
Abstract
As an important component of the dynamic tumor microenvironment, mesenchymal stem cells (MSCs) can interact with tumor cells to promote tumor growth. Treatment with tumor cell-derived exosomes can change the biological functions of MSCs. We want to study the mechanism by which exosomes derived from gastric cancer cells affect the biological functions of MSCs. After MSCs were treated with adenocarcinoma gastric cells (AGS) cell-derived exosomes, circular RNAs differentially expressed in MSCs were verified using existing RNA microarray results combined with quantitative real-time polymerase chain reaction (qRT-PCR). Then, circular RNAs were knocked down or overexpressed by plasmids, and the functions of circular RNAs were evaluated by Migration and invasion assay. Dual luciferase reporter assay was used to evaluate the potential mechanism of circular RNAs. After treatment with exosomes secreted by AGS, the results showed that some circular RNAs expressed by human adipose-derived MSCs showed significant differences. The elevated circ_0004303 promoted the migration and invasion of human adipose-derived MSCs in vitro. Circ_0004303 upregulated the expression of activated leukocyte cell adhesion molecule (ALCAM) by acting as a miR-148a-3p sponge, thereby enhancing the migration and invasion functions of human adipose-derived MSCs. Therefore, exosomes secreted by AGS can affect the expression of circular RNAs in human adipose-derived MSCs. Hsa_circ_0004303 can regulate the migration and invasion of human adipose-derived MSCs via the miR-148a-3P/ALCAM axis. This study suggests that tumor cells can promote the migration and homing of MSCs in adjacent tissues by secreting exosomes.
Introduction
Gastric cancer has the fifth highest incidence rate and the fifth highest mortality rate among cancers in the world [1]. Cases of gastric cancer in China alone account for 42.6% of the global incidence rate and 45% of the global mortality rate [2]. Although important progress has been made in the treatment of gastric cancer through conventional treatment combined with chemotherapy and targeted drugs, the results of many phase III clinical trials are less than satisfactory, and many challenges remain for advancing the current treatment of gastric cancer [3]. Further research is urgently needed to identify more effective diagnosis and therapeutic schedules.
The tumor microenvironment (TME) includes surrounding mesenchymal stem cells (MSCs), immune cells, extracellular matrix, fibroblasts, and lymphocytes [4], and the interactions among these components affect the occurrence and development of tumors. The composition and structure of TME vary with cancer types and patients. As an important component of the TME, human MSCs can be extracted from a variety of adult organs and tissues, such as adipose tissues, bone marrow, and peripheral blood. MSCs have differentiation ability of osteogenesis, chondroblast, and adipogenesis [5].
Adipose MSCs have been found in human gastric cancer. Adipose tissue exists in the gastric submucosa and subserosa. Adipose tissue stromal cells influence the progression of gastric cancer by increasing their growth/invasion and decreasing their apoptosis through MAPK activation in a COX-2-independent way [6]. Helicobacter pylori, as a part of the gastric cancer microenvironment, can promote the migration of human adipose-derived MSCs into the TME and transformation into cancer cells [7]. MSCs that are recruited into the TME can indirectly affect the growth and metastasis of tumor cells by secreting a variety of inflammatory cytokines and chemokines in the TME to regulate immune cells functions. MSCs can also play an important role in direct interaction with immune cells and tumor cells [8].
Tumor cells can modulate tumor stromal cells through intercellular communications. This process can be mediated through either direct cell contact or secreted signaling factors (cytokines, chemokines, and growth factors) and microvesicles [9]. Exosomes are extracellular vesicles that can be generated by all cells. They carry soluble factors such as nucleic acids, proteins, enzymes, lipids, and cytokines and serve significant information transmission functions between cells [10].
Recently, a growing number of studies have investigated the application of exosomes in the TME. Tumor cell-derived exosomes carry tumor-associated antigens (TAA), costimulatory molecules, and major histocompatibility complex components, which enable them to stimulate immune cells and promote antitumor responses [11]. Lung cancer cell-derived exosomes can activate the Toll-like receptor 2 (TLR2)/nuclear factor kappa B (NF-κB) signaling pathway and induce MSCs to be proinflammatory MSCs, thereby promoting tumor growth [9]. Compared with bone marrow-derived MSCs (BM-MSCs), MSCs cultured with melanoma cell-derived exosomes (TE-MSCs) significantly promoted tumor growth [12].
Circular RNAs are a subclass of noncoding RNAs, which have a covalently closed-loop structure [13]. The expression of circular RNAs is specific at different tissue and different stages of development. Since circular RNAs lack free 5′ and 3′ terminals and are not sensitive to nucleases, they are stabler than linear RNA [14]. With the development of RNA sequencing technology, there has been an increasing interest in the study of circular RNAs, which are closely related to many physiological and pathological processes. For example, circMTO1 acts as a sponge for miR-9 to upregulate p21 expression and inhibit hepatocellular carcinoma progression [15]. Circ- FBXW7 encodes a new protein, FBXW7–185aa, which competitively interacts with USP28 and releases FBXW7a to degrade c-Myc, thereby inhibiting glioma cell proliferation and slowing down the cell cycle [16]. However, research studying the role of circular RNAs in gastric cancer is still very limited.
In this study, a novel circular RNA was identified from the DDX60, termed circ_0004303. After human adipose-derived MSCs were treated with exosomes derived from gastric cancer cells, circ_0004303 was significantly upregulated, and the high level of circ_0004303 enhanced the migration and invasion abilities of human adipose-derived MSCs. With regard to the mechanism of this effect, we found that an elevated circ_0004303 level affected the functions of human adipose-derived MSCs by sponging miR-148a-3p to upregulate activated leukocyte cell adhesion molecule (ALCAM) expression.
Materials and Methods
Cell culture
Human adipose-derived MSCs were isolated as previously reported [17] from discarded adipose tissue from patients undergoing liposuction in the Plastic surgery department of Peking Union Medical College Hospital. They have been approved by the Ethics Committee of the Institute of Basic Medicine of the Chinese Academy of Medical Sciences and obtained the informed consent of the patients.
Gastric cancer cell line adenocarcinoma gastric cells (AGS) was purchased from the Cell Resource Center, Institute of Basic Medicine, Chinese Academy of Medical Sciences. AGS was cultured in Dulbecco's modified Eagle medium (DMEM)/F12 containing 10% fetal bovine serum in a cell incubator at 37°C with a constant temperature and 5% CO2.
Exosome extraction
Exosome extraction was performed as previously described [18]. In brief, AGS cells were cultured in serum-free DMEM/F12 medium for 36 h, and then the supernatant was collected and centrifuged at 3,000 rpm for 30 min to remove lifted cells. The supernatant was transferred to 100,000 Mw cutoff membrane (CentriPlus-70, Millipore) and centrifuged to extract exosomes by ultrafiltration method. The volume of supernatant was reduced from approximately 250–500 mL to <3 mL. The supernatant was washed twice with phosphate-buffered saline (PBS) until it was approximately colorless. The supernatant was filtered with a 0.22 m filter and preserved at −80°C.
