
Abstract
Nuclear reprogramming efficiency has been shown to be highly variable among different types of somatic cells and different individuals, yet the underlying mechanism remains largely unknown. Several studies have shown that reprogramming of fibroblasts into induced pluripotent stem cells (iPSCs) requires remodeling of mitochondria and a metabolic shift from an oxidative state to a more glycolytic state. In this study, we evaluated the nuclear reprogramming efficiency in relation to mitochondrial bioenergetic parameters of fibroblasts from seven different human individuals. Using the Seahorse extracellular energy flux analyzer, we measured oxygen consumption rate (OCR) profiles of the cells, along with their nuclear reprogramming efficiency into iPSCs. Our results showed that fibroblasts with the lowest mitochondrial spare respiratory capacity (SRC) had the highest nuclear reprogramming efficiency, opposed to fibroblasts with the highest mitochondrial SRC, which showed lowest reprogramming efficiency. Furthermore, we found that targeted fluorescent tagging of endogenous genes (MYH6 and COL2A1) by CRISPR/Cas9-mediated homologous recombination was accompanied by an increase in the SRC level of the modified fibroblasts and impaired reprogramming efficiency. Our findings indicate a negative correlation between high mitochondrial SRC in somatic cells and low reprogramming efficiencies. This type of analysis potentially allows screening and predicting reprogramming efficiency before reprogramming, and further suggests that nuclear reprogramming might be improved by approaches that modulate the SRC.
Introduction
S
All living cells need energy to perform their vital biological processes. Mitochondria are the powerhouse of the cell and, therefore, a central platform to produce energy for various cellular activities [8]. Mitochondria provide energy in the form of adenosine triphosphate (ATP) by oxidative phosphorylation (OXPHOS). OXPHOS is performed by the mitochondrial respiratory chain organized into five complexes (I–V) [9,10]. However, mitochondrial activities have also been shown to be associated with oxidative stress, cell death [11,12], aging [13,14], and human diseases [15 –17].
Recently, a growing body of evidence indicates an association between mitochondrial function and pluripotency [18 –21]. It has been demonstrated that somatic cells convert from an oxidative state to a more glycolytic state during the transition of differentiated cell status to pluripotency, which suggested a positive correlation between preferential production of lactate from glycolytic pyruvate and reprogramming efficiency, where somatic cells with higher lactate production displayed higher reprogramming efficiency [19].
A well-recognized mitochondrial bioenergetic parameter is the reserve or spare respiratory capacity (SRC), which is the difference between the maximum mitochondrial oxygen consumption rate (OCR), measured after uncoupling proton pumping by the electron transport chain and ATP synthesis, and the basal mitochondrial OCR. It has been indicated that SRC is regulated by cellular metabolic sensors [22]. Cells can utilize SRC to adapt to increased energy demand upon increase in workload or cellular activity. The mitochondrial SRC has been suggested to be critical for cellular survival and function [23 –27]. Previously, it has been reported that iPSCs and embryonic stem cells (ESCs) display reduced mitochondrial SRC, whereas somatic cells have a significantly higher SRC level [28]. This suggests that the somatic SRC status has to be restored to the pluripotent state upon establishment of pluripotency by nuclear reprogramming.
In this study, we used the Seahorse extracellular energy flux analyzer to determine mitochondrial bioenergetics parameters in human fibroblasts derived from seven different donors, and related the results to their reprogramming capacity. Our study indicates that somatic cells with lower mitochondrial SRC had higher nuclear reprogramming efficiency, and cells with higher SRC showed lower reprogramming efficiency. Furthermore, genetic modification of fibroblasts, which is accompanied with relatively long-term in vitro cultivation, may increase SRC and decrease the reprogramming efficiency. We also showed that, once pluripotency is established and disregarding earlier mitochondrial performance, the iPSCs produced from different fibroblast cells are similar in pluripotency and differentiation capacity.
Materials and Methods
Cell culture
Primary human dermal fibroblast cells were isolated as previously described [29], under the ethical permission (Danish Regional Committee on Health Research Ethics, H-4-2011-157). Fibroblasts were cultured in high glucose DMEM medium (BE12-604F, LONZA) with 10% fetal bovine serum, 2 mM
During the induction of human iPSCs, transduced cells were cultured on mitomycin-C-treated mouse embryonic fibroblasts (MEF) with iPSC induction medium (knockout DMEM (10829-018; Gibco), 20% KSR (10828-028; Gibco), 1% NEAA (11140-050; Gibco), 0.2% 2-mercaptoethanol (31350-010; Gibco), 1% GlutaMAX, 1% penicillin/streptomycin, and 10 ng/mL bFGF (PHG0026; Invitrogen).
Human iPSCs and ESCs (H9, generously provided by Associate Professor Mark Denham, Department of Biomedicine, Aarhus University) were maintained on VTN-N (A1413301; Life Technologies)-coated four-well dishes with Essential 8 medium (A1517001; Life Technologies) as described previously [30]. The medium was changed daily, and the iPSCs were manually passaged or using EDTA (0.05 mM) upon 80–90% confluence.
