
Abstract
Growth-arrest-specific 7 (Gas7) is preferentially expressed in the nervous system and plays an important role during neuritogenesis in vertebrates. We recently demonstrated that gas7 is highly expressed in zebrafish neurons, where it regulates neural development. The possibility that gas7 may also regulate the development of other tissues remains to be examined. In this study, we investigate the role of Gas7 in the development of craniofacial tissues. Knockdown of gas7 using morpholino oligomers produced abnormal phenotypes in neural crest (NC) cells and their derivatives. NC-derived cartilage maturation was altered in Gas7 morphants as revealed by aberrant sox9b and dlx2 expression, a phenotype that could be rescued by coinjection of gas7 mRNA. While rhombomere morphology remained normal in Gas7 morphants, we observed reduced expression of the prechondrogenic genes sox9b and dlx2 in cells populating the posterior pharyngeal arches, but the fundamental structure of pharyngeal arches was preserved. In addition, NC cell sublineages that migrate to form neurons, glial cells, and melanocytes were altered in Gas7 morphants as revealed by aberrant expression of neurod, foxd3, and mitfa, respectively. Development of NC progenitors was also examined in Gas7 morphants at 12 hpf, and we observed that the reduction of cell precursors in Gas7 morphants was due to increased apoptosis level. These results indicate that the formation of NC progenitors and derivatives depends on Gas7 expression. Our observations also suggest that Gas7 regulates the formation of NC derivatives constituting the internal tissues of pharyngeal arches, without affecting the fundamental structure of mesodermal-derived pharyngeal arches.
Introduction
G
The neural crest (NC) is a transient tissue of the vertebrate embryo. The NC contains pluripotent cells that are derived from the border of the neural and epidermal ectoderm, and which migrate through specific migration pathways in the embryo. Following induction, NC cells delaminate, migrate, and differentiate into a wide variety of cell types, including neurons, glial cells, and melanocytes, as well as cells involved in the formation of bones, cartilage, teeth, smooth muscles, and connective tissues (reviewed in Le Douarin and Kalcheim and Graham et al. [14,15]). During migration, NC cells divide and begin to express markers according to their respective lineage. They are then guided to their destinations where they differentiate into mature cellular structures. Although the wide variety of vertebrate facial morphologies indicate that a complex interplay occurs between intrinsic genetic NC cell programs and extrinsic environmental signals during morphogenesis, the initial patterns of NC cells and migration are conserved between species [16].
Zebrafish exhibit fast development, and mutants are widely available, making this species a powerful model for the study of human skeletal disorders of unknown etiology. Some of the molecular mechanisms involved in establishing and maintaining the migration of segmentally restricted NC streams into the PAs have been established (reviewed in Minoux and Rijli [16]). On the other hand, we know little about the mechanisms by which craniofacial NC streaming is maintained in those species where focal cell death in rhombomeres 3 and 5 is not observed, such as in zebrafish [17] and Xenopus embryos [18]. Zebrafish pharyngeal arches are derived from all three germ layers. The NC is covered by the ectoderm on the outside and the endoderm on the inside. Interactions between the endoderm and NC are also known to be important for cartilage formation. Between arches, the endoderm meets the ectoderm, forming the pharyngeal pouches, which later develop into gill clefts and the epithelial lining of the pharynx, thyroid, parathyroid, and thymus.
Developmental aberrations of the pharyngeal apparatus lead to birth defects such as Treacher Collins and DiGeorge syndromes in humans [19]. Studies in mice, chickens, frogs, and zebrafish showed that multiple signaling pathways involving BMPs, FGFs, Notch, and Wnt proteins, as well as retinoic acid, participate in the formation of NC cells [20 –26]. These signaling pathways induce the expression of NC-specific genes, including ap-2, foxd3, pax3, slug, and sox10 [27 –33]. Many transcription factors mediate the differentiation of NC cells. In zebrafish, the two Sox9 orthologs (Sox9a and Sox9b) serve as markers for chondrogenic NC cells [34]. Expression of sox9a/b correlates with that of col2a1 in chondrogenic elements [35]. The zebrafish sox9 orthologs exhibit partially overlapping expression patterns in the craniofacial cartilage, otic placodes, and pectoral appendages [36]. Both dlx1a and dlx2a play a role in patterning the proximal aspects of the mandibular and ceratohyal arches and during maturation of the arch cartilage elements [37].
Recently, zebrafish Gas7 was found to be essential, but not specific, for the development of sensory neurons in zebrafish [8]. The knockdown of Gas7 expression using morpholino antisense oligonucleotides reduced the number of neurons in the trigeminal and statoacoustic ganglions and produced deformed phenotypes, such as flattening of top of the head, altered somitogenesis, cardiac edema, and defects in pigment distribution, suggesting that Gas7 possesses multiple functions [8]. In this study, we used the zebrafish model to examine the role of Gas7 on craniofacial formation, specifically NC cells and branchial cartilages, during early development. We found that the NC formation is disturbed in Gas7 morphants, leading to defective development of the primordia of pharyngeal arches and nonectomesenchymal sublineages in zebrafish. The impairment observed in Gas7 morphants is associated with an increased apoptosis of NC progenitors.
Materials and Methods
Animals and ethics statement
Adult zebrafish (Danio rerio) and embryos were maintained in a laboratory breeding colony on a 14-h light–10-h dark cycle using the methods described by Westerfield [38]. Wild-type zebrafish (strain Tuebingen) were purchased from the Zebrafish International Resource Center (ZIRC, Eugene, OR). Wild-type or transgenic line Tg (fli1a:EGFP) embryos were maintained at 28.5°C and staged according to the number of somites and hours (hpf) or days postfertilization (dpf) [39]. All experiments were performed in strict accordance with standard guidelines for zebrafish studies and were approved by the Institutional Animal Care and Use Committee of Chang Gung University (IACUC approval No. CGU04-57 and CGU08-86).
