
Abstract
Wharton's jelly mesenchymal stromal cells (WJMSCs) are considered mesenchymal, multipotent, and capable of differentiating into cells of mesodermal origin. Ectodermal differentiation from mesenchymal cells has been recently reported. Herein, we show for the first time that we can generate cytokeratin 19-positive cells and hair-like structures from WJMSCs in vitro using 2 separate methodologies that utilize osteogenic media to induce WJMSCs to undergo osteogenic differentiation. In one method, WJMSCs were seeded on a matrix isolated from Wharton's jelly following decellularization. In the other method, WJMSCs were cultured to form spheroids. Our findings demonstrate that WJMSCs may have the capacity for ectodermal differentiation.
Introduction
MSCs from fetal mice dermal tissue, when placed in a 3-dimensional (3D) culture condition, formed spheroids from 50 to 200 mm in diameter [14]. These spheroids could be induced to form hair-like structures. Interestingly, the sphere formation was essential to creating new hair structures. Based on this work, we hypothesized that hair structures could be created from MSCs originating from Wharton's jelly if they were exposed to stimulating media and in a 3D structure. We produced 3D spheroids from WJMSCs using 2 methods, one with a matrix material and one matrix-free, to determine their ability to create hair structures. In both methods, we used osteogenic media to induce WJMSCs to undergo osteogenic differentiation. Our findings highlight the importance of mesenchymal bone differentiation in driving ectodermal differentiation.
Materials and Methods
WJMSC isolation
IRB approval was obtained for human umbilical cord collection and WJMSC isolation from the University of Kansas Medical Center (HSC No. 12129). The procedure described by Wang et al. was used for WJMSC isolation and expansion [16]. Following these procedures, WJMSCs were plated in cell culture flasks and expanded to 80%–90% confluence then passaged until the 4th to 7th passage (P4–P7). Expansion media was composed of low-glucose Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum (FBS) and 1% penicillin and streptomycin (pen/strep) all purchased from Life Technologies.
WJMSC culture and osteogenic differentiation to generate spheroids
WJMSCs in expansion media were loaded into 24-well nontissue culture-treated plates and incubated at 37°C and 5% CO2. For each well 1×106 cells/mL were cultured in 1 mL of expansion medium. After 2 days, the medium was changed to the osteogenic medium, consisting of low-glucose DMEM supplemented by 100 nM dexamethasone (Sigma), 5 mM B-glycerophosphate (Sigma), 10 nM 1α,25-hydroxyvitamin D3 (Biomol International), and 50 μg/mL ascorbic acid 2-phosphate (Sigma). Exposure to osteogenic differentiation media continued up to 5 weeks.
DWJM scaffold preparation
Umbilical cords were immediately collected after normal vaginal delivery and placed in a premade transfer solution made of physiologic saline containing Amphotericin B, pen/strep at 4°C. Within 72 h, decellularization was noted. Pieces of fresh Wharton's jelly matrix were aseptically dissected from the cord, separated from the blood vessels in pieces (∼5×30 mm), and decellularized in a custom apparatus. The decellularization process included multiple osmotic shock cycles using hypertonic and hypotonic solutions, a nonionic detergent (Triton-X), and an anionic detergent (sodium lauroyl succinate) as well as an enzyme digestion with recombinant endonuclease (Benzonase™). All process residuals were removed with 36 h of iterative solvent washouts (double deionized water and 40% ethyl alcohol) dialyzed against hydrophobic and ion exchange resin beads (IWT® TMD-8, Amberlite™ XAD16, Powex™ 550A Biobeads, all from Sigma-Aldrich).Imaging of the decellularized matrix revealed the matrix material devoid of cellular elements with a very minimal average percent dsDNA residual (data submitted elsewhere). Before seeding, under a laminar flow hood, DWJM fragments were transferred to a large Petri dish and covered with phosphate-buffered saline (PBS). Using a sterile 5-mm skin punch biopsy kit, 5 mm in diameter pieces were isolated, and then washed with PBS twice. The height of the DWJM pieces ranged from 2 to 3 mm. When the WJMSCs were ready to be seeded, the DWJM pieces were moved to a nontissue culture-treated plate.
DWJM cell seeding and osteogenic differentiation
To reseed the DWJM, samples of DWJM were immersed in an expansion medium for 1 day at 37°C with 5% CO2. The medium was subsequently removed from the scaffolds. P4-P7 WJMSCs were then resuspended in a fresh medium and seeded by repeated pipetting into the DWJM scaffold at a 2×104/mm3 DWJM. This was followed by incubation on an orbital shaker at 50 rpm. After 4 h, the medium was added to completely immerse the scaffold in a total of 1 mL. After 2 days, the medium was changed to the osteogenic medium as described previously. Exposure to osteogenic differentiation continued for up to 5 weeks.
Histology
Spheroids
Spheroids generated from these experiments were scraped from the bottom of the culture well and transferred to PreservCyt Solution® (Cytyc Corporation (Hologic)) and processed using Cellient Automated Cell Block System®. Eosin was applied and vacuum-drawn through the sample to visualize the cells. After isopropyl alcohol dehydration and xylene clearing, the samples were embedded in paraffin.
DWJM
DWJM material was fixed in either formalin or 4% paraformaldehyde, embedded in paraffin, sectioned, and stained as described.
Alizarin red stain
Alizarin red stain was performed using the American Mastertech alizarin red stain kit per the manufacturer's instructions.
Collagen I stain
For collagen I stain, the anticollagen I antibody (ab34710) supplied by Abcam was used. Following epitope retrieval with a heated citrate buffer, pH 6.0, slides were stained with a primary antibody 100 μL at 1:200 dilution for 30 min followed by FLEX EnVision anti-rabbit HRP detection for another 30 min and FLEX DAB for 5 min.
Immunohistochemistry
The cytokeratin 19 (CK19) monoclonal mouse anti-human antibody (Dako) was used for immunohistochemical staining. Epitope retrieval was performed in Biocare Decloaking Chamber (pressure cooker), under pressure for 5 min, using the Dako EnVision Flex high pH buffer. Endogenous peroxidases were blocked with 3% H2O2. CK19 (clone RCK108) was used as the primary antibody for 20 min, followed by FLEX EnVision HRP detection for 30 min and FLEX DAB for 5 min. As an enhancing reagent, 2% CoCl2 was added for 5 min. All of the previously mentioned antibodies and reagents used were purchased from Dako. A hematoxylin counterstain was used.
