
Abstract
Although the role of Notch has been studied extensively in the developing nervous system, the embryonic lethality of Notch pathway mutants has hindered studies in the adult brain. The creation of cre/lox-mediated conditional gain- and loss-of-function mice has allowed us to investigate the role of Notch signaling in adult neural stem and progenitor cells. We have determined that Notch signaling is important for conferring stem cell characteristics upon neural precursor cells. Knocking-out Notch signaling in vivo results in neural progenitors, leaving the subependymal niche and migrating along the rostral migratory stream to the olfactory bulb, while overexpressing Notch results in retention of cells in the subependyma. Further, increased Notch signaling in progenitor cells resulted in the expression of stem cell markers in vivo as well as conferring the characteristics of self-renewal and multipotentiality upon subsequent isolation in vitro. Similar to what has been reported from the embryonic brain, the overexpression of Notch in neural precursor cells in vitro increased the numbers of neurospheres from the adult brain. Finally, overexpression of Notch1 in pure populations of progenitor cells (excluding neural stem cells) isolated by fluorescence activated cell sorting led to the formation of multipotent, self-renewing neurospheres from the non-neurosphere forming fraction. Hence, Notch overexpression confers stem cell properties upon progenitor cells and demonstrates that Notch signaling not only preserves stem cell characteristics, but that it can confer stem cell characteristics upon a subset of progenitor cells.
Introduction
S
The Notch signaling pathway has been little studied in the adult brain despite the fact that the stem cell lineage is well characterized, thereby facilitating an examination of the role of signaling molecules [7,8]. Stem cells in the adult brain reside in the subependyma (SE) of the periventricular lining of the lateral ventricles (LV). The slowly dividing stem cells divide to give rise to rapidly dividing progenitor cells that migrate to the olfactory bulb (OB) along the rostral migratory stream (RMS) where they differentiate into OB interneurons. Notch is expressed in the neurogenic regions of the adult brain, including the SE, and the generation of transgenic mice with conditional activation or suppression has made it possible to overcome the early embryonic lethality of Notch mutants [9]. The adult brain provides an excellent model to test the intriguing possibility that Notch signaling plays a role not only in preserving stem cell identity into adulthood, but also the possibility that Notch signaling is sufficient to impart stem cell properties on more committed neural progenitor cells. This would be consistent with recent findings in other systems demonstrating that a small number of genes, including genes linked to stem cell fate in the respective systems, are sufficient to revert cells to early cell types in their lineage [10 –12].
We have used transgenic mice to manipulate Notch signaling in adult neural stem and progenitor cells both in vitro and in vivo. We found that Notch signaling increases stem cell numbers by preserving stemness in neural stem cells and by conferring stem cell properties upon progenitor cells.
Materials and Methods
Retrovirus preparation and infection
The constructions of replication-incompetent retrovirus vector pMXIE have been described previously [6]. A sequence encoding Cre recombinase with a nuclear localization signal was inserted into pMXIE to generate nlsCre-pMXIE (a kind gift from Derek van der Kooy's lab, hereafter called Cre-GFP-RV). A packaging cell line, PlatE, was transfected transiently with the retroviral plasmids and pVSV-G plasmid using Fugene6 (Roche) and cultured at 32°C. The supernatant containing viruses was filtered with a 0.45 μm filter and concentrated by ultracentrifugation. The viruses were titrated on NIH3T3 cells by counting green fluorescent protein-positive (GFP+) cells in serial dilutions. Viral concentrates were diluted to an equal concentration (1×107) and frozen in aliquots at −80°C until used. For in vitro infection, dissociated cells from adult IC-Notch1 [13], Notch1 cKO [14], or IC-Notch1 (−/−) littermate control mice SE derived neurospheres were infected with multiplicity of infection (MOI) ∼1.0 with Cre-GFP-RV at 37°C for 8 h. The retroviruses were then washed out and the cells were plated at clonal density (10 cells/μL). To assess self-renewal, primary infected neurospheres were mechanically dissociated into single cells and plate at clonal density (10 cells/μL). Stem cell self-renewal was assessed by identifying new neurospheres after a further 7 days in vitro. For in vivo infections, adult IC-Notch1, Notch1 cKO, or IC-Notch1 (−/−) littermate control mice received unilateral intraventricular injections of 1.0 μL of Cre-GFP-RV at 0.1 μL/min using a 30-gauge Hamilton syringe at the following coordinates: anterior 0.2 mm, lateral 0.8 mm relative to bregma, and depth 2.3 mm below the dura. The needle was left in the position for 10 min after the injection.
