
Abstract
Due to late diagnosis, often recurrence, formation of metastases and resistance to commonly used chemotherapeutics human ovarian carcinoma represents a serious disease with high mortality. Adipose tissue-derived mesenchymal stromal cells (AT-MSC) can serve as vehicles for therapeutic genes and we engineered AT-MSC to express either Herpes simplex virus thymidine kinase (HSVtk-MSC), which phosphorylates ganciclovir (GCV) to its toxic metabolites or yeast fused cytosine deaminase::uracil phosphoribosyltransferase (CD::UPRT-MSC), which converts 5-fluorocytosine (5-FC) to highly toxic 5-fluorouracil (5-FU). Here, we reported different responses of cytotoxicity mediated by CD::UPRT-MSC/5-FC treatment on human ovarian carcinoma cell lines—SKOV-3 and A2780 used in adherent or three-dimensional (3D) cell culture and we proved high potential of 3D model to predict results in our in vivo experiments. Both tumor cell lines showed similarly high chemosensitivity to the used treatment in adherent culture, but 3D model revealed severe discrepancy—only 36% of SKOV-3 cells but even 90% of A2780 cells were eliminated. This result served as a prognostic marker—we were able to achieve significantly decreased tumor volumes of subcutaneous xenografts of A2780 cells in nude mice and we prolonged tumor-free survival in 33% of animals bearing highly metastatic ovarian carcinoma after CD::UPRT-MSC/5-FC treatment.
Introduction
H
Mesenchymal stromal cells (MSC) are defined as a population of nonhematopoietic undifferentiated cells with self-renewal potential, capability of differentiation into various mature somatic cell types [5], their tropism to the site of wounds or chronic inflammation, and regenerative potential [6]. They are characterized by their adherence to plastic in culture, specific cell surface marker expression and capability of trilineage differentiation [7,8] and they can be readily obtained from various tissues [9,10]. Due to their preferential homing into tumor mass [11], easy harvest, isolation, expansion, efficient transgene introduction with viral vectors, and refractoriness to host immune response, MSC can be considered as very suitable vehicles to aim tumor cells [12,13]. MSC are able to migrate to primary or metastatic tumors derived from breast, glioma, colon, ovarian, prostatic, lung carcinoma, or melanoma upon systemic administration [14]. Due to this specific feature we tried to exploit the adipose tissue-derived MSC (AT-MSC) as cellular delivery vehicles for a gene-directed enzyme/prodrug therapy (GDEPT).
The main idea of GDEPT is to generate differences between healthy and tumor cells on biochemical level, using various delivery methods (viral-based, molecular, or cellular vectors) to bring a gene product, which converts a nontoxic prodrug to highly toxic metabolites in the vicinity of targeted tumor cells. These toxic compounds are taken up by neighboring cells (not expressing the suicide gene) via gap junctions or free diffusion across the cell membrane by a process named bystander effect (enhanced by host immune system or by uptake of apoptotic bodies containing toxic metabolites) [15 –19].
We genetically modified the AT-MSC to express gene for viral (Herpes simplex) thymidine kinase (HSVtk) or fusion yeast cytosine deaminase::uracil phosphoribosyltransferase (CD::UPRT). HSVtk belongs to one of the most intensively studied suicide gene systems: HSVtk catalyzes phosphorylation of nontoxic ganciclovir (GCV) to its phosphorylated form–GCV monophosphate and cellular kinases convert it to di- and tri-phosphate. GCV triphosphate (GCV-3P) can be incorporated into the molecule of DNA, which results in the termination of its synthesis and cell commits suicide via pathway [17,18]. GCV-3P is a charged molecule and a gap junctional intercellular communication (GJIC) between therapeutic and tumor cells is necessary to achieve therapeutic effect [20]. Tight gap junctions are formed from connexin proteins [21] and Matuskova et al. [16] showed that the AT-MSC are capable of GJIC formation thus rendering them suitable for HSVtk/GCV treatment. Nowadays, there are four completed clinical trials investigating the gene therapy using HSVtk/GCV to treat brain neoplasms (NCT00001328; phase 1), hepatocellular carcinoma (NCT02202564; phase 2), melanoma (NCT00005057; phase 1), and hematological malignancies (NCT01086735; phase 1 and 2). There is still ongoing clinical trial (NCT00005025; phase 2) using the gene therapy in treating refractory or relapsed ovarian epithelial cancer, fallopian tube cancer, or peritoneal cancer, trying to determine development of systemic immunity against the tumor in patients and any possible toxic effects of this gene therapy.
Another GDEPT uses bacterial (Escherichia coli) or yeast (Saccharomyces cerevisiae) gene encoding cytosine deaminase capable of converting an antifungal agent 5-fluorocytosine (5-FC) to a commonly used chemotherapeutic 5-fluorouracil (5-FU). The synthesized compounds are incorporated into nascent strands of DNA and RNA and they terminate the synthesis of nucleic acids, which results in cell death [22,23]. The enzymes involved in the 5-FU metabolism such as thymidylate synthase (TS), thymidine phosphorylase (TP), orotate phosphoribosyltransferase (OPRT), and dihydropyrimidine dehydrogenase (DPD) have been under focus of 5-FU response predictive biomarkers in many studies [24 –26]. The construction of bifunctional yeast fusion gene coding for CD::UPRT significantly improved the conversion of 5-FC to 5-FU and its metabolites further improving the therapeutic effect in various preclinical animal models for colon, breast, medullary thyroid carcinoma, melanoma, glioma, and others. The suicide gene therapy decreases the systemic side effects of 5-FU therapy and increases the local antitumor effect [12,27 –29]. Several positive results supported the pilot study of oral 5-FC and neural stem cells expressing gene for bacterial CD for treatment of recurrent high grade gliomas (NCT01172964).
We decided to examine the efficacy of the bystander effect mediated by two different enzyme/prodrug systems delivered by genetically engineered AT-MSC on two human ovarian carcinoma cell lines SKOV-3 and A2780.
Materials and Methods
Chemicals
All chemicals were purchased from Sigma-Aldrich® (St. Louis, MO), if not stated otherwise.