Morpholino and RNA injection
Morpholino antisense oligonucleotides (MO) were synthesized by Gene Tools (Corvallis, OR). Two MOs against gas7 were used, MO1 (5′-CTGTCTTTCTTTGACGATCTCACTC-3′ corresponding to nucleotides–36 to–61 located 5′ of the translation start site) and MO2 (5′-TGCTTGACTCTGACTGCTGCTTTGT-3′ corresponding to nucleotides–75 to–99) within the 5′-untranslated region. MO2 provided a better Gas7 knockdown efficiency than MO1 [8], and was therefore used in this study (gas7 MO). A standard MO control (5′-CCTCTTACCTCAGTTACAATTTATA-3′) was used in control injections. Capped RNA encoding the full coding sequence of Gas7 was prepared as before [8]. All injections were performed at the one- to two-cell stage, and mRNA (0.42 ng/embryo) or morpholinos (7.45 ng/embryo) were introduced into blastomeres.
Immunoblotting
Western blots on total protein extracts of embryonic zebrafish (48 hpf) were carried out essentially as before by Hung et al. [8]. Blots were incubated with antibodies directed against Gas7 [8] or glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (Santa Cruz Biotechnology, Santa Cruz, CA). Signal on the blot membrane was revealed using enhanced chemiluminescence according to the specifications of the supplier (Pierce, Rockford, IL).
Whole-mount cartilage staining
Cartilage was detected with Alcian blue staining using a protocol modified from Schilling et al. [40]. Zebrafish embryos (4 dpf) were fixed with 4% paraformaldehyde in PBS at 4°C overnight. After brief washing, embryos were bleached in 3% H2O2 for 1 h, followed by incubation in 0.1% solution of Alcian blue (dissolved in 80% ethanol/20% glacial acetic acid). The embryos were cleared with acidic ethanol, dehydrated, and stored in 90% glycerol.
Whole-mount in situ hybridization
Digoxigenin-UTP labeled riboprobes to detect gas7, sox9b, sox10, dlx2a, neurod, foxd3, and mitfa transcripts were synthesized according to the manufacturer's instructions (Roche, Basel, Switzerland), and an in situ hybridization was performed as described previously [8]. Embryos were treated with 1-phenyl-2-thiourea (PTU) to inhibit pigment formation and to facilitate imaging and staining for the whole-mount in situ hybridization. The color reaction was carried out using the NBT/BCIP substrate (Roche).
Whole-mount immunostaining
Fixed embryos were blocked in 5% goat serum, and antibodies against the proliferating cell marker phospho-histone 3 (pH3) (Millipore, Billerica, MA) were applied. The Alexa Fluor 488 fluorochrome-conjugated goat anti-mouse IgG (Invitrogen, Carlsbad, CA) was used as the secondary antibody. Embryos were mounted with the Vectashield mounting medium (Vector Laboratories, Burlingame, CA).
Cell death assay
Dehydrated embryos were rehydrated with reverse methanol gradient and then washed with PBST. After several washes with PBS, the embryos were subsequently incubated at 37°C for 1 h with a terminal transferase-mediated dUTP nick end-labeling (TUNEL) apoptosis detection kit (Roche). Embryos were then extensively washed with PBST and fixed with 4% paraformaldehyde in PBS at room temperature for 30 min.
Statistical analysis
Statistical analysis was performed with a two-tailed paired Student's t-test for single comparisons. Means are expressed ± SEM. All experiments were performed in triplicate.
Results
Knockdown of gas7 produces craniofacial defects in zebrafish
The efficiency and effectiveness of two morpholinos were described in our previous study [8], and both morpholinos produced identical phenotypes. Therefore, only the results on morpholino MO2 are presented here. While gas7 was mainly expressed in the zebrafish brain as shown by the in situ hybridization in 48 and 72 hpf embryos (Fig. 1A), we also detected the gas7 expression in the mesodermal tissues in the ventral head region (Fig. 1A, indicated by arrowheads), suggesting that Gas7 may mediate the development of cranial tissues derived from the NC or mesoderm. The effectiveness of gas7 MO was further confirmed by the observation that the expression of Gas7 protein was decreased in Gas7 morphants as revealed by immunoblotting (Fig. 1B, gas7 MO). Several defects were noted in Gas7 morphants, including aberrant tail curvature, cardiac edema, smaller eyes, shortened mandibular structure, and flattening of the top of the head (Fig. 1C).