Animals
Mice were 8–10 weeks of age. Transgenic IC-Notch1 [13] and Notch1 cKO [14] mice were bred in house. IC-Notch1 (−/−) littermate control mice were used as controls. CD1 mice (Charles River) were used to study survival in the ZVAD experiments. All animals were housed at the University of Toronto animal facilities and maintained in accordance with the Animal Care Committee and Institutional guidelines.
Isolation and culture of adult SE-derived neural stem cells
Adult brains were dissected as described previously [15]. Briefly, mice were killed via cervical dislocation and their brains were excised. Medial and lateral portions of lateral ventricle SE were dissected and placed in artificial cerebrospinal fluid. The tissue was digested with 1.33 mg/mL trypsin (Sigma), 0.67 mg/mL hyaluronidase (Sigma), and 0.2 mg/mL kynurenic acid (Sigma) at 37°C for 20 min. Tissue was transferred into a serum-free medium containing 0.7 mg/mL trypsin inhibitor (Roche) and triturated with a fire-polished Pasteur pipette. Cells were plated at 10 cells/μL in 24-well (0.5 mL/well) uncoated plates (Nunc) in a serum-free Dulbecco's modified Eagle's medium (DMEM)/F12 medium containing 10 ng/mL fibroblast growth factor (Sigma), 20 ng/mL epidermal growth factor (Sigma), and 2 ng/mL heparin (Sigma). ZVAD (0.1, 1, 10, and 20 μM) (R&D Systems) or BrdU (0.6 μM) (Sigma) were added to the media as indicated in the text. Cell survival was assessed by trypan blue exclusion as indicated in the text, using the Countess Automated Cell Counter (Invitrogen) according to the manufacturer's instructions.
Side-population cell analysis and flow cytometry
The side-population (SP) sorting was conducted as described previously with slight modifications [16]. Dissociated adult neurosphere cells were incubated in serum-free medium (DMEM:F12) supplemented with 5 μg/mL Hoechst 33342 (Sigma), and incubated at 37°C for 1 h. Cells were washed once in serum-free media and proprodium iodide (Molecular Probes) was added to a final concentration of 2 μg/mL to exclude dead cells from the analysis. For blocking experiments, verapamil was added at 50 μM final concentration (Sigma) at the beginning of Hoechst staining. Cells were analyzed on Dako Cytomation MoFlo equipped with a UV laser and a 488 nm laser. For noninfected cells, the sorting was gated on total live cells. For retrovirus infected cells, the sorting was either gated on total live cells or gated on total GFP+ live cells. Two populations, SP and non-SP, were sorted based on different lower blue and red fluorescence emissions. Sorted cells were either used for RNA isolation and reverse transcription-polymerase chain reaction (RT-PCR) or neurosphere culture. SP and non-SP cells were plated at clonal densities (10 cells/μL) in standard neurosphere conditions as described. The numbers of neurospheres were counted after 10–14 days in vitro.
RT-PCR
Total RNA was isolated using RNeasy extraction kit (Qiagen) and RT-PCR was performed using one-step RT-PCR kit (Qiagen) according to manufacturer's instructions. The sense and antisense primers were used as follows [6]: Jag1, sense 5′-GTCCACG GCACCTGCAATG-3′; antisense 5′-CAAGGTTTGGCCTCG CACT-3′; Notch1, sense 5′-CCAGCATGG CCAGCTCTGG-3′, antisense 5′-CATCCAGATCTGTGGCC CTGTT-3′; Hes1, sense 5′-AAAGACGGCCTCTGAGCACA-3′, antisense 5′-TCATG GCGTTGATCTGGGTCA-3′; Hes5, sense 5′-AAGTACCGTG GCGGTGGAGATGC-3′, antisense 5′-CGCTGGAAGTGG TAAAGCAGCTT-3′; Beta-actin, sense 5′-AGGCCAACCGTGAAAAGATGAC-3′; antisense 5′-GTACATGGTTGGTACCACCAGAC-3′. Primers were designed to encompass at least 1 intron to avoid false-positive amplification of contaminated genomic DNA. qPCR was conducted using the Solaris qPCR Gene Expression Assay (ThermoFisher Scientific/Dharmacon) according to the manufacturer's instructions and using proprietary primers for Nestin, Id1, and GAPDH (used as reference control) (ThermoFisher Scientific/Dharmacon).