Cell lines
Human ovarian carcinoma cell line SKOV-3 (ATCC HTB-77), SKOV-3/EGFP expressing enhanced green fluorescent protein, EGFP; all EGFP-positive cells were prepared according to Matuskova et al. [20] and human fibroblasts were cultured in high-glucose (4.5 mg/mL) Dulbecco's modified Eagle medium (DMEM; PAA Laboratories, GmbH, Pasching, Austria) supplemented with 5% fetal bovine serum (FBS; GIBCO® Invitrogen™, Carlsbad, CA); antibiotic/antimycotic mix: 10,000 IU/mL penicillin (BiotikaOArt. Lupca, Slovakia), 5 μg/mL streptomycin, 2.5 μg/mL amphotericin, and 2 mM glutamine (PAA Laboratories GmbH). Human ovarian adenocarcinoma cell line A2780 (ECACC 93112519) and A2780/EGFP were maintained in high-glucose DMEM supplemented with 10% FBS, antibiotic/antimycotic mix and glutamine. Human AT-MSC were isolated by plastic adherence technique as described previously [12]. The AT-MSC were cultured in low-glucose (1 mg/mL) DMEM (PAA Laboratories, GmbH) supplemented with 5% HyClone Advance Mesenchymal Stem Cell Growth Supplement (Thermo Scientific, Waltham, MA), 5% FBS and antibiotic/antimycotic mix (GIBCO Invitrogen Life Technologies, Carlsbad, CA). Cells were grown at 37°C in 5% CO2 humidified atmosphere.
Fluorimetric assay
Chemosensitivity to cisplatin and 5-FU was evaluated by fluorimetric assay as described previously [16]. Briefly, tumor cells were seeded on a black 96-well plate (Greiner BioOne, Monroe, NC) with a concentration gradient of cisplatin or 5-FU. The cells were washed thrice with phosphate-buffered saline (PBS) and the fluorescent signal was measured in PBS containing 0.2% Nonidet P40 at day 6 or 7 by PolarStar OPTIMA reader (BMG Labtech, Offenburg, Germany). Similarly, both SKOV-3/EGFP and A2780/EGFP were cocultured with AT-MSC, HSVtk-MSC, or CD::UPRT-MSC in the presence of concentration gradient of GCV or 5-FC; alternatively with different ratio of tumor cells to therapeutic AT-MSC in the presence of stable concentration of a prodrug and the cell's fluorescence was measured at day 3 or 7. All tests were performed in quadruplicates; results were expressed as percentage of relative fluorescence [average ± standard deviation (SD)] and fluorescence of untreated cells in standard culture medium without prodrugs was set to 100%.
Spheroid formation in 3D culture
Quadruplicates of 3.5 × 103 SKOV-3/EGFP or 4.5 × 103 A2780/EGFP cells were mixed with different ratio of CD::UPRT-MSC (labeled with DiI according to [30]) and seeded into 96-well ultra-low attachment plate (Corning 7007; Corning, Inc., Corning, NY) in 125 μL of standard culture media according to CellPlayer™ 96-well Kinetic 3D Spheroid Protocol (Essen BioScience, Welwyn Garden City, UK). Forty-eight hours later, 125 μL media with or without 5-FC was added to the wells and the growth of aggregates was monitored by IncuCyte ZOOM™ Kinetic Imaging System (Essen Bioscience) for 9 days. At the assay endpoint, the viability assay was performed on spheroids as described below.
Cell viability assay
The relative viability of the cells in the experimental setup of 3D aggregate's cultures was measured by the CellTiter-Glo™ Luminescent Cell Viability Assay (Promega Corporation, Madison, WT) and evaluated by the LumiStar GALAXY reader (BMG Labtech) at the experiment endpoint (day 9). Briefly, 96-well plate and all reagents were equilibrated to the room temperature, 50 μL of prepared CellTiter-Glo® Reagent was added to 50 μL of media containing aggregates in 3D culture plates, mixed, and placed on an orbital shaker in the absence of light to induce cell lysis. After stabilization of luminescent signal, 100 μL was transferred to a white 96-well plate (Greiner BioOne) and luminescence was recorded. Test was performed in quadruplicates; results were expressed as percentage of relative proliferation (average ± SD). Proliferation of untreated aggregates in standard culture media without a prodrug was set to 100%.
Triplicates of 4.5 × 103 A2780 cells were seeded on a white 96-well plate (Greiner BioOne) with different amount of AT-MSC or CD::UPRT-MSC in the presence of 50 μg/mL 5-FC. The relative viability of cells was evaluated by the CellTiter-Glo Luminescent Cell Viability Assay (Promega Corporation) at day 3. Results were expressed as percentage of relative proliferation (average ± SD) and proliferation of A2780 cells was set to 100%.
Evaluation of GJIC
GJIC of SKOV-3 and A2780 cells with AT-MSC was evaluated by using flow cytometry to detect fluorescent dye transfer between tumor cells and AT-MSC as described previously [30].
Quantitative reverse transcriptase–polymerase chain reaction analysis
Expression of OPRT, DPD, TS, and TP was determined by quantitative reverse transcriptase–polymerase chain reaction (qRT-PCR) from SKOV-3 and A2780 cells cultured in the absence/presence of 5-FU (IC50 concentration of 5-FU for SKOV-3 = 0.12 μg/mL; A2780 = 0.47 μg/mL) for 3 days.
Expression of Connexin 43 was also examined by qRT-PCR from SKOV-3, A2780, AT-MSC, and human fibroblasts.
Total RNA was isolated from 5 × 106 cells by NucleoSpin RNA II kit (Macherey-Nagel) and treated with RNase-free DNase (Macherey-Nagel). RNA was reverse transcribed with a RevertAid™ H minus First Strand cDNA Synthesis Kit (Thermo Scientific). Quantitative PCR was performed in 1× GoTaq® qPCR Master Mix (Promega Corporation), 0.1 μM primers (Table 1) and 100 ng of template cDNA on a CFX96™ Real-Time PCR Detection System (Bio-Rad, Hercules, CA) using the following protocol: activation step at 95°C for 2 min, 40 cycles of denaturation at 95°C for 15 s, 60 s annealing and polymerization at 60°C, and plate read for 5 s at 76°C followed by melt curve analysis. The size of PCR products was verified by 4% MetaPhor Agarose (Lonza, Rockland, ME) electrophoresis. Relative gene expression change was calculated according to the ΔΔCt method. GAPDH (glyceraldehyde 3-phosphate dehydrogenase) gene was used as a reference gene. Analysis was performed twice in triplicates and data are expressed as mean ± SD.
Cx43, Connexin 43; DPD. dihydropyrimidine dehydrogenase; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; OPRT, orotate phosphoribosyltransferase; TP, thymidine phosphorylase; TS, thymidylate synthase.