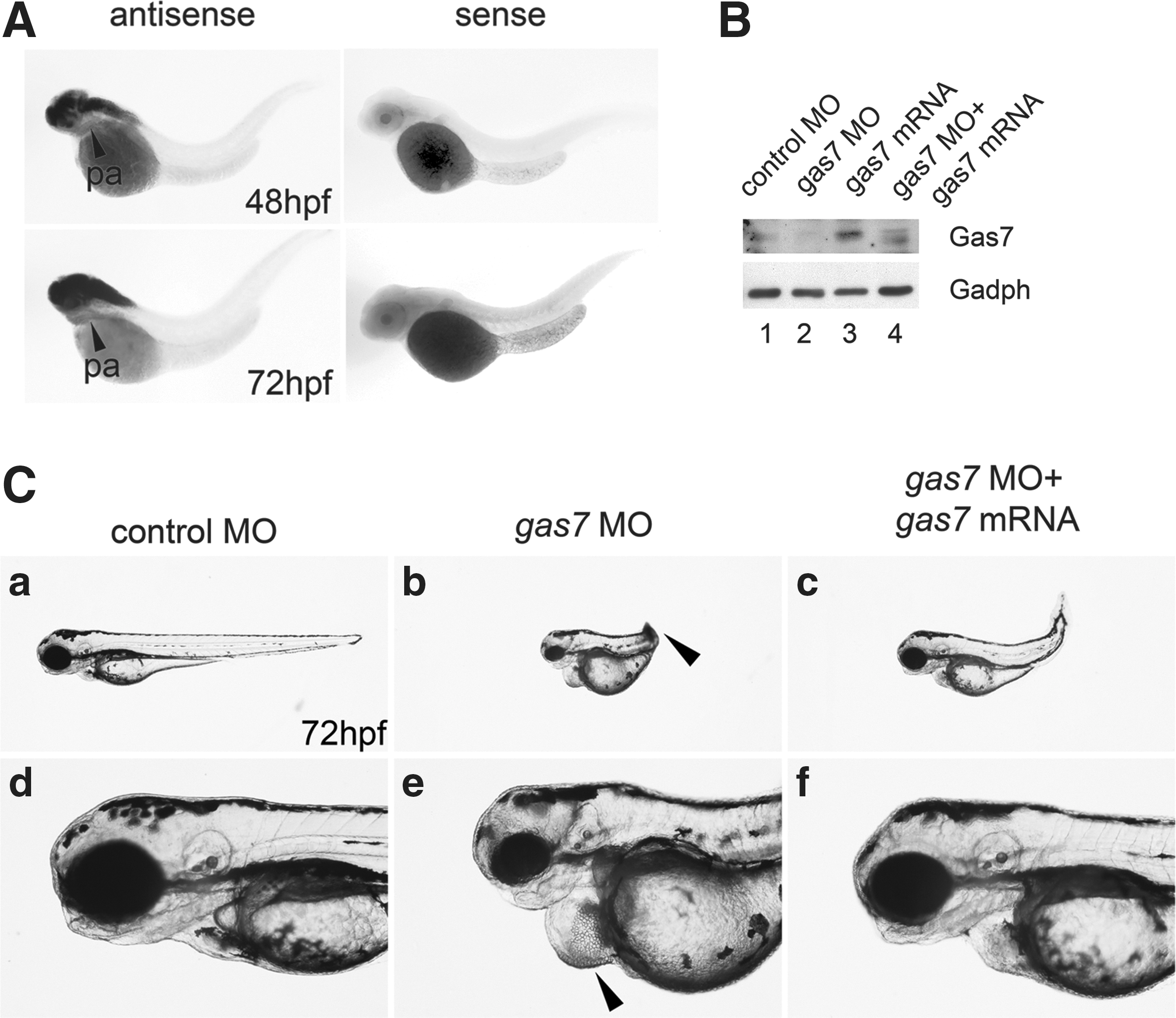
Phenotypes of gas7 knockdown in zebrafish.
To assess the effects of Gas7 deficiency on craniofacial skeleton defects, we examined the cartilage of Gas7 morphants using Alcian blue, which binds proteoglycan of the chondrogenic extracellular matrix (Fig. 2). Morphants were examined at 4 dpf to facilitate staining procedures and obtain an appropriate spatial architecture of cranial cartilages. A reduced Alcian blue staining was detected in pharyngeal cartilages of Gas7 morphants (Fig. 2). The bone development revealed by Alizarin red staining was also impaired in Gas7 morphants (Supplementary Fig. S1; Supplementary Data are available online at
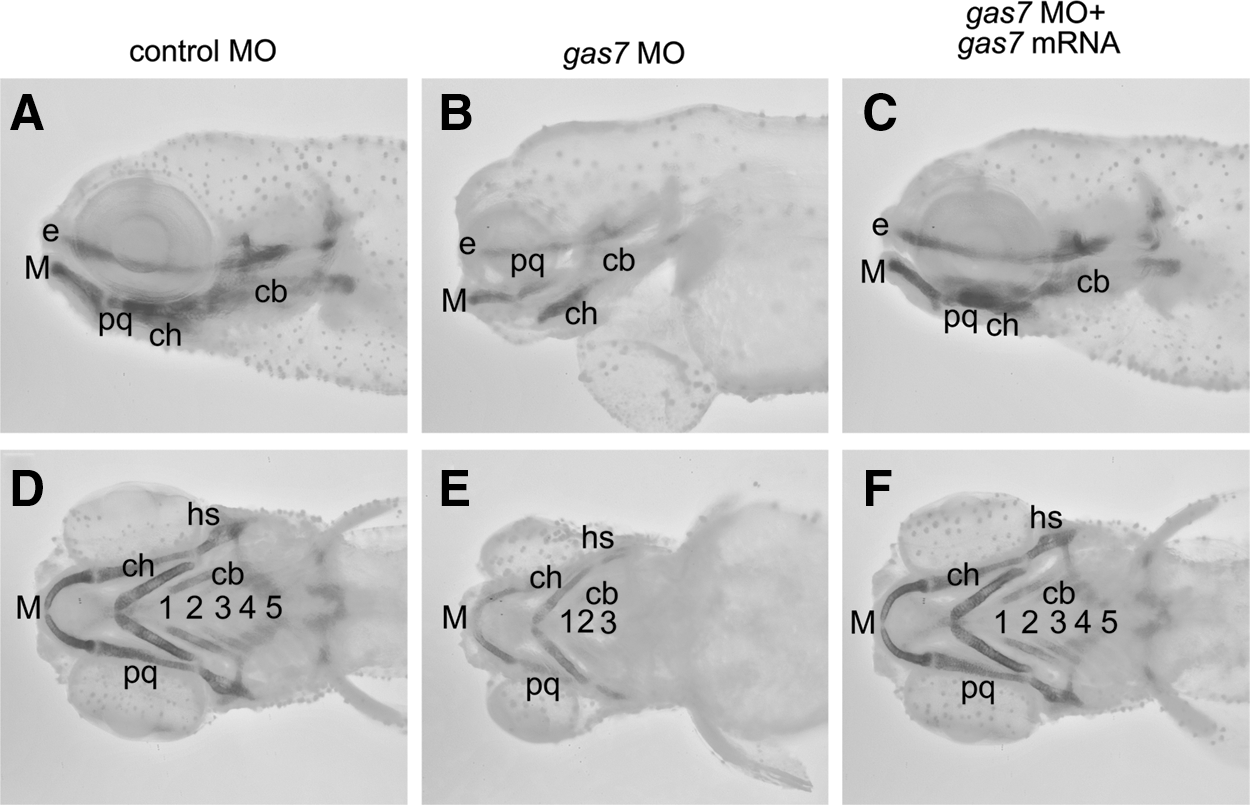
Effects of gas7 knockdown on craniofacial cartilage morphology. Cartilage patterning was assessed using Alcian blue staining. Lateral
In 48 hpf embryos, sox9b was expressed in the prechondrogenic condensation of the mandibular arch (Fig. 3A, indicated by number 1), hyoid arch (Fig. 3A, number 2), and the posterior branchial arch (Fig. 3A, numbers 3–6). We also examined Gas7 morphants at 48 hpf using sox9b and dlx2a as markers and found defects in the expression pattern of these genes in the lower jaw (Fig. 3A, B, D, E). These defects could be rescued by coinjection of gas7 MO with gas7 mRNA (Fig. 3C, F). Furthermore, we used fli1a:eGFP to examine the patterning of blood vessels, which is regulated by the structure of pharyngeal arches. We found that the patterning of fli1a-postive blood vessels was unaffected by gas7 morpholino (Fig. 3G–I), suggesting that Gas7 is not implicated in the