
Abstract
Spinal cord injury (SCI) is a devastating medical condition with profound social and economic impacts. Although research is ongoing, current treatment options are limited and do little to restore functionality. However, recent studies suggest that mesenchymal stem cell-derived exosomes (MSC-exosomes) may hold the key to exciting new treatment options for SCI patients. MSCs are self-renewing multipotent stem cells with multi-directional differentiation and can secrete a large number of exosomes (vesicles secreted into the extracellular environment through endocytosis, called MSC-exosomes). These MSC-exosomes play a critical role in repairing SCI through promoting angiogenesis and axonal growth, regulating inflammation and the immune response, inhibiting apoptosis, and maintaining the integrity of the blood–spinal cord barrier. Furthermore, they can be utilized to transport genetic material or drugs to target cells, and their relatively small size makes them able to permeate the blood–brain barrier. In this review, we summarize recent advances in MSC-exosome themed SCI treatments and cell-free therapies to better understand this newly emerging methodology.
Introduction
In recent years, the number of patients with spinal cord injury (SCI) has increased gradually. Because of the high medical expenses, complex conditions, treatment challenges, various complications, and low cure rate associated with SCI, it imposes a heavy burden on patients, their families, and society, making this condition a major challenge for researchers.
Stem cell therapy as a new strategy for the treatment of SCI has attracted much attention recently [1]. Stem cell therapy promotes the recovery of SCI mainly by replacing lost or damaged cells, providing nutritional support to neurons, protecting neurons, or improving the microenvironment in the spinal cord to allow the regeneration of damaged axons [2]. Mesenchymal stem cells (MSCs) are derived from various sources. Bone marrow, adipose tissue, umbilical cord blood, the placenta, and pulp contain large numbers of MSCs [3,4]. MSCs are the most commonly used stem cells in animal research and human clinical trials.
Exosomes are extracellular vesicles that can be released into the extracellular environment by all cells [5]. Exosomes can be easily isolated, are small in size, and can reach central nervous system (CNS) injuries by passing through the blood–brain barrier [6]. Therefore, they have a unique ability to treat diseases.
There have been many reports on the application of MSCs for SCI treatment. Recent studies have shown that the ability of MSCs to treat and repair an injured spinal cord is mainly related to the paracrine secretion of exosomes by MSCs, rather than to MSC substitution and multidirectional differentiation. This article briefly reviews the pathological process of SCI and provides information related to MSCs and exosomes. This article also discusses recent studies on the use of MSC-derived exosomes (MSC-exosomes) for the treatment of SCI. We attempt to clarify the repair mechanism of MSC-exosomes in the treatment of SCI and propose exploring the potential application value of MSC-exosomes in the field of cell-free therapy.
Spinal Cord Injury
SCI is a complicated process involving various factors, and its pathophysiological mechanism has not been fully elucidated. SCI involves primary SCI and secondary injury. Primary SCI is directly caused by an external force and occurs within a few minutes after injury, whereas secondary injury is induced by a cascade of factors for several weeks after injury [7].
Injury to the spinal cord caused by an external mechanical force is called primary injury. Primary SCI can lead to spinal cord laceration, contusion, compression, or concussion. It eventually causes the axonal rupture of spinal cord neurons and the destruction of blood vessels and cell membranes [8].
The pathophysiological changes related to secondary SCI are the result of many complicated mechanisms. Secondary SCI can be divided into acute, intermediate, and chronic stages according to the time after injury and the pathological mechanism (Table 1).
Pathophysiological Characteristics of Spinal Cord Injury
SCI, spinal cord injury; BSCB, blood–spinal cord barrier; CSPGs, chondroitin sulfate proteoglycans.
Although SCI can be alleviated through self-repair mechanisms, recovery from SCI requires external intervention. Limiting secondary SCI, promoting axon and myelin formation, and reconstructing neural circuits are the keys in treating SCI [9]. Early surgical decompression and drug treatment for inhibition or alleviation of secondary injury, combined with daily rehabilitation training and nursing to restore functions, are the main treatments for SCI [10,11].
Mesenchymal Stem Cells
Because MSCs are easier to obtain [22], they can be more easily isolated [23], have lower immunogenicity [24], and are subject to fewer ethical restrictions compared with other types of stem cells [25]; they are currently the focus of much research. Interest in MSCs mainly stems from their homing, proliferation, differentiation, secretion, and immunomodulatory abilities. However, because there are also some challenges associated with the application of MSCs, their feasibility and safety are unclear (Table 2).