Transmission electron microscopy
Purified exosomes were fixed with 1% glutaraldehyde in PBS (pH 7.4). After rinsing, a 20-uL drop of the suspension was loaded onto a formvar/carbon-coated grid, negatively stained with 3% (w/v) aqueous phosphotungstic acid for 1 min and observed by transmission electron microscope.
Western blot analysis
Proteins were extracted with radio immunoprecipitation assay (RIPA) lysis buffer with phenylmethylsulfonyl fluoride (PMSF), quantified by bovine serum albumin (BCA) Protein Assay Kit, separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis gel, and then transferred onto a polyvinylidene fluoride membrane. The membrane was incubated with the primary antibody and then incubated with the horseradish peroxidase (HRP)-conjugated secondary antibody for 1 h at room temperature. The primary antibodies were as follows: HSP70 (Santa Cruz), HSP90 (Santa Cruz) and CD63 (Cell Signaling Technology). Secondary (HRP)-conjugated antibodies were purchased from NeoBioscience. The immunodetection was then visualized by a chemiluminescent ECL reagent (Millipore).
Real-time PCR
Total RNA was extracted from cells using TRIzol reagent (Life Technologies) according to the manufacturer's protocol. Triplicate reactions were performed for each complementary DNA (cDNA) sample. The relative expression of each gene to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was calculated using the ΔCT method. The gene-specific primers are listed in Table 1.
Primers for Semiquantitative Reverse Transcription-Polymerase Chain Reaction and Quantitative Real-Time PCR
ALCAM, activated leukocyte cell adhesion molecule; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; IDO, indoleamine-2,3-dioxygenase; IL, interleukin; MCP-1, monocyte chemotactic protein 1; TGF-β, transforming growth factor beta; TNF-α, tumor necrosis factor alpha.
Exosome staining
Exosomes labeled with Dil dye was cocultured with MSCs. The uptake was observed in real-time using a living cell imager at different time points (from 1 to 24 h). After 24 h of uptake, the MSCs were fixed with 4% paraformaldehyde for 10 min. Then the nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI) for 5 min and the results were observed.
Cell transfection
AGS cells were seeded into six-well plates (5 × 104 cells/well) and cultured overnight to ∼70% confluence in a humidified atmosphere containing 5% CO2 at 37°C. The cells were then transfected with 100 nM of small interfering RNA (siRNA), overexpression vector or negative control for circular RNA purchased from GeneBiogist (Shanghai, China) by using Lipofectamine 3000 (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. At 48 h after transfection, the transfected cells were collected to analyze the efficiency of transfection by quantitative real-time polymerase chain reaction (qRT-PCR).
Transwell assay
Migration of AGS cells was assayed using chamber with 8-μm-pore filters. AGS cells were trypsinized and 5 × 104 AGS cells in serum-free medium were added to the upper chamber. Then, 1 mL media with 10% serum was added to the lower chamber. Cells were incubated for 18 h at 37°C, and then nonmigrating cells were removed with cotton swabs. Cells that migrated to the bottom of the membrane were then fixed with 4% paraformaldehyde for 20 minutes and stained with 0.2% crystal violet for 30 min, and then washed twice with PBS. Then stained cells were visualized under a microscope.
Luciferase reporter assay
The circ_004303 3′ untranslated region (3′-UTR) containing the potential binding site of hsa-miR-148a-3p was cloned into the pRL-TK vector (Invitrogen) by PCR method. In brief, cells were inoculated into a 24-well plate and were cotransfected with miR-148a-3p mimics or the negative control and the luciferase reporter vector as well as lipofectamine 2000 reagent (Invitrogen). After 48 h, cells were collected to measure Firefly and Renilla luciferase activity using the dual-luciferase reporter assay system (Promega).
Statistical analysis
Data are presented as mean ± standard deviation, which were representative at least in triplicate. The two-tailed Student's t-test was used to evaluate the significance of differences between two groups. Statistical analysis was performed using GraphPad Prism 5.0 (GraphPad Software, Inc., San Diego, CA). A value of P < 0.05 was considered to be statistically significant.
Results
Differentiation potential and phenotypic identification of human adipose-derived MSCs
The human adipose-derived MSCs isolated from adipose tissue grew adherently and showed a fusiform shape (Fig. 1A). Lipid formation, osteogenesis, and cartilage differentiation of human adipose-derived MSCs were determined by oil red O, alizarin red, and alkaline phosphatase staining. The results showed that human adipose-derived MSCs were capable of lipid formation, osteogenesis, and chondrogenic differentiation (Fig. 1B–D). Further analysis of the immunophenotype of stem cells indicated that CD90, CD29, CD44, and CD73 were positive (positive rate >95%), and HLA-DR, CD34, CD31, and CD106 were negative (positive rate <5%; Fig. 1E).
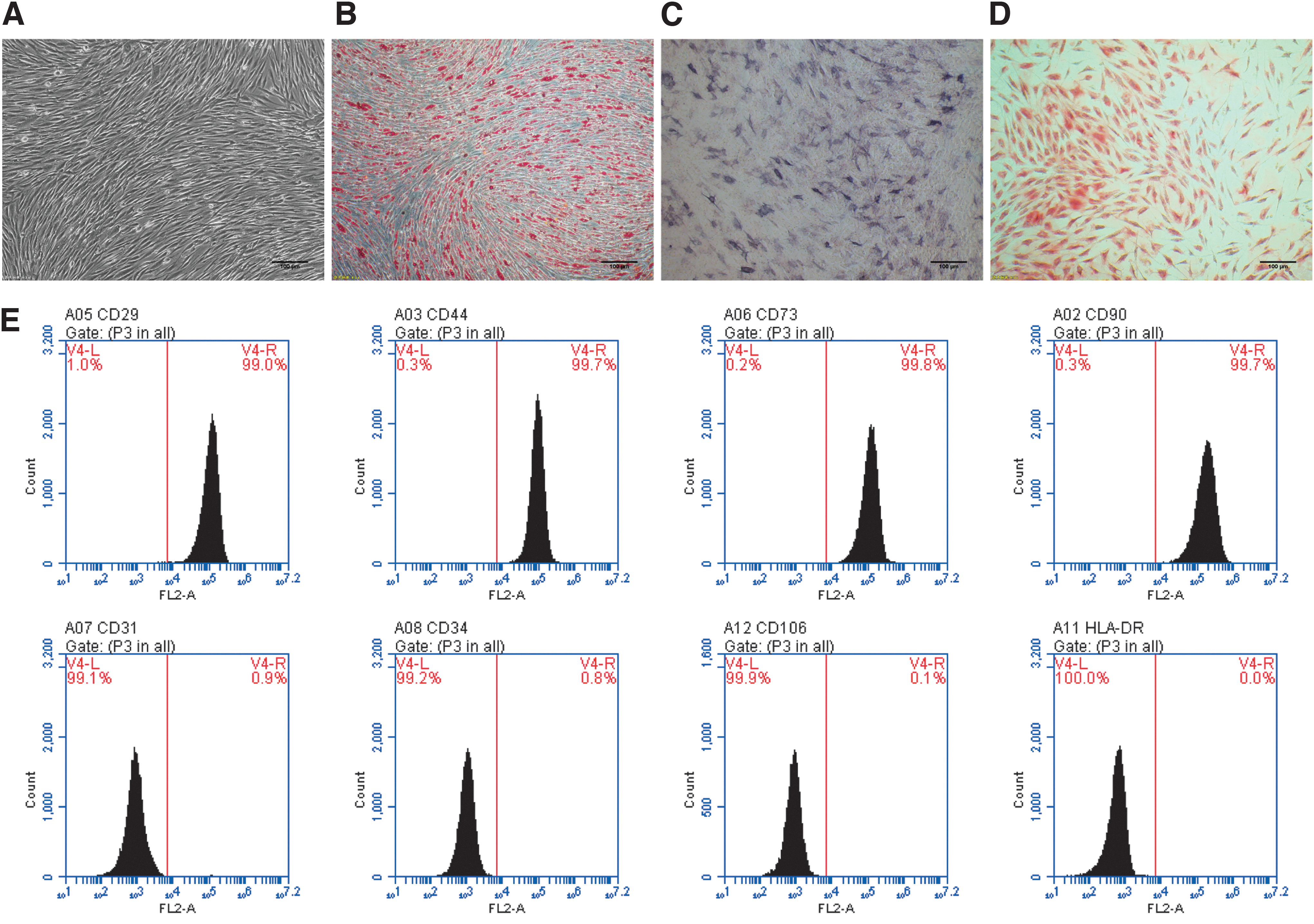
Differentiation potential and phenotypic identification of hAMSCs.
