
Abstract
Ischemic stroke represents the leading cause of adult neurological disability, with no effective therapeutic strategy. Stem cell transplantation promises a new promising for treating stroke, through cell replacement and cytokine paracrine. However, due to the effect of hostile immune microenvironment, the survival and differentiation of stem cells are limited in vivo. Furthermore, the delayed inflammatory response to stroke induced secondary neurological injury. IFN-γ as pro-inflammatory cytokine has the potential to protect stem cell population during inflammatory response, as well as stimulates neurogenesis of stem cells. The purpose of this study was to investigate whether co-injection of neural stem cells and IFN-γ can improve therapeutic outcomes in ischemic stroke model. In this study, we found that IFN-γ did not interfere with the proliferation of neural stem cells (NSCs) in vitro and induced levels of subsequent neuronal differentiation significantly superior to those of other four cytokines BDNF, VEGF, TGF-β, and IGF-1. Co-delivery of IFN-γ (concentration: 50 ng) enhanced the effectiveness of NSC transplantation therapy in ischemic rats. And combined IFN-γ treatment significantly increased neurogenesis in vivo, with more BrdU/DCX dual-positive cells found in ischemic areas. Moreover, co-treatment with IFN-γ and NSCs exerted additional neurological benefits compared with NSC transplantation alone. In conclusion, low concentration of IFN-γ can promote the functions of transplanted NSCs and facilitate their ability of neurological repair. Thus, our findings suggest that co-delivery of NSCs and IFN-γ without genetic modification may be an effective, simple, and novel approach for the treatment of ischemic stroke.
Introduction
I
Previous studies have revealed that neurogenesis and angiogenesis occur following neuronal injury, suggesting that endogenous recovery processes are initiated following stroke [4]. However, as these effects decrease over time, the capacity for endogenous recovery may be limited [5]. Stem cell-based approaches represent promising treatments for ischemic stroke and allow for a more dynamic biological response to the microenvironment [6]. Several experimental studies have demonstrated that transplantation of neural stem cells (NSCs) improves neurological function following ischemic stroke by increasing neurogenesis, regulating the secretion of trophic factors in the brain, mediating neuroprotection, modulating immune function, and stimulating angiogenesis [7,8]. Recent clinical trials involving patients with stroke have also reported improvements in neurological function following stem cell-based therapy and that such treatment was associated with no adverse effects [9,10].
Although transplantation of NSCs can ameliorate ischemic brain injury, the therapeutic benefits of transplantation are limited by poor survival of the grafted cells due to the hostile ischemic microenvironment, which may be caused by reactive oxidative stress, inflammation, and so on. [11 –13]. In addition, previous studies have revealed that grafted stem cells tend to differentiate into neuroglia rather than functional neurons [7,8,12,13]. Various stem cell gene modification and co-transplantation approaches have been investigated to enhance the efficiency of transplantation (eg, overexpression of pro-survival signaling molecules or paracrine factors of interest) [7,14]. However, even with such approaches, the transplanted stem cells exhibit relatively low rates of neuronal differentiation. Thus, a simpler and more effective approach for inducing neuronal differentiation in grafted stem cells in vivo is required.
Interferon-gamma (IFN-γ) is a mediator of the pro-inflammatory pathway and plays an important role in the postischemic brain [15]. IFN-γ not only contributes to the maintenance and initiation of the immune response in the post-ischemic brain but also exerts a significant effect on the differentiation of stem cells [16,17]. Under differentiating conditions, physiological concentrations of IFN-γ significantly enhance neuronal differentiation [18]. Moreover, IFN-γ protects NSCs against damage due to central nervous system (CNS) infection through immune-related bystander effects [17]. Interestingly, IFN-γ binds to extracellular vesicles (EVs) through Ifngr1 (IFN-γ receptor 1), which activates Stat1 in target cells [19]. This finding suggests that IFN-γ mediates communication between the grafted stem cells and the immune system of the host through EV-associated IFN-γ/Ifngr1 complexes [19]. Therefore, the present study aimed to determine whether co-delivery of IFN-γ and NSCs induces reprogramming of NSCs and attenuates damage following ischemic stroke, thereby enhancing the efficacy of stem cell transplantation therapy. We further aimed to elucidate the potential mechanisms underlying the effects of combined IFN-γ and NSC treatment.
Materials and Methods
Animals and NSC isolation cultivation
All animal procedures were conducted in accordance with the Care and Use of Laboratory Animals protocol approved by the Institutional Animal Care Committee of Zhongda Hospital, Southeast University. All Sprague-Dawley (SD) rats were purchased from Vital River Laboratory Animal Technology Co., Ltd. (China), maintained on a 12-h light/12-h dark cycle and given access to food and water ad libitum. Primary NSCs were extracted from the hippocampus of fetal rats (from pregnant 14 days female rat) according to established protocol. Cells were collected, and resuspension was done with complete medium (DMEM/F12 basal medium with 20 ng/mL epidermal growth factor, 20 ng/mL basic fibroblast growth factor, 2% B27, 2% N2, 1% Penicillin-Streptomycin, 2 nM Glutamate, and 2 μg/mL Heparin sodium) in the culture flask and put into 37°C, 5% CO2 incubator. Fresh medium was changed every 3 days and passaged at the 7th day. Then single cells were dissociated from neurospheres with Accutase Dissociation Reagent (Sigma) and gave rise to new neurospheres.
NSC proliferation identification and differentiation
To evaluate the proliferation of NSCs, cells were incubated in the flask or plates with complete medium. For examining the neural stem cell differentiation, stem cells were dissociated with Accutase and then plated at 1 × 105 cells per 24 well or 5 × 105 per six well coated with poly-D-lysine polymers (PDL; Sigma) in differentiation medium (DMEM/F12 supplemented with 2% FBS and 1% Penicillin-Streptomycin) for 7 days. At the appropriate time points, cells were fixed in 4% paraformaldehyde, after being washed in PBS followed by Triton X-100 (sigma). Then camber slides were blocked in 10% donkey serum solution (Jackson). Cells were then incubated with primary antibodies: rabbit anti-nestin, mouse anti-beta III-tubulin (Tuj1), goat anti Myelin oligodendrocyte glycoprotein (MOG) (1:1,000; Abcam), rabbit anti GFAP (1:100; Bioss), and goat anti Ki67 (1:200; Santa Cruz) overnight. Then the secondary antibodies donkey anti-rabbit-Alexa Fluor® 647, anti-goat-Alexa Fluor 488, and anti-mouse-Alexa Fluor 488 (1:800; Abcam) or donkey anti-mouse-Alexa Fluor 488 (1:800; Jackson) were added; after the nuclear marker DAPI adding, the slides were cover slipped for capturing on fluorescence microscope.