Characteristics and Functions of Mesenchymal Stem Cells
CXCR, C-X-C motif chemokine receptor; JAK, Janus kinase; MAPK, mitogen-activated protein kinase; MSC, mesenchymal stem cell; STAT, signal transducer and activator of transcription.
MSC-Exosomes
Extracellular vesicles can be divided into two categories, namely, ectosomes and exosomes [5]. Exosomes, which have a diameter of ∼40–160 nm [5], are vesicular bodies with a spherical lipid bilayer membrane [46] (Fig. 1). The biogenesis of exosomes involves a variety of pathways, including an endosomal sorting complex required for the transport (ESCRT)-dependent pathway and an ESCRT-independent pathway [47]. ESCRT comprises five different proteins [48]. ESCRT-0 recognizes ubiquitinated proteins, ESCRT-I/II/III mediates the sprouting of intraluminal vesicles, and the Vps4 complex is responsible for final membrane scission and/or ESCRT recycling [47]. Recently, it was reported that apoptosis-linked gene 2-interacting protein X-ray (ALIX), as a protein related to multivesicle bodies biosynthesis, can directly recruit ESCRT-III to late endosomes independently of other ESCRT interactions, but the binding between ALIX and ESCRT-III requires the presence of lysobisphosphatidic acid [49]. After exosomes are released, the biological activities of the receptor cells are regulated by the release of cargo into the receptor cell through interactions between transmembrane proteins and the receptors on the target cell, endocytosis, or direct fusion of the exosome with the receptor cell membrane [50].
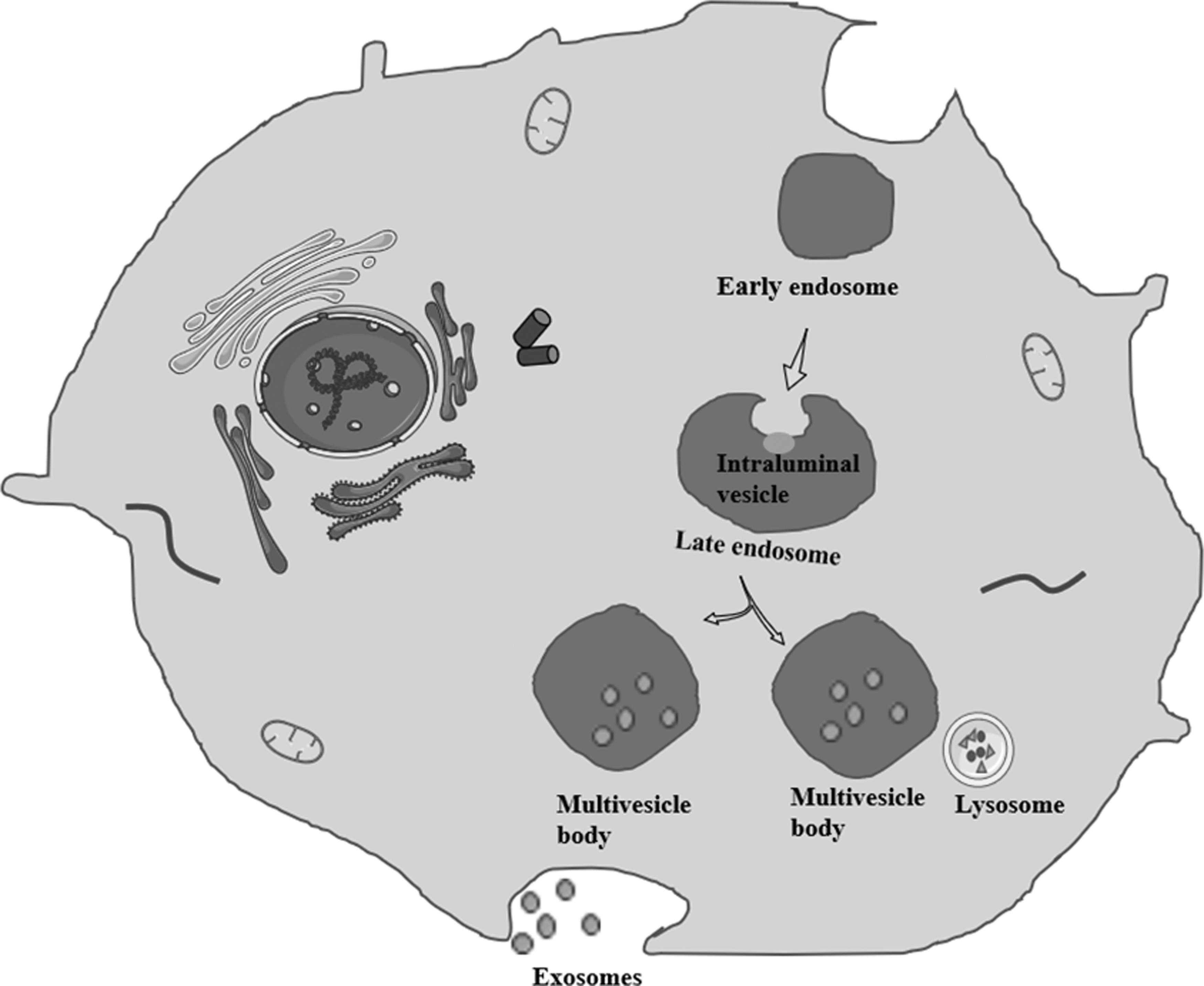
Exosomes biogenesis: exosome biogenesis begins with the formation of early endosomes through cell membrane invagination. Early endosome bud inward, develop into late endosomes, and wrap specific cargos to form intraluminal vesicles. Late endosomes that contain several intraluminal vesicles transform into multivesicle bodies. Multivesicle bodies may be degraded by lysosomes or may fuse with the plasma membrane and be secreted into the extracellular environment as exosomes.