morphogenesis of pharyngeal arches. The levels of sox9b and dlx2a expression were abnormal, while the patterning of pharyngeal arches was intact in Gas7 morphants, suggesting that Gas7 regulates the formation of NC derivatives constituting the internal tissues of pharyngeal arches, without affecting the fundamental structures of mesodermal-derived pharyngeal arches.
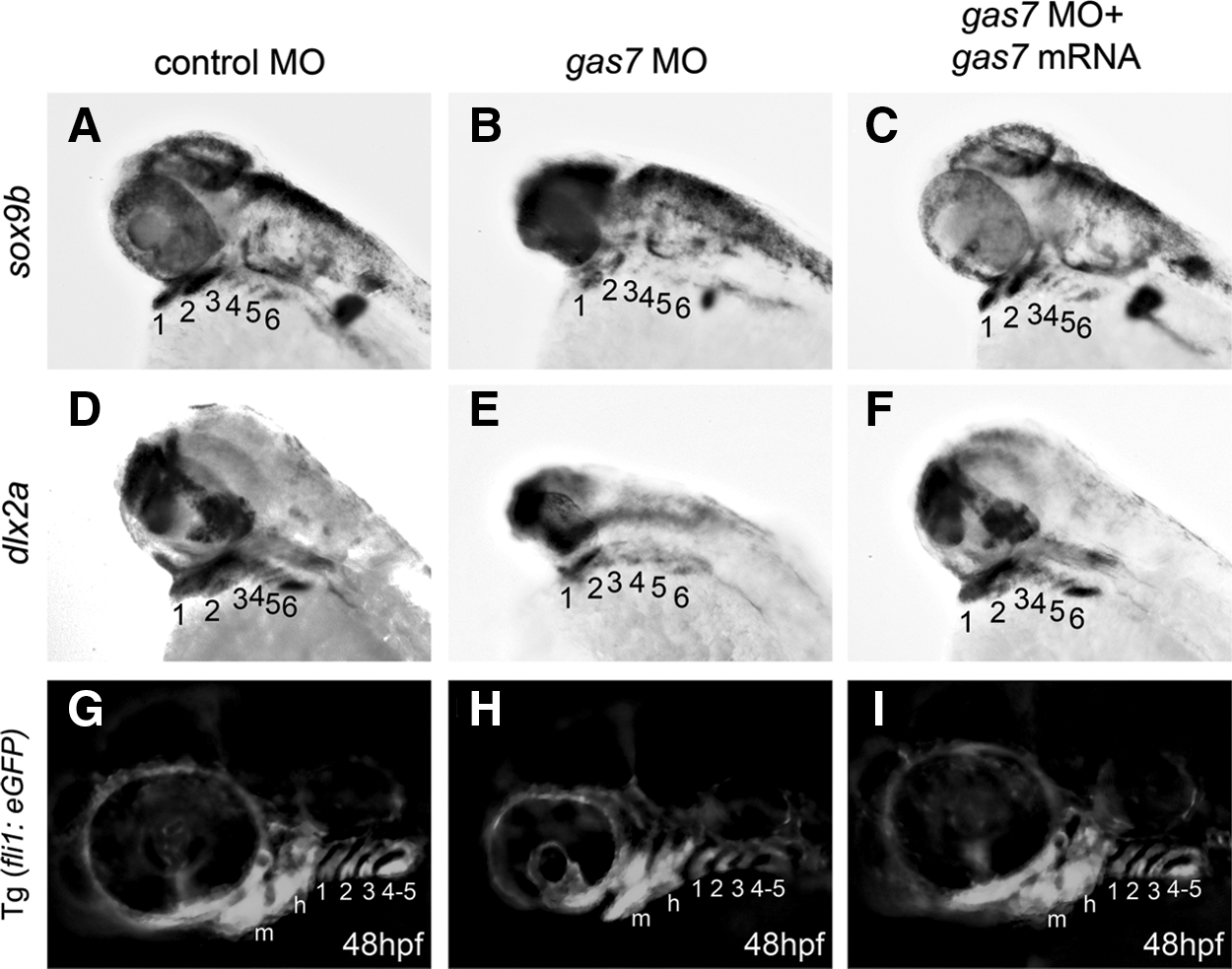
Effects of gas7 knockdown on craniofacial cartilage. Lateral views of 48 hpf larvae injected with control
Reduction of nonectomesenchymal and ectomesenchymal NC lineages in gas7 morphants
During early development, the zebrafish hindbrain is transiently partitioned into seven rhombomeres. The organization of rhombomeres is highly related to the numbers and migration patterns of NC cells [41,42]. Thus, the anteroposterior patterning of rhombomeres could conceivably alter the number of NC cells and their derivatives. To clarify whether the deformed craniofacial/chondrogenic phenotypes were caused by defective NC development or due to the abnormal development of rhombomeres, we analyzed the pattern of rhombomeres at 24 hpf, when the rhombomere structure can be clearly distinguished morphologically; we also used the molecular marker sox9b, which is located in the middle cells of each rhombomere [34]. Gas7 morphants displayed a reduced expression of sox9b in the otic primordium and along the rhombomere pattern (Fig. 4A, compare a and b for a representative morphology). However, sox9-positive cells were still located in the middle of each rhombomere, indicating that the pattern of rhombomeres was intact.