Cell viability/proliferation test (CCK-8 assay)
NSCs were seeded in 96-well plates for cell viability analysis. The cells were cultured with fresh medium containing 20 ng/mL concentration of interferon γ (IFN-γ), brain-derived neurotrophic factor (BDNF), vascular endothelial growth factor (VEGF), transforming growth factor-β (TGF-β), and insulin-like growth factor 1 (IGF-1) stabilized for 3 or 5 days and then CCK-8 (KeyGen, China) was added into each pore. After incubation, absorbance value of each pore was colorimetrically determined. Proliferation rate = (OD450 of cytokine group − OD450 of control group)/OD450 of control group × 100%.
Quantitative RT-PCR
Total RNA from NSC preconditioning by IFN-γ was isolated using TRIzol reagent (Invitrogen). RNA was subsequently treated with RNase-free DNase I (Omega). Synthesis of cDNA was done using the Transcriptor First Strand cDNA Synthesis Kit from TIANGEN (China) according to the manufacturer's instructions. Quantitative RT-PCR was carried out using ABI Detection System with FastStart Universal SYBR Green Master of Roche. The PCR primers are Ascl1, Forward primer, 5′-CCCACCATCTCCCCCAACTA-3′, reverse primer, 5′-CTGTCAGCCCCTCAGAACCA-3′, Nestin, Forward primer, 5′-GGGGAATCTGATGGGTTTGC-3′, reverse primer, 5′-GTACCGGGTCTCCTCTTTGGG-3′, Stat1, forward primer, 5′-CCTGTGGTACAACATGCTGGTG-3′, reverse primer, 5′-TTGGTGACTGACGAAAACTGCC-3′, GAPDH, forward primer, 5′-TTCCTACCCCCAATGTATCCG-3′, reverse primer, 5′-CATGAGGTCCACCACCCTGTT-3′.
Western blot analysis
To analyze protein levels, equal amounts of total proteins were subjected to SDS-PAGE and transferred onto polyvinylidene difluoride membranes (PVDF; Millipore). Membranes were then blocked with milk and probed with IFN-γ, (1:400; Santa Cruz), Ascl1, Tuj1 (1:1,000; Abcam), p-Jak1, p-Stat1 (1:1,000; CST), and PCNA, GAPDH (1:400, 1:2,500; Bioss) primary antibodies. Subsequently, membranes were incubated with horseradish peroxidase-conjugated secondary antibodies for 1 h and then reacted with an enhanced ECL substrate (Pierce).
Transient middle cerebral artery occlusion
Adult male SD rats (240–280 g) were subjected to transient focal cerebral ischemia by intraluminal middle cerebral artery blockade with a suture method. Briefly, rats were anesthetized with chloral hydrate. Blunt dissection was performed to isolate the common carotid artery (CCA), the internal carotid artery (ICA), and the external carotid artery. Then nylon thread was introduced into the CCA, fed distally into the ICA, and advanced to the origin of the MCA. The insertion depth was approximately 18 ± 0.5 mm above the CCA bifurcation. The nylon thread was then withdrawn after 90 min ischemia. The incision was then sutured closed, and the rat was allowed to wake in a fresh cage. The rectal temperature was controlled at 37°C with a homeothermic blanket during surgery.
Intracerebral administration of NSCs and/or recombinant IFN-γ
Rats were treated with chloral hydrate as anesthetic and then placed securely in stereotaxic apparatus. The NSCs and/or IFN-γ were then stereotactically transplanted into the striatum of the infarcted hemisphere 3 days after the stroke onset. The transplantation site was positioned along the anterior–posterior axis into the cortex: 0.5 mm posterior to the coronal suture, 3 mm lateral to the sagittal suture, and 4.5 mm deep from the dura. Deposits were delivered at 2 μL/min, and the syringe was kept in place for additional 5 min after injection before removal. Rats were randomly divided into five groups, including sham operation group, PBS group (10 μL PBS), IFN-γ group (50 ng in 10 μL PBS), NSC group (8 × 105 cells in 10 μL PBS), and NSCs combined IFN-γ group (8 × 105 cells and 50 ng IFN-γ in 10 μL PBS), n = 15 rats/group. 5-Bromo-2′-deoxyuridine (Brdu, 50 mg/kg, twice every day; Sigma) was intraperitoneally administered into rats after transplantation for 14 consecutive days.
Behavioral test
Neurobehavioral tests were performed before middle cerebral artery occlusion (MCAO) and 1, 3, 7, 14, and 28 days after MCAO and transplantation by two investigators who were blinded to the experimental groups. Modified neurologic severity scores (mNSS) of the rats (n = 6–15 rats/group) were graded on a scale of 0 to 18, which is a composite of motor, reflex, and balance tests. The higher the score the more severe injury. With the rotarod test (n = 6–15 rats/group), after 3 days of training before stroke, the mice were placed on the rotarod cylinder, and the time the animals remained on the rotarod was recorded. The speed was slowly increased from 4 to 40 rpm within a period of 5 min. The trial was ended if the animal fell off the rungs or gripped the device and spun around for two consecutive revolutions. Data were analyzed as the average duration of three trials on the rotarod.
Infarct volume analysis
Rats' brains (n = 5 rats/group) were removed on the 28th day after MCAO and transplantation, sectioned into six equidistant slices, and incubated with 2% 2,3,5-triphenyltetrazoliumchloride (TTC; Sangon Biotech) solution to visualize the infarct area.
Double immunofluorescence
The brain sections were dried at 37°C, after being washed followed by Triton X-100 (Sigma). Then camber slides were blocked in donkey serum solution (Jackson). Cells or sections were then incubated with primary antibodies: goat anti-Brdu (1:100; Santa Cruz), rabbit anti-nestin and rabbit anti DCX (1:1,000; Abcam), and rabbit anti GFAP (1:100; Bioss) diluted in blocking solution. The second day, the secondary antibodies donkey anti-rabbit-Alexa Fluor 647/488 and anti-goat-Alexa Fluor 488/555 (1:800; Abcam) were added, followed by DAPI adding. After washing, the slides were cover slipped for capturing on fluorescence microscope.
TUNEL assay
Apoptotic cells were detected with TUNEL staining according to the manufacturer's instructions (Roche). Cells with yellow-brown granules in the nucleus were regarded as apoptotic cells.