Immunocytochemistry
Seven days after primary culture, single neurosphere colonies were mechanically dissociated into single cells, and the cells were divided into 3 wells of a 24-well culture plate (Nunclon) coated previously with Matrigel (0.6 mg/mL in serum-free medium) (Becton-Dickinson) in 0.5 mL/well of the medium containing 1% fetal bovine serum (Gibco). After 7 days in vitro, cells were fixed with 4% paraformaldehyde for 20 min at room temperature, and then each of the 3 wells was processed using one of the following antibodies: anti-βIII tubulin (1:400; Sigma), anti-GFAP (1:400; Sigma), and anti-O4 (1:400; Chemicon/Millipore), followed by appropriate Alexa secondary antibodies (Roche). Cultures were counter stained with the nuclear stain Hoechst 33258 (1 μg/mL; Sigma). For brain sections, mice were overdosed with anesthetic and perfused transcardially with 4% paraformaldehyde. Brains were removed and postfixed overnight, cryoprotected with 20% sucrose in phosphate buffered saline at 4°C, and then cryosectioned (20-μm sections). Sections were immunostained using mouse monoclonal anti-GFP (Chemicon/Millipore) at 1:500. The numbers of retrovirally labeled (GFP+) cells were counted throughout the entire hemisphere from the OBs rostrally and extending caudally to the rostral crossing of the anterior commissure. Cells were grouped into 3 categories based on their location: (1) SE cells in the periventricular region of the LV, (2) RMS cells, which are found along the pathway from LV to OB, and (3) OB cells confined to the OB. For further analysis of brain sections, anti-ki67 (1:300; Abcam), anti-GFAP (1:400; Sigma), and anti-DCX (1:500; Abcam) antibodies were used with Alexa secondary antibodies (Roche).
Statistical analysis and experiment design
All data are presented as mean±SEM. P values for t-test or analysis of variance are indicated in the text. For in vivo experiments, n values indicate the number of mice. For in vitro experiments, n values indicate the number of independent experiments performed. Individual experiments used the pooled cells of >2 mice and represent the average of triplicate wells.
Results
Active Notch1 signaling confers stem cell characteristics upon progenitor cells in vivo
In the adult brain, stem cells comprise ∼0.5% of the cells in the SE. These slowly dividing stem cells have an estimated cell cycle time of ∼15 days [17] and divide asymmetrically to give rise to rapidly dividing progeny with an estimated cell cycle time of 13 h in the rodent brain [18]. In vivo lineage analysis reveals that the majority of progenitor cells die, while those that survive migrate rostrally and differentiate into OB interneurons [18]. We confirmed the expression of Notch1 (receptor), Jagged1 (ligand), and Hes1/5 (target genes) in the SE using RT-PCR on dissected SE tissues (Supplementary Fig. S1A; Supplementary Data are available online at
We took advantage of 2 transgenic mice strains to examine the role of Notch signaling in the adult brain: (1) the IC-Notch1 mouse, which coexpresses the constitutively active intracellular domain of Notch1 (NICD) as well as an enhanced GFP reporter after Cre-mediated excision [13] and (2) the Notch1 conditional knockout (Notch1 cKO) mouse that has lox P flanked Notch1 (thereby permanently knocking out Notch1 after Cre-mediated excision) [14]. We modulated Notch signaling in the SE in vivo by stereotactically injecting a retrovirus expressing Cre-recombinase and GFP (Cre-GFP-RV) directly into the right LV of IC-Notch1 +/− or Notch1 cKO+/+ transgenic mice. Littermate IC-Notch1 −/− mice serving as controls (Fig. 1A). As progenitor cells constitute the vast majority of dividing cells (>99%) and are rapidly proliferating in the SE, the retrovirus should exclusively transfect progenitor cells [15,17,]. To verify that the retrovirus infected progenitor cells and not the stem cells, we dissected control (IC-Notch1 −/−) mice on days 3 and 12 postinfection. We examined every neurosphere that formed and found that none of the neurospheres contained GFP+ cells (n>5 animals/group). Further, we stained brain sections from each of the 3 groups (IC-Notch1, Notch1 cKO, and control) 3 days postinfection and found that >40% of the GFP+ cells were colabeled with the progenitor marker DCX in each group (n>300 GFP+ cell, n=3 animals/group) (Supplementary Fig. S1A). Finally, we did not see any infected cells colocalize with the stem cell marker GFAP at this time point (n>300 GFP+ cell, n=3 animals/group). Accordingly, we predict that if Notch signaling is important for stem cell identity, in vivo transfection to enhance Notch signaling would lead to reduced migration of transfected SE cells and a lengthening of the cell cycle time, both of which are characteristic of neural stem cells. Conversely, we would predict that if endogenous Notch1 is important for preserving stem cell characteristics, deleting it would decrease the retention of transfected cells in the SE. To test these hypotheses we quantified the numbers of GFP-expressing (GFP+, retrovirally transfected) cells in the SE, RMS, and OB at 3, 6, and 12 days after Cre-GFP-RV injection (Fig. 1A, B). Notably, modifying Notch signaling did not dramatically alter the general pattern of labeled cells, in that GFP+ cells were never observed outside the SE, RMS, and OB. In addition, in contrast to previous work showing that Notch promotes a glial cell fate [20,21], no GFP+/GFAP+glial cells were observed in the OB of either transgenic