Identification of AGS cell-derived exosomes and observation of exosomes entering human adipose-derived MSCs
Exosomes were isolated from AGS cells using ultrafiltration. Transmission electron microscopy showed that the exosomes were microvesicles with a diameter of about 40–100 nm (Fig. 2A). Western blot analysis showed that AGS cell-derived exosomes expressed HSP70, HSP90, and CD63 (Fig. 2B). NanoSight analysis showed that AGS cell-derived exosomes were uniform in size (Fig. 2C). To examine whether AGS cell-derived exosomes could be ingested by human adipose-derived MSCs, AGS cell-derived exosomes were labeled with DIR dye and cocultured with human adipose-derived MSCs. The process of exosomes uptake was observed in real-time, and the exosomes were observed in the cytoplasm of human adipose-derived MSCs after 10 h under a microscope (Fig. 2D). The nuclei of human adipose-derived MSCs were stained blue with Hoechst33342 dye (Fig. 2E). These results indicated that human adipose-derived MSCs could effectively ingest AGS cell-derived exosomes.
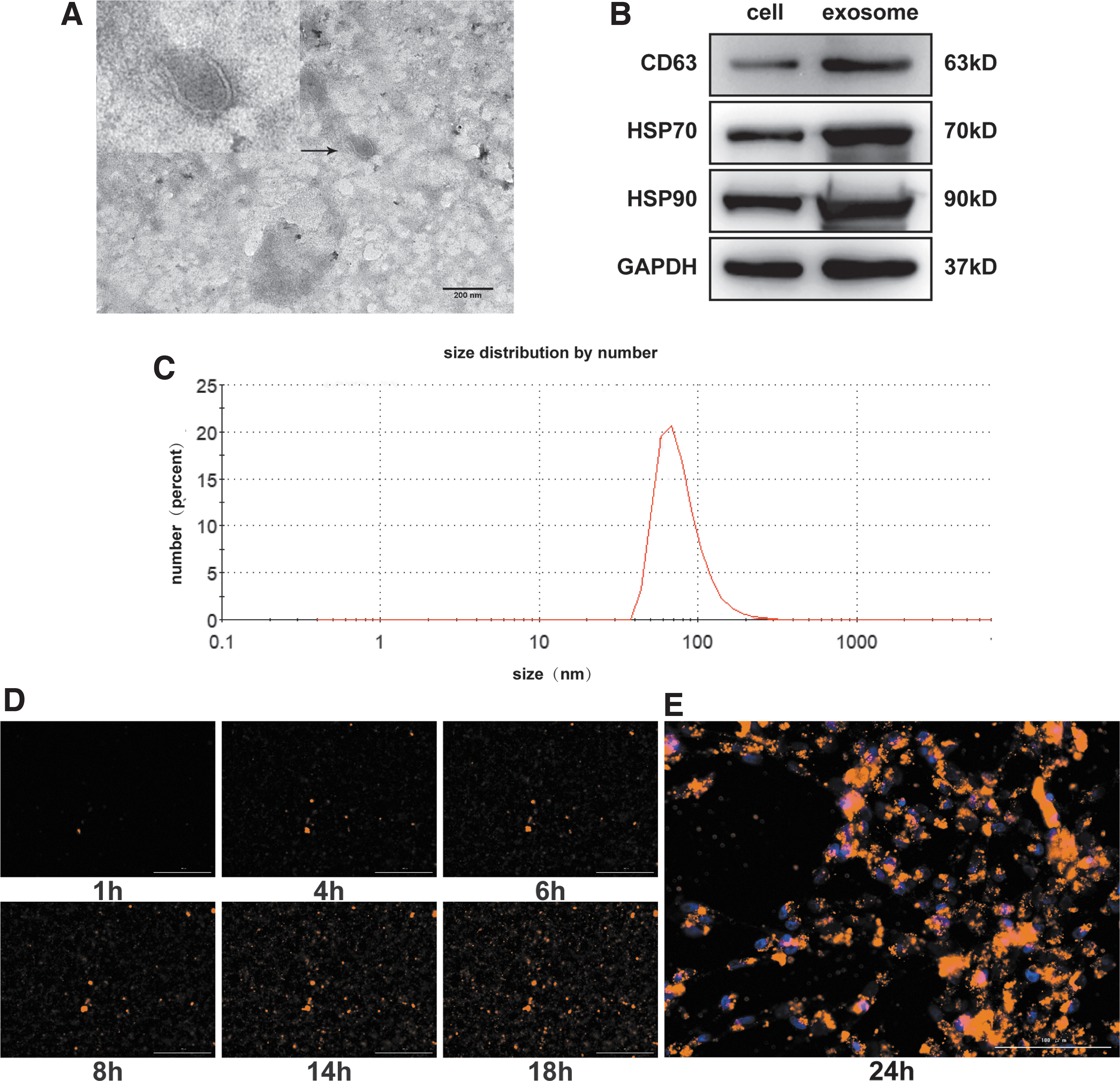
Characterization of exosomes secreted by AGS cancer cells.
Time and concentration dependence of the effects of AGS cell-derived exosomes on human adipose-derived MSCs
Previous research has found significant differences in inflammatory cytokines secreted by human adipose-derived MSCs after treatment with AGS cell-derived exosomes [19]. This study further verified this phenomenon. QRT-PCR results showed that the expressions of proinflammatory cytokines interleukin (IL)-6 and IL-8, chemokines monocyte chemotactic protein 1 (MCP-1) and IP-10, and anti-inflammatory cytokines indoleamine-2,3-dioxygenase (IDO) secreted by human adipose-derived MSCs were significantly upregulated after the treatment of AGS cell-derived exosomes (Fig. 3A).