Immunohistochemical analysis
Briefly, the brain slices were washed with double distilled water and then put into the Nissl staining solution for about 15 min. The slices were then washed with double distilled water again, followed by dehydrating with 95% ethyl alcohol for 5 s. Then, the slices were dehydrated twice with 75% ethyl alcohol for 1 s. The slices were finally put into the xylol twice for 3 min before observation. Then for apoptotic assay, after blocking with BSA (Sigma), brain sections were incubated with TUNEL staining according to the manufacturer's instructions, with slight modification. Cells with yellow-brown granules in the nucleus were regarded as apoptotic cells.
Statistical analyses
Values are expressed as the mean ± standard deviation (SD). Statistical analyses were performed using t-tests, one-way or two-way ANOVA, followed by Mann–Whitney test, Welch's correction, nonparametric Kruskal–Wallis test, and Bonferroni post-tests using GraphPad 5.0 Software. The significance of the differences between different groups was tested by variance analysis (with P < 0.05 as significant and P < 0.01 as very significant).
Results
IFN-γ does not affect NSC proliferation but promotes neuronal differentiation in vitro
Primary NSCs isolated from the hippocampal tissue of fetal rats formed nonadherent neurospheres in vitro (Fig. 1A), and these neurospheres significantly expressed the NSC-specific marker Nestin (Fig. 1B). To induce cellular differentiation, neurospheres were cultured in differentiation medium (DMEM/F12 supplemented with 2% FBS) for 7 days after the withdrawal of growth factors. Cells began to migrate away from neurospheres, and staining with Tuj1, GFAP, and MOG revealed that the cells had differentiated into neurons, astrocytes, and oligodendrocytes, respectively (Fig. 1C). Staining results thus confirmed the identity of neural stem cells.
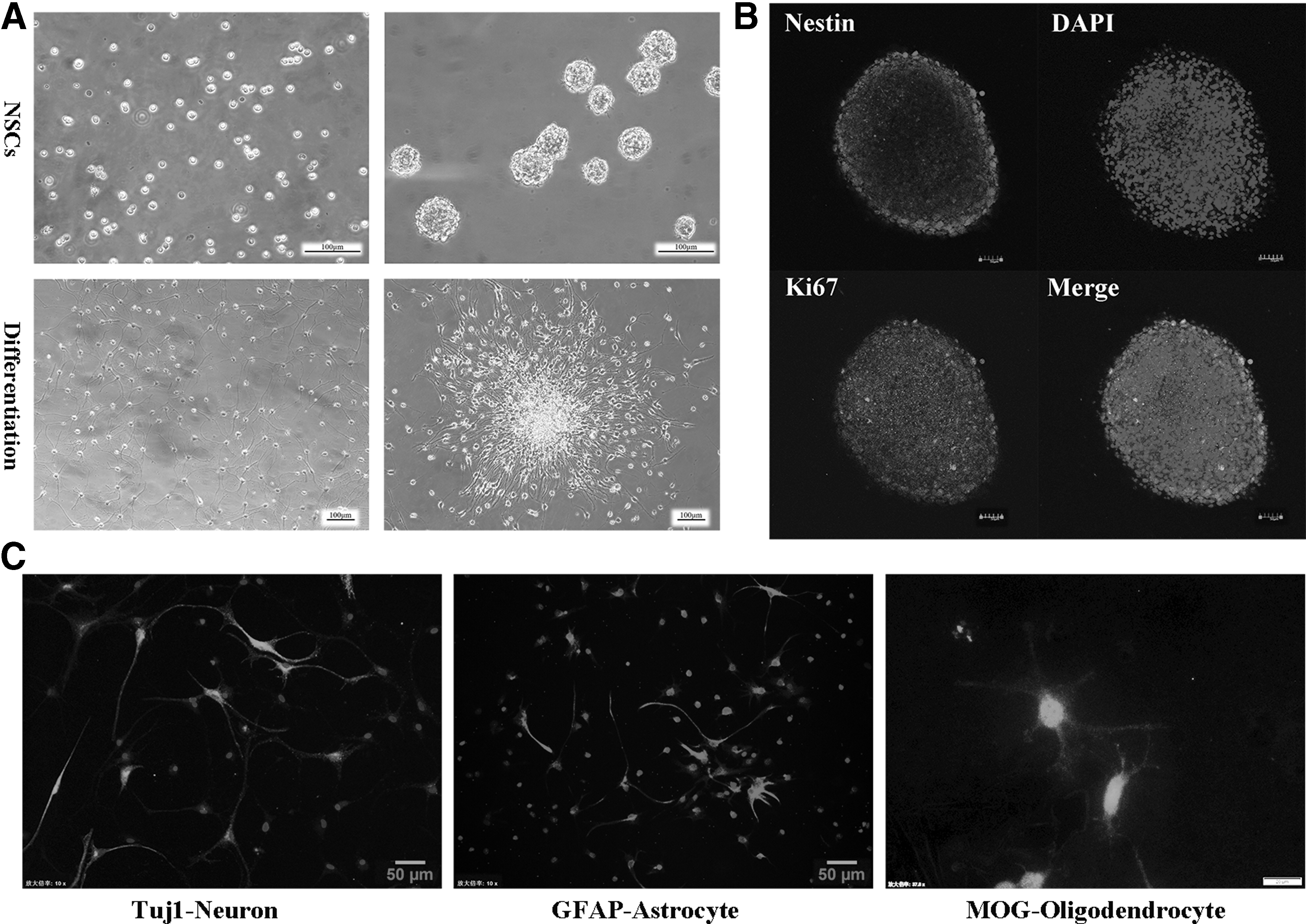
The characteristics and differentiation of hippocampal-derived neural stem cells obtained from fetal rats. Upper right
To examine whether IFN-γ exerted effects on stem cell proliferation and the generation of new neurospheres, we compared the effects of IFN-γ treatment with those of four other cytokines (BDNF, VEGF, TGF-β, and IGF-1; concentration: 20 ng/mL each). Each treatment was applied to dissociate NSCs, which were subsequently allowed to proliferate for 5 days. Our results (Fig. 2A) indicated that none of the five cytokines exerted adverse effects on cell viability. Although all treatments resulted in proliferation and neurosphere formation, the neurospheres formed following BDNF and VEGF treatment were larger than those observed following the remaining treatments, and interestingly, IFN-γ treatment promoted the generation of branches from NSC neurospheres. It suggested that IFN-γ exerts no obvious cytotoxic effects on NSCs.