Mitochondrial bioenergetic measurements
To assess mitochondrial respiration in cells, an XF96 extracellular flux analyzer (Seahorse Bioscience) was used to measure OCR and extracellular acidification rate (ECAR). Fifteen thousand fibroblasts or 5000 iPSCs/ESCs were seeded to each well of a FX96 cell culture microplate and incubated overnight at 37°C in a humidified atmosphere of 5% CO2. One hour before the assay, culture medium was changed into unbuffered DMEM supplemented with glutamine, sodium pyruvate, and glucose and the culture plate was incubated in a non-CO2 incubator at 37°C.
The XF Cell Mito Stress Test Seahorse Bioscience was used to measure the key parameters of mitochondrial respiration, using specific mitochondrial inhibitors and uncouplers. Oligomycin, FCCP, and a mixture of rotenone/antimycin A were injected in subsequential manner into each well following the manufacturer's recommendations. First, the level of oxygen consumption was measured without any additions, which represents the basal OCR. Next, oligomycin was injected to inhibit ATP synthase (complex V) and OCR was recorded. To determine the maximal OCR, the uncoupler FCCP was injected. Finally, a mixture of rotenone/antimycin A was injected to inhibit the flux of electrons through complex I and III, and thus shutting down mitochondrial respiration (Fig. 1A). After the assay, all measurements were normalized to total protein concentration using Bradford protein assay (Bio-Rad).
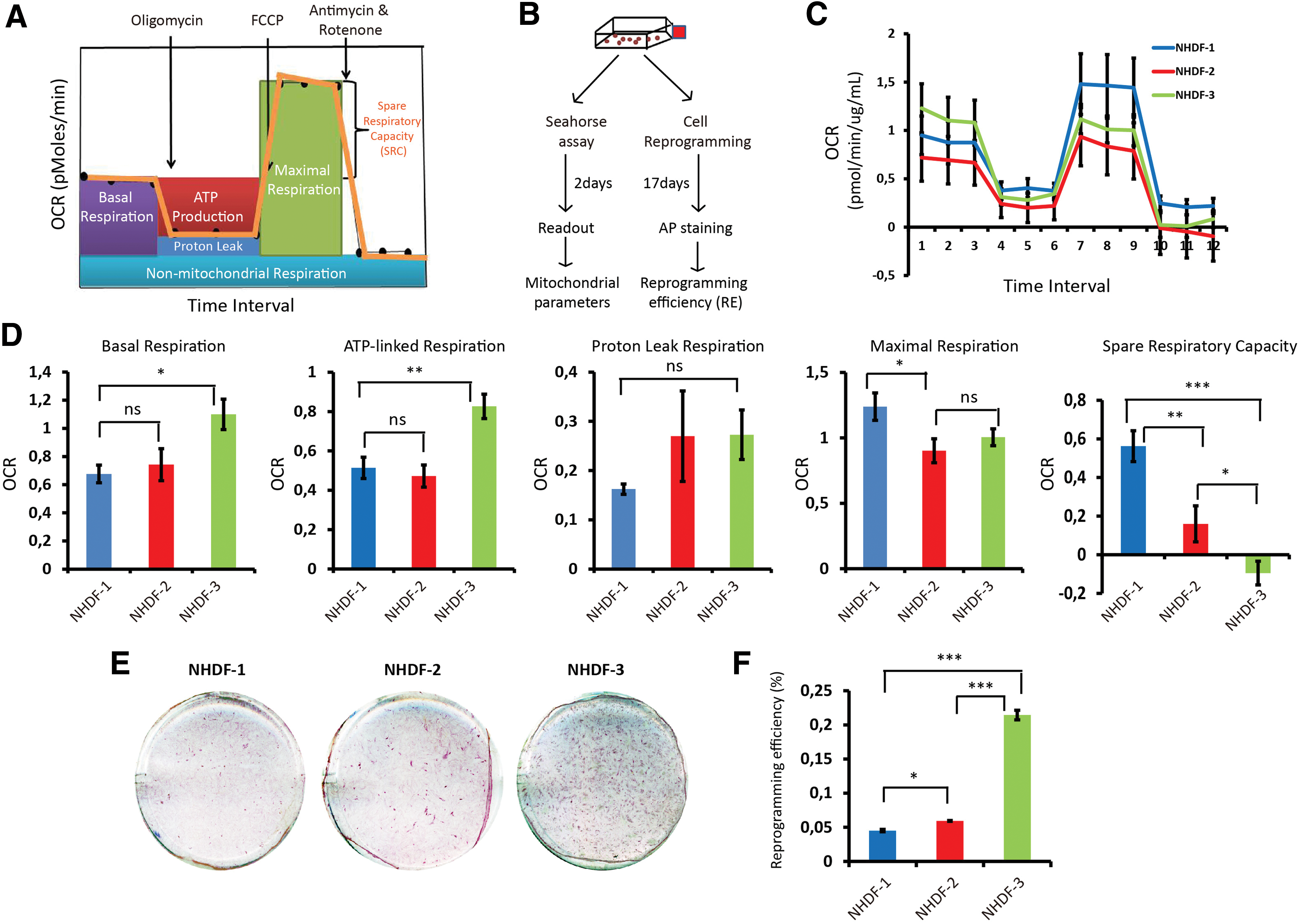
Bioenergetic profiles of three different fibroblasts and their iPSC reprogramming efficiency.