In vivo data
Six to eight-weeks-old athymic female nude mice (Balb/c nu/nu) were used in accordance with the institutional guidelines under the approved protocol. Project was performed in the approved animal facility (license No. SK PC 14011) as approved by the Institutional Ethic Committee and by the national competence authority (State Veterinary and Food Administration of the Slovak Republic, registration No. Ro 3108/14-221) in compliance with the Directive 2010/63/EU of the European Parliament and the European Council and the Regulation 377/2012 on the protection of animals used for scientific purposes.
Mice were injected with 2.5 × 106 SKOV-3/EGFP cells alone or co-injected with 2.5 × 105 CD::UPRT-MSC in 100 μL PBS into the flanks on both sides subcutaneously (s.c.). Animals (n = 6) were treated with 500 mg/kg/day i.p. (Ancotil, Valeant, Poland) of 5-FC starting on day 0 or 3 for 14 consecutive days. Mice were sacrificed on day 28 after the tumor cells administration, when the control-untreated animals had to be sacrificed. Tumor volume was calculated from caliper measurements according to formula: volume = (length × width2)/2. Results were evaluated as average of tumor volume ± standard error (SE).
Alternatively, mice (n = 3–4 per group) were inoculated with 2 × 106 A2780 cells alone or co-injected with 4 × 105 AT-MSC or CD::UPRT-MSC in 100 μL PBS s.c. into the flanks on both sides. The administration of prodrug 5-FC (i.p.) started on day 0 for 14 days and mice were sacrificed when the tumor volume exceeded 500 mm3. Results were evaluated as median tumor volume ± SE.
Athymic female nude mice (n = 4–6 per group) were injected with 5 × 106 SKOV-3/EGFP cells in 100 μL PBS i.p. (day 0) and 2 × 106 CD::UPRT-MSC were administered i.p. at the day 7, 14, and 21 after injection of tumor cells. The treatment with 5-FC (i.p.) started concomitantly with injection of therapeutic AT-MSC for 14 consecutive days. All animals had to be sacrificed due to moribund state at day 27 and examined for the presence of metastases in abdominal cavity.
Alternatively, mice (n = 6–12 per group) were injected with 2.5 × 106 A2780/EGFP cells in 100 μL PBS i.p. (day 0) and 1 × 106 CD::UPRT-MSC were administered i.p. at day 7, 14, and 21 after injection of tumor cells. The treatment with 5-FC (i.p.) started as described previously. Mice were monitored for disease-free survival; they were sacrificed at the moribund state and examined for the presence of metastases.
Statistical analysis
Normality of distribution was tested by the Shapiro–Wilk test. Differences between more than two groups were tested by one-way analysis of variance and by Bonferroni's test if equal variance was assumed or Tamhane's test if equal variances were not assumed (as a post hoc procedure when needed). P values with P < 0.05 were considered as statistically significant. The Kaplan–Meier survival times of groups of mice were compared with the control group using a log-rank test (IBM® SPSS® Statistics, version 23.0).
Results
Previously, we have reported the antitumor efficiency of HSVtk-MSC/GCV or CD::UPRT-MSC/5-FC treatment on various types of tumors cells—melanoma (A375), breast carcinoma (MDA-MB-231), cervical adenocarcinoma (HeLa), glioblastoma (8-MG-BA) [16], and medullary thyroid carcinoma (TT) [31] and we also performed experiments using the combination of both types of gene therapy on tumor cell lines [30]. This study focused on comparison of two different types of GDEPT mediated by therapeutic AT-MSC on human ovarian carcinoma cell lines with metastatic potential.
HSVtk-MSC/GCV treatment on human ovarian carcinoma cells
As the functional GJIC is an important prerequisite for TK/GCV effectiveness, we examined the expression of Connexin 43 in tumor cells and AT-MSC by qRT-PCR (Fig. 1A). Our data showed the ability of tumor cells to communicate via tight gap junctions after direct coculture with AT-MSC, which was confirmed by functional dye transfer assay (Fig. 1B, C). We observed a shift of both cell lines to Calcein AM+ DiI+ region (double positivity) (region 3 on graphs), so both tumor cells SKOV-3 and A2780 can exchange metabolites via gap junctions. This functional GJIC predicted that the treatment with HSVtk-MSC/GCV could be effective due to transfer of toxic metabolite GCV-3P from AT-MSC via connexin channels to tumor cells as it was previously shown by Kucerova et al. [13]. Therefore, we cocultured both tumor cell lines with HSVtk-MSC in the presence of a concentration gradient of GCV to directly evaluate bystander effect in elimination of tumor cells (Fig. 1D–F). SKOV-3 cells were more sensitive to the treatment compared to A2780 (IC50 was not reached), but higher proportion of HSVtk-MSC in co-culture did not eliminate more SKOV-3 cells (data not shown).