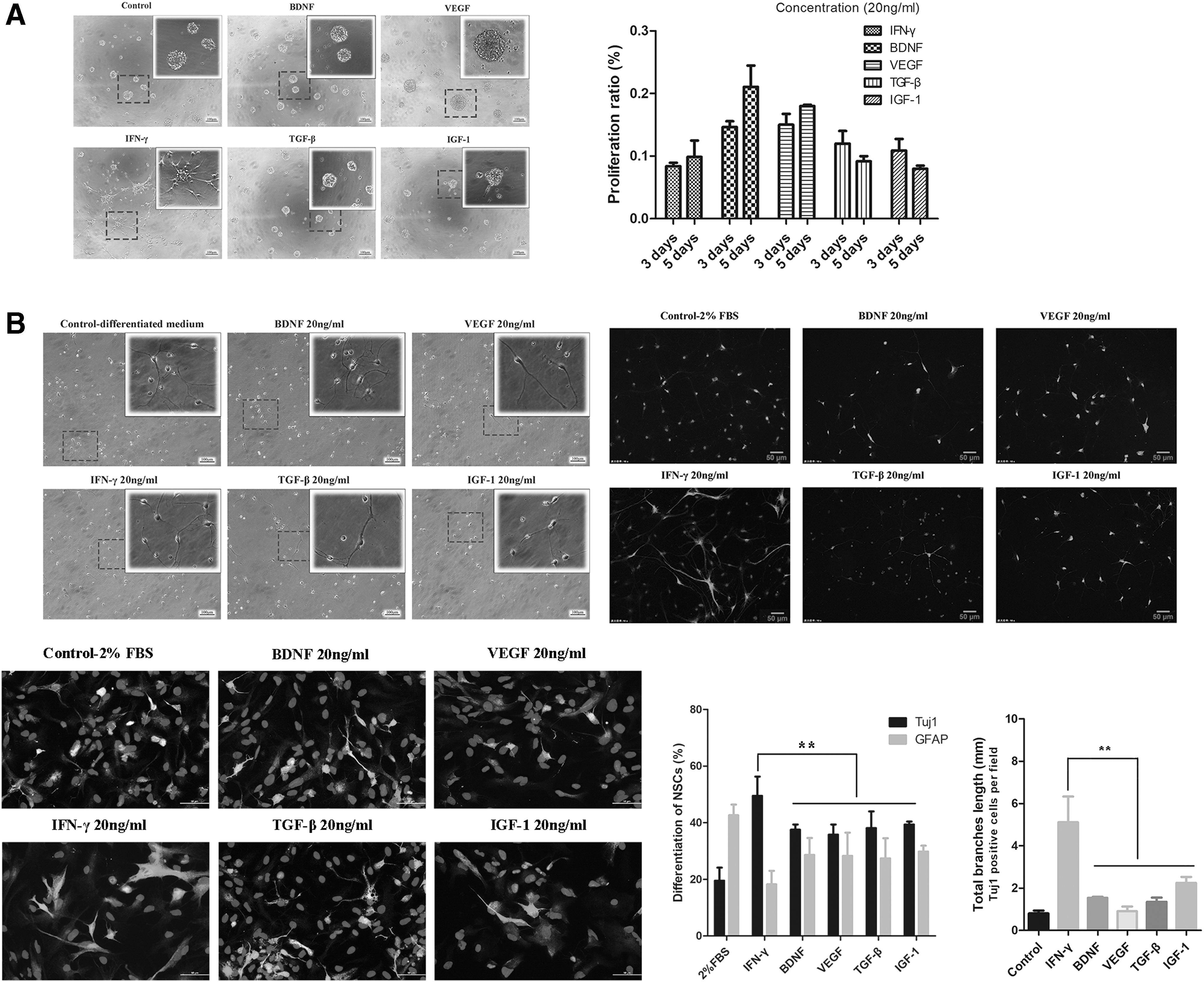
IFN-γ did not affect NSC proliferation but promoted neuronal differentiation in vitro through the Stat1 pathway.
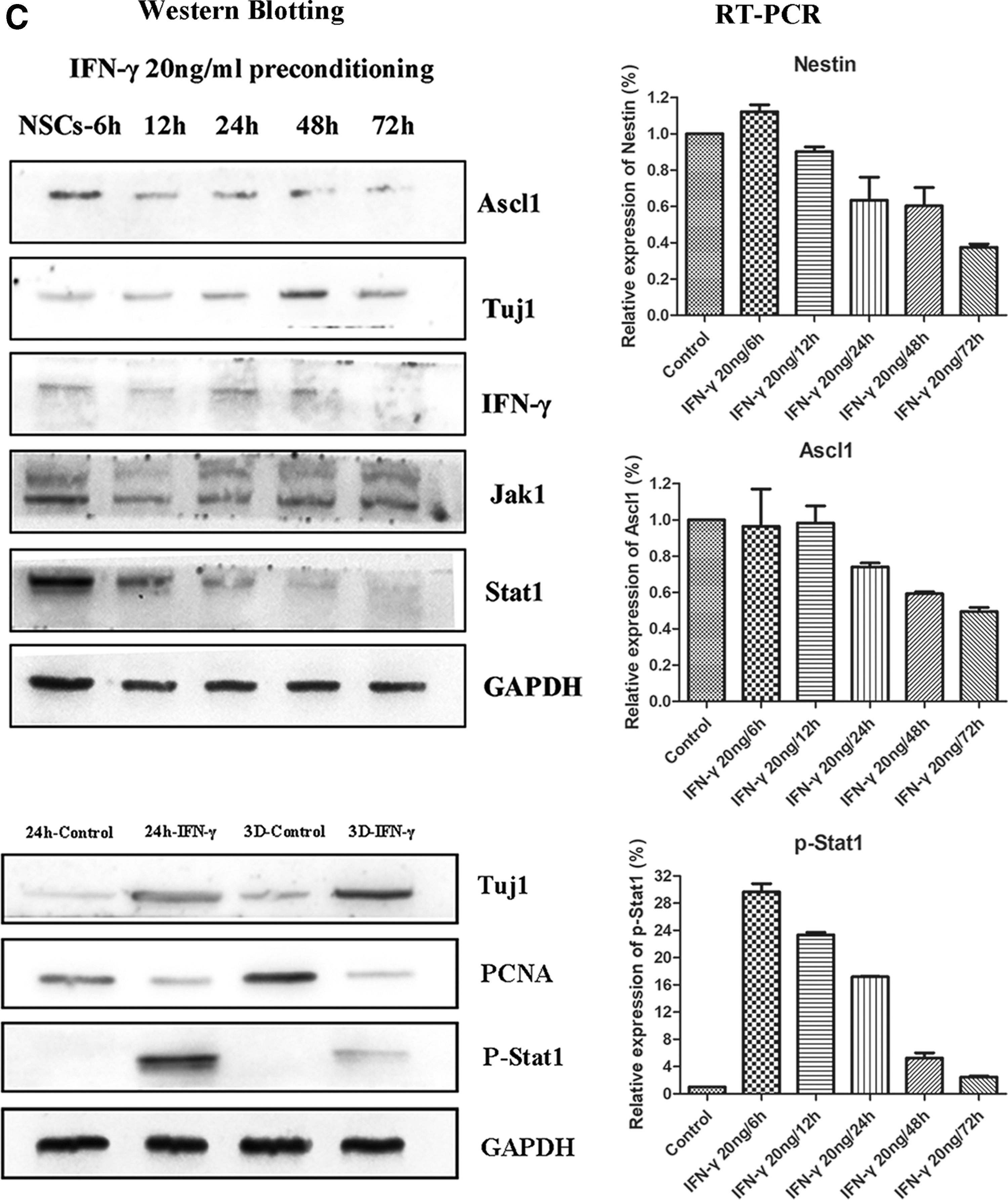
We then aimed to determine whether these cytokines exerted effects on NSC differentiation. Five cytokines, including IFN-γ, were added to the differentiation medium in which NSCs were cultured. While none of the five cytokines influenced the differentiation of NSCs, IFN-γ treatment significantly increased the ratio of Tuj1-positive neurons and the total branch length of Tuj1-positive cells on day 3, relative to levels observed in controls or following treatment with any other cytokines (Fig. 2B, P < 0.01). We further analyzed the different protein expressions of NSCs at different time points following IFN-γ culturing. Western blotting experiments revealed that IFN-γ stimulation downregulated the expression of PCNA, a specific marker of cell proliferation (Fig. 2C). However, IFN-γ significantly increased the generation of neurons (Tuj1 expression) from 6 to 72 h following application. This was primarily reflected in an early increase in neuronal numbers after IFN-γ preconditioning.