Human iPSC induction
Induction of human fibroblasts into iPSCs was performed by lentiviral-mediated nuclear reprogramming, as described previously with minor modifications [31]. Briefly, a polycistronic lentiviral plasmid (pRRL.PPT.SF.hOCT3/4.hKLF4.hSOX2.hMYC.dTOMATO (LV-OKSM-Tomato), a generous gift from Axel Schambach [32]), containing the four reprogramming transcription factors (OCT4, KLF4, SOX2, and C-MYC) and a fluorescent marker gene in a single transcript, was packaged into lentiviral particles by transient cotransfection with third-generation lentiviral packaging plasmids into HEK293T cells and concentrated by ultracentrifugation as described previously [33]. Viral titer was determined using the HIV-1 p24 ELISA Kit (XpressBio; XB-1000).
One day before transduction, 1.5 × 10(5) fibroblasts were seeded per well in a 0.1% gelatin-coated six-well plate. The cells were transduced with the pRRL-OKSM virus (10 ng, p24 measuring viral titer) in the presence of 8 μg/mL polybrene, here defined as day 0. On day 1, the medium was replaced with standard fibroblast medium and changed every 2 days. On day 4, the transduced cells were trypsinized, and 1 × 10(5) cells were seeded per well on mitomycin-C-treated MEFs in fibroblasts medium in six-well plates. From day 5, the medium was changed to human iPSC induction medium and changed daily. From day 14, colonies with round and compact morphologies appeared. On day 17, ESC-like, alkaline phosphatase (AP)-positive and TRA-1-60-poisitive colonies were counted as primary iPSCs. Primary iPSC colonies were manually picked for expansion onto fresh feeder cells or feeder-free dishes for further characterization.
Alkaline phosphatase staining
For quantification of iPSC reprogramming efficiency, alkaline phosphatase (AP) staining was performed to define the number of iPSC colonies (Sigma, Alkaline Phosphatase Staining Kit, 86) following the manufacturer's instructions. Briefly, on day 17 after induction, cells were immersed in fixation solution for 5 min, followed by 3× washes with deionized water, incubating in alkaline dye solution for 15 min, and 3× washes with deionized water. The individual ESC-like and AP-positive colonies were visualized under microscope. The number of iPSC colonies was counted. Reprogramming efficiency was calculated on the basis of the number of AP-positive colonies divided by the initial number of reprogramming cells. Three independent reprogramming experiments were performed for each NHDF.
Fluorescent live-cell imaging of pluripotent marker TRA-1-60
Live cell imaging of the cell surface pluripotent marker TRA-1-60 was performed as described by the Stem Cell Antibody Kits for Live Cell Imaging (Cat. no. A25618, Life Technologies).
Cell proliferation rate assay by the Click-iT® EdU Alexa Fluor® 488 Imaging Kit
Quantification of fibroblasts cell proliferation rate was conducted using the Click-iT® EdU Alexa Fluor® 488 Imaging Kit (C10337, Thermo Fisher Scientific) according to the manufacturer's instructions. Briefly, fibroblasts were seeded in six-well plates (1.0 × 105 cells/well) 24 h before changing to fresh growth medium (negative control) or growth medium containing 10 μM EdU for 2 h. Then cells were harvested by trypsinization and subjected to EdU staining according to the manufacturer's protocol. The cells were washed with 500 μL 1% bovine serum albumin (BSA)-PBS, incubated with 100 μL Click-iT fixative for 15 min in the dark, washed with 500 μL 1% BSA-PBS, resuspended with 100 μL 1× component E, stained in labeling cocktail for 30 min, washed with 500 μL 1× component E, and analyzed by flow cytometry (LSRFortessa); 10,000 events were recorded for each sample and data were analyzed with FlowJo 10.
Cell proliferation rate assay by WST-1
Fibroblasts (5,000 cells/well) were plated to a 96-well tissue culture plate and cultured for 48 h. One hour before the assay, Cell Proliferation Reagent WST-1 was added (10 μL/well) to the cells already cultured in 100 μL/well (1:10 final dilution). The Formazan dye formed through the cleavage of stable tetrazolium salt WST-1 by metabolically active cells was quantified with a scanning multiwell spectrophotometer at ab450nm.
2-deoxyglucose treatment
2-deoxyglucose (2DG) was purchased from Sigma (D8375-10 MG). The working concentration of 2DG used was 1 mM for both fibroblasts and fibroblasts under reprogramming.
Immunofluorescence staining of pluripotent markers in iPSCs
For immunofluorescence staining of pluripotent markers, the iPSC colonies were cultured on VTN-coated four-well Chamber Slides (Lab-Tek; 177437) in E8 medium. When the iPSC colonies were 60–70% confluent, the E8 medium was removed, and the cells were washed once with PBS. The iPSC colonies were fixed with 4% PFA for 30 min at room temperature, followed by two rinses in PBS and permeabilization with 0.3% Triton X-100-PBS for 30 min, and subsequent blocking with 5% donkey serum for 30 min. Afterward, the cells were incubated with primary antibodies against OCT4 (ab27985, 1:200 dilution; Abcam), NANOG (ab80892, 1:100 dilution; Abcam), SSEA-3 (ab16286, 1:100 dilution; Abcam), SSEA-4 (ab16287, 1:100 dilution; Abcam), or TRA-1-60 (ab16288, 1:100 dilution; Abcam) at 4°C overnight. Cells were then washed with PBS three times and incubated with a secondary antibody for 2 h in the dark. The secondary antibodies used were donkey anti-goat IgG Alexa Fluor 594, donkey anti-rabbit IgG Alexa Fluor 488, goat anti-rat IgM Alexa Fluor 594, goat anti-mouse IgG Alexa Fluor 594, and goat anti-mouse IgM Alexa Fluor 594 (all from Life Technologies, 1,000 × dilution). Nuclei of cells were visualized with DAPI staining for 10 min. The slides were mounted and images were captured with a fluorescence microscope at the indicated magnification (100×).