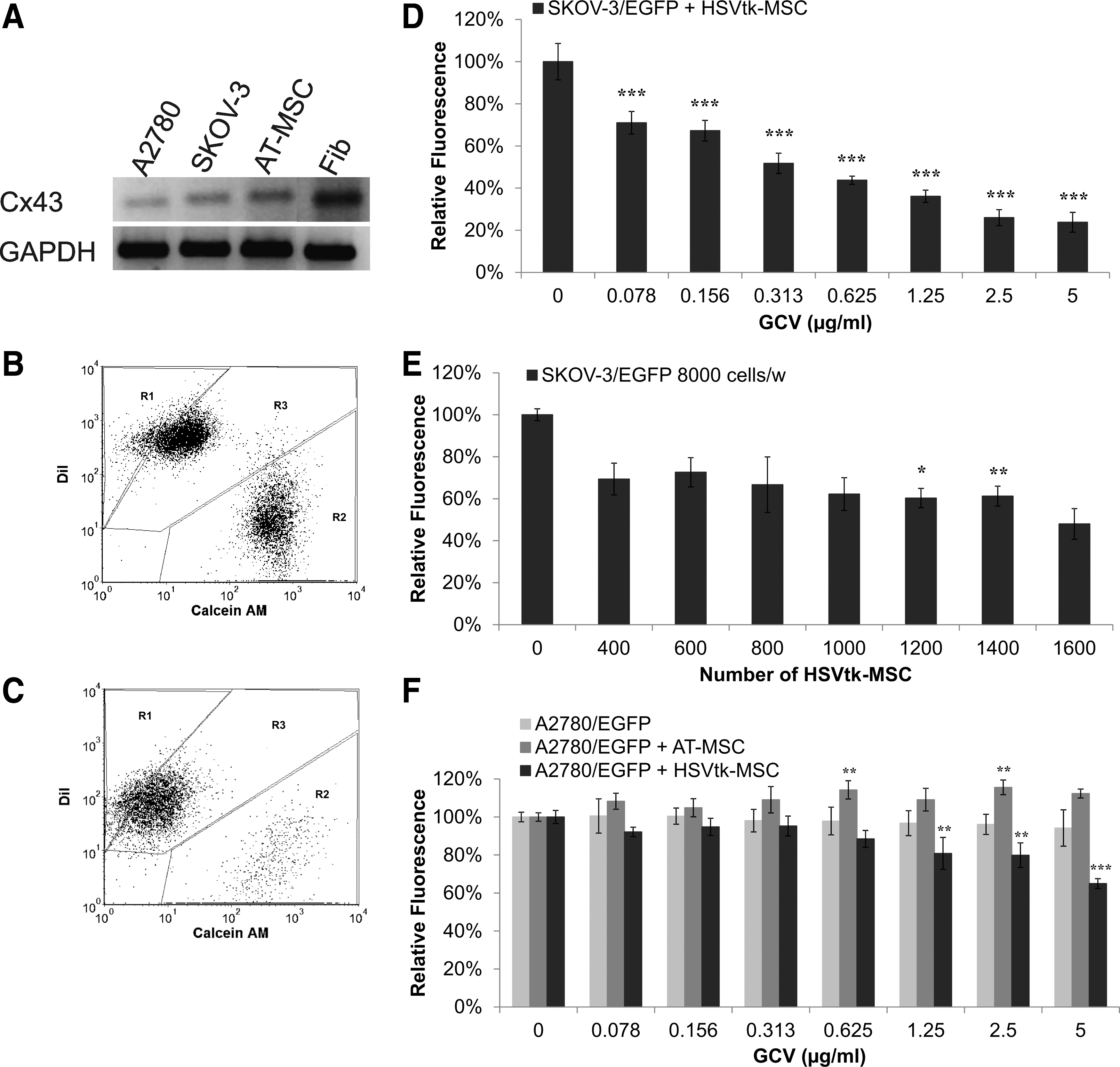
HSVtk-MSC/GCV did not eliminate ovarian tumor cells.
CD::UPRT-MSC/5-FC treatment on tumor cells in monolayer culture
As previously mentioned [32], we showed that SKOV-3 cells were chemoresistant (IC50 = 0.718 μg/mL) and A2780 cells were chemosensitive (IC50 = 0.069 μg/mL) to the commonly used chemotherapeutic platinum salt—cisplatin (Fig. 2A). To examine whether CD::UPRT-MSC/5-FC treatment is efficient to eliminate ovarian cancer cells we performed the experiment using the concentration gradient of 5-FU on both tumor cell lines. Contrastingly, both tumor cell lines showed similar chemosensitivity to 5-FU (IC50 for SKOV-3 = 0.149 μg/mL; IC50 for A2780 = 0.376 μg/mL) (Fig. 2B). The expression analysis of qRT-PCR showed different levels of enzymes involved in the metabolism of 5-FU (Fig. 2C). SKOV-3 cells after treatment of 5-FU slightly increased expression of TS and decreased expression of TP. The levels of DPD and OPRT did not change compared to untreated cells. Treated A2780 cells showed almost threefold higher expression of TS, and twofold higher expression of OPRT compared to untreated cells. Regardless of differences in intracellular levels of the 5-FU metabolizing enzymes, both ovarian cancer cell lines exhibited similar cytotoxic responses, which were comparable to other tumor cell lines tested [16].
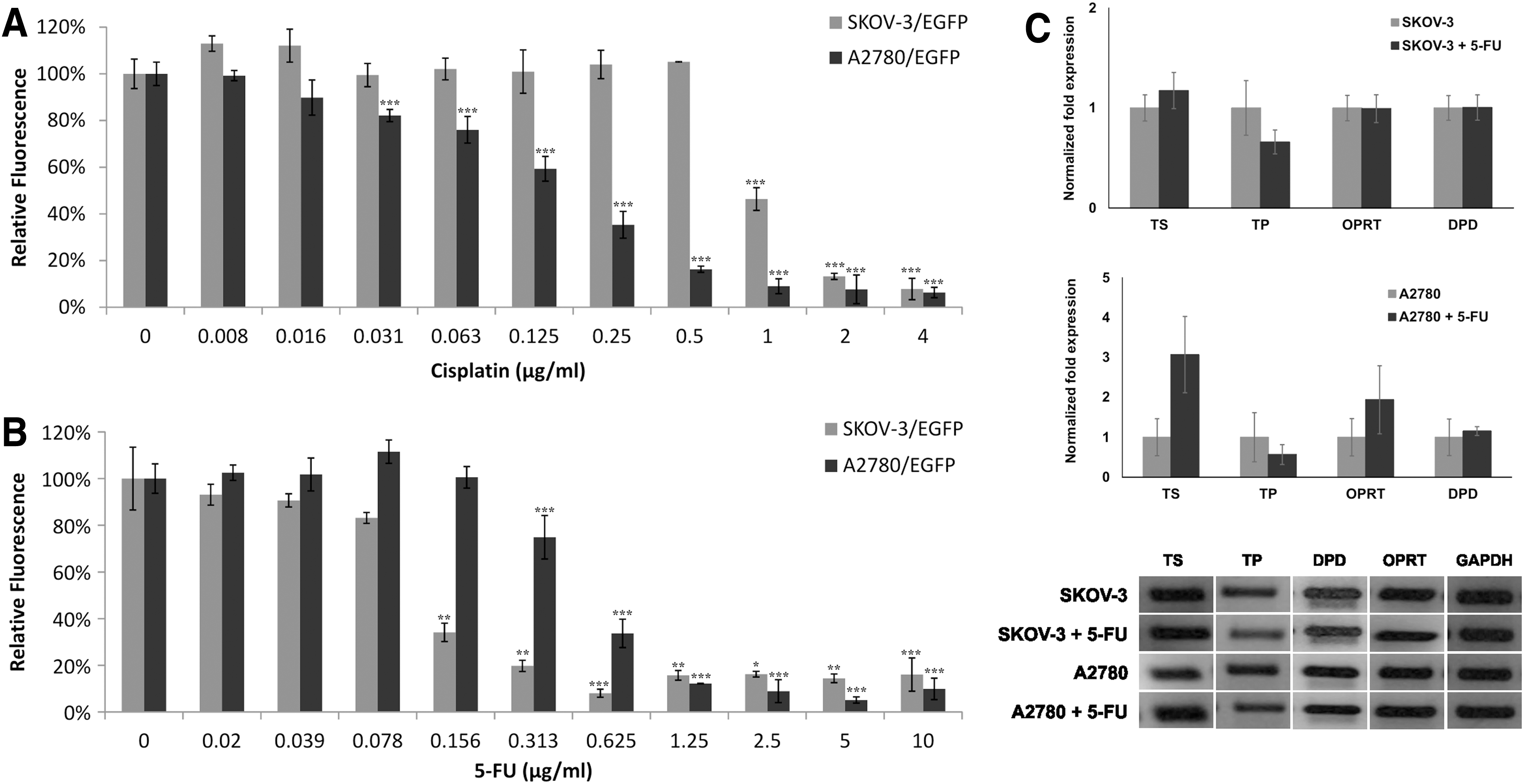
Ovarian cancer cells differ in drug sensitivity.