The cytokeratin 15 (CK15) monoclonal rabbit anti-human antibody (Abcam) was used for immunohistochemical staining. Epitope retrieval was performed in Biocare Decloaking Chamber (pressure cooker), under pressure for 5 min, using the citrate pH 6.0 buffer. Endogenous peroxidases were blocked with 3% H2O2. CK15 (clone EPR1614Y) was used as the primary antibody for 60 min, followed by Dako EnVision anti-rabbit HRP detection for 30 min and Dako DAB for 5 min. A hematoxylin counterstain was used. Slides were reviewed using the Olympus BX40 microscope and pictures were obtained using the DP72 digital camera.
Fluorescence-activated cell sorting
For the WJMSC phenotype, the following primary and secondary antibodies (BD Biosciences) were used: anti-CD34-phycoerythroin (PE), anti-CD45-PE, anti-CD105-PE, anti-CD73-PE, anti-CD90-PE, and mouse IgG1-κ-PE isotype control. Flow cytometry was performed using FACScan (BD) and the data were analyzed using CELLQuest Version 3.2.1 (BD).
Western blot
Spheroids were lysed in the Radio ImmunoPrecipitation Assay (RIPA) buffer (Sigma-Aldrich), separated by SDS-PAGE gel electrophoresis, transferred to an ImmobilonPolyvinylidenedifluoride membrane (Millipore), and immunoblotted with a primary antibody against CK19 (No. MAB3238, 1:1,000; Millipore) and a secondary anti-mouse IgG-HRP (No. 7076, 1:40,000; Cell Signaling). ImageJ software was used to calculate the western blot's band density.
Results
Characteristics of decellularized WJ matrix
To evaluate DWJM for any remaining cells or any CK19-positive cells in specific, we stained DWJM samples for CK19. H&E stained sections revealed no intact cells and no CK19-positive cells were detected in DWJM (Fig. 1).
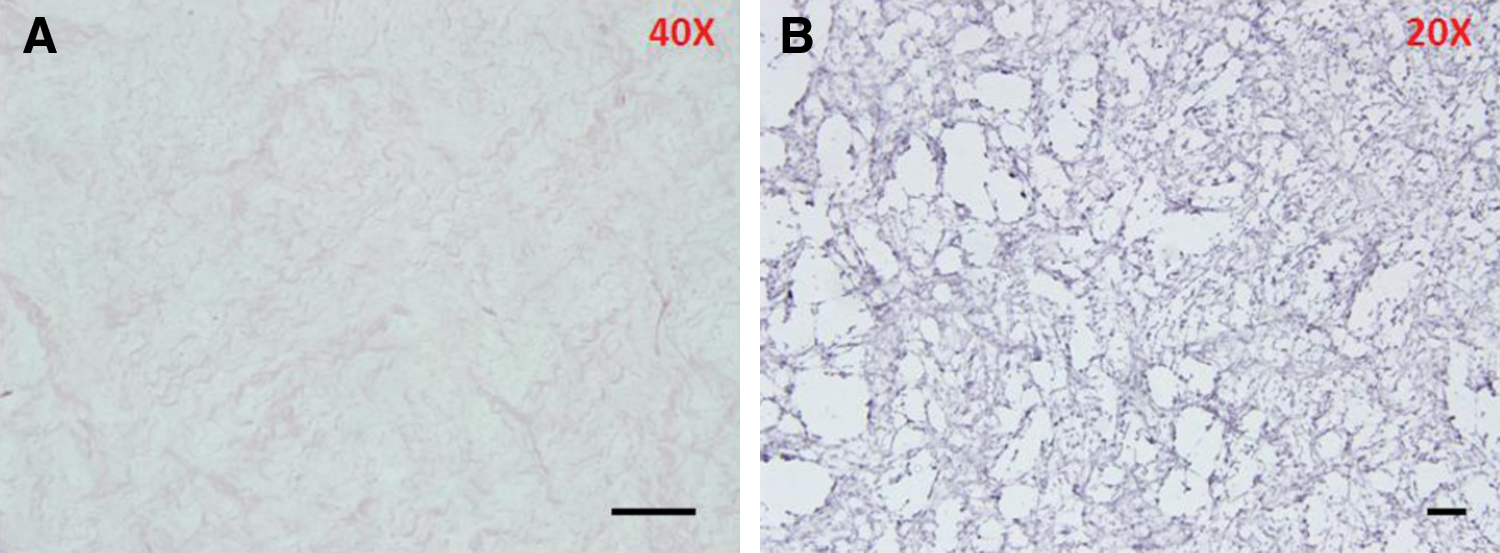
Decellularized Wharton's jelly matrix before seeding. This H&E stained section of DWJM
WJMSCs expressed MSC phenotype
Isolated and expanded WJMSCs were evaluated for the characteristics of MSCs as defined by the International Society for Cellular Therapy's recommended minimum criteria [17]. For immunophenotype, flow cytometry tests confirmed that the isolated and expanded WJMSC population was negative for CD34 and 45 and positive for CD73, CD90, and CD105, consistent with the MSC profile (Fig. 2). In addition, the isolated WJMSCs were selected based on their adherence to plastic surfaces in culture flasks. Finally, the isolated WJMCs demonstrated evidence of osteogenic differentiation based on positive alizarin red staining (described later) following their exposure to the osteogenic differentiation medium. Together, these findings support the conclusion that the isolated and expanded WJMSCs represent true MSCs.