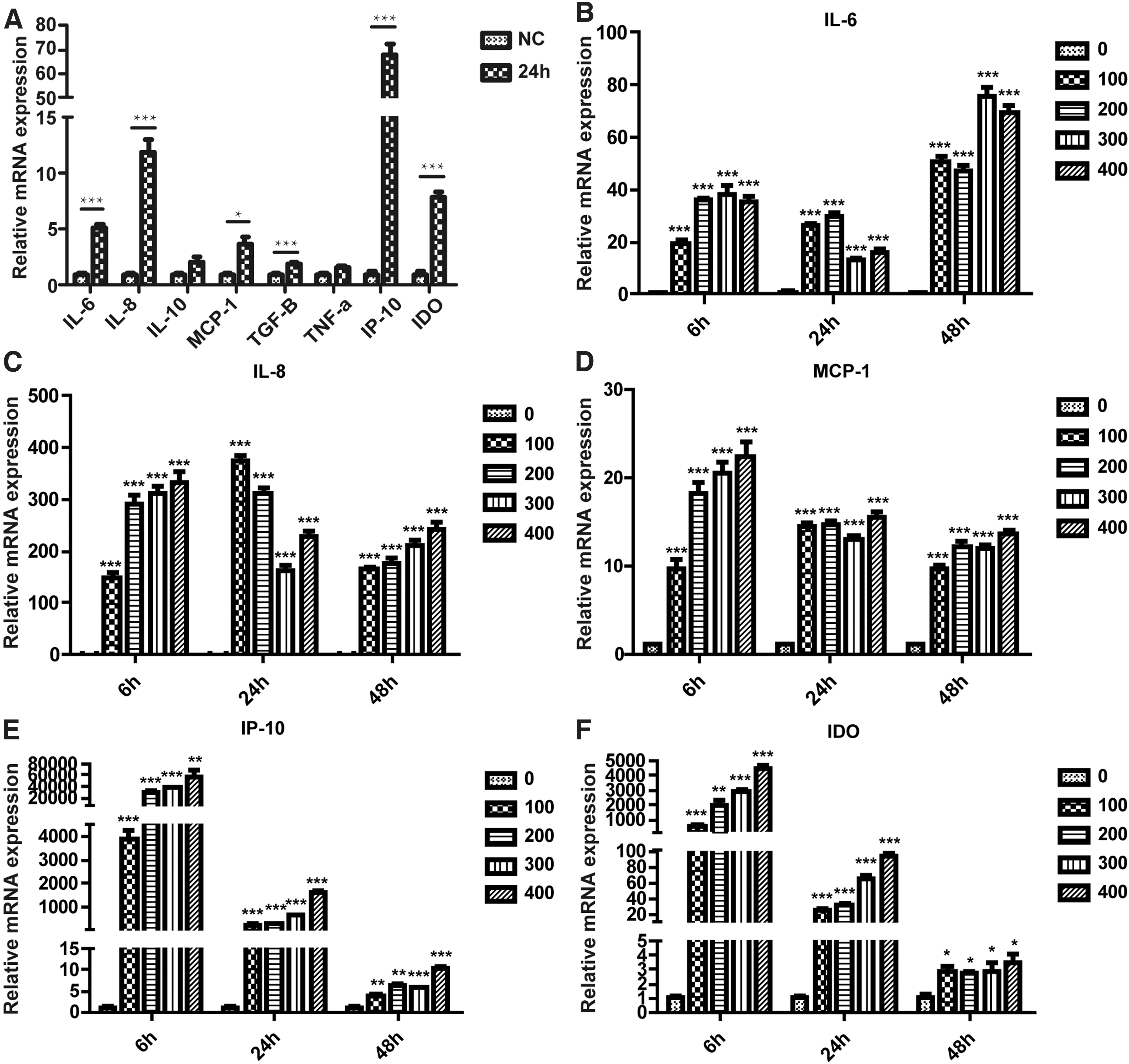
Effects of AGS cell-derived exosomes on secretion of inflammatory cytokines in hAMSCs.
To explore whether AGS cell-derived exosomes have time-dependent and concentration-dependent effects on human adipose-derived MSCs, we treated human adipose-derived MSCs with different concentrations of AGS cell-derived exosomes for 6, 24, and 48 h. The qRT-PCR results (Fig. 3B–F) showed that different concentrations of exosomes hardly affected the expression levels of inflammatory factors at the same time point. With the extension of time, the mRNA levels of IL-6 secreted by human adipose-derived MSCs showed an increasing trend, while the mRNA levels of IL-8, MCP-1, IP-10, and IDO secreted by human adipose-derived MSCs showed a decreasing trend. Combined with the treatment conditions and the expression of inflammatory factors reported in literature, the concentration and time of treatment were finally determined to be 200 ng/μL and 24 h.
Effects of AGS cell-derived exosomes on human adipose-derived MSCs gene expression profile
Existing gene microarray results showed that a total of 135,750 differentially expressed genes were detected before and after treating human adipose-derived MSCs with AGS cell-derived exosomes for 24 h (P ≤ 0.05) [19]. Compared with the control group, 69,091 genes were upregulated, and 66,659 genes were downregulated in the human adipose-derived MSCs treated with exosomes.
Gene Ontology (GO) analysis showed that these differentially expressed genes were mainly related to proteasome activator complex, apical plasma membrane, chloride ion channel complex, and ICE protease-activating factor inflammatory complex (cellular component); type I interferon signaling pathway, defense against virus, neuron migration, response to virus, and interferon-β (biological process); and double-stranded RNA binding, 2′–5′-oligosadenylate synthase activity, and olfactory receptor activity were associated with single-stranded RNA binding (molecular function) (Fig. 4A). Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis showed that it was mainly enriched in NOD-like receptor signaling pathway, RIG-I-like receptor signaling pathway, NF-κB signaling pathway, and Toll-like receptor signaling pathway (Fig. 4B).