Coculturing of tumor cells and AT-MSC or CD::UPRT-MSC in the presence of 5-FC prodrug showed higher response of SKOV-3 to bystander effect of CD:UPRT-MSC/5-FC treatment than A2780 cells in vitro (Fig. 3A, B). Remarkably, the ratio 1:160 of CD::UPRT-MSC to SKOV-3 was efficient enough to eliminate 60%–80% of tumor cells (Fig. 3C). On the contrary, after coculture of A2780 with CD::UPRT-MSC a higher concentration of 5-FC was necessary to reach IC50 and even the ratio of therapeutic cells to tumor cells was much lower (1:23–1:17) to obtain the same percentage (60%–80%) of eliminated A2780 cells like in the case of SKOV-3 cells (Fig. 3C). Higher number of CD::UPRT-MSC could not eliminate higher proportion of tumor cells by bystander effect in adherent cocultures (Fig. 3D).
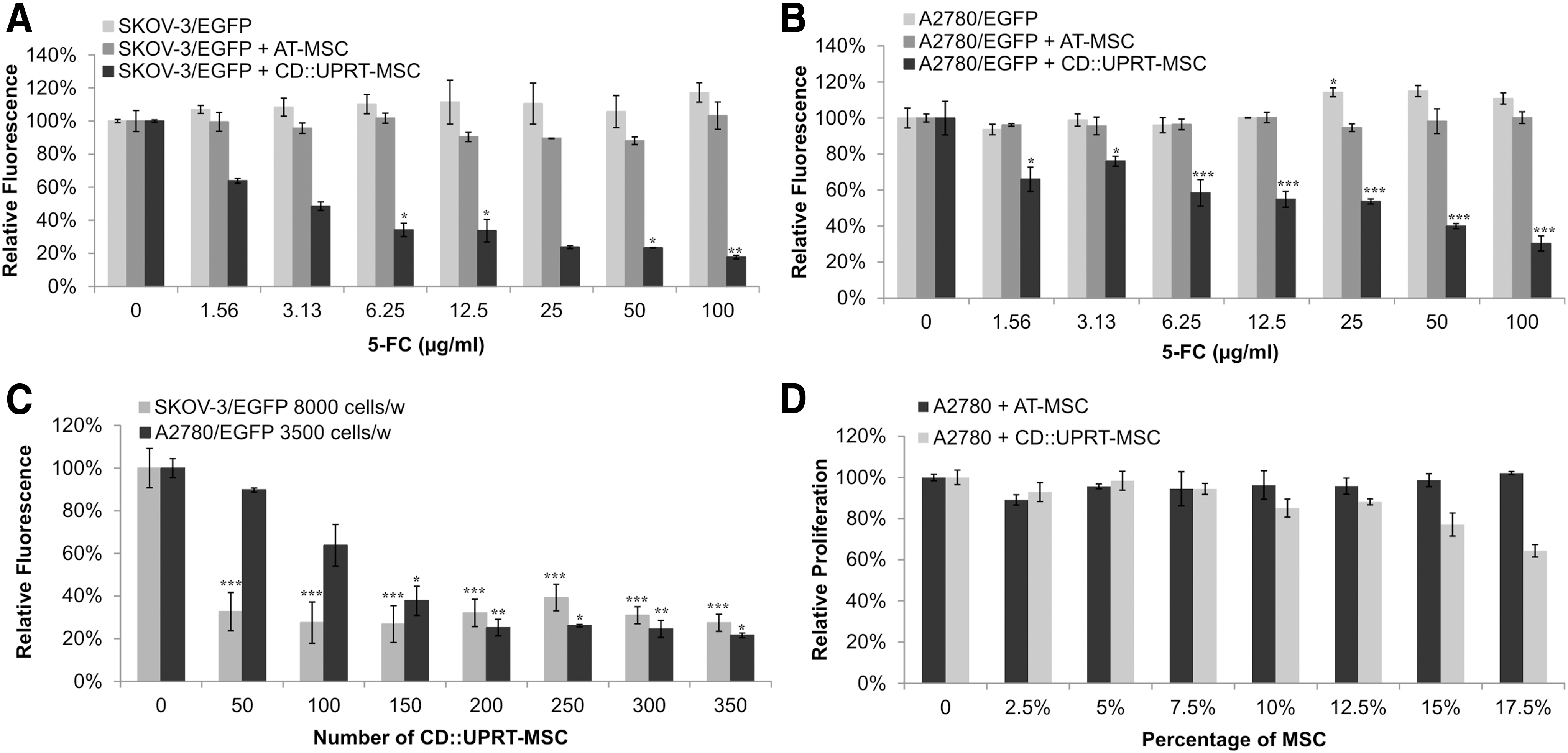
CD::UPRT-MSC/5-FC treatment shows high efficiency in A2780 cells in adherent cultures in vitro.
CD::UPRT-MSC/5-FC treatment on tumor cells in 3D culture predicts in vivo experiments
Based on our observation that 3D cultures can serve for prediction of therapeutic outcome [33] we decided to employ a 3D model for evaluation of treatment efficiency on SKOV-3 and A2780 cells. We compared the data from two-dimensional (2D) experiments with the 3D model of tumor cells treated with CD::UPRT-MSC/5-FC. We found significant differences in viability between SKOV-3 and A2780 cells after 3D culture (Fig. 4A, B). SKOV-3 cells were not able to form dense spheroid-like structures with/without CD::UPRT-MSC (data not shown), the structures were disintegrated, most of the CD::UPRT-MSC were on the surface of the aggregates and elimination of tumor cells was not efficient in comparison to controls even with high ratio of therapeutic cells to tumor cells (data not shown). We also measured viability of aggregates by chemiluminescent assay (evaluation of ATP content) at the experiment endpoint (Fig. 4C) and we reached only 36% decrease in aggregate's volume consisting the ratio 10:1 of SKOV-3/EGPF to CD::UPRT-MSC compared to control. Based on this outcome, the 3D modeling predicted poor response to the treatment mediated by therapeutic AT-MSC when the metastatic disease was propagated in vivo. A2780 cells formed very dense spheroid-like structures with CD::UPRT-MSC in the center within 48 h (Fig. 4B). Our data confirmed that CD::UPRT-MSC immensely supported the growth of A2780 if cultivated in standard media (as observed also in 2D model). We observed significant reduction of the aggregate's viability after addition of 5-FC. According to the performed viability assay, there was 90% reduction of aggregate's viability in comparison to controls, when ratio 1:5 CD::UPRT-MSC to A2780 was used in the presence of 5-FC (Fig. 4D; P ≤ 0.001).