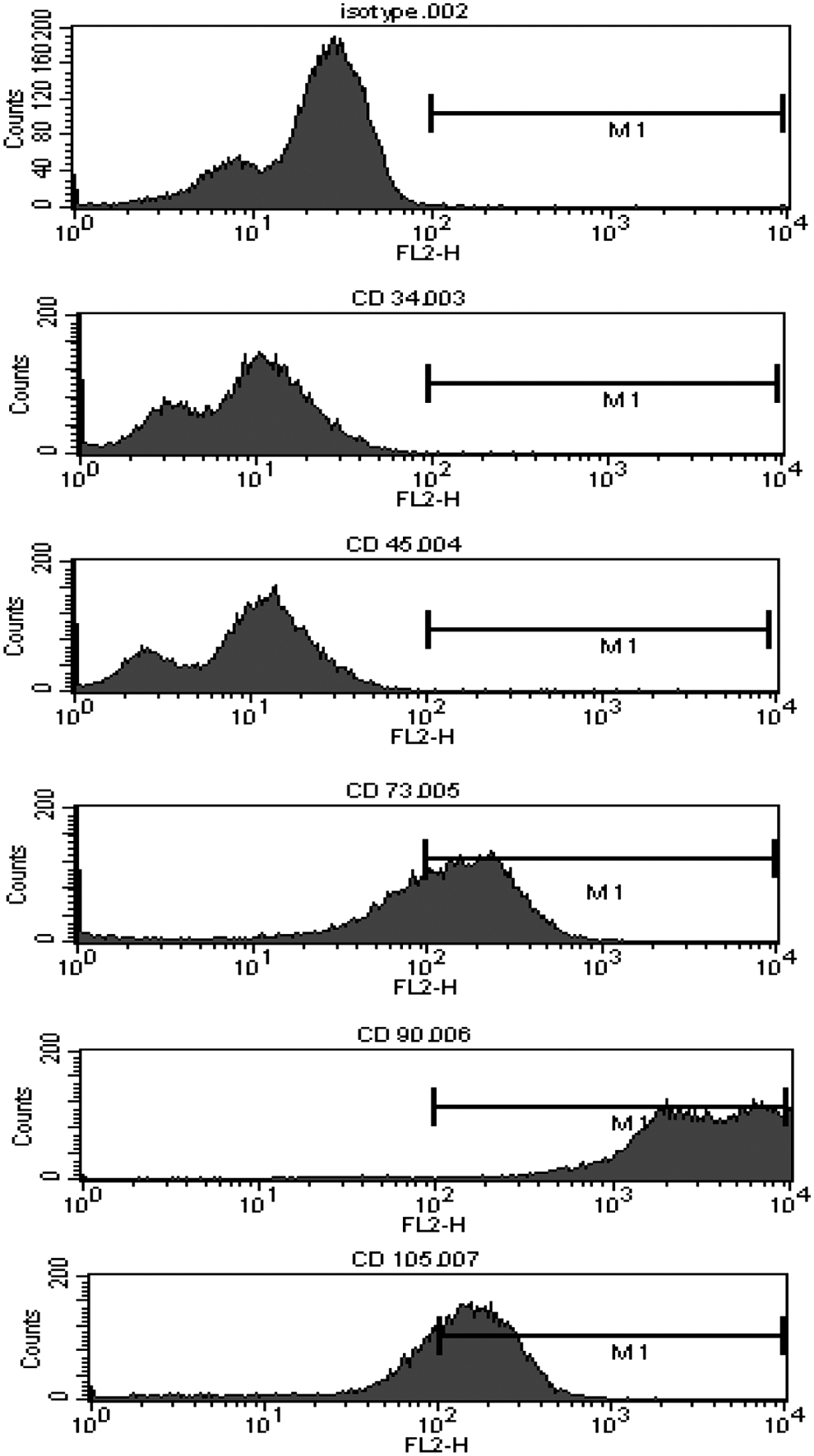
Flow cytometry profile of expanded Wharton's jelly mesenchymal stromal cells (WJMSCs). WJMSCs following expansion and before use in our experiments were CD34- and CD45-negative, while positive for CD73, CD90, and CD105.
Detection of hair-like structures during osteogenic differentiation in DWJM
WJMSCs were seeded on DWJM followed by osteogenic differentiation. Two weeks following induction of osteogenesis, hair-like structures were reproducibly protruding to or through the outer layer of the matrix material (Fig. 3). In some areas, these structures appeared thicker and straight protruding through the outer layer of cells (Fig. 3A). The structures were around 100 μm in width. In other areas, the hair-like structures appeared thinner and coiled underneath the outer layer of the matrix material (Fig. 3B).The thinner structures were around 30 μm in width. We evaluated these structures under a dissecting microscope. We confirmed our earlier findings and noticed the hair-like structures again. They were either protruding through the outer layer of the matrix in the case of the thicker and the more straight structures (Fig. 3C–E), or simply caused an outside protrusion (Fig. 3F). These structures were ≤100 μm in diameter, which is consistent with the diameter of hair as reported in the literature [18].
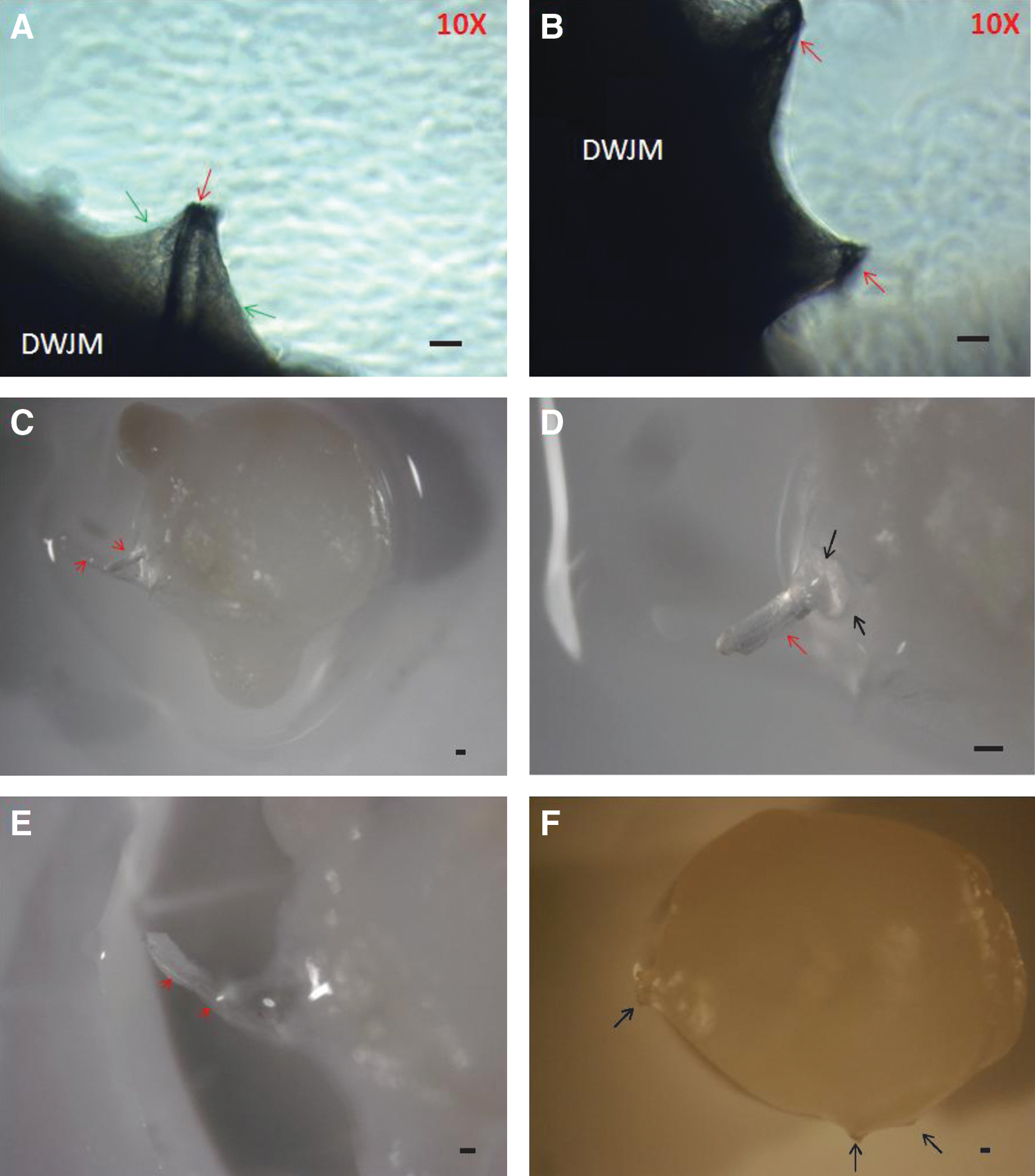
Patterns of hair-like structures in DWJM. Hair-like structures were observed growing out of DWJM 2 weeks following WJMSC seeding and osteogenic induction. In phase-contrast microscopy pictures
Hair-like structures in WJMSC spheroids underwent ostoegenic differentiation
To rule out an effect of the DWJM, the cells were cultured without the DWJM. WJMSCs were cultured in nontissue culture plates; similar to earlier experiments, we induced osteogenic differentiation using the same osteogenic medium components. In the WJMSC spheroids a similar phenomenon occurred, where hair-like structures were found on histologic sections of these spheroids (Fig. 4). The hair structures were never found in the core of the spheroid, but rather were always located close to the surface. Within the same peripheral region of the spheroid and adjacent to the hair-like structures were large regions of calcification (red arrows). Staining with alizarin red clearly highlighted the areas of calcification, also found along the periphery of the spheroid (Fig. 4).