Exosomes contain a large number of complex molecules, such as proteins, lipids, and nucleic acids [51,52] (Fig. 2), which are related to the source cell [53]. Exosomes are rich in lipids that form lipid rafts [54]. The exosomes of different cell types have different lipid compositions [53]. There are also significant differences between exosome lipids in different areas of the same cell [55]. This indicates that even within the same cell, exosomes may be synthesized through different pathways. The proteins in exosomes are mainly involved in metabolism, energy pathways, cell growth, communication, and transportation [56]. The isolation and purification of exosomes affect the biological functions of exosome proteins [57]. Although the specific proteins and their levels expressed by exosomes from different cell types vary, some of these proteins are common surface markers and are widely used to identify exosomes [58]. In addition to carrying proteins and lipids, exosomes contain various nucleic acids, such as mRNA, miRNAs, and other noncoding RNAs (ncRNAs). Among these nucleic acids, miRNAs are rich in exosomes and have therefore received increasing attention. The content of miRNA in exosomes is different from that in the source cell [59]. The content of miRNAs in exosomes during hypertension, cancer, nervous system diseases, and other diseases is significantly different from that in exosomes under physiological conditions [60 –62]. Therefore, the miRNA content can be used as a marker for early diagnosis. In addition, exosomes can be used as efficient carriers to deliver specific miRNAs to receptor cells [63], allowing them to participate in a variety of physiological and pathological processes [64].
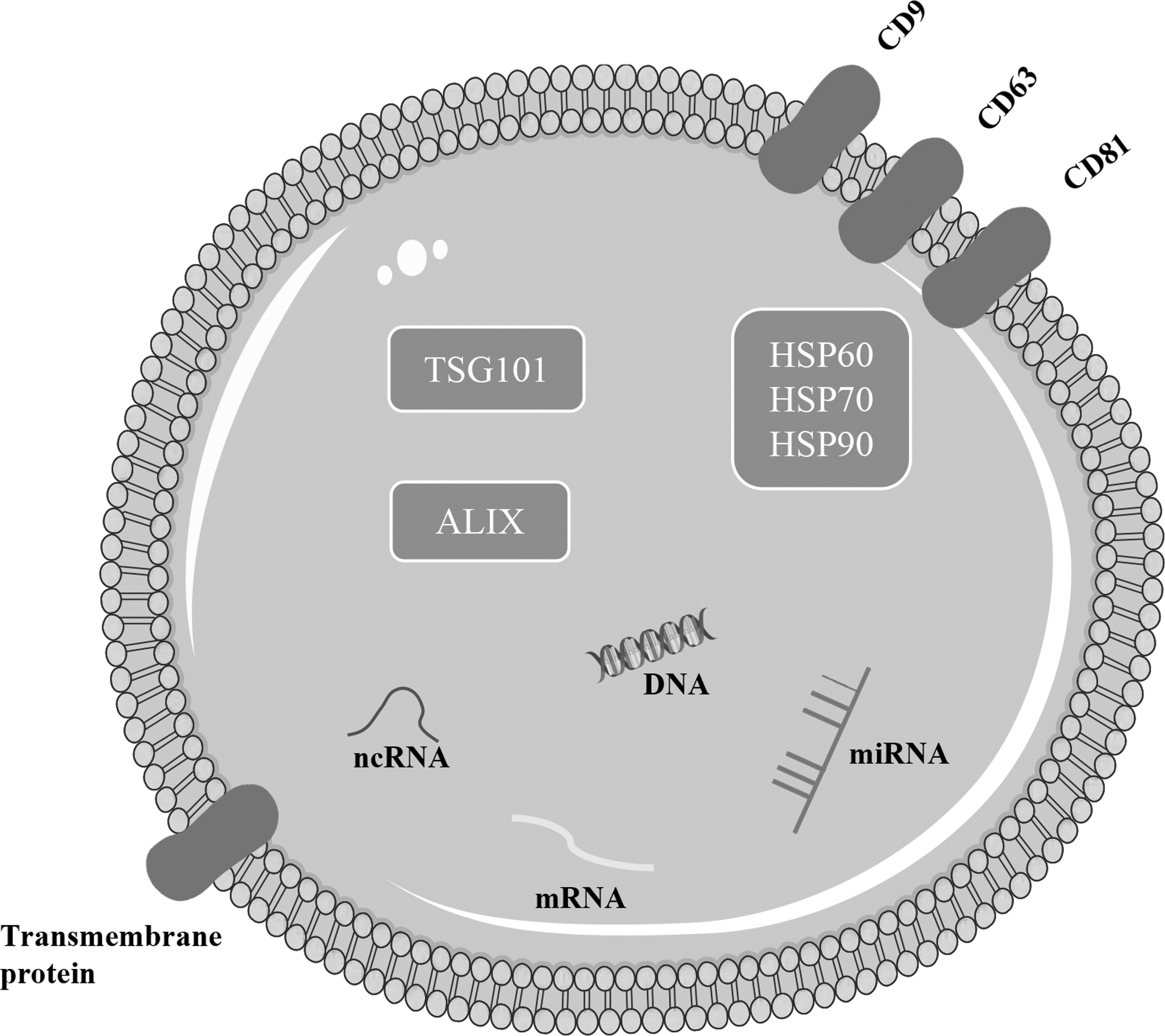