Embryoid body formation and differentiation
Embryoid body (EB) formation and differentiation were performed as previously described [34] with small modifications. Briefly, iPSCs were dissociated with PBS-EDTA (0.5 mM) for 5 min at 37°C, and 100,000 cells were replated per well onto 24-well ultra-low attachment plates (Corning), and cultured in E8 medium for 1 week with medium change daily. The mature EBs with round and aggregating morphology were transferred to 0.1% gelatin-coated four-well chamber slides with differentiation medium (high glucose DMEM medium supplemented with 10% FBS, 2 mM
Quantification of lentiviral transduction efficiency
The fibroblasts were seeded onto six-well plates (triplicate for each fibroblast line) at a cell density of 1.5 × 10(5) cell/well. Twenty-four hours later, the cells were transduced with lentivirus, expressing the green fluorescent protein gene (a titer of 3 ng p24), supplemented with polybrene (8 μg/mL). Twenty-four hours after transduction, the transduced cells were changed to fresh fibroblast medium. Seventy-two hours after transduction, transduction efficiency was analyzed by fluorescence imaging and flow cytometry analysis. For flow cytometry analysis, cells were trypsinized, washed once with 2% FBS-PBS, fixed in 1% formaldehyde for 10 min, washed with 2% FBS-PBS twice, and resuspended into 500 μL PBS. Ten thousand events were recorded for each sample and analyzed with FlowJo 10.
Statistics
Data are presented as the mean ± SD. Statistical analyses among different groups were performed using one-way ANOVA test with Bonferroni correction test. Asterisks represent P values of <0.05 “*,” <0.01 “**,” and <0.001 “***.”
Results
Nuclear reprogramming efficiency is associated with somatic mitochondrial SRC
To investigate mitochondrial bioenergetic parameters in human fibroblasts, we selected the XF96 extracellular flux analyzer, which measures OCR, a marker of OXPHOS flux, in a large-scale and real-time setting (96 wells). The bioenergetics profile, including basal respiration, ATP-linked respiration, proton leak, maximal respiration, and SRC can be calculated in a single assay (Fig. 1A).
To investigate the correlation between mitochondrial respiratory function and nuclear reprogramming efficiency, we selected normal human dermal fibroblasts (NHDFs) derived from three healthy individuals (NHDF-1, NHDF-2, and NHDF-3) (Table 1). For each NHDF, the cells were subjected to the Seahorse assay and reprogramming concordantly (Fig. 1B). All three NHDFs responded to the treatments with oligomycin, FCCP, and rotenone/antimycin A (Fig. 1C). When we calculated the different OCR rates for basal respiration, ATP-linked respiration, proton leak, maximal respiration, and SRC, we observed considerable variation between the three NHDFs (Fig. 1D). Notably, NHDF-3 cells consistently displayed higher basal respiration level, ATP-linked respiration, and proton leak than NHDF-1 and NHDF-2 (Fig. 1D). In contrast, the SRC level of NHDF-3 was significantly lower than those of NHDF-1 and NHDF-2 (P < 0.01, ANOVA).
When analyzing the nuclear reprogramming efficiency, we first measured the transduction efficiency of these three NHDFs using an enhanced fluorescent protein (EGFP) lentivirus to evaluate whether the fibroblasts differed in their capacity to take up lentivirus. The transduction efficiency in all NHDFs was similar, with an efficiency >70% (Supplementary Fig. S1A; Supplementary Data are available online at
Validation of the negative correlation between mitochondrial SRC and nuclear reprogramming efficiency using four additional fibroblast lines
To further validate the tentative correlation between mitochondrial SRC and reprogramming efficiency, we performed bioenergetic measurements and reprogramming of fibroblasts from further four different individuals (NHDF-PK236, NHDF-PK237, NHDF-PK241, and NHDF-PK242, Table 1). These four NHDFs, together with an iPSC line previously generated in our group [31] and a human embryonic stem cell line (H9ESC) were subjected to Seahorse bioenergetic measurements (Fig. 2A, B). The results showed that the relative SRC levels of PK237 and PK241 were significantly higher (almost two-folds) than those of PK236 and PK242 (P < 0.001). We also confirmed that both iPSCs and ESCs have very low SRC levels (even negative OCR value) in accordance with previous reports [19,35]. In parallel, the four NHDFs were reprogrammed into iPSCs, and their respective reprogramming efficiencies were determined. All four NHDFs were successfully reprogrammed to ESC-like colonies at day 17 after transduction. AP staining revealed prominently higher reprogramming efficiencies for PK236 and PK242 compared with PK237 and PK241 (0.35% and 0.33% for PK236 and PK242, and 0.09% and 0.11% for PK237 and PK241, P < 0.001) (Fig. 2C, D). These results further support our finding that somatic mitochondrial SRC is negatively correlated with reprogramming efficiency.