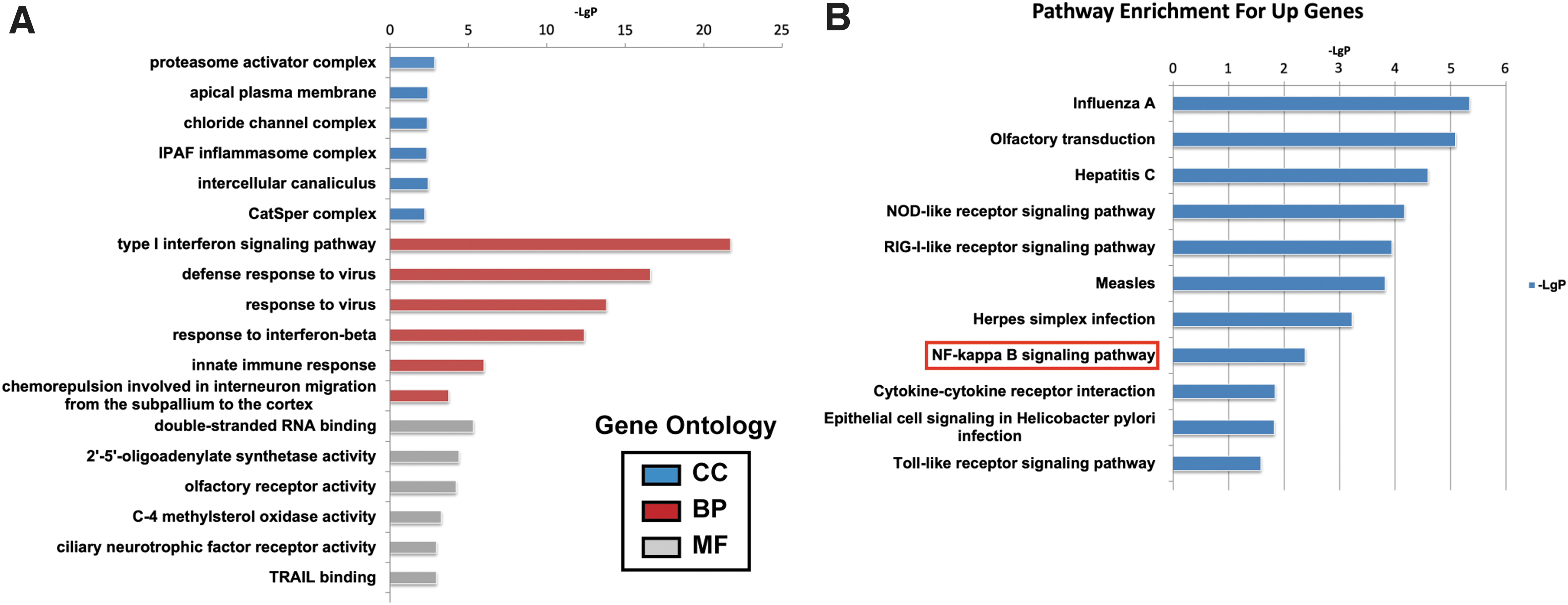
Effects of AGS cell-derived exosomes on hAMSCs gene expression profile.
Analysis of DDX60-derived circular RNAs in human adipose-derived MSCs
The gene chip results showed that after treating human adipose-derived MSCs with AGS-derived exosomes for 24 h, the circular RNAs expressed by human adipose-derived MSCs showed significant differences (Supplementary Table S1). Further screening of differentially expressed circular RNAs in the microarray results by qRT-PCR showed that six circular RNAs were significantly upregulated (Fig. 5A) and three circular RNAs were significantly downregulated (Fig. 5B). Gene microarray results showed that there was a significant difference between the expressions of hsa_circ_0004303 in human adipose-derived MSCs before and after the treatment of AGS cell-derived exosomes (Fig. 5C), and the differential expression of hsa_circ_0004303 had a certain influence on the biological functions of human adipose-derived MSCs.
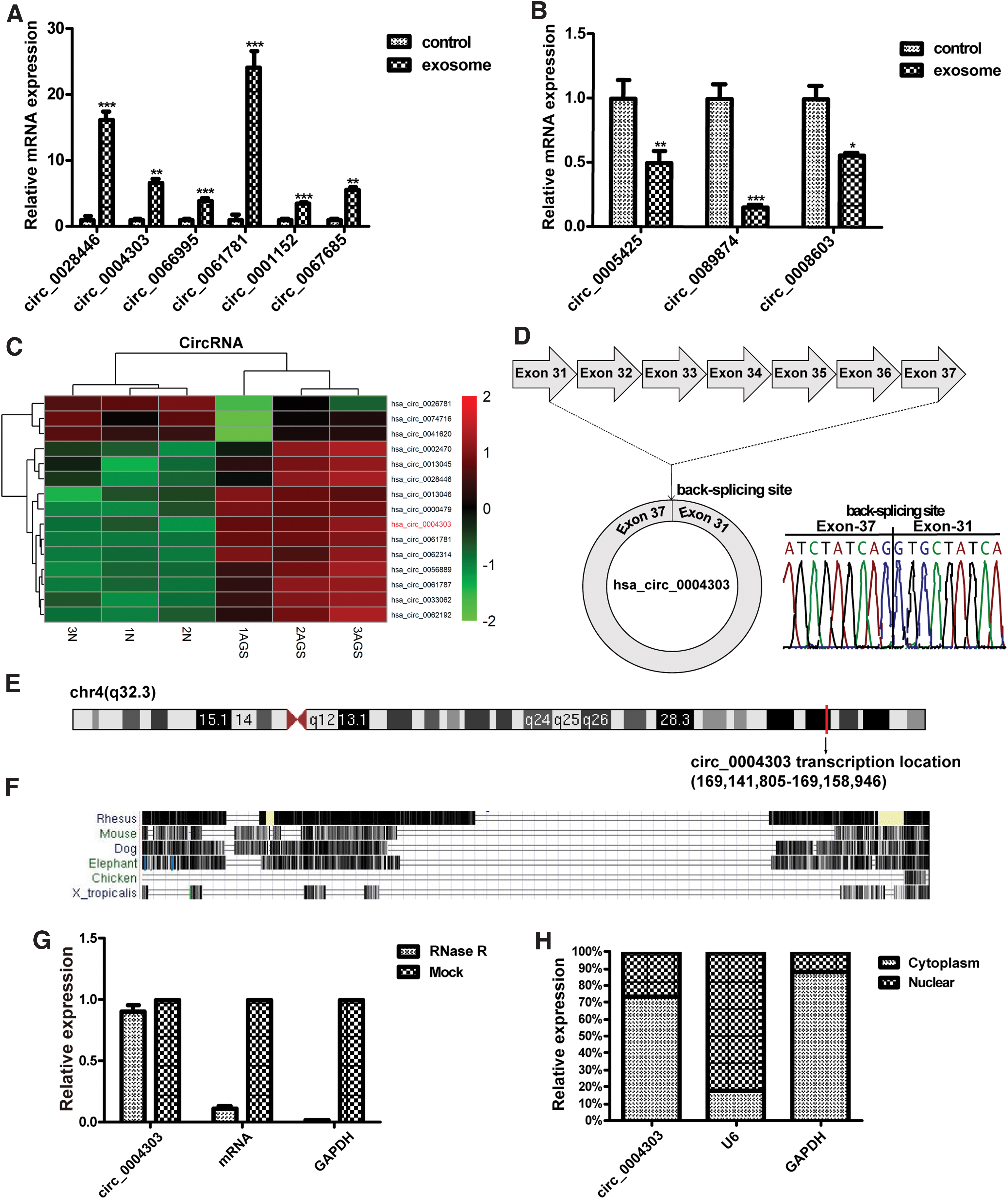
Expression and identification of circ_0004303.