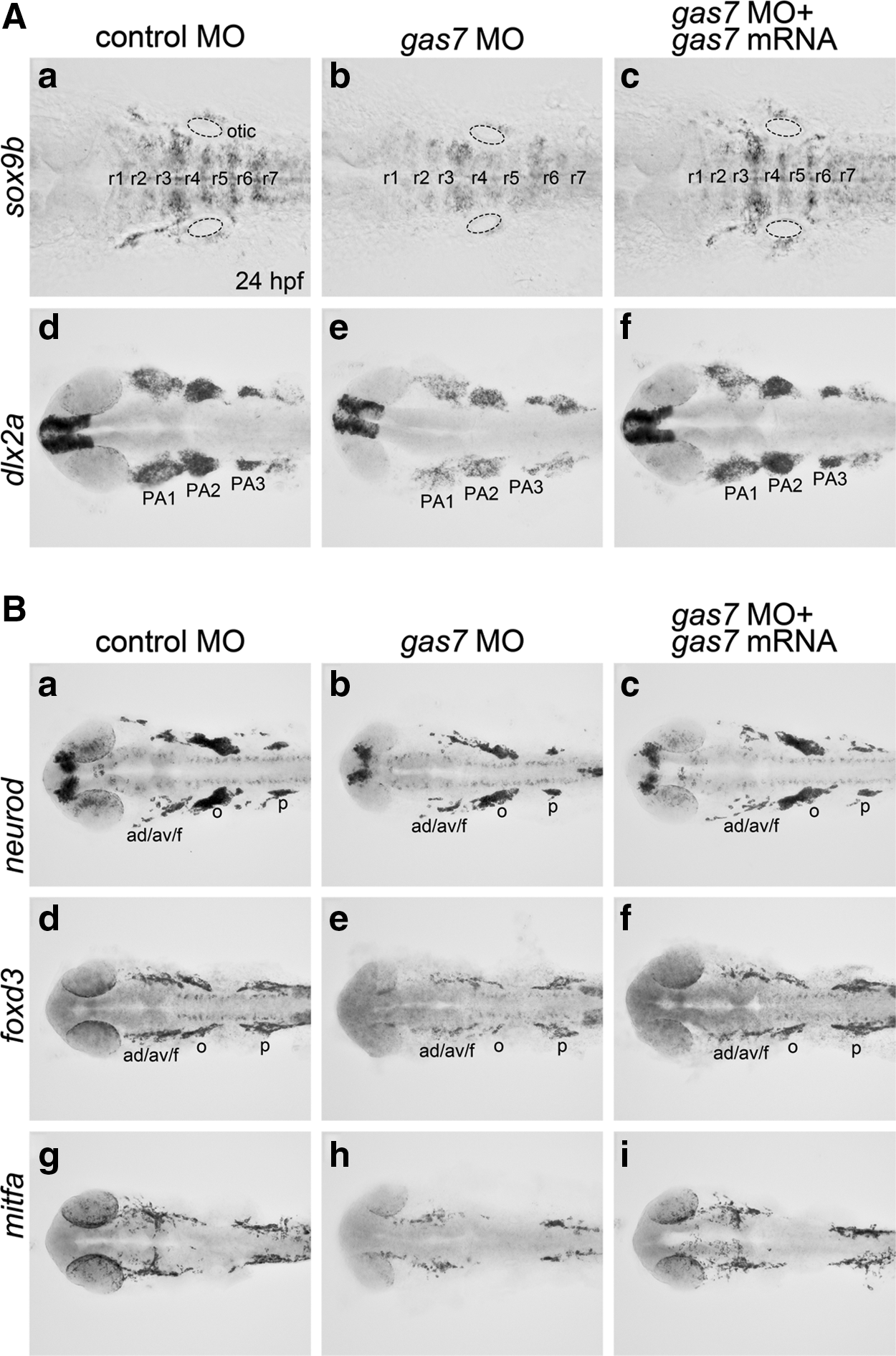
Effects of gas7 knockdown on NC cell lineages in 24 hpf larvae.