Three-dimensional culture of tumor cells shows different cytotoxic responses to bystander effect mediated by CD::UPRT-MSC in the presence of 5-FC.
To investigate whether the 3D culture of human ovarian carcinoma cells correctly predicted the effectiveness of the CD::UPRT-MSC/5-FC treatment, we have injected SKOV-3 and A2780 cells s.c. and i.p. into nude mice. Subcutaneous administration of SKOV-3 cells resulted in the growth of xenotransplants when tumor cells were injected alone or with CD::UPRT-MSC (Fig. 5A). When mice were treated with 5-FC starting at day 0, tumors significantly decreased volume in comparison to untreated controls (P ≤ 0.01). The treatment initiated 3 days postinoculation resulted in no effect in animals inoculated with SKOV-3 and CD::UPRT-MSC. We did not find any visible signs of metastases in peritoneal cavity in both treated and untreated animals.
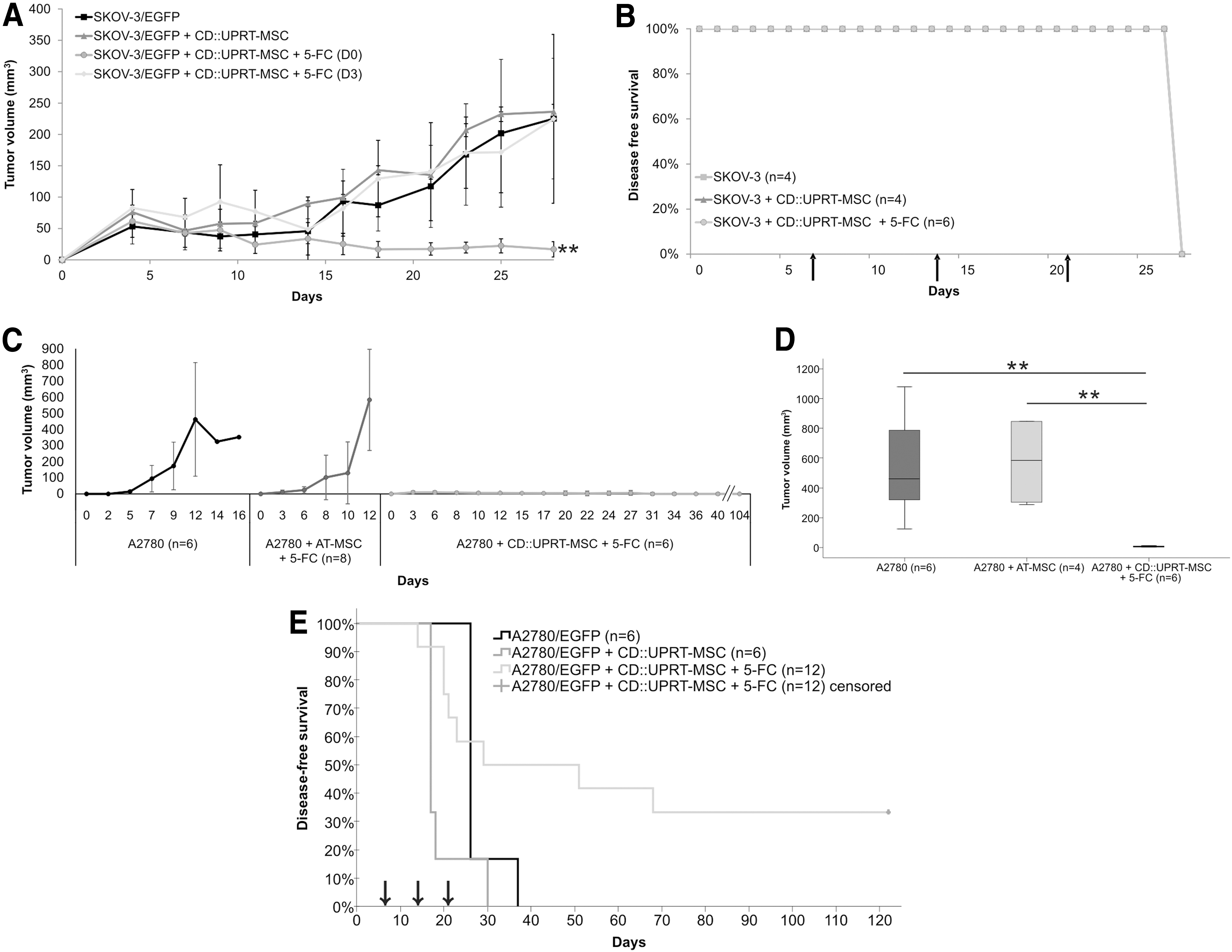
Long-term disease-free survival in 33% of metastases-bearing mice treated with CD::UPRT-MSC/5-FC.
SKOV-3 cells possess metastatic ability and therefore we injected them i.p. into mice, to investigate whether CD::UPRT-MSC/5-FC treatment will be effective in elimination of metastatic dissemination of tumor cells (Fig. 5B). Although we have administered the therapeutic AT-MSC three times i.p. (day 7, 14, and 21) and we treated animals with 5-FC, there was no difference among groups of animals examined for the presence of metastases. We had to euthanize all of them on the same day (28) due to moribund state—all mice had abdominal cavity full of blood, countless number of small-sized metastases attached to the intestine, peritoneum, and pleura as described by the group of Shaw et al. [34]. We did not observe any significant changes in the weight during the experiment; the moribund state slowly appeared ∼24 h before the animals had to be sacrificed.