There were also many differentially expressed genes involved in cell migration, immunity, inflammation, cell proliferation, and angiogenesis (Supplementary Fig. S1). The sequence and gene localization of circ_0004303 were obtained from the UCSC database. It was found that circ_0004303 was 875bp in length and comprised exon 31–37 of the DDX60 gene (chr4: 169141805-169158946). The head-to-tail splicing structure of circ_0004303 was verified via Sanger sequencing (Fig. 5D). The UCSC database showed that circ_0004303 is located in the q32.2 region of chromosome 4 (Fig. 5E), and that this gene is not conservative (Fig. 5F). Furthermore, we found that circ_0004303 was less sensitive to RNase R than the linear DDX60 in human adipose-derived MSCs (Fig. 5G). Nuclear cytoplasmic separation experiments showed that circ_0004303 was mainly located in the cytoplasm, which provided an idea for exploring the mechanism of its functions (Fig. 5H).
circ_0004303 regulates the migration and invasion ability of human adipose-derived MSCs
We further verified whether exosomes derived from AGS cells have an effect on the migration and invasion abilities of human adipose-derived MSCs. Results showed that the migration and invasion abilities of human adipose-derived MSCs were enhanced after treating with AGS cell-derived exosome (Fig. 6A, B). To explore the biological functions of circ_0004303, we conducted a series of in vitro experiments. We used siRNAs specifically targeting the circ_0004303 back-splice junction site to knock down circ_0004303, circ_0004303 expressions were significantly downregulated by circ_0004303 siRNAs compared with the control siRNA (designated control) in human adipose-derived MSCs (Fig. 6C). Migration and invasion assays showed that knocking down of circ_0004303 reduced the migration and invasion abilities of human adipose-derived MSCs (Fig. 6D, E). By using a plasmid, we succeeded in overexpressing circ_0004303 in human adipose-derived MSCs (Fig. 6F). Migration and invasion assays showed that the overexpression of circ_0004303 increased the migration and invasion ability of human adipose-derived MSCs (Fig. 6G, H).
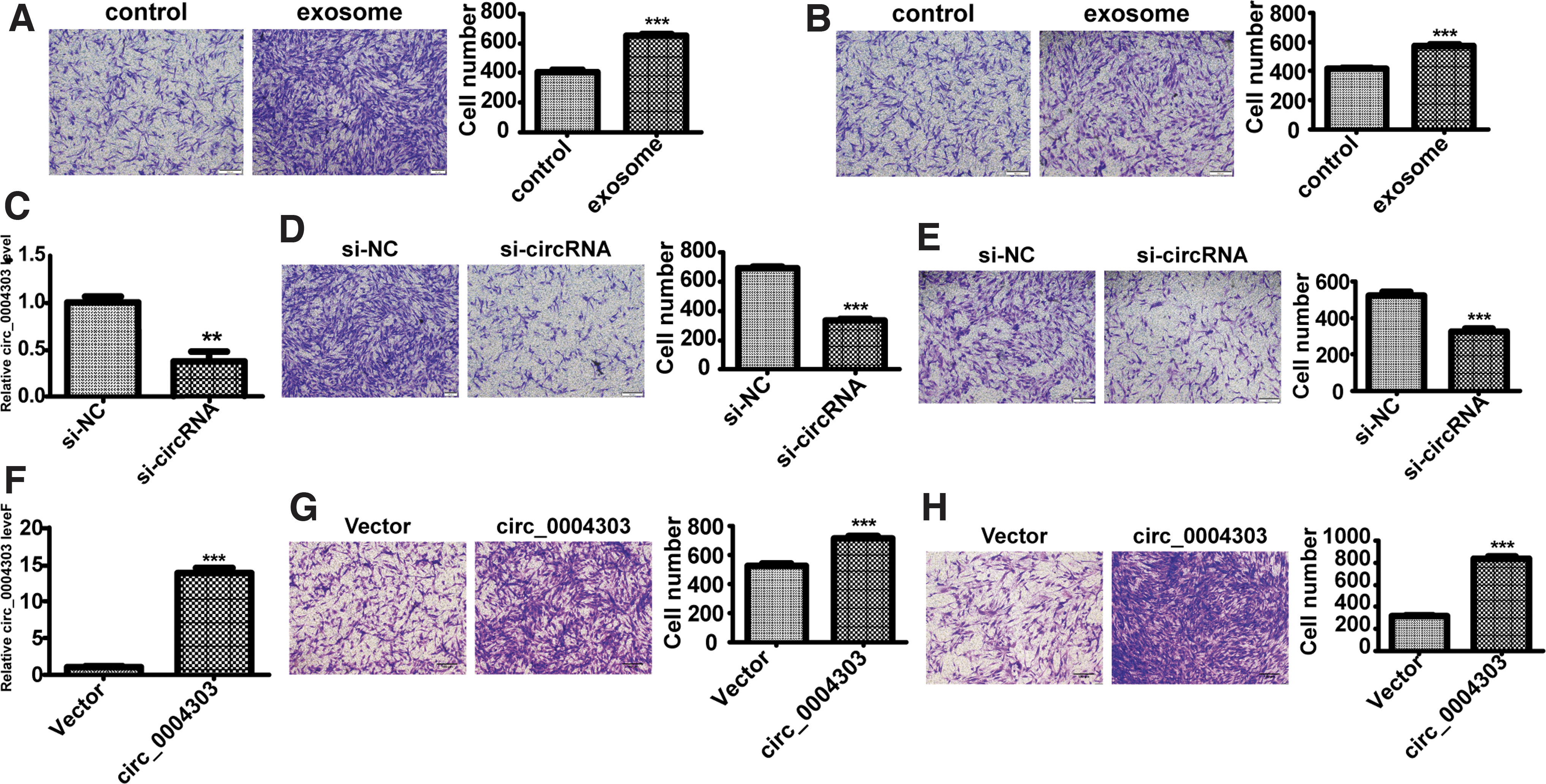
circ_0004303 regulates the migration and invasion ability of hAMSCs.
circ_0004303 regulates the migration and invasion abilities of human adipose-derived MSCs via sponging miR-148a-3p
A growing number of studies have shown that circular RNAs mainly function as miRNA sponges. Therefore, we speculated that circ_0004303 may act as a sponge of certain miRNAs that might play a role in the transformation of N-MSC to T-MSC. Through the Nuclear/Cytosol Fractionation assay, we demonstrated that circ_0004303 was mainly localized in the cytoplasm of human adipose-derived MSCs. Potential targets of circ_0004303 were predicted using the StarBase database, and some miRNAs, including miR-148a-3p, were found to have binding sites in circ_0004303 region (Fig. 7A). To further verify the interaction between miR-148a-3p and circ_0004303, we carried out luciferase reporter assays by transfecting HEK-293T cells with luciferase reporter vectors (containing the wild type or mutant sequence of circ_0004303 target). The Luciferase reporter activity was significantly decreased by the miR-148a-3p mimics in cells transfected with the wild-type sequence compared to those transfected with the mutant sequence (Fig. 7B, C).