strain at any time-point examined. In all strains the absolute number of GFP+ cells was similar at day 3 (142±12 control; 117±10 Notch1 cKO; 101±21 IC-Notch1, total GFP+ cells/brain). As predicted, the GFP+ cells were localized to the SE at this early time (Fig. 1A, B). Further, there was no difference in cell death within the SE as determined with caspase-3 staining (not shown). Together, these data indicate that rate of infection and cell survival were similar in transgenic and control mice. We examined the distribution of the GFP+ cells over time and found that in control mice, 15.2%±1.5% of the wild-type GFP+ cells remained in the SE (ie, ∼85% of GFP+ cells were found in the RMS or OB) by day 6. In the Notch1 cKO mice, a smaller proportion of the cells remain in the SE by day 6 (8.3%±0.9%), with more found on the RMS, indicating that the progenitor cells leave the SE sooner in the absence of Notch1. In contrast, IC-Notch1 mice had 43.7%±15.3% of the GFP+ cells confined to the SE at day 6, indicating that Notch overexpression increased the number of cells that were retained in the SE. In all strains the total number of GFP+ cells increased between day 3 and 6 in the SE (5.8±0.94-fold increase vs. 6.4±0.18-fold increase vs. 4.2±1.0-fold increase in control; Notch1 cKO; IC-Notch1, respectively). Interestingly, the expansion of GFP+ cell numbers in the IC-Notch1 mice was significantly less than controls or Notch1 cKO mice, consistent with a decreased rate of proliferation or increased cell death within the GFP+ cells. By day 12 the total numbers of GFP+ cells declined significantly in the Notch1 cKO and control mice (67.3%±8.0% in control; 65.1%±3.8% in Notch1 cKO), with no significant decrease observed in the Notch1-IC mice (Fig. 1A, B). In addition, the majority of GFP+ cells were found in the OB of all groups, indicating that cells are still able to migrate and differentiate even in the absence of Notch1. Most interesting is the finding that in the IC-Notch1 mice there was a 2-fold increase in the percent of GFP+ cells in the SE compared to littermate controls (17.1%±8.2% vs. 8.7%±0.9%, IC-Notch1 vs. littermate control) (Fig. 1B) corresponding to a 2.8-fold increase in the absolute number of labeled cells in the SE of IC-Notch1 mice compared to wild-type controls. Hence, a subset of the infected cells are retained in the SE for longer times in the IC-Notch1 mice relative to Notch1 cKO or controls.

Activation of Notch signaling in adult neural progenitors in vivo.
We next sought to determine if any of the Notch infected cells that remain in the SE after 12 days also have the cardinal stem cell properties of self-renewal and multipotentiality. To this end, the SE of Cre-GFP-RV-infected IC-Notch1 mice were dissected on day 12 and plated in the standard neurosphere assay. Neurospheres are clonally derived colonies of stem and progenitor cells that arise from a single neural stem cell [22]. Given that Cre-GFP-RV infects the abundant, rapidly dividing progenitor cells in the SE, we predicted a complete lack of GFP+ neurospheres from IC-Notch1 (−/−) littermate control-infected mice. Indeed, we never saw any GFP+ neurospheres from control mice, consistent with our earlier observation indicating that the retrovirus did not infect any stem cells in vivo. However, we did observe GFP+ neurospheres from IC-Notch1 mice infected brain (Fig. 1C). These GFP+ neurospheres derived from IC-Notch1 mice could be dissociated and passaged to generate secondary neurospheres (demonstrating the stem cell property of self-renewal) and generated neurons and glia upon differentiation in vitro (demonstrating multipotentiality) (data not shown). The frequency of formation of GFP+ neurospheres from IC-Notch1 mice infected brain was ∼1 neurosphere per 300 in vivo infected cells, indicating that only a subpopulation of the progenitor cells are able to give rise to neurospheres in response to Notch signaling. Consistent with this, in vivo immunostaining of day 12 brain sections revealed that a small fraction of the GFP+ cells (1.5%±0.2%) in the IC-Notch1 SE expressed markers of neural stem cells including the expression of GFAP (expressed in neural stem cells but not in progenitor cells [23,24] and Ki67, which labels cells that are not in G0, including neural stem cells [25]. Notably, no GFP+/GFAP+/Ki67+ triple-positive cells were present in the littermate controls (Fig. 1D). To determine if the activation of Notch signaling was causing or preventing the cells from leaving the cell cycle, we determined the number of GFP+ cells that retained the expression of Ki67, a marker of cells that are in a proliferative state (although not necessarily dividing). We found that by day 12, the proportion of GFP+ cells expressing Ki67 was lower than on day 3 in both IC-Notch1 and control mice (9.4%±6.7% vs. 11.1%±8.4% decrease in proportion of GFP+ cells that are Ki67+, IC-Notch1 vs. control, n=3 animals/group). However, there was no difference between the 2 treatment groups (Supplementary Fig. S1B). This indicates that Notch overexpression does not directly affect the cells ability to remain in cycle in the SE. Together, these data are consistent with the hypothesis that Notch overexpression can confer stem cell properties upon a subpopulation of the progenitor cells. To further examine the hypothesis that Notch signaling plays a role in regulating stem cell identity, we next examined neural stem and progenitor cells in vitro using the neurosphere assay.