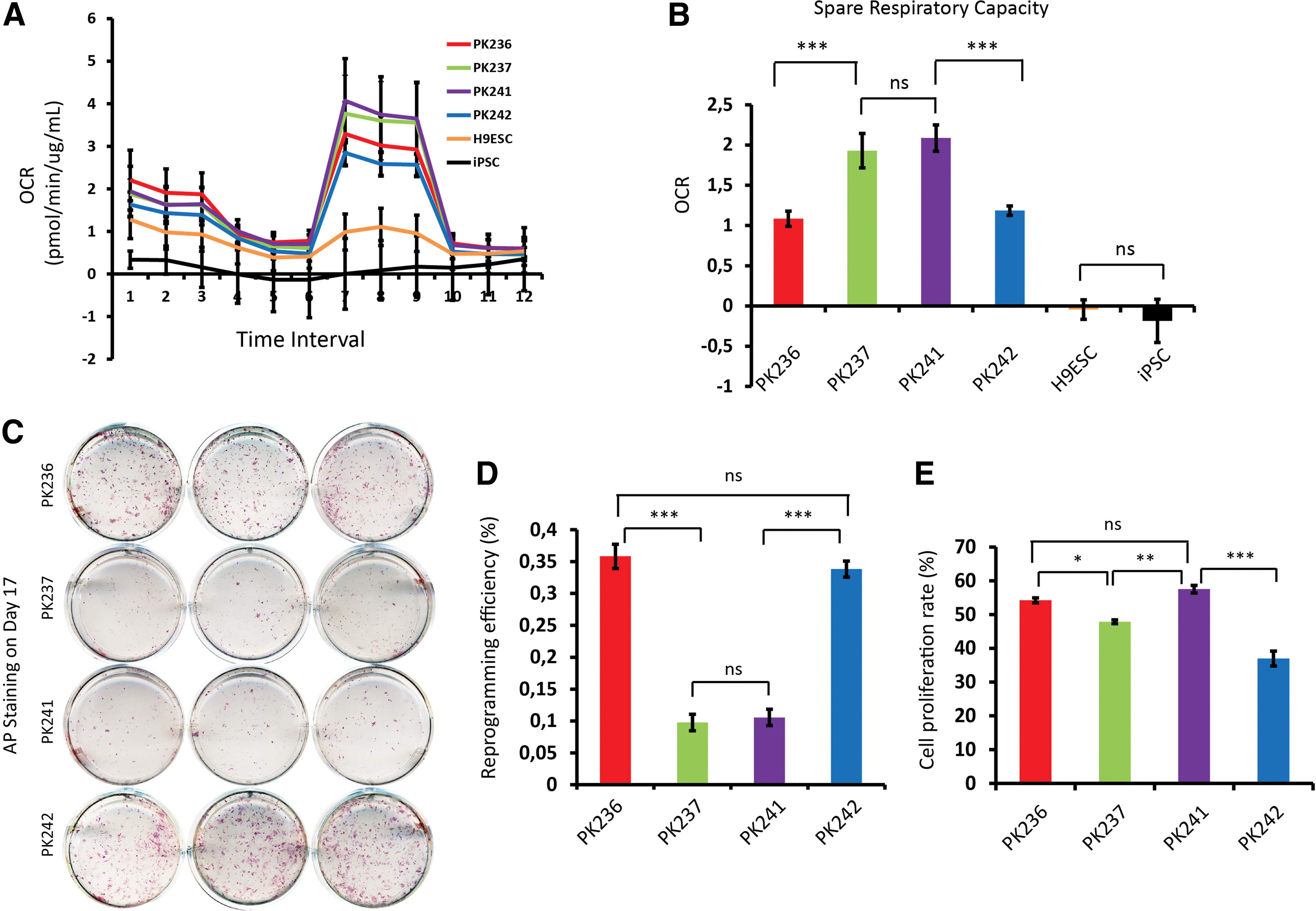
Validation of reverse correction of spare respiratory capacity and reprogramming efficiency in another four NHDFs.
A previous study reported that the cell proliferation rate is negatively correlated with overall reprogramming efficiency [36]. Using a highly sensitive EdU-based proliferation assay, our results revealed that the proliferation rate of NHDF-PK242 (37% ± 2.19%) was significantly lower (P < 0.01, ANOVA) than NHDF-PK236 (54% ± 0.7%), NHDF-PK237 (48% ± 0.6%), and NHDF-PK241 (57% ± 1.1%) (Fig. 2E and Supplementary Fig. S3). However, no such negative correlation between proliferation rate and reprogramming efficiency was observed for NHDF-PK236. Thus, to further validate the correlation between mitochondrial SRC and reprogramming efficiency, we treated fibroblasts with a molecule targeting energy production, 2DG, which has been reported to impair reprogramming efficiency [19]. Treatment of cultured fibroblasts with 2DG (1 mM) significantly inhibited the cell proliferation rate (P < 0.001) and increased SRC (P < 0.01) (Supplementary Fig. S4). Consistent with Folmes et al. we observed significantly (P < 0.001) decreased reprogramming efficiency through treating fibroblasts undergoing reprogramming with 2DG (Supplementary Fig. S4). However, the dramatic decrease in reprogramming efficiency by 2DG treatment may have resulted from the effects of 2DG on both cell proliferation and SRC. Nonetheless, our results further support a correlation between mitochondrial SRC and reprogramming efficiency.