IFN-γ activates the p-Stat1 signaling pathway during NSC differentiation
To further identify the roles of IFN-γ during NSC differentiation, we assessed the downstream signaling pathway of IFN-γ in vitro using NSC cultures. We examined the expression of phosphorylated Jak1 and Stat1 at different time points following IFN-γ preconditioning. Western blotting (Fig. 2C) experiments revealed that Jak1 and Stat1 were activated by IFN-γ beginning at 6 h following application. Expression of Jak1 was sustained until 72 h, although Stat1 expression gradually decreased following the 6-h mark. QRT-PCR (Fig. 2C) results verified that Stat1 activation reached a peak at 6 h, following which gradual decreases were again observed. Similar trends of expression were observed for Nestin and Ascl1 (markers of NSCs and activated NSCs, respectively). However, there was no further change in the expression of Jak1. These findings suggest that IFN-γ induces the differentiation of NSCs through activation of the Stat1 pathway.
Co-administration of IFN-γ and NSCs exerts synergistic effects on poststroke neurological impairments in vivo
We then co-injected IFN-γ and NSCs into a rat model of ischemic stroke. Neurological performance was evaluated from days 1 through 28 based on mNSS and the results of Rotarod tests. The NSC and co-transplantation groups exhibited significant improvements in mNSS on days 14 to 28, relative to those observed in the PBS control group. Moreover, greater improvements in functional outcomes were observed in the co-injection group than in rats treated with NSC alone (Fig. 3A, n = 11 rats/group, P < 0.05). Significant functional improvement in Rotarod performance was observed from days 7 to 28 in the transplantation group, relative to that observed in the PBS control group. And improvements were again greater in the co-administration group than in rats treated with NSC alone from days 7 to 28 (Fig. 3B, n = 11 rats/group, P < 0.05). Although our findings indicated that intracerebral transplantation of NSCs alone can alleviate functional impairments, co-treatment with NSCs and IFN-γ further reduced functional deficits. Therefore, co-injection of IFN-γ and NSCs may exert synergistic effects in vivo, leading to sustained improvements in functional outcomes.
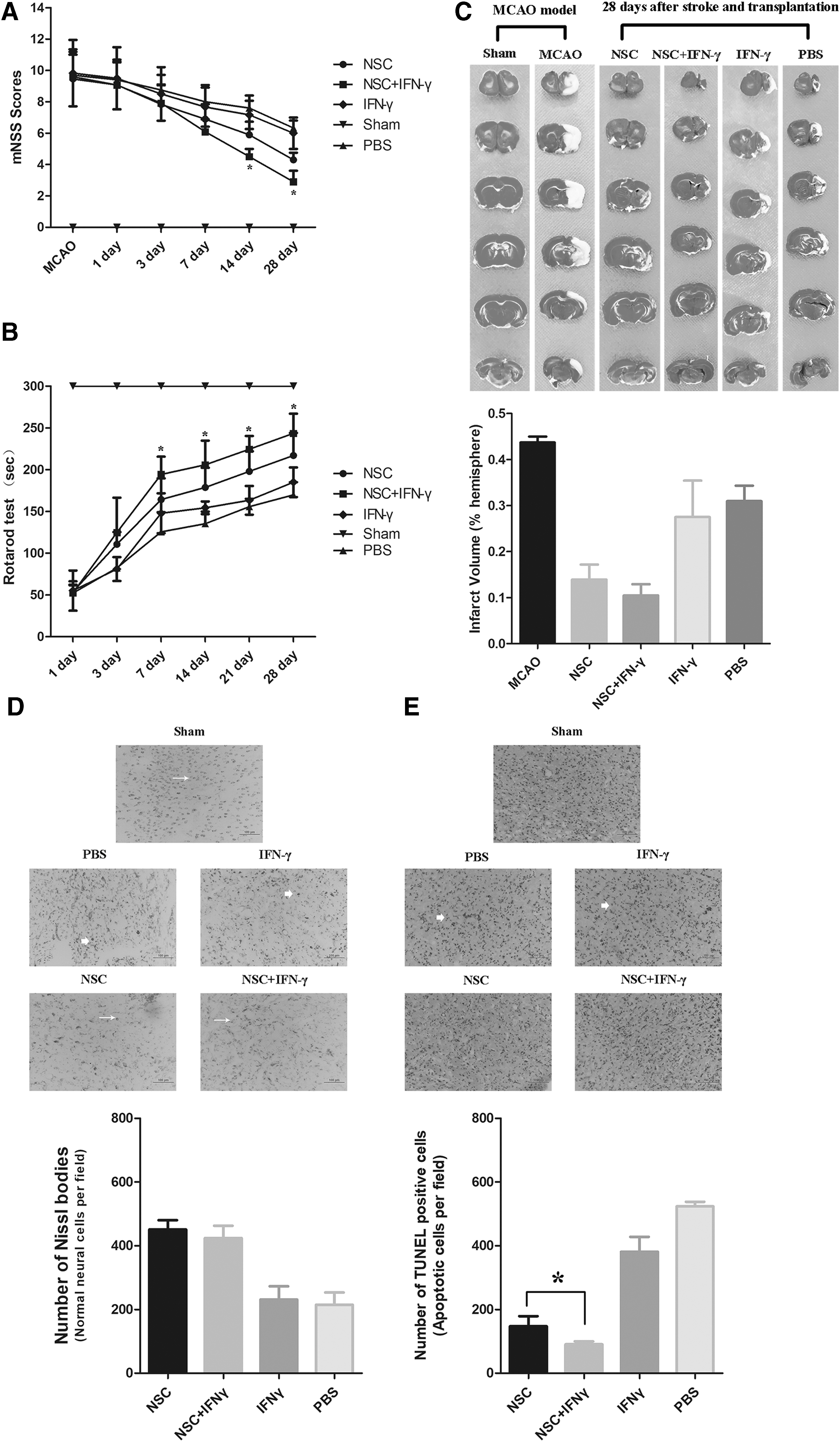
Co-administration of IFN-γ and NSCs results in greater functional recovery and lesser neuronal apoptosis of poststroke rats. The mNSS
Co-treatment with IFN-γ and NSCs does not reduce infarct size compared to NSCs alone
