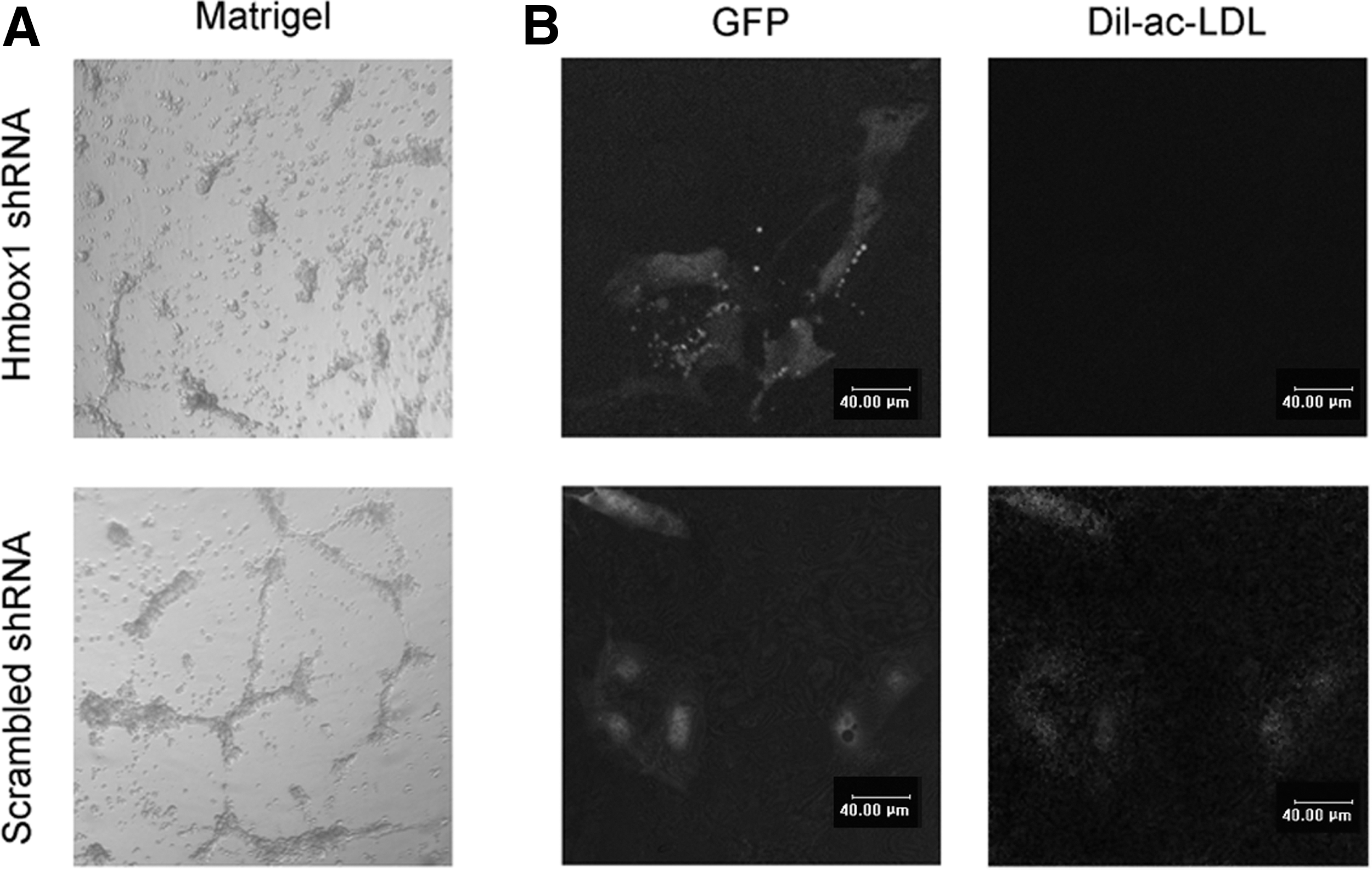
R-ABO failed to induce ESC differentiation into functional endothelial cells when Hmbox1 was knocked down. When Hmbox1 was knocked down in ESCs, R-ABO at 10 μM could not promote vascular structure formation at the tenth day
R-ABO elevated the FGF-2 protein level via Hmbox1
To understand whether R-ABO-induced differentiation of ESCs into VECs was through the classical proangiogenic pathways, we detected the expression of VEGF, FGF-2, and PDGF-BB, which are the important proangiogenic factors, by western blot. As shown in Fig. 6A and B, the expression of FGF-2 was most significantly increased at the fourth day compared with the control group. To further study the relationship between Hmbox1 and FGF-2, we examined the expression of FGF-2 during the differentiation of ESCs induced by R-ABO after Hmbox1 was knocked down by its shRNA. The results showed that FGF-2 could not be upregulated by R-ABO if Hmbox1 was knocked down (Fig. 6C, D). These data suggested that Hmbox1 acted upstream of FGF-2 in R-ABO-induced ESC differentiation into VECs.