The molecular composition of exosomes: exosome lipids are derived from the cell membrane, and the lipid structure of the bilayer membrane protects the contents of the exosome. Exosomes contain a variety of proteins, including membrane proteins, such as tetraspanin family proteins (CD9, CD63, and CD81). Exosome proteins mainly include the MVB biosynthesis-related proteins apoptosis-linked gene 2-interacting protein X-ray (ALIX), tumor susceptibility gene 101 (TSG101) proteins, and heat shock proteins (HSP60, HSP70, and HSP90). In addition, exosomes also contain genetic materials such as DNA, miRNA, mRNA, and noncoding RNAs (ncRNAs).
MSC-exosomes are mainly derived from bone marrow, adipose, and umbilical cord tissues. The cardiolipin content in MSC-exosomes is very high [53]. MSC-exosomes can also express iconic tetraspanin family members (CD9, CD63, and CD81), heat shock proteins (HSP) 70, and proteins associated with multivesicle bodies [ALIX and tumor susceptibility gene 101 (TSG101)] [65]. miRNAs in MSC-exosomes are mainly involved in angiogenesis and tissue remodeling [66]. Among small RNAs, tRNAs are found at the highest levels in MSC-exosomes, and the types of tRNAs in MSC-exosomes from different sources vary [59]. The biological function of MSC-exosomes is similar to that of MSCs [67,68], but MSC-exosomes are more stable and do not induce tumorigenicity or immune rejection. MSCs produce more exosomes than cells from other sources [69]. Therefore, as alternatives to cell therapy, MSC-exosomes have great potential to promote recovery after injury.
Effects of MSC-Exosomes on SCI: The Underlying Mechanisms
At present, few clinical trials of MSC-exosomes in repair of SCI have been carried out, but there are many related preclinical studies being conducted (Table 3). Studies have shown that MSC-exosomes repair injured spinal cord tissue mainly by inducing angiogenesis and axon formation, regulating inflammation and the immune response, inhibiting apoptosis, and maintaining the integrity of the blood–spinal cord barrier (BSCB) (Fig. 3).