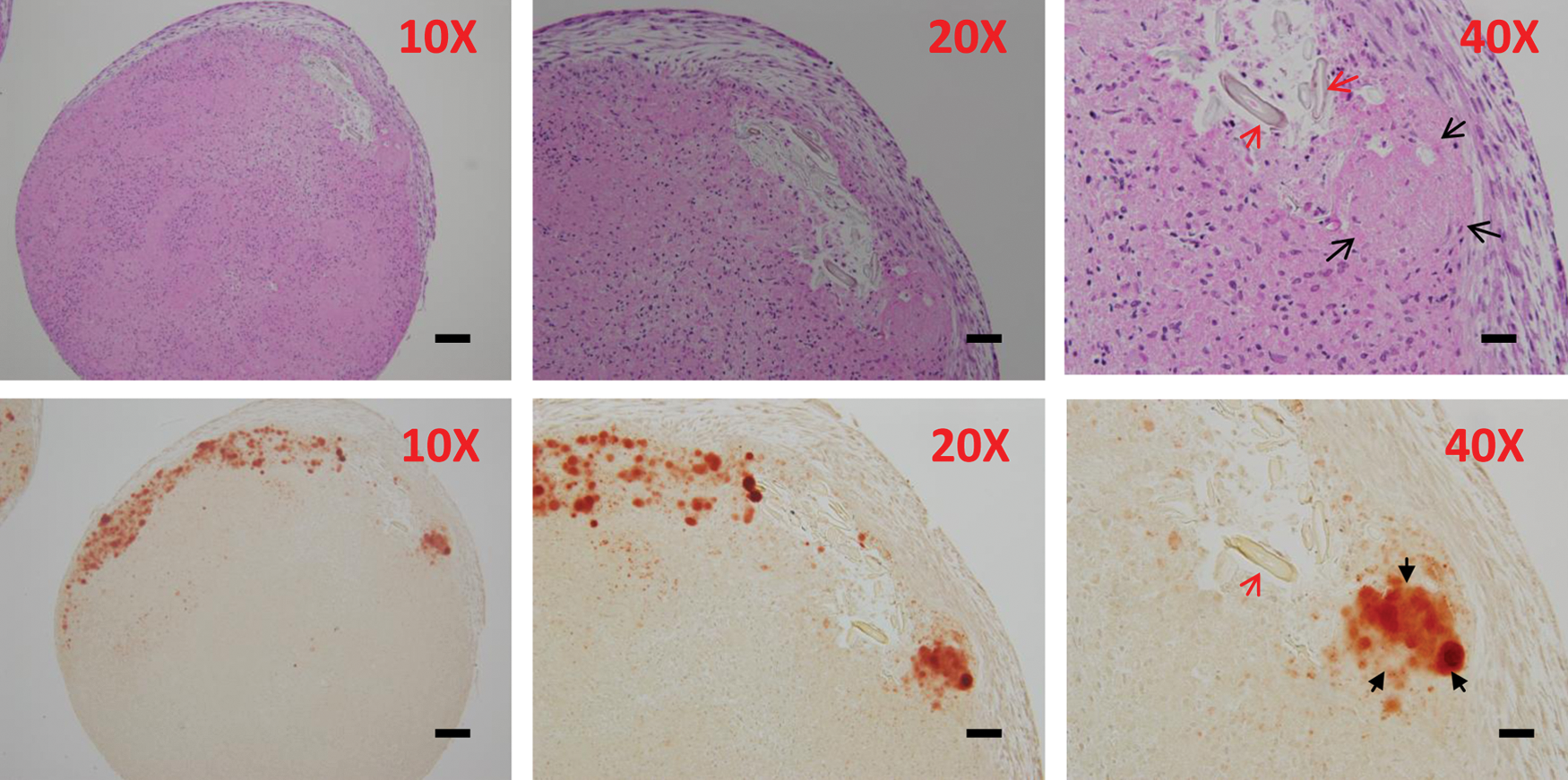
Hair structures and CK19-positive cells in osteogenic differentiated WJMSC spheroids. In the upper panel, H&E stained slides demonstrated areas of calcification (black arrows) in close proximity to hair-like structures (red arrow) within the osteogenic differentiated spheroids. In the lower panel, alizarin red stained slides demonstrated areas of mineralization (black arrows) within the calcified areas. The areas of mineralization appeared to be in close proximity to the hair structures (red arrow). Scale bar represents 100, 50, and 25 μm in 10×, 20×, and 40×. Color images available online at
Expression of CKs in cells associated with hair-like structures
Since hair is an ectodermal structure made of keratins by hair follicles, expression of CK15 and CK19 can be used as biomarkers of hair follicles. CK19 is considered a marker of hair follicle stem cells [19]. Likewise, CK15 is considered one of the markers of the bulge cells in human hair follicle [20]. These bulge cells have properties of stem cells as they have high proliferative capacity and are multipotent capable of regenerating ectodermal structures, such as hair follicles, sebaceous glands, and epidermis [21].