We also administered A2780 cells (2 × 106) s.c. into nude mice alone or co-injected with AT-MSC or CD::UPRT-MSC and treated with 5-FC (i.p.). We showed that s.c. co-administration of A2780 cells with AT-MSC has led to immense growth of tumor volume, the penetrance was 100% and tumor take rate 8/8 in contrast to the control group. The penetrance in tumor cell alone group was only 50% (tumor take rate: 6/12) and in CD::UPRT-MSC co-injected group was 75% (tumor take rate: 6/8). The graph (Fig. 5C) depicts tumor growth kinetics of only growing tumors of mice in all groups at the indicated time points until the mice had to be euthanized. The combination of A2780 cells with CD::UPRT-MSC/5-FC has led to significantly decreased tumor volume in mice and also the overall survival was prolonged for more than 3 months in comparison to control animals. We compared the medians of growing tumor volume of each group at the day 12 (Fig. 5D) and we found a significant difference (P < 0.005) between the control group and the group treated with CD::UPRT-MSC/5-FC. The median tumor volume of control group was 462 mm3 in comparison to the treated group whose median of tumor volume was only 6 mm3.
Based on the efficiency demonstrated on s.c. model in vivo we also performed i.p. injection of A2780/EGFP cells into nude mice. We confirmed again that CD::UPRT-MSC alone can strongly support the growth of ovarian cancer cells in comparison to tumor cells alone (Fig. 5E). We examined all mice for the presence of metastases and we usually found one big and many small-sized tumors spread across the whole abdominal cavity attached to intestine, peritoneum, pleura, liver, and ovaries with no presence neither of ascitic fluid nor the blood. We observed prolonged disease-free survival in the group that received the CD::UPRT-MSC and were treated with 5-FC. We were able to achieve long-term tumor-free survival in 33% of the animals successfully treated from aggressive ovarian cancer as they were able to survive for more than 120 days. The animals, which did not show any signs of the disease, were examined at the experiment endpoint by necropsy and confirmed to be tumor-free.
Discussion
The ovarian carcinoma is the most lethal of all the gynecological malignancies. The major obstacle in the treatment represents the resistance to commonly used chemotherapeutics or insufficient delivery of efficient concentration of drugs to the tumor sites [35]. New therapeutic modality could be the exploitation of MSC in GDEPT [13,16,20,30,36,37]—they are immuno-privileged; due to their tumor tropism they can migrate and integrate into tumor mass and also track microscopic metastases [38,39]. This approach can facilitate direct chemotherapy in the vicinity of tumor cells, patients could be spared the adverse effects accompanying systemically administered chemotherapeutics (eg, 5-FU) and, moreover, the therapeutic AT-MSC undergo cell death in the presence of the prodrug [13]; therefore this GDEPT can be considered as a safe therapeutic approach. There are also many studies describing eradication of ovarian carcinoma cells using MSC encoding for anticancer genes.
Zheng et al. [40] genetically modified human placenta-derived mesenchymal stem cells using adenoviral vector carrying gene for human recombinant endostatin. These MSC possess migratory potential toward A2780 cells in vitro and they could significantly suppress the tumor growth in vivo after their i.p. administration into nude mice. The engineered MSC showed inhibition of tumor growth, tumor cell proliferation, and angiogenesis and Zheng et al. claimed a profound antitumor and antimetastatic ability of constructed MSC in a human ovarian carcinoma model. The group of Zhang et al. [41] used human umbilical cord mesenchymal stem cells expressing interleukin-21 (IL-21) via lentiviral vector on SKOV-3 xenograft-bearing mice. These MSC could markedly inhibit ovarian cancer growth and IL-21 may stimulate innate immune response (augmenting natural killer cells cytotoxicity). They speculated that modified MSC play an antitumor effect through regulating Wnt signaling pathway. Dembinski et al. [42] modified mesenchymal stem cells encoding gene for IFN-β via adenoviral transduction. They tested the efficacy of MSC-IFN-β using SCID mice i.p. injected with ovarian tumor cells OVCAR-3 or SKOV-3. They administered i.p. MSC-IFN-β once per week for 5 weeks and they could reach significantly extended or prolonged survival in mice bearing established OVCAR-3 or SKOV-3 xenografts treated with therapeutic MSC. Nowadays, there is a clinical study (NCT02530047), which will try to find the proper dose of human mesenchymal stem cells with interferon-β that can be given to patients with ovarian cancer.
In our experiments we used genetically modified AT-MSC to examine their therapeutic potential to eliminate SKOV-3 and A2780 cells in two different experimental setups in vitro and in vivo. We compared adherent and nonadherent in vitro and s.c. and metastatic in vivo experiments using CD::UPRT-MSC/5-FC treatment on tumor cells.
The capability of both tumor cell lines to form GJIC predicted sensitivity to HSVtk-MSC/GCV treatment in our model. Umhauer et al. [43] describes tight gap junctions as plasma membrane channels, which permit cells to exchange small molecules and ions. Even though several human ovarian adenocarcinoma cell lines miss the major subunit Connexin 43, we showed that both SKOV-3 and A2780 cells possess functional GJIC (Fig. 1A–C). Connexin 43 is inevitable for successful GJIC between tumor cells and HSVtk-MSC and according to our data both tumor cell lines can respond to treatment in the presence of GCV. We assume that low efficiency of HSVtk-MSC/GCV treatment on A2780 cells can be explained by the mechanism of action of GCV. This molecule after its phosphorylation creates toxic GCV-3P, which incorporates into the replicating DNA (not into RNA), and thus only dividing cells are destroyed. The quiescent cells can avoid cytotoxicity and they can grow after removing this chemotherapeutical agent from their environment. The same inefficiency of HSVtk-MSC/GCV treatment showed Matuskova et al. [16], when they cocultured melanoma cells A375 with HSVtk-MSC/GCV. Despite the fact, that A375 cells are able to communicate via tight gap junctions, the efficiency of the used treatment was limited because of the active multidrug-resistant protein 4 (MRP4; ABCC4). This protein was responsible for active efflux of GCV out of the cells [44].
Due to a different mechanism of action and target molecules another widely used approach of GDEPT exploits cytosine deaminase/5-FC treatment. Kucerova et al. [31] showed that therapeutic AT-MSC expressing fused yeast CD::UPRT in the presence of 5-FC can eradicate chemoresistant medullary thyroid carcinoma cells and this gene therapy is also promising for destroying melanoma or breast carcinoma cells [16,27]. Hartkopf et al. [45] used oncolytic measles vaccine virus expressing gene for CD::UPRT (MeV-SCD) to infect ovarian cancer cells lines SKOV-3 and OAW42 and primary cell cultures derived from malignant ascites of patients with ovarian cancer. They reached significant cell elimination after addition of 5-FC in stable cell lines but it was necessary to use higher concentration of a prodrug to reach comparable levels of cell death in primary cultures. The group of Jiang et al. [46] used human umbilical cord blood-derived MSC (hUCB-MSC) carrying double suicide gene encoding for CD and HSVtk. They clearly showed tumor tropism of genetically modified hUCB-MSC toward SKOV-3 cells and also efficient tumor cell killing while cocultured with therapeutic hUCB-MSC in the presence of 5-FC and GCV.