Our findings indicated that combined treatment with IFN-γ and NSCs exerts synergistic effects on neurological recovery in a rat model of ischemic stroke. We then investigated whether co-transplantation of IFN-γ and NSCs reduces infarct volume or structural damage following ischemic stroke. Twenty-eight days after stroke and transplantation, TTC staining revealed that infarct size had significantly decreased by approximately 65.8% and 54.8% in the co-transplantation and NSC-only groups, respectively (Fig. 3C). Although this reduction in infarct size in the co-transplantation group was also more than in the NSC-alone group, it had no statistic difference (n = 5 rats/group, P > 0.05). Thus, these findings suggest that intracerebral injection of IFN-γ just enhances the neurological functions of NSC transplantation in vivo.
Co-delivery of IFN-γ and NSCs reduces the death of neural cells and increases cell survival
Cerebral ischemia usually results in large areas of neuronal necrosis or damage at the site of ischemia. Thus, we investigated whether co-injection of IFN-γ and NSCs influenced the survival of grafted cells in vivo. NSC was injected into the ischemic penumbra with or without IFN-γ 3 days after stroke onset, and cell death in the ischemic regions was assessed 3 days after transplantation. The extent of necrosis/damage to neuronal cells was significantly lower in rats treated with NSCs than in those treated with PBS (control); however, no significant difference was observed between the co-transplantation and NSC-only groups (Fig. 3D, n = 5 rats/group, P > 0.05). These findings suggest that transplantation of exogenous NSC attenuates the degeneration and necrosis of ischemic cells in rats with brain ischemia.
We then examined whether co-injection of IFN-γ and NSCs influences neuronal apoptosis in rats following ischemia through TUNEL staining. Our results indicated that, 3 days after stroke and transplantation (the same time point with above), the number of TUNEL-positive cells in both the NSC-only and co-transplantation groups was much lower than that in the PBS group. Moreover, the number of TUNEL-positive cells was significantly lower in the co-administration group than in the NSC-only group (Fig. 3E, n = 5 rats/group, P < 0.05). These results indicate that NSC treatment reduced necrosis and apoptosis of neurons in ischemic rats and that co-injection of IFN-γ may enhance the effects of NSC transplantation on neuronal apoptosis.
We further examined the expression of BrdU/Nestin in double-positive cells 3 days after stroke and transplantation. Immunofluorescence staining revealed extensive distribution of the grafted NSCs toward the border of the ischemic lesion in both the co-transplantation and NSC-only groups (Fig. 4A). However, the number of BrdU/Nestin double-positive cells in the co-transplantation group was not less than that in the NSC-only group (Fig. 4A, n = 5 rats/group, P > 0.05). These results indicate that grafted NSCs survived in the ischemic brains and that IFN-γ treatment did not affect the survival of grafted NSCs in vivo.
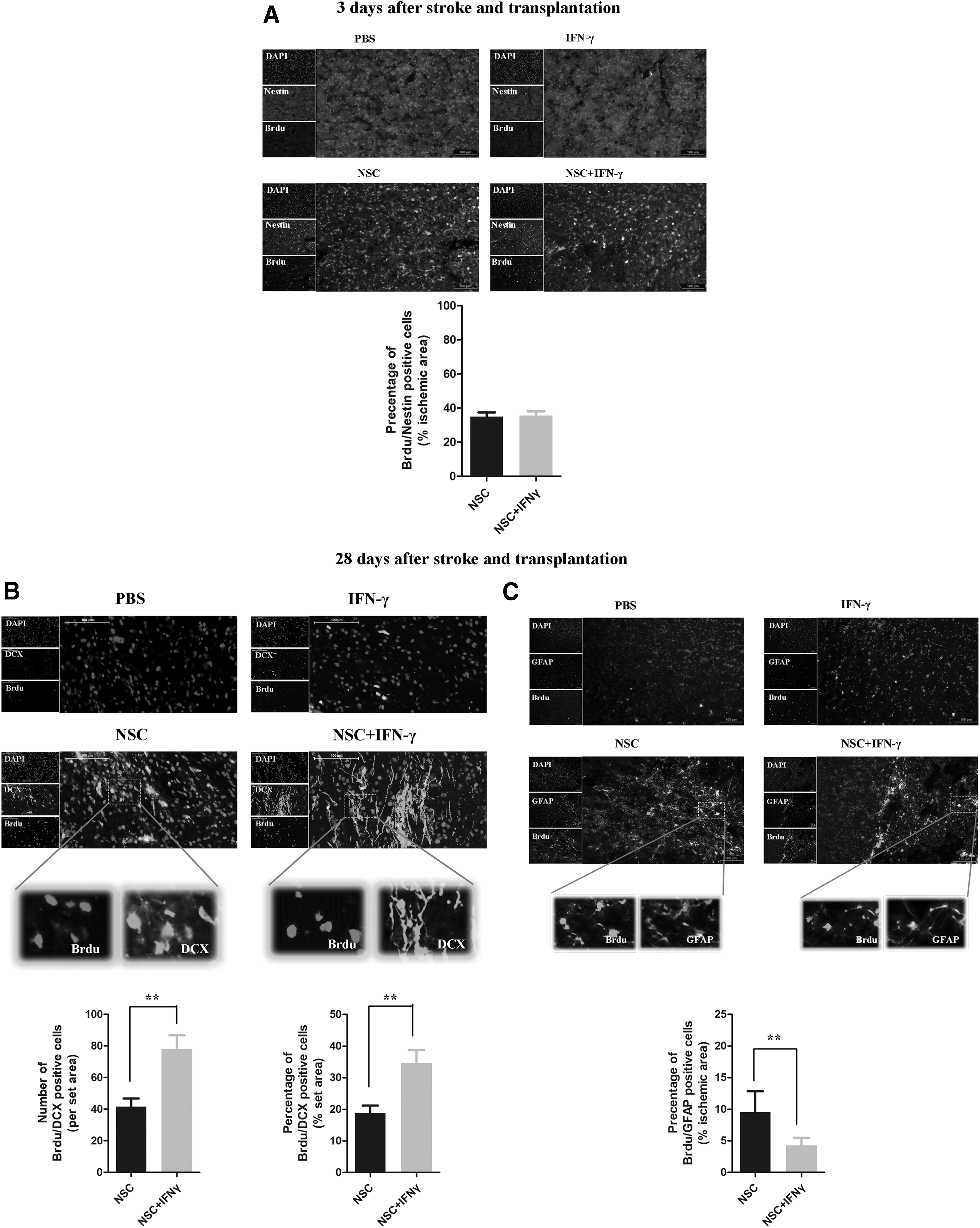
IFN-γ did not interfere with transplanted NSC survival in vivo, and co-administration of NSCs and IFN-γ promoted neural graft survival and differentiation of grafted NSCs.