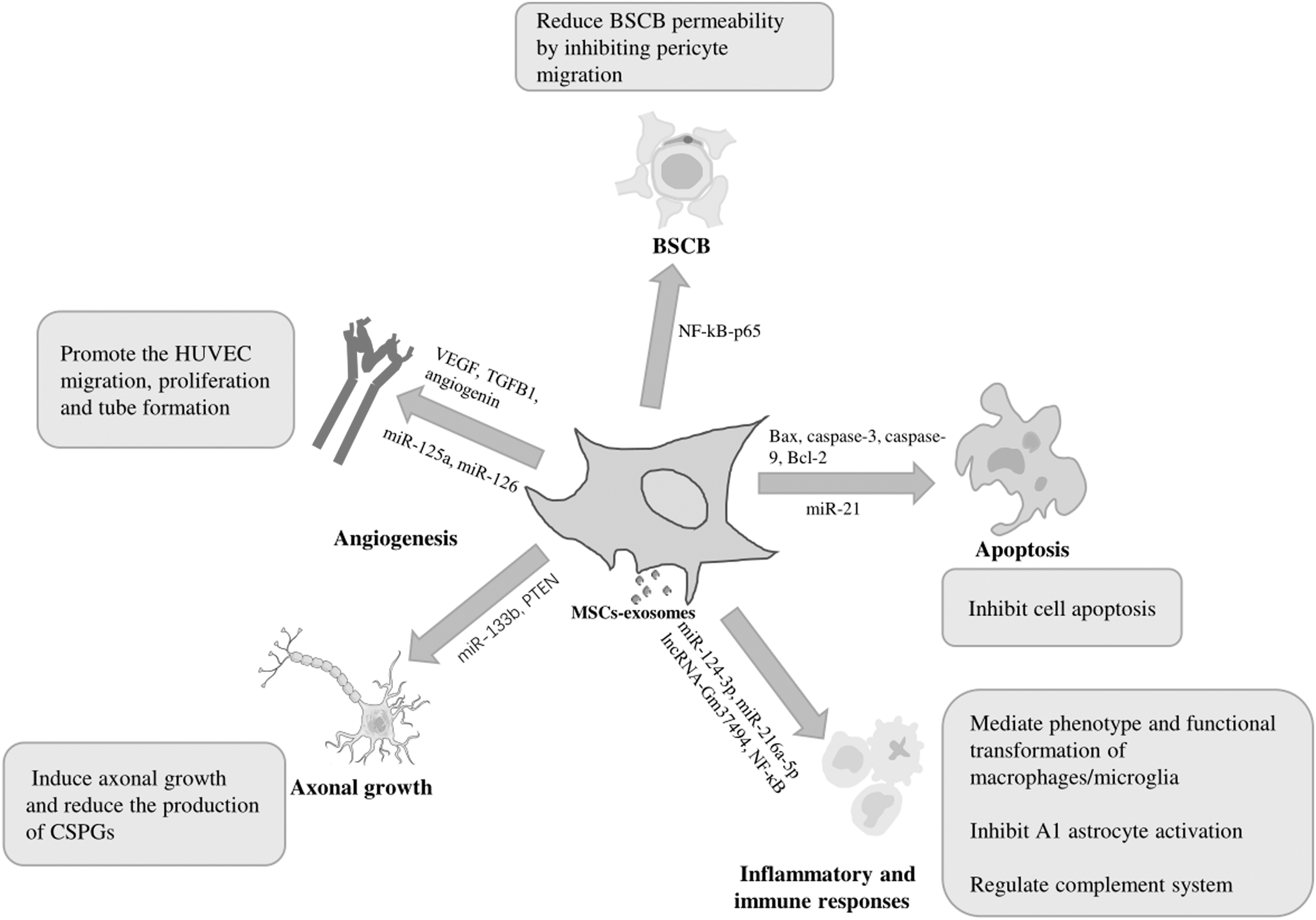
Effects and mechanisms of MSC-exosomes on SCI. BSCB, blood–spinal cord barrier; CSPGs, chondroitin sulfate proteoglycans; HUVEC, human umbilical vein endothelial cells; lncRNA, long noncoding RNA; MSC-exosomes, mesenchymal stem cell-derived exosomes, NF-κB, nuclear factor kappa B; PTEN, phosphatase and tensin homolog; SCI, spinal cord injury; TGFB1, transforming growth factor-beta-1; VEGF, vascular endothelial growth factor.
Current Preclinical Studies of Mesenchymal Stem Cell-Exosomes for Spinal Cord Injury Repair
BSCB, blood–spinal cord barrier; PTEN, phosphatase and tensin homolog.