Notch1 regulates neurosphere number in vitro
Neurospheres are comprised of a mixed population of stem (<1%) and progenitor (>99%) cells [26]. Similar to the SE in the adult brain, SE-derived neurospheres express Notch receptor (Notch1) ligand Jagged1, and the target genes Hes1/5, as detected using RT-PCR (Supplementary Fig. S2A). To examine the role of Notch signaling in pure populations of neural precursors, neurosphere cultures were first established from IC-Notch1 and littermate controls, and then neurospheres were collected, dissociated, and replated in neurosphere conditions in the presence of Cre-GFP-RV. We observed a >50% increase in the numbers of neurospheres from IC-Notch1 cells infected with Cre-GFP-RV compared to Cre-GFP-RV-infected littermate control mice (Fig. 2A). This increase in neurosphere number is consistent with the in vivo observation that Notch-infected progenitor cells, which would not otherwise form multipotent, passagable neurospheres, were able to do so with active Notch signaling. Indeed, if Notch conferred a change in cell-cycle time or a change in the mode of stem cell division, neither of these changes would increase the numbers of neurospheres that formed. Hence, the data support the hypothesis that Notch overexpression confers stemness upon progenitor cells. Consistent with a role for Notch signaling in stem cell identity, qPCR analysis reveals increased expression of Nestin and Id1, genes associated with stem and progenitor cell identity [27], in the IC-Notch1-infected cells as compared to control (Supplementary Fig. S2B).
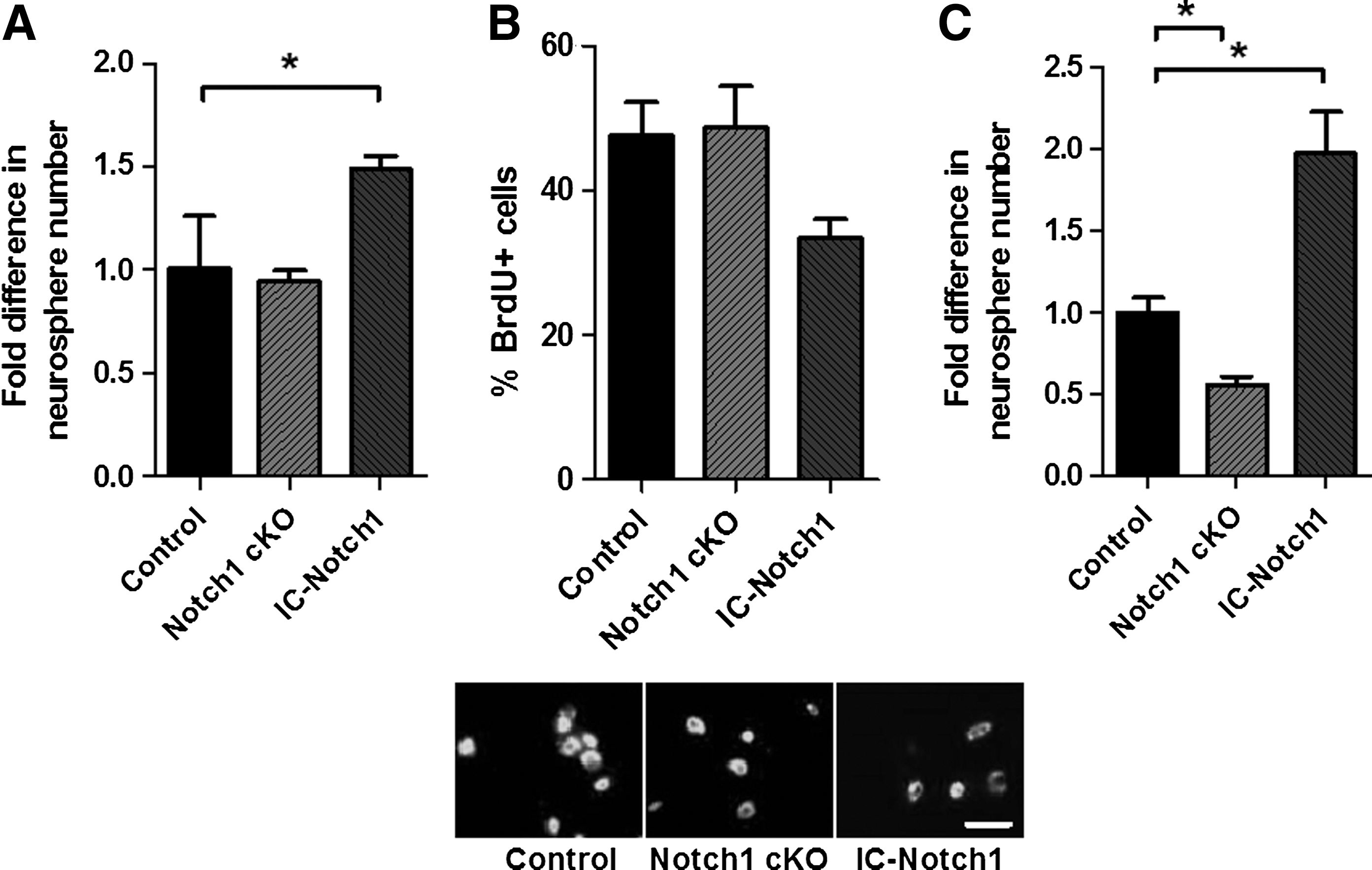
Modulating Notch signaling in vitro.
One alternative explanation for the increased numbers of neurospheres observed in vitro is that Notch expression enhanced neural stem cell survival. To examine a possible change in survival in response to Notch signaling, we compared the number of Cre-GFP-RV and control GFP-RV-infected cells 8 h after plating from IC-Notch1 mice. We observed no change in the overall number of surviving cells (10.3%±0.3% vs. 