To confirm that the pattern of rhombomeres was affected by the gas7 knockdown, we labeled 24 hpf embryos with EphA4, a patterning marker of rhombomeres 3 and 5 [43]. We observed that the placement or compartmentalization of the rhombomeres in Gas7 morphants was normal (Supplementary Fig. S2). These results suggest that the gas7 knockdown caused defective sox9b-positive cells without affecting the pattern of rhombomeres. The specificity of this knockdown could be demonstrated by coinjection of gas7 MO with gas7 mRNA (Fig. 4A, compare a, b and c). We also found that gas7 MO reduced the sox9b expression in the cluster of cells adjacent to hindbrains. These cells are sox9b-positive NC cells that are migrating out from the neural tube, and which contribute to cranial cartilages. These results suggest that gas7 deficiency downregulates the sox9b expression in NC-derived mesenchymal cells. Reduced dlx2a levels (another marker for the cranial NC cells) were also noted at pharyngeal arches in the morphants (Fig. 4A, compare d and e). In addition, we observed a reduced number of total crestin-positive cells, a pan-crest cell marker, in Gas7 morphants at the 12- and 14-somite stages (Supplementary Fig. S3). These results suggest that NC progenitors may be affected by gas7 MO.
The migrating cranial NC cell population contains multipotent progenitor cells as well as two major lineages. These are the nonectomesenchymal lineage, which generates neurons, glial cells, and melanocytes, and the ectomesenchymal lineage, which gives rise to osteoblasts, chondrocytes, and muscle cells [17,44]. We examined the Gas7 morphants at 24 hpf, a time at which most NC derivatives have already differentiated, and we observed that gas7 MO injection downregulated the expression of the nonectomesenchymal lineage as revealed by markers of NC-derived neurons (neurod, Fig. 4B, a–c), glial cells (foxd3, Fig. 4B, d–f), and melanocytes (mitfa, Fig. 4B, g–i) at 24 hpf, suggesting that all NC derivatives were affected by the gas7 knockdown at 24 hpf.
Defects in NC cell formation in gas7 morphants
NC progenitors are induced during gastrulation and then begin to delaminate following migration and differentiation after expressing specific genes. One of the earliest genes expressed during the initial stage is sox10, a transcription factor characteristic of NC progenitor cells. NC progenitors express sox10 and sox9 between 10.5 and 12 hpf. We studied the expression of sox10 and sox9b to assess the NC development in Gas7 morphants. There was a dramatic reduction in the expression of sox10 and sox9b at 12 hpf in Gas7 morphants (Fig. 5A, representative images are shown in a–c and d–f, respectively). The specificity of this knockdown was demonstrated by coinjection of gas7 MO with gas7 mRNA, which significantly restored the expression levels of sox10 and sox9b (Fig. 5A, compare b and c; e and f), indicating that the Gas7 expression is essential for the development of NC progenitors.
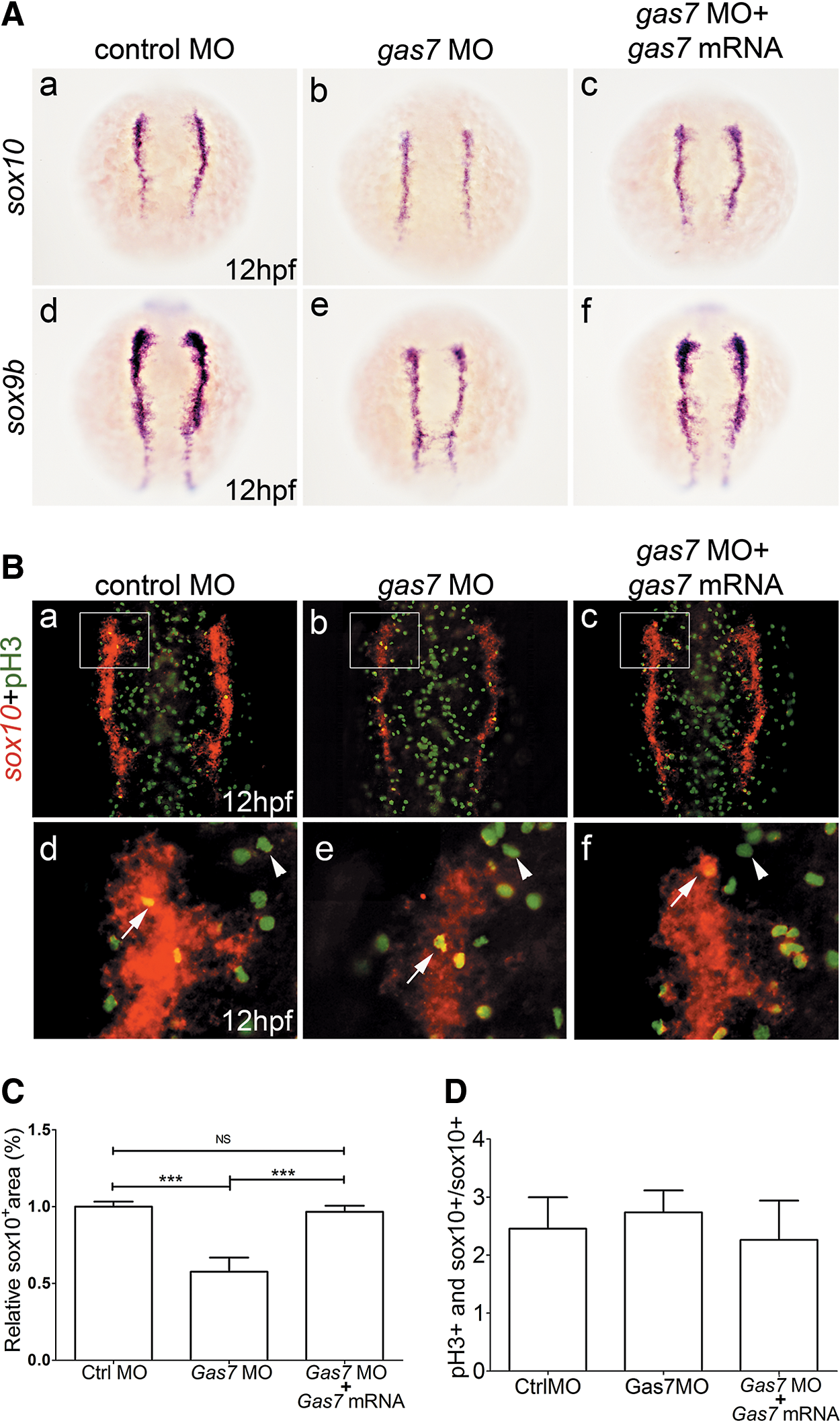
Effects of loss of gas7 on NC cell survival and proliferation.