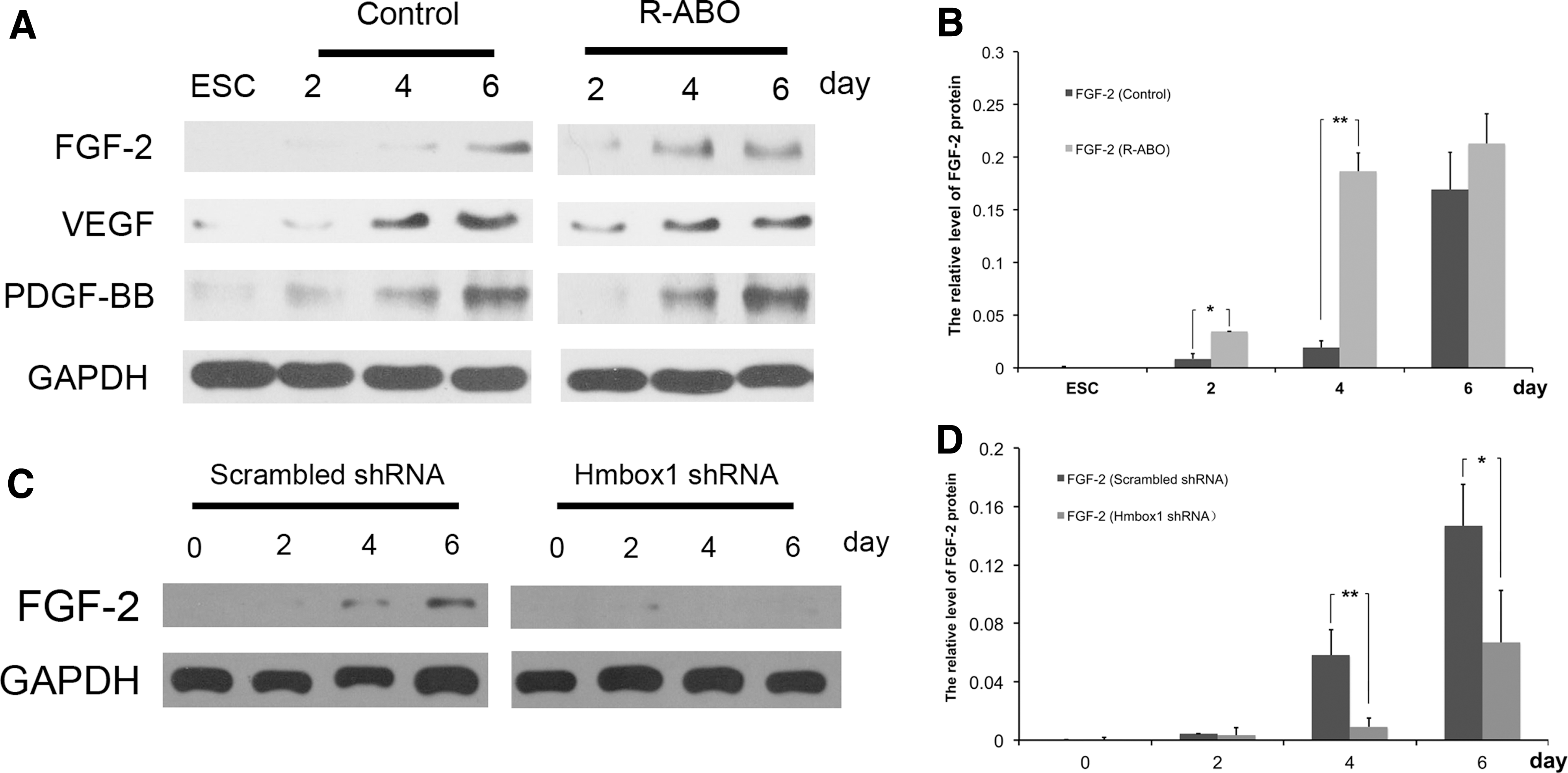
Hmbox1 acted upstream of FGF-2 in R-ABO-induced ESC differentiation into VECs.