Validation of iPSC pluripotency and SRC
Despite differences in reprogramming efficiency, all iPSCs generated from the four NHDFs had typical ESC-like morphology (Supplementary Fig. S5). To investigate whether the iPSC colonies retained pluripotent stem cell properties, we expanded one iPSC colony derived from each of the four parental fibroblasts (hereafter referred to as iPSC-PK236, iPSC-PK237, iPSC-PK241, and iPSC-PK242). First, using a Live-Cell Stem Cell Characterization Kit, we validated the positive expression of the pluripotency marker TRA1-60 in all four iPSC colonies maintained on E8-VTN feeder-free system (Supplementary Fig. S5). Furthermore, iPSC-PK236, iPSC-PK237, iPSC-PK241, and iPSC-PK242 were analyzed for expression of the pluripotency markers NANOG, OCT4, SSEA3, SSEA4, and TRA-1-60 by immunofluorescence staining. All four iPSC colonies were positive and indistinguishable from each other regarding the staining patterns for the investigated pluripotency markers (Fig. 3A). We further analyzed the iPSCs for their capability to differentiate into the three germ layers. Due to the limited resource and ethical considerations, we investigated the three-germ-layer differentiation capacities of iPSCs by means of EBs instead of teratoma assay. EBs were generated for each iPSC line, followed by a 3-week spontaneous differentiation. All iPSC-PK236, iPSC-PK237, iPSC-PK241, and iPSC-PK242 cell lines could differentiate into the three defined germ layers: ectoderm (TuJ1, neuron-specific class III beta-tubulin), endoderm (AFP), and mesoderm (SMA) (Fig. 3B), suggesting that the generated iPSCs were indeed pluripotent, and no differences in their pluripotent capacity could be observed. We also investigated the SRC of two iPSC lines (iPSC-PK241 and iPSC-PK242) originating from fibroblasts with high and low SRC, respectively, together with one control iPSC line. Consistent with what we observed above (Fig. 2B), both iPSC-PK241 and iPSC-PK242 displayed a negative SRC value similar to the control iPSCs (Fig. 3C). Our data are consistent with the notion that pluripotent stem cells are reset to a low SRC state, independent from the somatic SRC state observed in the fibroblasts they are derived from. This further supports the notion that mitochondrial metabolism plays an important role in nuclear reprogramming [37].

Validation of iPSC pluripotency.
In vitro gene editing of fibroblasts could increase SRC and decrease reprogramming efficiency
Fluorescent tagging of endogenous genes can be carried out in either fibroblasts or iPSCs. In a cellular disease model project we had previously C-terminally tagged the MYH6 gene in human fibroblasts NHDF-1 with an EGFP tag using CRISPR/Cas9 to introduce a cardiomyocyte marker that can be easily monitored [38]. When we attempted to reprogram the resulting CRISPR/Cas9-edited MYH6 fibroblasts, we could not obtain any fully reprogrammed iPSCs (only 2–3 partially reprogrammed colonies). In contrast, the original unedited NHDF-1 fibroblasts, reprogrammed in parallel, produced substantial number of iPSC clones (Fig. 4A, B). Since we had observed that somatic mitochondrial SRC is related to nuclear reprogramming efficiency, we wondered if genetic modification of fibroblasts, which is frequently accompanied with extended cultivation of the fibroblasts in vitro, had caused a change in SRC. Indeed, measurements of mitochondrial bioenergetics in the CRISPR/Cas9-edited MYH6 fibroblasts (MYH6) and the parental unedited fibroblasts (NHDF-1) revealed a significantly higher SRC level in the MYH6 cells compared with NHDF-1 (P < 0.05, Fig. 4C, D).