MSC-exosomes promote angiogenesis
Angiogenesis is crucial for SCI repair. Local blood vessel loss and blood–brain barrier disruption caused by injury can lead to ischemia and inflammation, which can cause comprehensive injury to spinal nerve tissue [89].
Vascular endothelial cells are important components of blood vessel walls. MSC-exosomes can upregulate the expression of key genes involved in angiogenesis in human umbilical vein endothelial cells (HUVECs). Vascular endothelial growth factor (VEGF) plays a key role in the MSC-exosome-mediated proliferation and migration of HUVECs [90]. MSC-exosomes derived from induced pluripotent stem cells are directly taken up by HUVECs. There, they activate the expression of VEGF, angiogenin, and transforming growth factor-beta 1 (TGFB1) to promote the migration, proliferation, and tube formation of HUVECs [91]. After SCI, the spinal cord tissue is often in a hypoxic state. Hypoxia-treated MSC-exosomes are more easily taken up by HUVECs compared with normal MSC-exosomes and can also increase VEGF expression in HUVECs by activating the protein kinase A signaling pathway [92].
MSC-exosomes can release endogenous angiogenic miRNAs and transfer exogenous angiogenic miRNAs to HUVECs to regulate the target gene of the miRNA. Human adipose-derived MSC-exosomes are rich in miR-125a. miR-125a taken up by HUVECs inhibits the expression of delta-like ligand 4 and promotes angiogenesis by specifically binding to the 3′ untranslated region of delta-like ligand 4 [93]. miR-126-modified MSC-exosomes can also exert a similar proangiogenic effect, mainly by inhibiting the ability of miR-126 to directly target phosphoinositide-3-kinase regulatory subunit 2 and sprouty-related EVH1 domain-containing protein 1 in HUVECs [82].
MSC-exosomes promote axonal growth
The regeneration of injured axons or the stimulation of the growth of spare axons at the injury site is of great significance for the recovery of spinal cord function after SCI [94].
MSC-exosomes can alleviate nerve conduction disorders by inducing axonal growth and reducing the production of chondroitin sulfate proteoglycans (CSPGs). MSC-exosomes can promote the axonal growth of cortical neurons in vitro. The distal axon length increases by 33% and 24%, respectively, in cortical neurons cultured with MSC-exosomes for 24 and 48 h compared with control neurons [95]. However, the internalization of MSC-exosomes into neurons and distal axons requires the core protein-soluble NSF attachment protein receptor, which mediates vesicle fusion at synapses [95]. Whether glial scarring hinders axonal regeneration is controversial. Recently, it was suggested that preventing or ablating astrocyte reactivity in the scar does not stimulate axonal growth or inhibit axonal regeneration. The main mechanism by which axonal regeneration is inhibited is the generation of CSPGs [96]. In rats subjected to spinal cord contusion and injected with MSC-exosomes, CSPG deposition in the spinal cord lesion area is significantly lower, and axonal regeneration is nearly 80% higher than that in untreated injured rats [18].
In addition, MSC-exosomes carrying specific miRNAs that mediate axonal growth can better promote axonal growth compared with MSC-exosomes carrying an empty vector. In a rat model of SCI, the injection of miR-133b-modified MSC-exosomes increases the expression of growth-related protein 43 and neurofilament, indicating axonal regeneration [74]. miR-133b increases the expression of extracellular-regulated protein kinases 1/2, signal transducer and signal transducer and activator of transcription (STAT) 3, and cAMP-response element binding protein, to affect axon regeneration-related pathways [74]. By downregulating the high expression of phosphatase and tensin homolog (PTEN) after SCI, the intranasal delivery of PTEN gene-silenced MSC-exosomes can target damaged sites to promote axonal regeneration [76]. The PTEN-mammalian target of rapamycin (mTOR) pathway is a key factor in axonal regeneration [97]. The inhibition of PTEN can activate the synthesis of proteins downstream of mTOR signaling and promote the regeneration of injured CNS axons [98]. Therefore, MSC-exosomes can be used as efficient miRNA and small interfering RNA carriers for CNS diseases.
MSC-exosomes regulate inflammatory and immune responses in SCI
MSC-derived exosomes can communicate with the inflammatory microenvironment to play a neuroprotective role and can adjust the complementary systems to promote spinal cord function recovery.