Neurogenesis is enhanced in rats following co-injection of IFN-γ and NSCs
We then analyzed the differentiation profiles and proliferation capacity of the grafted NSCs in ischemic regions. Double-immunofluorescent staining for the proliferation marker BrdU and lineage-specific phenotype markers demonstrated that the grafted NSCs differentiated into neurons and astrocytes 28 days after stroke and transplantation (Fig. 4B). A greater percentage of neurons and astrocytes was detected in rats treated with NSCs. Notably, intracerebral injection of IFN-γ combined with NSC transplantation resulted in a sustained postischemic increase in the number or percentage of BrdU/DCX double-positive cells (Fig. 4B, n = 5 rats/group, P < 0.01) relative to that observed in the NSC-only group, as well as a decrease in the percentage of BrdU/GFAP dual-positive cells (Fig. 4C, n = 5 rats/group, P < 0.01). These findings indicate that co-transplantation of IFN-γ and NSCs enhances the expression of BrdU positive cells co-localized with the neuronal marker DCX in ischemic regions. Thus, our results suggest that co-administration of IFN-γ significantly stimulates grafted NSCs into neuronal differentiation. No tumor genesis was observed following NSC grafting.
IFN-γ/Stat1 signaling pathway regulated the activation and neurogenesis of NSCs in vivo
We then investigated the expression of IFN-γ protein and related signaling molecules in the ischemic brain following NSC transplantation with or without co-injection of IFN-γ through western blotting. As shown in Fig. 5, IFN-γ levels were upregulated in the IFN-γ-only and co-transplantation groups, relative to levels observed in controls. Furthermore, the expression of p-Stat1 was upregulated in the co-injection group, indicating that the IFN-γ/Stat1 signaling pathway was activated by co-injection of IFN-γ in vivo.
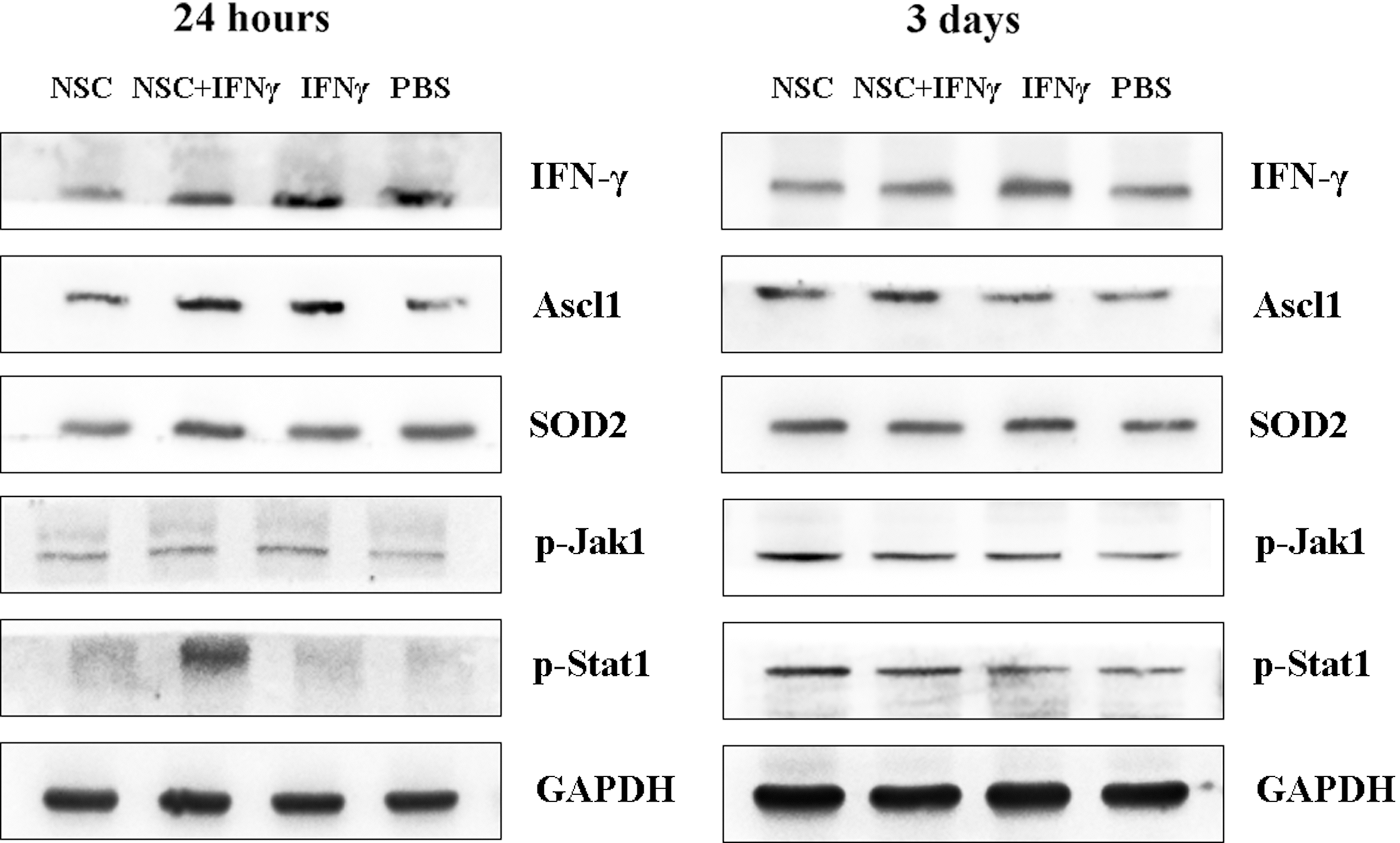
Co-delivery of IFN-γ activated the Stat1 signaling pathway in vivo. Western blotting results indicated that the protein levels of IFN-γ and its downstream signaling target p-Stat1 were upregulated in the co-transplantation group (NSCs and IFN-γ) at 24 h and 3 days after transplantation. Furthermore, the expression of Ascl1 and SOD2 was also increased in the co-transplantation group. GAPDH was used as the loading control.