An increased cell death or reduced proliferation of NC cells may be responsible for the disruption in NC development following the gas7 knockdown and consequently may cause craniofacial defects. To investigate these possibilities, we stained proliferating cells using immunolabeling with anti-phospho-histone 3 antibody (pH3); we also double-labeled NC cells using an in situ hybridization for sox10. A reduction of sox10-positive cells was noted in the gas7 MO group (Fig. 5B, C, P < 0.001). The specificity of this knockdown was demonstrated by coinjection of gas7 MO with gas7 mRNA. However, there was no difference in the level of pH3-sox10-double positive cells between the gas7 MO and control groups (Fig. 5D). Thus, the gas7 knockdown does not affect the proliferation of NC precursors.
Induction of apoptosis in NC cells by gas7 MO
To evaluate whether cranial NC cell reduction in Gas7 morphants is due to an increased apoptosis, we used the TUNEL assay to visualize apoptotic cells, and apoptotic NC cells were identified by colocalization with the sox10-positive signals at 12 hpf (Fig. 6A, indicated with arrowhead). Statistical analysis revealed a significant increase in apoptosis of NC cell precursors (sox10-positive cells) in Gas7 morphants (Fig. 6B, P < 0.01). The apoptosis induced by gas7 MO could be partly rescued by coinjection of gas7 mRNA (Fig. 6B, P < 0.05). To further confirm that the apoptotic cells were of NC origin, we also double stained the embryos at 10-somite stages with crestin using in situ hybridization. Apoptotic crestin-positive cells were increased in the morphants and were partly restored following the overexpression of gas7 mRNA (Supplementary Fig. S4). Since cell proliferation was not affected in Gas7 morphants, the decreased number of cells observed in this case could be caused by a defect in the regulation of apoptosis.
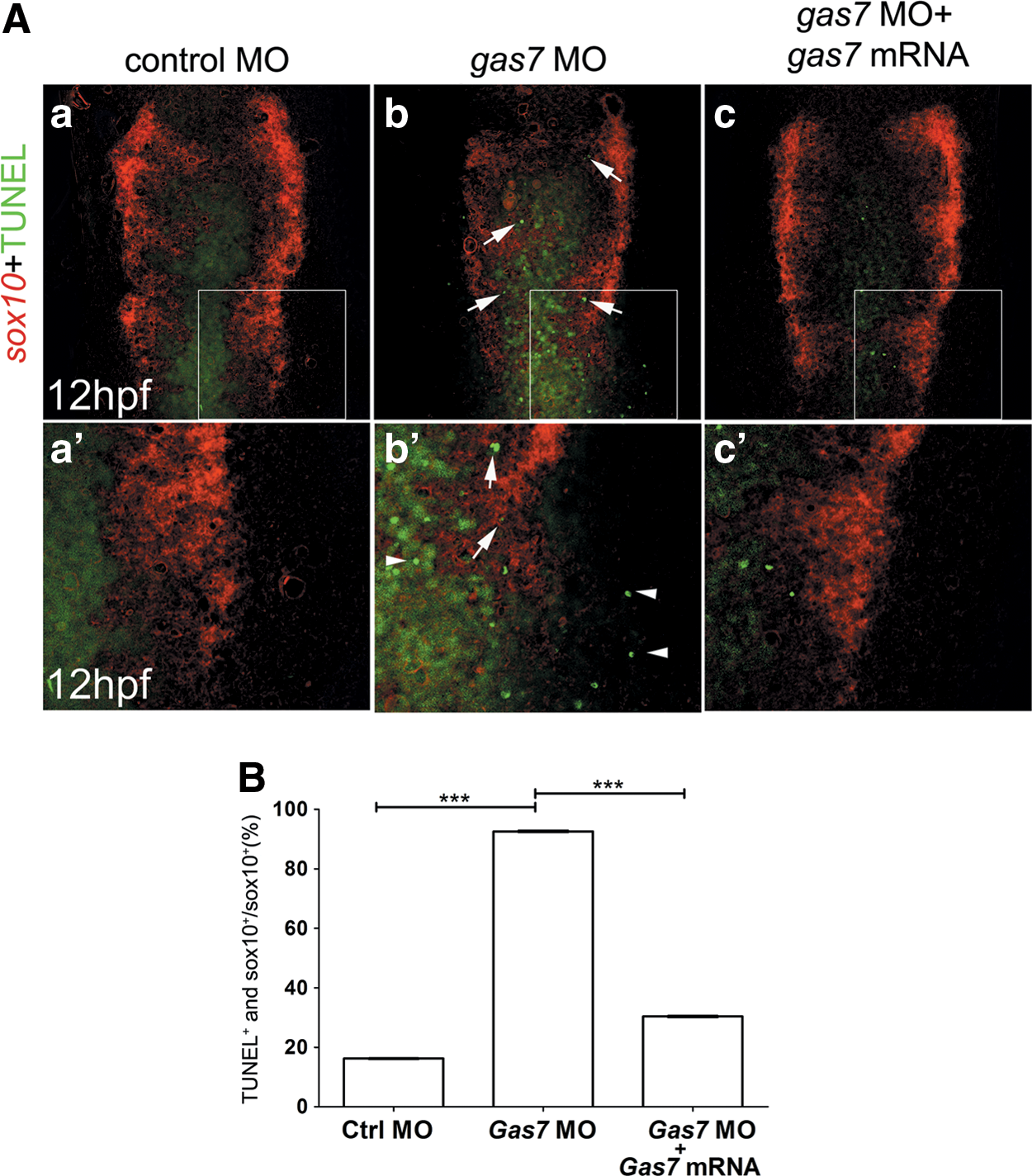
Effects of loss of gas7 on NC cell apoptosis. Sox10 in situ hybridization was carried out in combination with TUNEL assay on 12-hpf stage embryos.