In our study, we also pointed out the use of MSC in cancer gene therapy. We showed that strong bystander effect of CD::UPRT-MSC/5-FC can effectively eliminate ovarian cancer cells but our data also confirmed that nontherapeutic MSC have promoted the ability of SKOV-3 and A2780 cells to grow more than tumor cells alone. We confirmed this protumorigenic effect of AT-MSC not only in adherent cultures but also in 3D culture and in vivo experiments; likewise, it was demonstrated by Kucerova et al. in the melanoma model [47]. There are many reviews [9,48,49] that sum the supportive and suppressive role of MSC in tumor biology, which depends on a very complex and not well-known tumor-MSC crosstalk. Nevertheless, in our further investigation we will try to reveal why the residual living population of tumor cells remains resistant to the treatment and the cause why the higher amount of therapeutic AT-MSC in the presence of a prodrug does not eliminate more tumor cells but it can support their growth.
Adherent culture is the most traditional design of many experiments in cancer biology, but it possesses a lot of limitations and it remains rather artificial [33]. Spheroid formation exhibits histological and physiological features very similar to those of solid tumors and in the review of Cesarz and Tamama [50] is mentioned that the responses of tumor cells to the treatment in vivo is better replicated in 3D culture than in 2D adherent culture and that also tumor 3D formations may possibly simulate circulating tumor aggregates and mimic the tumor microenvironment. Our study has focused on CD::UPRT-MSC/5-FC treatment on SKOV-3 and A2780 multicellular spheroids monitored by cell kinetic imaging system. We have observed significant differences in responses to treatment when we compared control and treated A2780 cells. Data showed that 90% of tumor cells were destroyed and we observed the aforementioned immense increase of aggregate's volume while cultured in control media, confirmed by chemiluminescent assay. SKOV-3 cells did not react on treatment, which has correlated with in vivo data. According to previous reports [51,52], we assume that the cause of the chemoresistance can be the presence of cancer stem cells (CSC) or due to increased expression of ATP-binding cassettes (ABC transporters), which can efflux the toxic metabolites out of the tumor cells. CSC is a well-known term for s subpopulation of tumor cells linked with tumor initiation, increased proliferation, occurrence of relapse, formation of metastases, and chemoresistance to commonly used chemotherapeutics. ABC transporters are responsible for the transport of substrate across the membrane against a concentration gradient [53,54], and their overexpression in tumor cells is one of the major causes of multidrug resistance (MDR). Nowadays, they are numerous studies investigating the ways of blocking their efflux function (eg, tyrosine kinase inhibitors; phosphodiesterase-5 inhibitors) or their use with conventional chemotherapeutics to increase the response to the anticancer treatment [55,56]. Currently, there are several ABC transporters involved in MDR of ovarian carcinomas effluxing platinum derivates or taxanes (eg, ABCA1; ABCA3; ABCC2; ABCC6; ABCC10, ABCG2) (reviewed in Ween et al. [57]). SKOV-3 cells have increased expression of ABCG2 in comparison to A2780 (data not shown) and as the further aim we will examine the subpopulation of CSC in SKOV-3 and A2780 cultures and determine their expression of ABC transporters. The use of three-dimensional (3D) models fills the gap between 2D cultures and in vivo experiments and indeed, we clearly showed that 3D model can predict the efficiency of elimination of tumor cells in experimental animals.
We showed decreased average tumor volume in mice after s.c. administration of SKOV-3 cells with CD::UPRT-MSC in the presence of 5-FC (since the day 0) in comparison to other groups. We observed significant difference between the control group and animals treated on the same day as they were injected with tumor cells. There is an experiment performed by the group of Nouri et al. [58], where they exploited CD::UPRT-MSC/5-FC treatment on SKOV-3 s.c. xenografts in nude mice. Once the tumors reached adequate volume, animals were injected intratumorally with therapeutic AT-MSC once per week (six doses) and consequently treated with 5-FC. The data showed significant inhibition of tumor growth in the treated group, but complete eradication was not reached. Nouri et al. suggested for achieving complete tumor remission that either higher amount of therapeutic MSC could be used or use of other GDEPT system would bring some improvements. According to our results, we assume that more important than aforementioned reasons is the right timing of administration of a prodrug into animals [59]. Also the 3D model of SKOV-3 cells predicted the result from in vivo experiment, when tumor cells were injected in mice i.p. and the CD::UPRT-MSC/5-FC treatment was used. Even the i.p. administration of 5-FC did not prevent formation of SKOV-3 metastases. All mice had to be sacrificed on the same day and in all of them metastases were present within the whole abdominal cavity.
Notably, the use of CD::UPRT-MSC/5-FC treatment has shown remarkable strong antitumor results in treating s.c. and metastatic tumors established from A2780 cells. The treatment decreased the volume of tumors (s.c.) and prolonged the disease-free survival of animals (i.p.) compared to untreated controls.
Taken together, this study has been focused on two different systems of the gene therapy mediated by genetically modified MSC on ovarian carcinoma cell lines. We compared adherent and 3D in vitro models of CD::UPRT-MSC/5-FC treatment and we proved the predictive potential of 3D cultures whose result can be expected in in vivo experiments. We emphasize that the use of therapeutic AT-MSC in combination with 5-FC decreased the tumor volume and prolonged the disease-free survival in mice with highly metastatic ovarian cancer.
Footnotes
Acknowledgments
We thank Z. Kozovska for help with qRT-PCR, M. Bohac for providing material for AT-MSC isolation, E. Durinikova and S. Miklikova for isolation and expansion of AT-MSC, and M. Dubrovcakova and V. Frivalska for the excellent technical assistance. This study was supported by the VEGA grant nos. 2/0087/15 (L.K.) and 2/0171/13 (M.M.) and by the Slovak Research and Development Agency under the contract nos. APVV-0230-11 and APVV-0052-12. The experiments on the CFX96 Real-Time PCR Detection System and the IncuCyte ZOOM were enabled with the kind help and financial support from the Cancer Research Foundation RFL2009 and RFL2012.
Author Disclosure Statement
No competing financial interests exist.