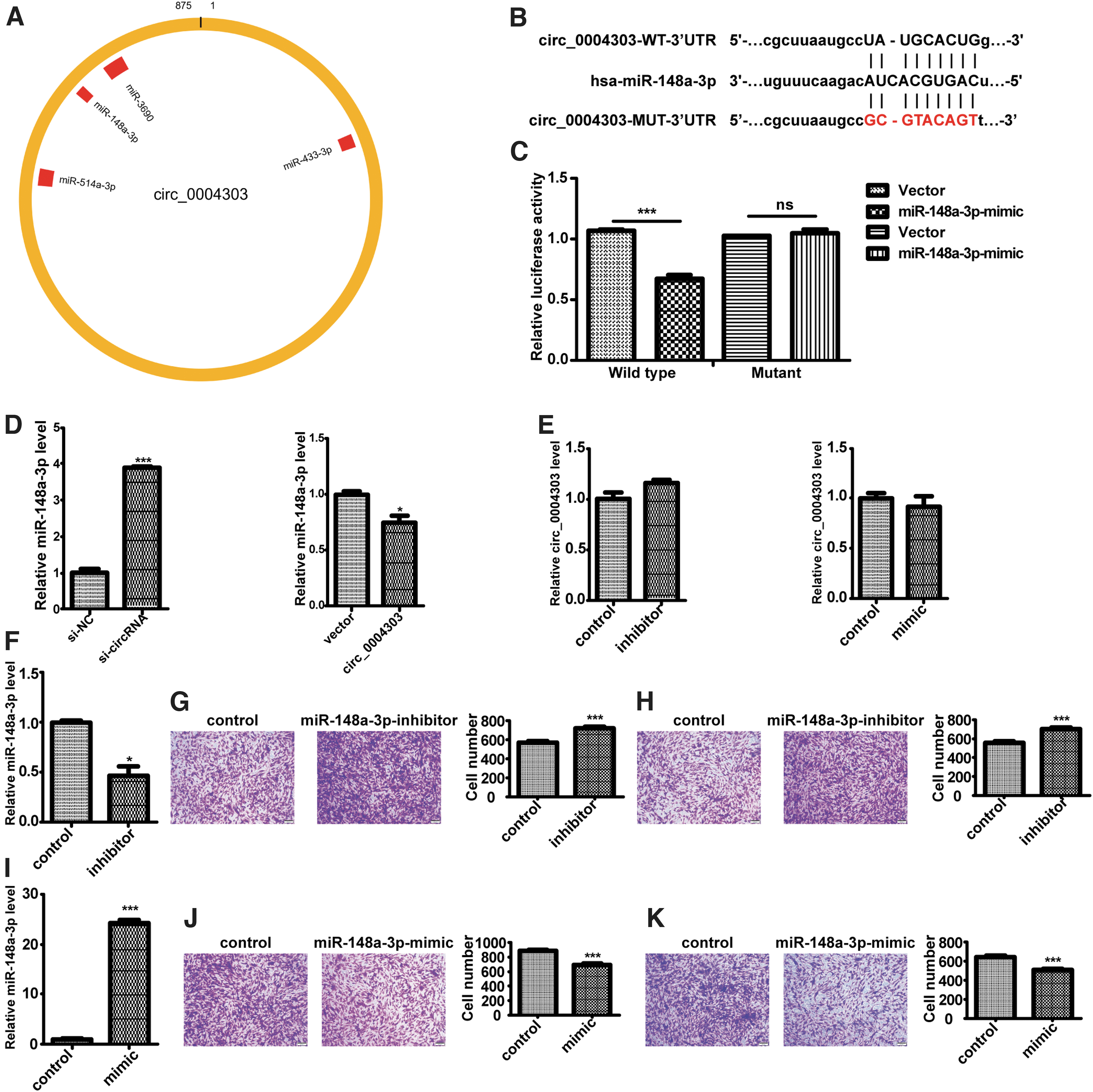
circ_0004303 regulates the migration and invasion ability of hAMSCs via sponging miR-148a-3p.
After knocking down circ_004303, the expression level of miR-148a-3p was significantly upregulated, and after overexpressing circ_004303, the expression level of miR-148a-3p was significantly downregulated (Fig. 7D). However, the expression level of circ_004303 remained unchanged after miR-148a-3p being upregulated or downregulated (Fig. 7E). Then, miR-148a-3p inhibitor was used to knock down the expression of miR-148a-3p (Fig. 7F). Migration and invasion assays showed that knockdown of miR-148a-3p increased the migration and invasion abilities of human adipose-derived MSCs (Fig. 7G, H). MiR-148a-3p was overexpressed using miR-148a-3p mimics (Fig. 7I). Migration and invasion assays showed that the overexpression of miR-148a-3p reduced the migration and invasion abilities of human adipose-derived MSCs (Fig. 7J, K).
Taken together, our results provide evidence that circ_004303 can directly bind to miR-148a-3p in human adipose-derived MSCs, and that miR-148a-3p can regulate the migration and invasion abilities of human adipose-derived MSCs.
circ_0004303 upregulates the level of ALCAM by acting as a miR-148a-3p sponge, and ALCAM is involved in regulating the migration and invasion abilities of human adipose-derived MSCs
Four databases (including miRWalk, TargetScan, miRTarBase, and miRDB) were used to predict the potential targets of miR-148a-3p, and 40 mRNAs were overlapped in these four databases (Fig. 8A). ALCAM has been reported to promote tumor progression [20 –22]. Therefore, we speculated that circ_0004303 mainly acts as a miR-148a-3p sponge to regulate the expression of ALCAM, thereby affecting the progression of gastric cancer. In addition, mRNA and protein levels of ALCAM significantly decreased after circ_0004303 downregulation, and significantly increased after circ_0004303 upregulation (Fig. 8B).