Spheroid structures expressed CK19-positive cells in a diffuse pattern
In WJMSC spheroids, CK19-positive cells were expressed in an abundant although diffuse fashion (Fig. 5). These CK19 expressing cells appeared to coalesce producing a centrally located clear area on cross sections (Fig. 5). In the case of WJMSC spheroids, we confirmed CK19 expression by western blot analysis (Fig. 5). Interestingly, the expression of CK19 clearly increased over time following exposure to osteogenic media (Fig. 5). These findings suggest that exposure of WJMSCs in their spheroids to ostoegenic differentiation media led to further differentiation of WJMSCs to CK19 positively expressing cells.
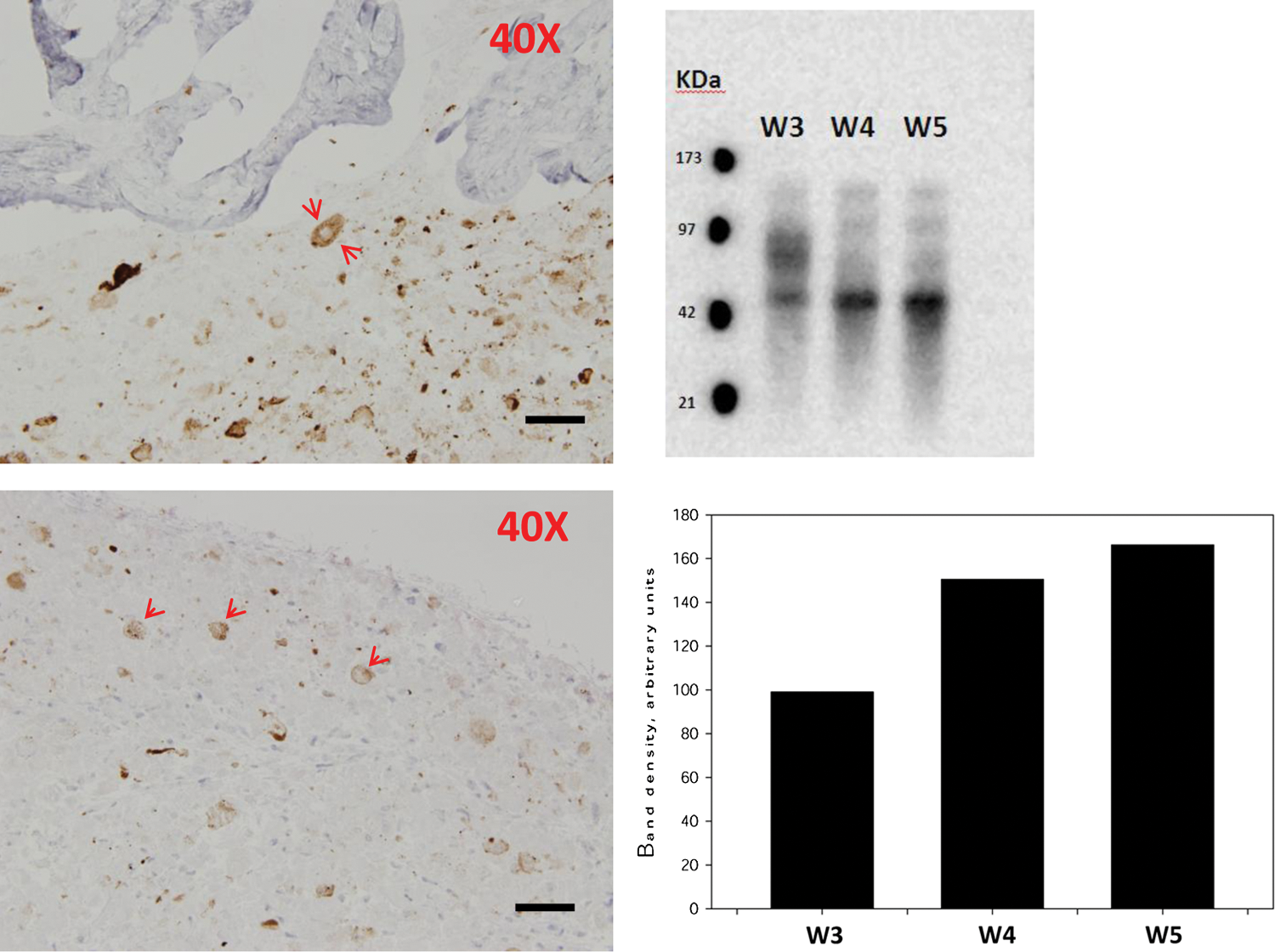
CK19-positive cells in DWJM and spheroids resemble hair follicle structure. CK19-positive cells coalesced and formed round structures with central clearings, resembling hair follicle structure in both DWJM (left upper) and spheroids (left lower) (red arrows point to these structures). Scale bars represent 50 μm. CK19 expression was confirmed by western blot at 3, 4, and 5 weeks of osteogenic differentiation time points (right upper). A band with approximate molecular weight of 40 KDa was detected. The calculated western blot's band density for weeks 3, 4, and 5 of osteogenic differentiation using ImageJ is shown (right lower). Band density for CK19 expression clearly increased over time. Color images available online at
DWJM expressed CK19-positive cells in a more discrete fashion
In DWJM, CK19 expression was predominantly closer to the surface of the DWJM and in discrete areas between the surface and what appears to be a bone matrix material evident by collagen I staining (Fig. 6E). We stained for collagen I as it is considered the most abundant bone matrix protein [22]. Alizarin red staining, on the other hand, was negative in DWJM. However, the Alizarin red stained sections brought out the large structures of CK positivity with central clearing. In some sections, there were no indications of hair structures, although the CK19 expressing cells were seen. These CK19-positive cells appeared coalesced and produced a centrally located clear central area resembling the structures seen in spheroids.