13.1%±1.9% surviving, Cre-GFP-RV vs. littermate controls, n=2 independent experiments, P>0.05). These data suggest that cell survival does not account for the increased numbers of neurospheres; however, given that stem cells account for only a small fraction of the neurosphere-derived cells, this data cannot formally rule out the possibility that Notch1 enhances survival exclusively in the neural stem cell fraction. To determine if here is a selective cell survival effect when Notch is overexpressed, we used ZVAD (a pan-caspase inhibitor) [28] to block cell death in vitro. SE-derived cells were plated in neurosphere media in the presence of a range of ZVAD concentrations (0.1–25 μM). At 48 h after plating, the number of surviving cells was counted (using trypan blue exclusion). The presence of ZVAD significantly increased the number of surviving cells, with the maximum effect occurring at a ZVAD concentration of 10 μM (45.0%±0.5% increase vs. 0 μM ZVAD in the number of live cells relative to the cell numbers at 1 h postplating, n=3 independent experiments, P<0.05). However, the number of neurospheres surviving at day 7 postplating was not changed by the presence of ZVAD at any concentration examined, including 10 μM, which showed the greatest overall cell survival (48.6±1.8 neurospheres/5,000 cells vs. 51.4±4.0 neurospheres/5,000 cells in control vs. 10 μM ZVAD, n=3 independent experiments, P<0.05). These findings suggest that Notch signaling does not increase neurosphere number simply by increasing stem cell survival.
When neurospheres from Notch1 cKO mice were infected with the Cre-GFP-RV to ablate Notch1, there was no change in the numbers of neurospheres that formed compared to GFP-RV-infected control (Fig. 2A). Hence, endogenous Notch signaling does not affect the ability of stem cells to form neurospheres. In all instances, GFP+ neurospheres were multipotent and the treatments had no effect on the differentiation potential in that the frequency of astrocytes, oligodendrocytes, and neurons were similar between groups (Supplementary Fig. S2C).