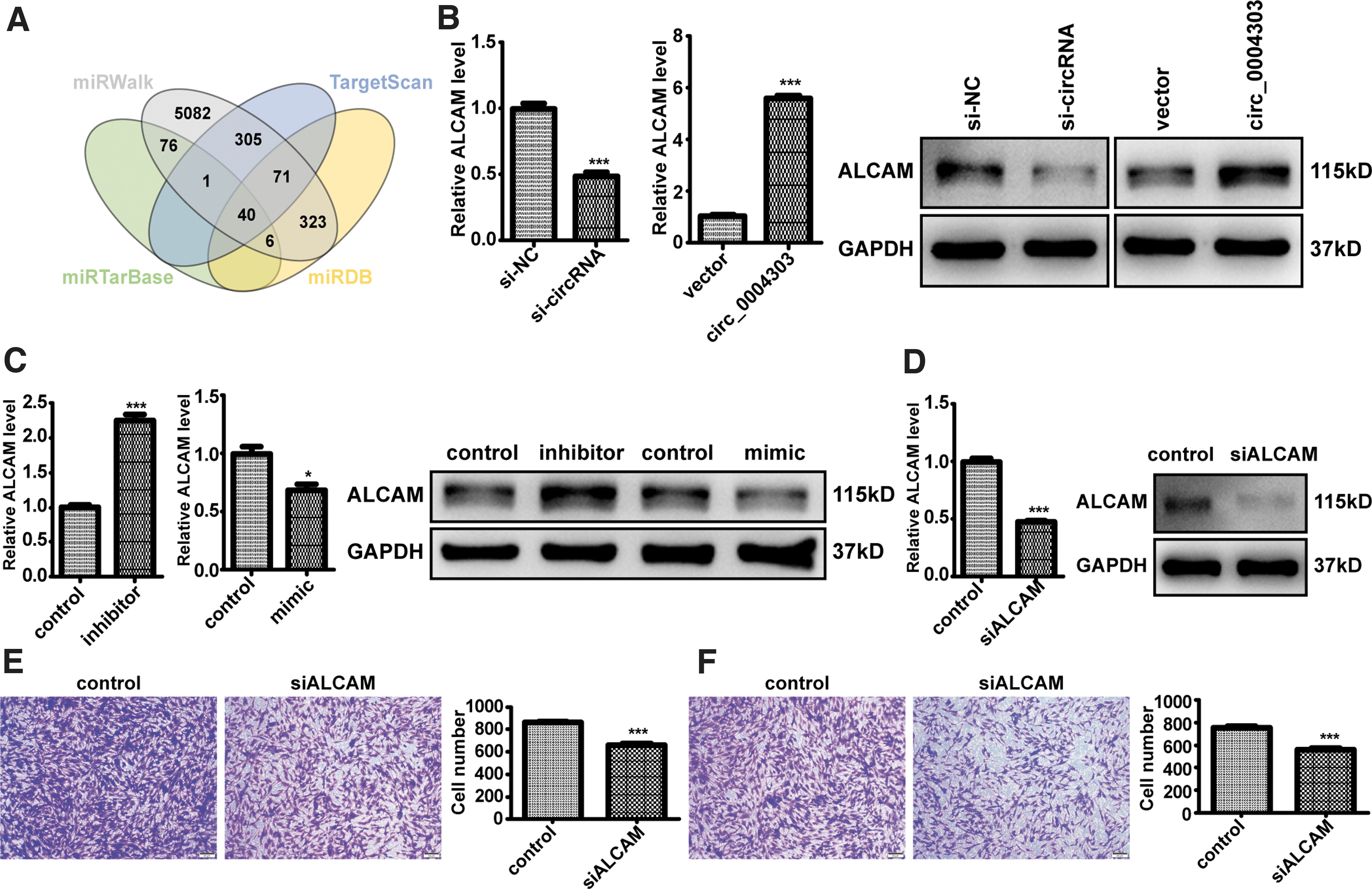
circ_0004303 upregulates the level of ALCAM by acting as a miR-148a-3p sponge, and ALCAM is involved in regulating the migration and invasion ability of hAMSCs.
Moreover, mRNA and protein levels of ALCAM significantly increased after miR-148a-3p downregulation in human adipose-derived MSCs, and significantly decreased after miR-148a-3p upregulation in human adipose-derived MSCs (Fig. 8C). The mRNA and protein levels of ALCAM were significantly reduced after human adipose-derived MSCs were transfected with shRNA specifically targeting ALCAM (Fig. 8D). Migration and invasion assays showed that knockdown of ALCAM reduced the migration and invasion abilities of human adipose-derived MSCs (Figs. 8E, F). In conclusion, our results provide evidence that circ_0004303 upregulates the level of ALCAM by acting as a miR-148a-3p sponge, and ALCAM can regulate the migration and invasion ability of human adipose-derived MSCs.
Discussion
To date, a substantial body of research has shown that exosomes secreted by tumor cells influence the functions of MSCs, but the effects and the mechanisms of such effects vary depending on the sources of the MSCs and the exosomes. Our study found that exosomes of AGS can affect the gene expression of human adipose-derived MSCs and enhance their migration and invasion ability. Results have shown that the expression of some circular RNAs is significantly different after human adipose-derived MSCs being treated with exosomes secreted by AGS for 24 h. Through a series of in vitro experiments, we further identified that circ_0004303 could regulate the migration and invasion abilities of human adipose-derived MSCs. This study enriches the research field of circular RNAs in gastric cancer.
Studies have shown that circular RNAs can play an important role through different modes, such as acting as sponges to absorb miRNA, binding to proteins, translating into polypeptides or proteins, and generating pseudogenes. Competitive endogenous RNA (ceRNA) has many miRNA binding sites and can act as miRNA sponge to regulate the activity of miRNA on its target genes [23]. Circular RNA Cdr1as regulates the stability of miR-7 in neurons, while miR-671 regulates the level of Cdr1as. Cdr1as and miRNAs directly interact with it play an important role in sensorimotor gating and synaptic transmission [24]. circHIPK3 acts as a miR-124 sponge to inhibit miR-124 activity and regulate cell growth [25].
In this study, it was found that circ_0004303 was mainly distributed in the cytoplasm through nucleocytoplasmic separation assay. Database prediction combined with dual luciferase reporter gene assay further confirmed that circ_0004303 acted as a sponge for miR-148a-3p. MiRNA is an important posttranscriptional regulator, which leads to decreased mRNA stability by binding to the target site of mRNA 3′-UTR [26]. By using database prediction combined with literature review, the target gene ALCAM of miR-148a-3p was obtained. These findings indicate that circ_0004303 can act as a ceRNA to regulate ALCAM gene expression as a miR-148a-3p sponge, thereby affecting the biological functions of human adipose-derived MSCs.
ALCAM (CD166) belongs to the immunoglobulin superfamily and consists of five immunoglobulin-like domains [27]. ALCAM is expressed in a variety of cells, such as neurons, fibroblasts, endothelial cells, and so on [28]. ALCAM-mediated cell adhesion is essential in different physiological and pathological processes, especially in terms of leukocyte exudation, immune synapse stabilization, T cell activation and proliferation, and tumor growth and metastasis [29]. The overexpression of ALCAM promotes the progression of malignant mesothelioma, and ALCAM can be used as a potential therapeutic target for malignant mesothelioma [21]. ALCAM can promote the migration, invasion, and metastasis of tumor cells and thus can be used as a marker for the recurrence of endometrial cancer [30].
In the tumor tissue microenvironment, ALCAM plays an integrated role in regulating cell adhesion, growth, invasion, and proteolysis [31]. Knockdown of ALCAM significantly increases the growth and migration ability of gastric epithelial cells HFE145 [32]. High levels of ALCAM on the membrane of gastric cancer cells and in the serum of patients with gastric cancer are signs of poor prognosis in patients with gastric cancer [33]. The level of ALCAM in the serum of patients with gastric cancer is significantly higher than that in the control group, and the level of ALCAM in the serum can be used as a diagnostic indicator for patients with gastric cancer [34]. In this study, we found that circ_0004303 acts as a miR-148a-3p sponge to reduce the inhibition of miR-148a-3p on ALCAM expression. The reduction of ALCAM can reduce the migration and invasion abilities of human adipose-derived MSCs.
Conclusion
In conclusion, our study illustrates the mechanism by which circ_0004303 regulates the biological function of hAMSCs, and that the expression of ALCAM in hAMSCs will affect the migration and invasion ability of hAMSCs and further affects the homing of hAMSCs to tumor tissue. The biological characteristics of MSCs homing to tumors will change and form phenotypes that promote tumor progression.
Availability of Data and Materials
All data generated or analyzed during this study are included either in this article or in the Supplementary Materials and Methods, Tables, Figures, and Figure Legends files.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from The National Key Research and Development Program of China (2016YFA0101000,2016YFA0101003,20 18YFA0109800); CAMS Innovation Fund for Medical Sciences (2017-I2M-3-007, 2017-2017-I2M-3-006); Supported by the 111 Project (B18007) National Natural Science Foundation of China (81672313, 81700782).
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.