Discussion
In this study, we show that the knockdown of gas7 in zebrafish produces phenotypes in craniofacial structure, including sharpened/protruding mandibular cartilage and shortened ceratohyal and ceratobranchial cartilage. These structures are developed from NC-derived chondrogenic cells. Gas7 may be required during a specific time period before NC differentiation and/or during migration. To examine whether NC progenitors were also affected before cranial chondrogenesis in Gas7 morphants, we stained embryos to monitor sox10 expression. While sox10 was expressed in most cranial NC progenitors (6-somite stage or12-hpf), sox10-positive cells were greatly reduced in Gas7 morphants (Fig. 5). The role of Gas7 in regulating apoptosis has also been demonstrated in cultured neuroblasts [45]. These results indicate that Gas7 is essential for cell survival and that a deficient expression of this gene results in cell apoptosis.
Defects in NC progenitors at early stages in Gas7 morphants may impair the development of NC derivatives such as branchial cartilages and thus the craniofacial structure. In addition, NC cells were able to migrate in the absence of Gas7, as seen in the pharyngeal arches and other derivatives (Figs. 3 and 4). The reduced lateral migration of NC cells observed in Gas7 morphants may not be only due to reduction in the number of NC precursors. However, a sudden reduction of sox10-positive NC progenitors was observed at the 6-somite stage in Gas7 morphants and this resulted in deformed cartilage (Fig. 5). In the absence of Gas7, the number of crestin-positive NC cells was also greatly reduced at the beginning of and during migration (Supplementary Fig. S3). At the later stage, the migratory NC cells failed to express sox9b at certain regions along the hindbrain (Fig. 4). Specifically, we observed that NC cells in the hindbrain commenced migration, but fewer reached their final destination at the posterior pharyngeal arches and differentiated into cartilage. Thus, Gas7 deficiency may also impair the migration and differentiation of NC. Direct evidence will require more experiments to track NC progenitors and their cell lineage maturation, including specification, migration, and differentiation.
In addition to defects in chondrogenesis, Gas7 morphants exhibited an altered development of neurons, glial cells, and melanocytes. In zebrafish, the specification of pigment cells from NC progenitors occurs before migration [46]. During migration, melanoblasts continue to proliferate and are susceptible to environmental cues for patterning. After the knockdown of Gas7, embryos displayed a reduced number and disorganized distribution of melanocytes (Fig. 4B and Supplementary Fig. S5). In addition, the level of NC-derived neurons of the dorsal root ganglion was reduced by the gas7 knockdown [8]. In general, Gas7 morphants showed a prominent defective development in both nonectomesenchymal and ectomesenchymal NC lineages. This observation can be simply explained by the loss of NC progenitors. Therefore, NC-derived cells are highly affected in Gas7 morphants, suggesting that Gas7 is essential for NC development.
NC cells arise from the border of the neural ectoderm and migrate to multiple targets in embryos [14]. Throughout their migration and differentiation, NC cells undergo cell division [47]. In zebrafish, peripheral neurons are formed from early migrating and lateral premigratory NC cells, while cranial cartilage and pigment cells are derived from late-migrating medial crest cells [18]. Precise regulation of NC cell proliferation is crucial for the developmental processes of NC cells. The present and our previous study revealed that Gas7 may regulate sox9-dependent transactivation of chondrogenic genes and chondrogenic differentiation in primary culture of human mesenchymal stem cells [48]. We also found that Gas7 is required for neuronal development in zebrafish [8] and in mammalian cell cultures [4,6,7]. Taken together, these results suggest that Gas7 plays multiple important roles in vertebrates, such as in chondrogenesis and neurogenesis.
The mechanisms underlying the regulation of embryonic NC development by Gas7 are important for our understanding of NC-related pathologies and are still being investigated. In addition, adult NC-derived stem cells persist in various locations of the postnatal organism. For example, epidermal-NC stem cells (EPI-NCSC) are multipotent stem cells, which are located in the bulge of hair follicles. EPI-NCSC can undergo self-renewal and generate all major NC derivatives, including neurons, nerve-supporting cells, smooth muscle cells, bone/cartilage cells, and melanocytes [49]. New medical applications may eventually be developed based on knowledge of the roles of stem cells present throughout the body. Our findings that Gas7 regulates NC-cell-associated embryonic development suggest that this gene may play an important role in the regeneration of tissues and organs in adults and may lead to novel clinical applications in this field.
Footnotes
Acknowledgments
We thank colleagues of the Zebrafish Core Laboratory for technical assistance. This work was supported, in part, by Chang Gung University (CMRPD34019) (to YCC) as well as by Chang Gung Memorial Hospital (BMRP071), and the Foundation for the Advancement of Outstanding Scholarship (to CCKC).
Author Disclosure Statement
The authors declare no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.