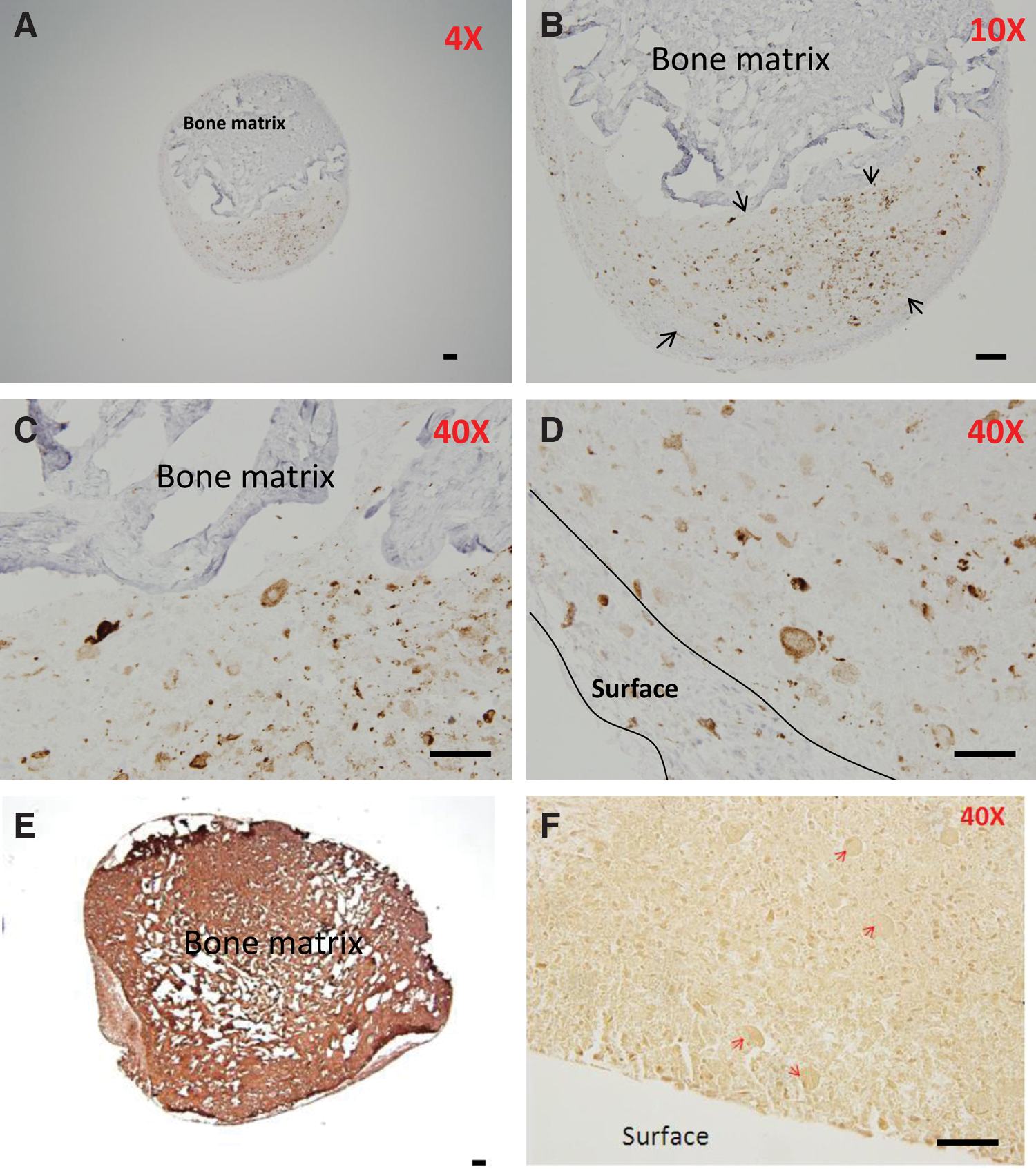
Pattern of CK19 expression in DWJM following osteogenic differentiation. In these pictures, CK19-positive cells appeared to concentrate predominantly in 2 areas; around the DWJM structure as seen in
In DWJM tissue sections, bulbous structures were noted. These structures that resemble the hair follicle communicated with the outside of the DWJM structure (Fig. 7). These areas were closely associated with CK19 expressing cells (Fig. 7). Hair-like structures were associated with CK19-positive cells. These hair-like structures were either straight or coiled (Fig. 8), but strongly positive for CK19 in the case of the latter. Positive staining for CK19 confirms that the structures represent keratinized material. On histologic sections, the straight hair ranged from 4 to 53 μm in width and the coiled hair ranged from 12 to 24 μm in width.
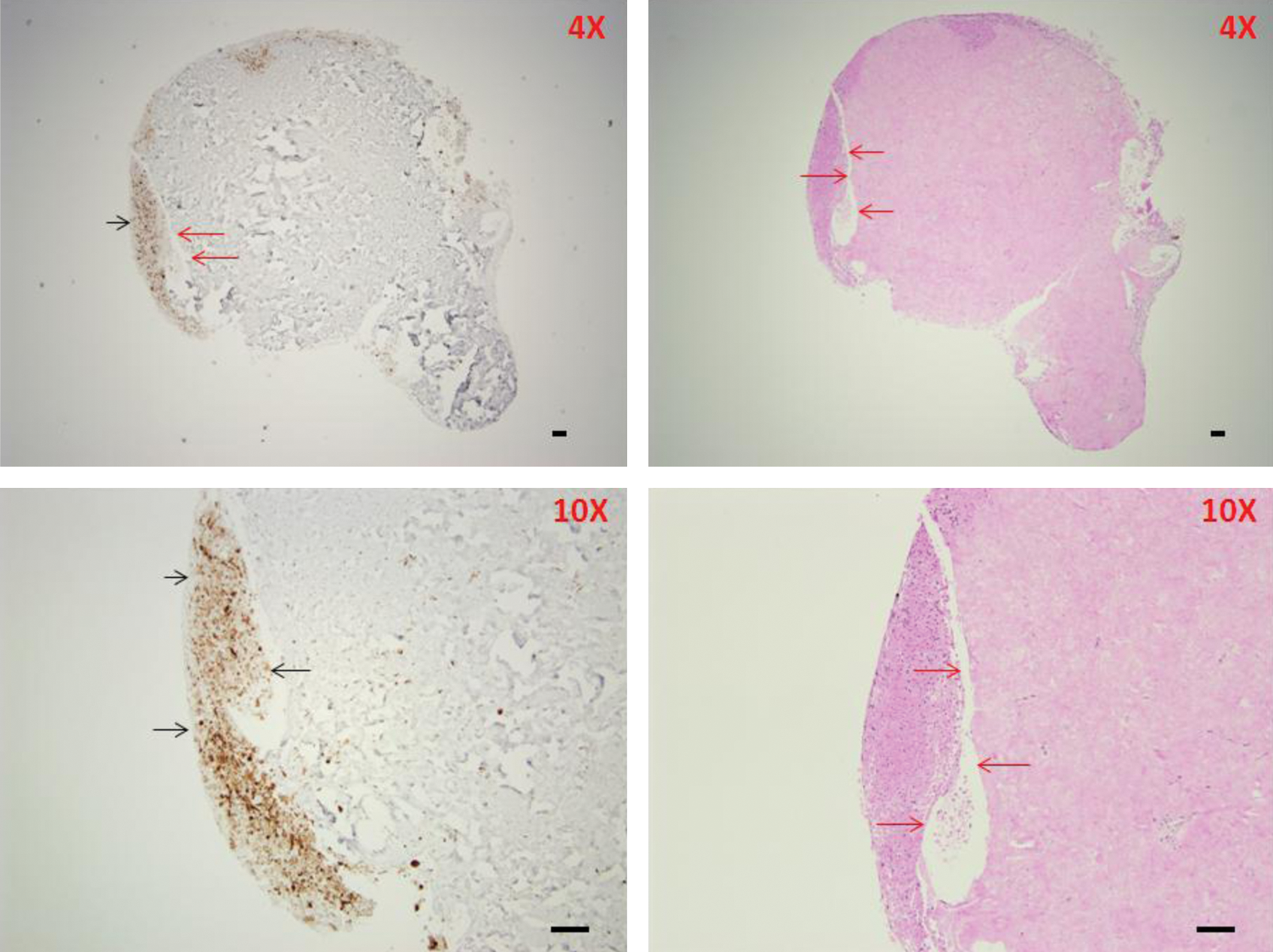
Areas of defects in DWJM resembling hair follicle structure associated with CK19 expression. Defect areas (outlined by red arrows) within the DWJM were seen in H&E-stained and CK-stained sections. These areas resembled hair follicle structure. The hair structures were not identified in