Discussion
The present study aimed to determine whether co-treatment with IFN-γ and NSCs improves therapeutic functional outcomes in a rat model of ischemic stroke. Our findings indicated that co-delivery with IFN-γ enhanced the effectiveness of NSC transplantation therapy in ischemic rats and that co-treatment significantly increased neurogenesis and enhanced functional recovery in vivo.
Previously, we summarized the effect sizes of NSC therapy in experimental models of ischemic stroke [7]. Our meta-analysis and systematic review indicated that NSC transplantation significantly improves both structural and functional outcomes and that the extent of such improvements is associated with the time of NSC administration and the source of NSCs (immunogenicity) [7]. Typically, neuronal differentiation of transplanted NSCs in the CNS is relatively sparse; however, the capacity of the ischemic brain for recovery depends primarily on neurogenesis within the injured regions [20]. Thus, elucidating the differentiation pathways of both endogenous and exogenous NSCs may aid in the development of novel therapeutic strategies for stroke.
Brain ischemia is characterized by a local inflammatory response and the accumulation of inflammatory cells at the ischemic site. Moreover, a delayed immune/inflammatory response has been associated with secondary neural injury following stroke [21,22]. There are studies indicating that pretreatment of NSCs with BDNF can mediate neuroprotection [23,24]; combined treatment with BDNF and NGF increases the number of neurons expressing in vitro NSC cultures [25]. And BDNF pretreatment or gene transfer is known to enhance chemotaxis and adhesion of transplanted NSCs, thereby promoting cell engraftment in vivo and improving functional outcomes following stroke [26 –28]. Another study observed that VEGF exerted neuroprotective effects with regard to the survival of stem cells and neuronal cells [29]; preconditioning of NSC cultures with VEGF also significantly increased the neuronal fraction in vitro [30,31], as well as cytokines TGF-β1 and IGF-1 also can promote the long-term proliferation and maintenance of NSCs in vivo [32 –35]. Although these cytokines (such as BDNF, VEGF, or TGF-β1 and IGF-1) may improve neurological function in the ischemic brain, the extent of improvement is limited, mainly because neuronal generation from transplanted NSCs was a little in vivo.
Recent evidence has indicated that the pro-inflammatory cytokine IFN-γ may protect the NSC population during CNS inflammation and stimulate neurogenesis from NSCs and also suggested that IFN-γ is involved in several endogenous repair processes in the nervous system [16 –19]. In the present study, we observed that IFN-γ did not interfere with the proliferation of NSCs in vitro and induced levels of subsequent neuronal differentiation significantly superior to those of the cytokines BDNF, VEGF, TGF-β, and IGF-1. These findings indicate that IFN-γ may regulate the directional differentiation of grafted NSCs. We further observed that co-administration of IFN-γ and NSCs into rats with ischemic stroke significantly improved neurological function relative to levels observed in rats treated with NSCs alone and IFN-γ alone. But the combination group did not further reduce the infarct volume; may be the number of rats or transplanted NSCs was not enough. Our findings also suggested that IFN-γ did not affect the survival of NSCs in vivo and when co-injected with NSCs significantly increased neurogenesis and the percentage of BrdU/DCX double-positive cells, possibly through the IFN-γ/Stat1 signaling pathway.
The level of IFN-γ is normally elevated following ischemic injury, and altered levels of IFN-γ may affect the outcome of treatment [36]. Although Ahn et al. [37] found that IFN-γ inhibited the neuronal differentiation of mouse neural progenitor cells, our results were consistent with others [38,39], which showed that IFN-γ enhanced neuronal differentiation of NSCs and inhibited astrocyte generation, with no cytotoxicity. Furthermore, it also revealed that neurite and dendrite outgrowth of neurons were strongly associated with the presence of IFN-γ [18,40]. Interestingly, IFN-γ also can promote the communication of NSCs with the microenvironment by binding to EVs. It suggested that uncombined IFN-γ may bind to EVs through EV-associated IFN-γ/Ifngr1 complexes (through Ifngr1 activated Stat1) in target cells, thereby guiding the grafted stem cells to communicate with the host immune system [19].
Conclusions
In summary, our findings support the notion that NSCs represent a promising therapeutic strategy for the treatment of various neurological diseases, including ischemic stroke. IFN-γ not only can produce an inflammatory/immune response at the site of injury but also can promote intercellular molecule exchange between grafted stem cells with host immune cells [19]. The primary implications of the present study are as follows: (1) IFN-γ culturing did not affect the phenotype of NSCs but promoted neuronal differentiation of NSCs through the IFN-γ/Stat1 signaling pathway; (2) IFN-γ co-delivery promoted neuronal generation of grafted NSCs in the ischemic environment; and (3) co-administration of IFN-γ and NSCs into rats with ischemic stroke can accelerate improvements in behavioral performance. However, our study also has some limitations, such as the phenotype of differentiated neurons (inhibitory GABAergic or excitatory glutamatergic), the reconnection of neural circuits, and the relation with EVs (or exosomes) induced by IFN-γ; this critical information will be explored the next time. In summary, our findings suggest that co-delivery of NSCs and IFN-γ without genetic modification may be an effective, simple, and novel approach for the treatment of ischemic stroke.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (grant numbers 81471780 and 81671819) and Jiangsu Science and Technology Project (grant number BL2014076).
Author Disclosure Statement
The authors declare that they have no competing financial interests.