We observed that IC-Notch1-derived neurospheres were smaller than control neurospheres, consistent with a reduction in total cell number, potentially the result of a decrease in progenitor cell proliferation (50.4±3.5 μm diameter vs. 63.3±5.3 μm diameter, IC-Notch1 vs. control. n>100 neurospheres, 2 independent experiments. P<0.05). We directly examined the effect of Notch signaling on the proliferation of the cells by exposing primary neurosphere-derived cells from 4 day cultures to BrdU then fixing the cells 4 h post-BrdU. Although ablating Notch1 (Notch1 cKO) had no effect on proliferation, we observed a significant decrease in proliferation in the infected Notch1-overexpressing cells (from IC-Notch1) (Fig. 2B), indicating that Notch signaling decreases progenitor cell proliferation, consistent with our in vivo observations.
We reasoned that if Notch signaling is important for stem cell identity, it would increase the number of self-renewing divisions undergone by Notch1-overexpressing cells (ie, stem cell progeny that overexpress Notch are more likely to be maintained as stem cells). To assess self-renewal within the adult stem cell population, we dissociated and passaged the IC-Notch1 and littermate control-derived neurospheres [29,30]. Greater than 95% of the neurospheres in the cultures were derived from retrovirally infected cells (GFP+). We observed a 2.0-fold increase in the numbers of secondary neurospheres derived from the IC-Notch1 versus control groups. Conversely, when neurospheres from the attenuated Notch signaling group (Cre-infected Notch1 cKO) were passaged, we observed a 44.6% decrease in secondary neurosphere formation, as compared to GFP-RV control-infected neurospheres (Fig. 2C). Hence, Notch overexpression increases self-renewal. However, increased self-renewal alone cannot explain the increase in primary neurosphere numbers observed in the IC-Notch1 cultures since a single stem cell only generates 1 neurosphere, regardless of the initial mode of division [22,31]. Accordingly, these data are consistent with our in vivo findings that a subpopulation of progenitor cells in the SE acquire the stem cell properties of self-renewal and multipotentiality after Notch overexpression.
Notch1 expression confers stem cell characteristics upon progenitor cells in vitro
Definitively demonstrating that Notch signaling confers stem cell properties upon progenitor cells is difficult due to the lack of a specific marker that unambiguously and exclusively labels stem cells. Since we cannot achieve prospective isolation of stem cells upon plating, we took advantage of the SP analysis. This assay was originally reported to isolate a highly enriched population of hematopoietic stem cells from adult bone marrow [32], and we have demonstrated it to be a simple and effective means by which to enrich for neural stem cells from populations of neurosphere-derived cells [16]. Consistent with our previous report, we found that the SP fraction from adult-derived neurospheres comprises less than 1.0% of the live cells sorted but contains 99% of the neurosphere forming stem cells (Fig. 3A). Hence, while the SP fraction represents a mixed population of stem and progenitor cells, the non-SP fraction is a virtually pure population of progenitors.

Active Notch1 increases the SP fraction and converts non-SP cells into neurosphere-forming cells.
The in vitro studies and the in vivo lineage tracking both demonstrate that Notch overexpression leads to changes in progenitor cells, conferring upon them stem cell characteristics such as an ability to form clonal, multipotent, self-renewing neurospheres and a slower cell cycle time. A prediction from these findings is that the relative size of the SP fraction would be increased from populations of neural precursor cells with activated Notch signaling. Neurospheres derived from IC-Notch1 transgenic mice were dissociated and transduced with Cre-GFP-RV to overexpress active Notch in the cells. After 7 days, the resulting neurospheres were dissociated and SP sorted. As predicted, we observed a 6-fold increase in the proportion of SP cells from neurospheres derived from transgenic mice, compared with neurospheres derived from littermate control mice that also expressed GFP following transfection (Fig. 3B–D). Cells from both groups had similar infection efficiency (ranging from 12% to 15% GFP+ cells) and sorting with the GFP gate did not change the SP profile, confirming that the increased size of the SP fraction was due to active Notch signaling (Fig. 3D). The addition of verapamil during SP sorting to selectively block the ABC transporter eliminated the SP fractions in both cases (data not shown) [16,32]. Our second prediction is that if Notch confers stem cell properties upon progenitor cells, then the non-SP fraction from GFP+ IC-Notch1 cells would give rise to neurospheres. We observed virtually no neurospheres from the control group (0.3 neurospheres/5,000 cells). Consistent with our prediction, we observed a significant 6.8-fold increase in the number of GFP+ neurospheres from the non-SP fraction of IC-Notch1 mice (Fig. 3E, E'). We confirmed that the GFP+ neurospheres displayed the cardinal stem cell properties of multipotentiality and self-renewal (Fig. 3F). Overall, these in vitro data demonstrated that constitutively active Notch signaling preserves the stem cell properties in neural stem cells and confers stem cell properties upon progenitor cells in vitro. We conclude that Notch signaling is an important regulator of neural stem cell identity.