these sections. We felt that specimen processing lead to their expulsion from DWJM. CK19 expressing cells were associated with these defects (black arrows). Objective lens magnifications are included in the graphs. Scale bars represent 100 μm. Color images available online at
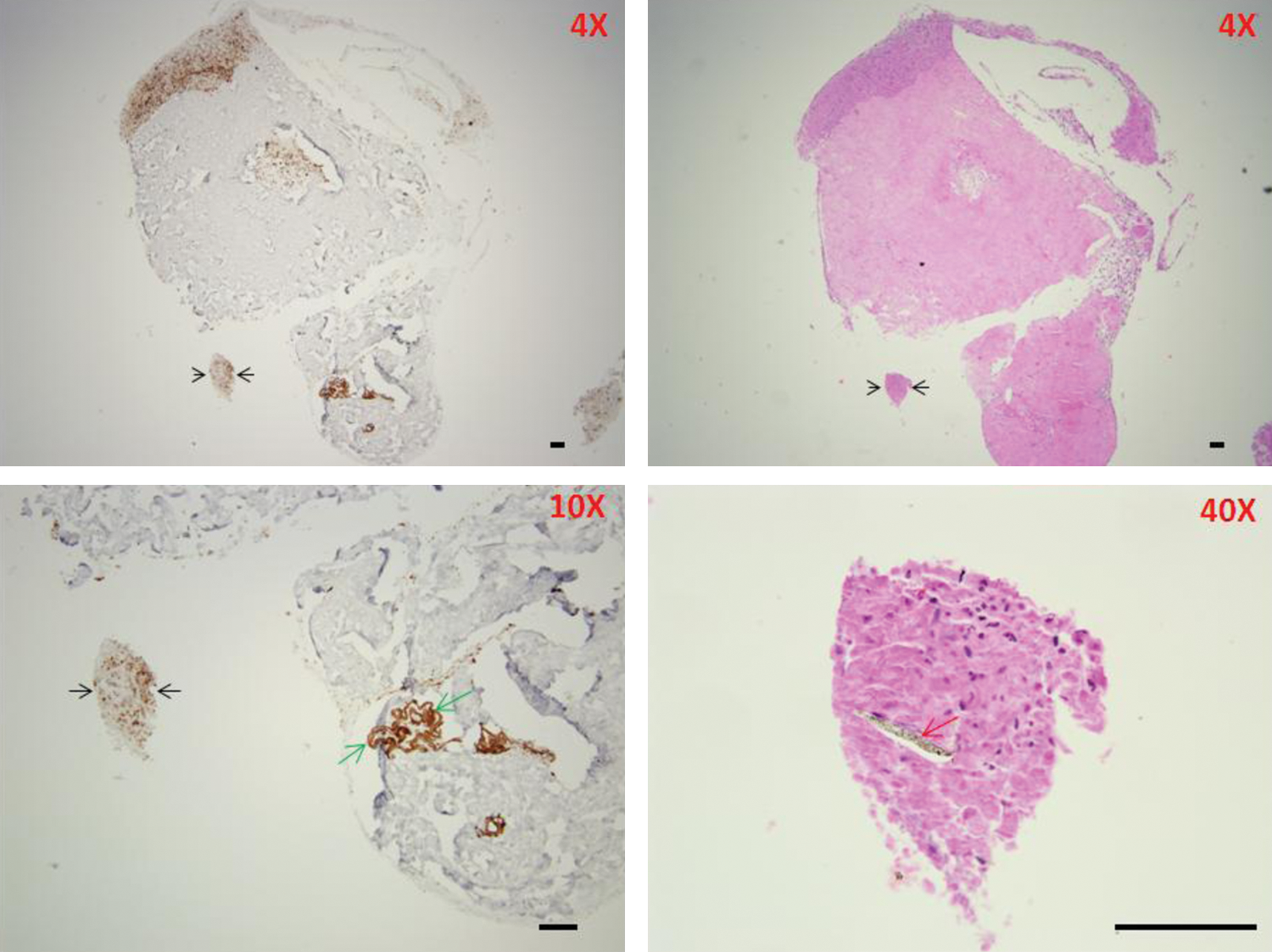
Hair-like structures identified in DWJM associated with CK19 expression. A CK19 strongly expressing collection of cells (black arrow) was found to include a straight hair-like structure (red arrow). On the other hand, a coiled CK19-positive hair-like structure was observed in another section (green arrows). Scale bars represent 100 μm. Color images available online at
CK15 and CK19 expressing cells in DWJM
Interestingly, very few cells/structures stained positive for CK15. These CK15-positive cells/structures were located in sections, where the more abundant CK19-positive cells were observed (Fig. 9). This spatial closeness is a clue that these cells might truly reflect hair follicle cells. In addition, coiled hair-like structures were seen in close proximity to CK19-positive cells, which provides another clue that these were hair follicles capable of producing keratinized material.