MSC-exosomes mediate the phenotype and functional transformation of macrophages/microglia
Microglia are macrophages in the CNS. Activated microglia and macrophages are also called macrophages/microglia because they are not easily distinguished [99]. Macrophages/microglia have high plasticity. Different subpopulations of macrophages/microglia, namely, M1 proinflammatory macrophages and M2 anti-inflammatory macrophages, are present at the site of SCI [100]. The ratio of these two phenotypes is important for SCI repair.
MSC-exosomes can be specifically localized to the site of SCI in rats and can bind to M2 macrophages [71], thereby increasing the production of anti-inflammatory cytokines. In a mouse model of SCI, a tail vein injection of MSC-exosomes can trigger the transformation of macrophages from the M1 phenotype to the M2 phenotype [72]. Activated M2 macrophages downregulate the release of tumor necrosis factor (TNF)-α, macrophage inflammatory protein-1α, interleukin (IL)-6 and interferon (IFN)-γ to reduce inflammation in the injured area and increase the expression of IL-4 and IL-10 to promote healing of the injury site [72]. However, the mechanism through which MSC-exosomes mediate the phenotypic transformation of macrophages was proven to require stimulation by proinflammatory factors. After preactivation of the proinflammatory cytokines IFN-γ/TNF-α, MSC exosomes acquire the ability to shift M1 macrophages to the M2 phenotype. This process may be related to miRNAs in MSC-exosomes, such as miR-34a-5p, miR-21, and miR146a-5p [101].
MSC-exosomes can regulate macrophage/microglial polarization through regulation of pathways by exogenous miRNAs and long ncRNAs (lncRNAs). Endoplasmic reticulum stress is induced after SCI [102], and miR-124-3p can inhibit the expression of the endoplasmic reticulum to nucleus signaling 1, which is a major regulator of endoplasmic reticulum stress [87]. Exogenous miR-124-3p is transferred to macrophages through MSC-exosomes and enhances the polarization of M2 macrophages by inhibiting the expression of endoplasmic reticulum to nucleus signaling 1 [87]. In addition, MSC-exosomes carrying miR-216a-5p activate phosphatidylinositide 3-kinases (PI3K)/serine/threonine kinase (AKT), inhibit the Toll-like receptor 4/nuclear factor kappa B (NF-κB) signaling pathways, and transform M1 microglia into M2 microglia [83]. MSC-exosomes can also carry lncRNAs with nerve repair functions to target cells. MSC-exosomes modified by lncRNA-Gm37494 can shift the M1/M2 phenotype of microglia/macrophage M1/M2 phenotypes by inhibiting miR-130b-3p and promoting the expression of peroxisome proliferator-activated receptor γ [103].
MSC-exosomes inhibit A1 astrocyte activation
Neuroinflammation and ischemia in the CNS can alter the phenotypes of A1 and A2 reactive astrocytes, respectively [104]. A1 astrocytes, which are induced by IL-1α, TNF-α, and C1q secreted by microglia, lose the ability to promote neuronal survival, synapse formation, growth, and phagocytosis and can also lead to neuronal and oligodendrocyte death [105]. However, A2 astrocytes promote neuronal survival and tissue repair [104].
MSC-exosomes can exert anti-inflammatory and neuroprotective effects by inhibiting the activation of NF-κBp65, thereby reducing the number of A1reactive astrocytes. MSC-exosomes can reduce the expression of the A1 astrocyte marker glial fibrillary acidic protein and its complement C3 in astrocytes [73]. They can also reduce the expression of the NF-κB-related protein p-IkBα and nuclear p-p65, but the expression of NF-κB inhibits protein IkB-α from regulating the NF-κB signaling pathway [73].
MSC-exosomes regulate the complement system
Both classic and alternative complement pathways can be activated after SCI [106]. The activation of these pathways may aggravate the inflammatory process and secondary injury to the spinal cord.
MSC-exosomes can be used as complementary inhibitors to reduce complementary mRNA expression while inhibiting the NF-κB signaling pathway and promoting SCI recovery. In SCI rats injected with MSC-exosomes, the mRNA levels of complements C1q, complement factor H, C3, C4b, C6, C5, mannan-binding lectin, and complement factor P are lower than those in uninjected SCI rats, and SCI is alleviated [77]. Complements C1q and C3 are related to the NF-κB signaling pathway [107,108]. The secondary inflammatory response caused by SCI is regulated by NF-κB [109]. Inhibiting NF-κB can reduce the inflammatory response and promote functional recovery after SCI. In addition, MSC-exosomes can downregulate p-NF-κB and phosphorylated-IκBα levels and inhibit SCI-induced NF-κB subunit expression [77]. Therefore, targeting key complements with MSC-exosomes may be an effective method for alleviating SCI.