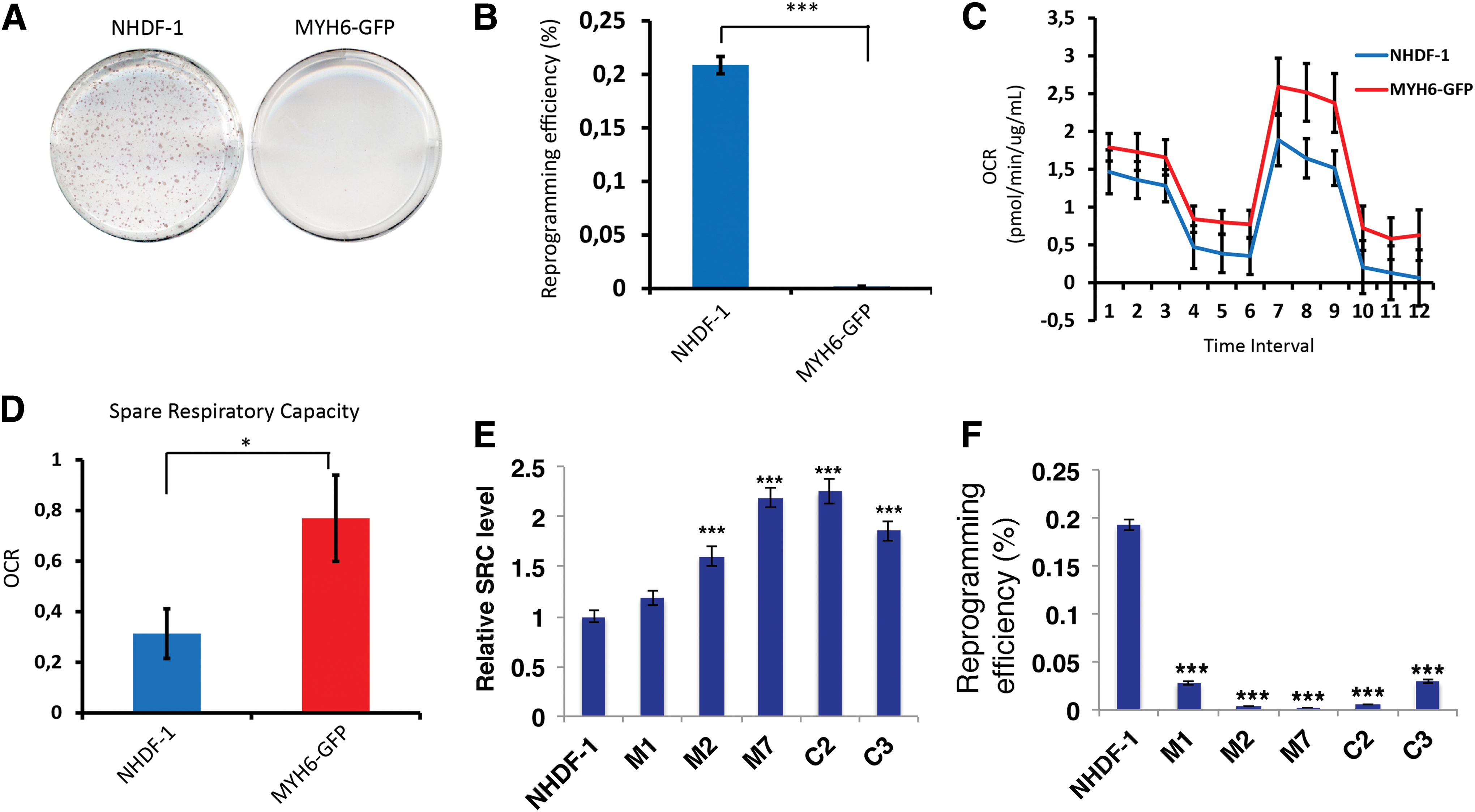
In vitro long-term genetic modification of fibroblasts increases spare respiratory capacity and decreases reprogramming efficiency.
To gather further evidence that this is not due to single clonal variation, we tested five additional gene-edited fibroblast cell clones: Three clones edited by CRISPR-mediated tagging of MHY-6 (M1, M2, M7) and two clones modified by CRISPR-mediated tagging of COL2A1 (C2 and C3) generated by us previously [38]. The SRC levels were significantly (P < 0.001, ANOVA with Bonferroni testing) higher in M2 (1.6-fold), M7 (2.2-fold), C2 (2.3-fold), and C3 (1.8-fold) compared with the parental fibroblast NHDF-1 (Fig. 4E), as well as decreased reprogramming efficiency (Fig. 4F). However, the reprogramming efficiency of fibroblast clone M1 was also greatly decreased, while there is only a slight increase, but not significant (P = 0.07, ANOVA) SRC compared with NHDF1 (Fig. 4E). This suggests that in addition to SRC, other unknown factors arising during the establishment of clonal gene-edited fibroblasts, a process requiring long-term in vitro cultivation, hamper reprogramming efficiency.
Discussion
In the present study, we used the XF96 extracellular flux analyzer to evaluate iPSC generation efficiency in relation to mitochondrial function and activity. Mitochondrial cellular bioenergetics have been shown to play a vital role in nuclear reprogramming [37,39]. We have characterized the bioenergetic profiles of fibroblasts to uncover the correlation between SRC and reprogramming efficiency. We assessed OCR in fibroblasts, and our findings indicate that fibroblasts with decreased ability to respire when treated with FCCP had greater reprogramming capacity.
Many factors have been identified influencing reprogramming efficiency, such as cell proliferation rate [36], TGF-beta signaling [40], WNT signaling [41], epigenetic remodeling [42] etc. Pluripotent stem cells are characterized by their self-renewal capacity and unattenuated cell proliferation capacity. Previous studies have implicated that signaling pathways regulating cell proliferation also regulate metabolic pathways [43,44]. In this study, we observed a negative correlation between cell proliferation rate and overall reprogramming efficiency for NHDF-PK242 in accordance with a previous report [36]. However, we did not observe such correlation for NHDF-PK236, which indicates the complexity of multifactorial determination of overall reprogramming efficiency. Similar to cell proliferation rate, the negative correlation between reprogramming efficiency and SRC is clearer for these fibroblast lines with extreme high or low SRC levels, which again confirm the complex correlation between mitochondrial function and nuclear reprogramming [36]. Thus, it is likely that such an SRC range exists that SRC is not the major determinant of reprogramming efficiency. This might explain the correlation between SRC and reprogramming efficiency for NHDF-1 and NHDF-2 (Fig. 1 and Supplementary Fig. S2).