Discussion
The role of Notch has been studied extensively in the developing nervous system; however, the embryonic lethality of Notch pathway mutants has hindered studies aimed at examining its role in the adult brain. The creation of cre/lox-mediated conditional gain- and loss-of-function mice has enabled investigation of Notch signaling in the adult neural stem cell lineage. By studying this well characterized lineage, we have determined that active Notch signaling is important for preserving or enhancing the stem cell property of self-renewal and in addition, Notch signaling confers stem cell characteristics upon the progeny of neural stem cells. In vitro, Notch overexpression increased the number of cells that gave rise to multipotent, self-renewing neurospheres without affecting cell survival. Notch1 overexpression in progenitor cells (non-SP cells) was sufficient to convert the progenitors into multipotent and self-renewing neurosphere forming cells. In vivo studies showed that the loss of Notch signaling led to cells leaving the stem cell niche (the SE) and migrating onto the RMS sooner than is observed in control brains. Conversely, overexpressing Notch1 leads to a higher proportion of cells being retained in the SE for longer times (up to 12 days in this study). Moreover, the progenitor cells with enhanced Notch signaling in the adult brain formed multipotent and self-renewing neurospheres when isolated in vitro, something that was never observed in control mice. Together, these data indicate that not only does Notch signaling preserve stem cell characteristics, but also it can confer stem cell characteristics upon a subset of progenitor cells.
When Notch was overexpressed in the progenitor cells, we observed a decrease in proliferation at the population level, but it was not clear if this decrease occurred uniformly in all cells, or if only a subpopulation decreased the rate of proliferation and/or left the cell cycle. What is clear is that only a subpopulation of progenitor cells took on the stem cell characteristics of SE retention and neurosphere formation when Notch1 was overexpressed. The reason that only a subset of the cells responded to Notch in this manner is unclear. Based on work in the field of induced pluripotent stem cells where it has been shown that as stem cells become progressively more specified (tissue specific), more manipulations are required to revert them back to pluripotency [33], we hypothesize that only the progenitor cells that are the direct progeny of the neural stem cell (ie, the transit amplifying cell) can be reverted to a stem cell like state by the single manipulation of Notch overexpression.
Throughout this work we found modest, yet significant, effects when Notch1 is knocked-out both in vitro and in vivo. Notably, in our studies we knocked Notch1, the principle Notch signaling receptor in the neural stem and progenitor cell populations, however other Notch receptors exist, including Notch2, 3 and 4 [2]. Hence, focusing of a single receptor may lead to an underestimation of the importance of Notch signaling in neural precursor cells. Indeed, a critical role for Notch signaling in adult neurogenesis was demonstrated using nestin-CreERT2-mediated deletion of RBPj, a downstream mediator of all Notch receptors, demonstrating that the absence of Notch signaling leads to a loss of the stem cell population and an eventual cessation of SE derived neurogenesis [34].
Cells overexpressing or lacking Notch1 still migrated properly and a similar number reached the OB. Also, contrary to the reported effects of Notch signaling on neurite outgrowth in vitro and in vivo [35 –37], no differences were observed in neuronal morphology between transgenic and wild type animals (not shown). In the case of the Notch1 knockouts, this may simply be a result of redundant Notch family member proteins as discussed above. In the Notch1 overexpression mice, this lack of difference may indicate that Notch does not play a role in neuron formation in the adult brain, or that Notch signaling is blocked or outcompeted by other factors in the OB. Indeed, a previous work using Notch1 cKO in the subgranular zone of the hippocampus observed a related phenomenon that a change in the environment was able to overcome the effect of Notch knockout in the more committed doublecortin-positive neural progenitor cells, but not the upstream Nestin-positive progenitor cells [37].
There is considerable interest in stimulating endogenous neural stem and progenitor cells to promote repair after CNS injury or to reverse CNS dysfunction. The small number of stem cells present in the brain may be a limitation in recruiting neural stem cells for repair; hence, strategies to expand the stem cell pool for mobilization may prove to be important (reviewed in Ref. [39]). Accordingly, an understanding of the pathways that regulate the transition from stem cell to more restricted progenitor cell is an important consideration.
Overall, this study demonstrates that in the adult brain, Notch signaling is important for both preserving stem cell identity and the ability to confer stem cell properties upon a subpopulation of neural progenitor cells. This understanding lends insight into the mechanisms that regulate stem cell identity.
Footnotes
Acknowledgments
We thank Wenjun Xu, Mike Cooke, and Nikolaos Mitousis for their contributions. This work was funded by operating grants to CMM from the Heart and Stroke Foundation and the Stem Cell Network. D.P. is the recipient of a CIHR fellowship.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.