MSC-exosomes inhibit cell apoptosis
Apoptosis in secondary SCI leads to irreversible changes in neurons and aggravates damage. Therefore, it is necessary to suppress neuronal apoptosis.
MSC-exosomes reduce apoptosis by regulating apoptosis-related proteins and miRNA transfer. MSC-exosomes can significantly downregulate the apoptosis-related markers Bax, caspase-3, and caspase-9 in rats after SCI, increase the level of the antiapoptotic protein Bcl-2 and further enhance the Wnt/β-catenin signaling pathway to exert an antiapoptotic effect [78]. In addition, MSC-exosomes can also reduce neuronal apoptosis by promoting neuronal autophagy. MSC-exosomes can activate autophagy and increase the number of autophagosomes [86]. Consistent with in vitro findings, MSC-exosomes increase the expression of the autophagy-related proteins Beclin-1 and LC3-II in SCI rats on the third day after treatment but decrease the protein expression of P62 [86].
As one of the miRNAs that is significantly upregulated after SCI, miR-21 can downregulate the expression of PTEN, programmed cell death factor 4 (PDCD4), and FasL. It also inhibits neuronal apoptosis and plays a protective role. MSC-exosomes transfected with miR-21 can inhibit the expression of PTEN/PDCD4 and reduce nerve cell apoptosis [81]. In addition, miR-21-5p exerts an antiapoptotic effect that is also related to the decreased expression of FasL, which is a proapoptotic gene [110]. Of note, miR-21 deficiency in obese rats is caused by insulin resistance, and secreted MSC-exosomes have no significant effect on SCI rats [79]. The above results further prove that MSC-derived exosomes can alleviate SCI by inhibiting apoptosis. The functions of MSC-exosomes carrying specific miRNAs that inhibit apoptosis are affected under certain pathophysiological conditions.
MSC-exosomes reduce the BSCB permeability by inhibiting pericyte migration
Pericytes are indispensable for the formation and function of the BSCB. The loss of barrier integrity leads to increased permeability and the penetration of toxic products into the spinal cord, resulting in edema and death of spinal cord neurons [111].
MSC-exosomes injected into the injury site of contusion SCI rats are taken up by pericytes and downregulate NF-κBp65 in these cells to inhibit pericyte migration, improve the coverage of pericytes, and reduce the permeability of the BSCB [75]. Previous studies have explored how MSC-exosomes maintain the integrity of the BSCB after SCI. Thus, targeting the BSCB is a promising treatment strategy for SCI.
Conclusions
Taken together, these data suggest that MSC-exosomes are new tools for SCI repair. However, before they can be used for clinical applications, the many challenges related to the tissue sources, isolation, purification, amplification, and joint application of MSC-exosomes need to be addressed.
MSC-exosomes from various tissue sources contain different inclusions and thus have distinct abilities to repair damage. In practical applications, MSC-exosomes should be chosen according to the functions of different source tissues. The physical and chemical properties of MSC-exosomes obtained through different separation methods are also different. It is necessary to establish a quick, cheap, and simple standardized separation and purification procedure to generate MSC-exosomes with a high yield and purity without impacting the biological activity of the exosomes. Obtaining a sufficient number of MSC-exosomes for repair is also a challenge. Physiological stimuli or small-molecule modulator pretreatment can increase the secretion of MSC-exosomes [112]. However, the physical and chemical properties, composition, and function of MSC-exosomes obtained using these methods need to be investigated in depth. Because of the limitations related to the application of MSC-exosomes alone, many studies currently use MSC-exosomes in combination with biological materials or use MSC-exosomes as gene or drug delivery vehicles. Thus, the combined effects of MSC-exosomes and the mechanism of targeted delivery need to be further elucidated.
In conclusion, we believe that the combination of MSC-exosomes and SCI has infinite potential. Research on the detailed mechanism and further evaluation of the safety and effectiveness of MSC-exosomes and SCI will promote the development of this field.
Footnotes
Acknowledgment
The authors thank American Journal Experts, LLC, for editing the English text of a draft of this article.
Author Disclosure Statement
The authors declare that they have no conflicts of interest.
Funding Information
This study was supported by the Visiting Scholar Program sponsored by the China Scholarship Council (201906915019), the National Natural Science Foundation of China (no. 81871794), and the Beijing Municipal Health Commission (Jing2019–2).