Our findings raise the question whether treatment of mitochondria targeting chemical molecules that modulate mitochondrial SRC could influence reprogramming efficiency. In our pilot experiment (Supplementary Fig. S4), we observed that treatment of fibroblasts with 2DG increased mitochondrial SRC and decreased reprogramming efficiency. However, since we also observed decreased cell proliferation by 2DG treatment, future studies are still required to systematically investigate the effects of other mitochondria-targeting drugs, their effects on SRC, and reprogramming efficiency, such as whether SRC-lowering drugs could increase reprogramming efficiency. In addition, our observation of the great difference of SRC levels between somatic cells and iPSC cells suggests that the mitochondrial SRC has undergone resetting from a somatic state to a pluripotent state during reprogramming. Although not investigated in this study, it is still unclear about the dynamic changes of mitochondrial SRC during the reprogramming process, which should be investigated in future studies. Nevertheless, our study collectively suggests a potential novel strategy for improving reprogramming efficiency by modulating mitochondrial SRC in somatic cells.
In this study, we observed that fibroblasts, which have been subjected to genetic modification in vitro have increased SRC and lower reprogramming efficiency. Our study also showed that iPSCs and human ESCs, characterized by their pluripotency and self-renewal capacity, have very low or even negative mitochondrial SRC level, implicating the correlation between SRC and cell senescence. Since we have consistently observed increased mitochondrial SRC and impaired reprogramming for six clonal fibroblast lines edited by two independent CRISPRs, the impaired reprogramming efficiency is unlikely to be directly related to gene editing or the potential off-target effects of CRISPR. Rather, one possible cause of the impaired reprogramming efficiency in gene-edited fibroblasts might be related to the long-term in vitro cultivation. Another study also showed that late passage fibroblasts produced fewer reprogrammed colonies [7]. Furthermore, a recent study demonstrated that SRC increases with age in control fibroblasts [45].
An early study discovered that mitochondrial SRC correlates with cell survival and cells with higher SRC are more resistant to cell death and generate less reactive oxygen species (ROS) [22]. The generation of gene-edited fibroblasts has been a process of selecting cells with higher cell survival. This explains why the SRC levels were significantly increased in the gene-edited clonal cells.
In addition, SRC reflects cellular levels of AMPK/mitochondrial mass [22]. During energy or oxidative crises (oligomycin/FCCP), AMPK becomes activated by AMP/ROS and increases mitochondrial acetyl-CoA supply and oxygen consumption [46]. C-MYC, which we overexpress during reprogramming to induce the Warburg effect and cell proliferation, has opposing effects on metabolism as compared with AMPK (AMPK is catabolic and C-MYC is mostly anabolic). AMPK also decreases C-MYC activity indirectly through inhibition of mTOR. It is, therefore, likely that high cellular levels of AMPK may override C-MYC expression and suppress the Warburg effect and reprogramming. In that way, cellular levels of AMPK/SRC determine cellular reprogramming capacity during C-MYC overexpression. A recent study showed that C-MYC cooperates with AMPK to inhibit ATP depletion and oxidative stress during highly proliferative conditions [47]. As such, a fine-tuned balance between C-MYC and AMPK activities is needed to support reprogramming, and AMPK/SRC levels could be used to screen for reprogramming efficiencies in cells. The fact that the correlation between SRC and reprogramming efficiency is strongest at extreme low SRC and high SRC may also reflect the fine-tuned balance between AMPK and C-MYC activities during reprogramming; too low AMPK cannot support C-MYC function during proliferation, and too high AMPK may override C-MYC function. Also, long-term cultivation increases ROS and depletes ATP, which may induce AMPK/SRC to a level sufficient to override the C-MYC/Warburg effect and thereby decrease reprogramming efficiency. Consequently, drugs to support ATP production and decrease ROS may be useful to increase reprogramming efficiency in the gene-edited cells.
In conclusion, we found that the SRC correlates negatively with reprogramming efficiency. Interestingly, another group [25], previously performed mitochondrial stress test on fibroblasts and their corresponding pluripotent cells. It was obvious that pluripotent cells responded less to FCCP treatment compared with their corresponding fibroblasts. Indeed, our fibroblast cells with the highest reprogramming efficiency had extremely low mitochondrial SRC in a manner similar to iPSCs and ESCs, with the authentic ESCs having the lowest SRC. Our results showed that fibroblast cells with extremely reduced SRC had the highest efficiency of iPSC generation. The present study suggests the possibility for predicting the reprogramming efficiency by measuring the mitochondrial SRC. This approach can be utilized to optimize reprogramming strategies to be more effective and less time consuming. Furthermore, based on the observation that 2DG affects both mitochondrial SRC and reprogramming efficiency, our study suggests that modulating SRC by mitochondria-targeting drugs could be an alternative strategy to improve reprogramming efficiency of somatic cells in the future.
Footnotes
Acknowledgments
This project is supported by grants from the Danish Council for Independent Research—DFF-1337-00128 [Y.L.], DFF-1335-00763 [Y.L.], DFF-4004-00548 [R.K.J.O], Lundbeckfonden (Lundbeck Foundation)—R173-2014-1105 [Y.L.], R151-2013-14439 [L.B.], R126-2012-12448p [P.B.], R126-2012-12456 [L.A.], and the BrainStem project from Innovation Fund Denmark. Y.Z. and R.A. S. are supported by PhD scholarship from the Faculty of Health at the Aarhus University. Y.Z. is also supported by the China Scholarship Council. The authors thank the FACS Core facility of the Aarhus University for the technical help with all flow cytometry analyses.
Author Disclosure Statement
The authors have no conflicts of interests to declare.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.