Discussion
Previous studies have demonstrated the spontaneous differentiation of various hESC lines to VECs in 2D and 3D differentiation systems. In a 3D differentiation system, the spontaneous differentiation rate of ESCs into VECs is only 1%–3% [22,23], whereas with the methods based on Matrigel and conditioned media, 5%–10% of the resulting culture were endothelial progenitor cells [24]. The culture of human ESCs under 5% O2 conditions for 5 days promoted expression of endothelial markers in a significant proportion of the cells. The expression of markers characteristic of more mature endothelial cells was detected, including CD31 (−11.6%) and vWF (−31.9%), when the cultures were analyzed at later time points (8–15 days) [25]. A recent report showed that DNA methyltransferase inhibition by 5′-aza-2′-deoxycytidine (aza-dC) induced mouse ESC differentiation into VECs, which was the first report of a significant induction of ESC differentiation into VECs that did not involve the use of endothelial growth factors. The results showed that the proportion of cells expressing VE-cadherin protein, a specific marker of mature VEC, was 16% in ESCs treated with aza-dC. In our study, we found that a chiral small molecule R-ABO could induce ESC differentiation into VECs. On the basis of acetylated low-density lipoprotein uptake assay results, the proportion of ESCs that differentiated to functional VECs was 45.4% after treatment with R-ABO for 10 days. According to the FACS analysis results, the differentiation rate of ESCs into VECs induced by R-ABO for 10 days was 32.13%. Thus, the data suggest that our differentiation method is much more efficient at generating functional VECs.
Previously, we found that Hmbox1 was a key factor in BMSC differentiation into VECs [9]. However, the function of Hmbox1 in ESC differentiation into VECs is unknown. In this work, we provide the new evidence for Hmbox1 regulating ESC differentiation into VECs. Furthermore, we found that Hmbox1 acted upstream of a classical FGF-2 proangiogenic pathway in R-ABO-induced ESC differentiation into VECs. In our next work, we will clarify the regulation mechanism of Hmbox1 on the classical proangiogenic pathway.
In differentiation of human ESCs, ITGB4 was downregulated [16]. In keratinocytes, loss of alpha6 integrins could lead to an increase in expression of differentiation genes [26]. Our data showed that ITGB4 was decreased during the differentiation of ESCs. These data suggest that ITGB4 might be the key factor in the differentiation of ESCs. In our further investigation, we will focus on the relations of Hmbox1, ITGB4, and FGF-2 in ESC differentiation into functional VECs.
In summary, our data suggest that R-ABO is a novel tool for ESC differentiation into functional VECs, and Hmbox1 that acts upstream of FGF-2 is a key regulator in this differentiation process. These findings provide information on a novel target and a new platform for further investigating the gene control of ESC differentiation to VECs.
Footnotes
Acknowledgments
This work was financially supported in part by the National Natural Science Foundation of China (No. 90813022, 20972088, 31070735, 31000510, and 81021001), the National 973 Research Project (No. 2011CB503906), China Postdoctoral Science Foundation of special funding (201104637), the Natural Science Foundation of Shandong Province (ZR2010CZ004), and the Independent Innovation Foundation of Shandong University (2009JC007, 2009GN033, and 2009TS083).
Author Disclosure Statement
No competing financial interest exists.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.