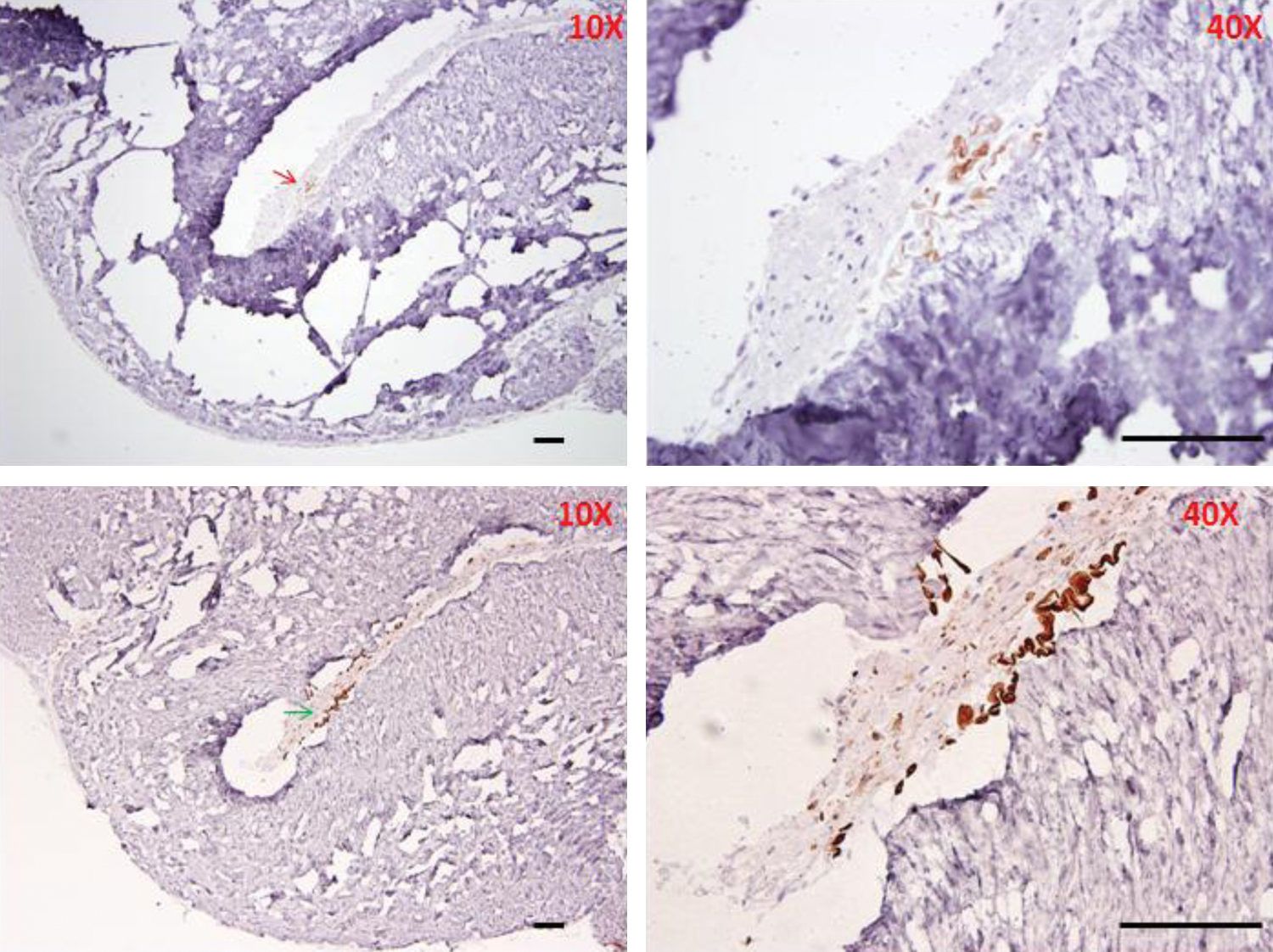
CK15 and CK19 staining of DWJM sections. The upper panel represent CK15-stained sections and the lower panels represent CK19-stained sections of DWJM. In these figures, CK15-positive cells/structures (red arrows) appear to be seen in the same sections, where CK19-positive cells/structures (green arrows) were identified. However, the frequency of CK15-positive cells appears to be less than CK19-positive cells in these sections. Scale bars represent 100 μm. Color images available online at
Discussion
In these experiments, we report for the first time that CK19-positive cells and hair-like structures can be produced from WJMSCs with or without using a matrix material. In the first methodology, we generated CK19-positive cells diffusely arranged within spheroids. On the other hand, using a matrix material, we generated CK-positive cells localizing to certain areas of the DWJM. In some areas of spheroids, we observed that these cells were close to hair-like structures, an association that led to the conclusion that the generated CK19-positive cells might reflect hair follicle cells. Generating epithelial cells from mesenchymal cells has been recently reported [10 –13]. For example, Baer et al. generated epithelial cells from adipose-derived adult MSCs [12]. Wan et al. also reported the generation of epithelial cells from bone marrow-derived MSCs [13].
To provide an explanation for such phenomena, the link between bone and hair formation is important. In our experiments, we found the hair structures localized in close proximity to areas of calcification and mineralization, evidenced by alizarin red stain in spheroids and to areas of collagen I matrix material in DWJM. There is some evidence suggesting that bone formation is connected to hair growth [23]. In a study by Mundy et al., the use of proteasome inhibitors stimulated both bone formation and hair growth utilizing the same mechanisms. Hedgehog-BMP-Wnt was suggested as the common molecular mechanism. We believe that the DWJM provided a CK19-positive cell–collagen I matrix interface that promoted hair growth. This interface resembles the epithelial–mesenchymal interaction that is essential for morphogenesis of various organs, including hair follicles [24]. In our case, CK19-positive cells represent the epithelial component and the collagen I matrix with osteogenic differentiated cells represent the mesenchymal component.
We are not the first to report that WJMSC differentiated into an ectodermal structure. First, CK-positive WJMSCs have been reported in the literature, especially in the perivascular space around the umbilical blood vessels [2]. In addition, a subpopulation of WJMSCs was also found to express primitive stem cell markers [25]. Finally, Yoo et al. generated dermal papilla-like tissue (DPLT) from umbilical cord MSCs following exposure to the dermal papilla forming medium. These generated DPLT, in turn, self-aggregated [26]. When transplanted to athymic mice, these DPLT induced new hair follicle formation. Collectively, these observations and our findings provide evidence that a population of WJMSCs might be capable of ectodermal differentiation. These findings and ours provide a clue that MSCs from umbilical cord tissue can generate hair. In our experiments, we generated CK19-positive cells arranged in what appears to be a hair follicle closely associated with hair. To generate these cells, we followed a totally different procedure that is based on the use of the osteogenic differentiation medium.
Shimizu et al. reported that the sphere formation was necessary for MSCs to develop the hair-inducing capacity [14]. In our case, we recognize the importance of sphere formation in the case of spheroids, though it was not necessary in the DWJM. It may be that it is not the actual spheroid that is essential, but that any 3D organization of the cells lends itself to hair development. Additionally, we found that osteogenic differentiation was the common factor in the 2 methodologies described in our case. In summary, we have found that the 3D WJMSC could be differentiated into hair-like structures using osteogenic differentiation media.
Footnotes
Acknowledgments
We thank Lynda Bonewald for her advice on osteogenic differentiation and her review of the manuscript. We also thank Marsha Danley who prepared the specimens for histologic examination, Lesya Novikova who helped with dissecting microscopy pictures, and Anna Ludlow who helped in proof-reading and formatting the manuscript. O.S.A. is a recipient of a research career award by the Office of Scholarly, Academic & Research Mentoring (OSARM) at home institution.
Author Disclosure Statement
No competing financial interests exist.