
Abstract
Cancer-associated fibroblasts (CAFs) are a key component of the tumor microenvironment and are essential for tumorigenesis and development. Regorafenib is a multikinase inhibitor that targets CAFs and suppresses tumor growth. In this study, we investigated the effects of regorafenib on gastrointestinal CAFs and the underlying molecular mechanisms. First, we established two in vivo tumor models, the cancer cell line HCT116 with and without mesenchymal stem cells (MSCs), and treated them with regorafenib. We found that application of regorafenib potently impaired tumor growth, an effect that was more pronounced in tumors with a high stromal ratio, thus demonstrating that regorafenib can inhibit CAF proliferation and induce CAF apoptosis in vivo. Moreover, we showed that regorafenib affected macrophage infiltration by reducing the proportion of CAFs in tumors. Second, we induced MSCs into CAFs with exosomes to establish an in vitro model. Then, we used 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt and flow cytometry to detect the effects of regorafenib on proliferation and apoptosis of CAFs and western blot to determine the expression level of apoptosis-related proteins. We found that regorafenib inhibited proliferation of CAFs and induced apoptosis in CAFs in vitro. Furthermore, western blot results showed that regorafenib downregulated the expression of B cell lymphoma-2 (Bcl-2) and concurrently upregulated the expression of Bcl-2-associated X (Bax), and regorafenib inhibited the phosphorylation pathway of AKT in CAFs. In conclusion, our results provide a model in which regorafenib induces CAF apoptosis by inhibiting the phosphorylation of AKT and regorafenib affects macrophage infiltration by reducing the proportion of CAFs in tumor tissues.
Introduction
Gastric cancer and colorectal cancer are two particularly pervasive cancers in the world and their morbidity and mortality have been rising rapidly [1,2]. Improvements in surgery and chemotherapy have improved survival in patients with gastrointestinal tumors, but the clinical effect is still unsatisfactory. Therefore, it is imperative to develop effective drugs against gastrointestinal tumors.
Receptor tyrosine kinases (RTKs) are single-pass transmembrane receptors with cytoplasmic tyrosine kinase domains. Abnormal activation of RTKs is one of the driving forces of human carcinogenesis. In the past decades, small-molecule inhibitors targeting RTKs have been developed for treatment of a variety of cancers [3]. Among these inhibitors, regorafenib is a novel oral multikinase inhibitor that directly inhibits tumor growth and indirectly affects stromal regulation and angiogenesis by inhibiting the activity of various tyrosine kinases such as c-kit, VEGFR, PDGFR, and FGFR [4,5].
The effects of regorafenib on stromal cells have been preliminarily described in basic studies [6]. Moreover, the antitumor activity of regorafenib against a range of tumors has been shown in clinical studies. In 2012, regorafenib was approved by the US Food and Drug Administration (FDA) for treatment of patients with colorectal cancer and gastrointestinal stromal tumors. Its effectiveness and safety have been confirmed [7,8].
Later, the antitumor activity of regorafenib was further evaluated in patients with gastric cancer, and the clinical trial result showed that regorafenib potentially suppressed the progression of gastric cancer [9]. Recently, it has been reported that the PD-1 inhibitor and regorafenib have a synergistic clinical effect, but the mechanism remains an enigma.
The tumor microenvironment is a complex network, consisting of cancer-associated fibroblasts (CAFs), immune cells, mesenchymal stem cells (MSCs), and extracellular matrix (ECM), which plays a pivotal role in tumor development [10]. CAFs are a major component of the tumor microenvironment with versatile functions. For instance, CAFs promote tumor progression by secreting a variety of proteins, growth factors, and chemokines [11,12].
Moreover, cross talk between CAFs and immune cells has been reported. For example, immunomodulatory molecules produced by CAFs are critical agents for recruitment of Tregs and tumor-associated macrophages (TAMs) [13]. Notably, when activated, CAFs start to express two CAF-specific proteins, α-smooth muscle actin (α-SMA) and fibroblast activation protein (FAP), which are commonly used as biomarkers for CAFs in basic research [14].
Regorafenib belongs to the multikinase inhibitor family and effectively targets stromal cells. Currently, only sporadic studies have focused on the effects of regorafenib on CAFs. Therefore, to advance our understanding of how regorafenib affects CAFs, we investigated the effects of regorafenib on gastrointestinal CAFs and explored the underlying mechanisms. This study not only provides add-on evidence for regorafenib's effectiveness in treating cancers but also presents new perspectives to develop new therapies.
Materials and Methods
Cell culture
Human adipose-derived MSCs (hAMSCs) were isolated and cultured in accordance with the Declaration of Helsinki. The protocol has been approved by the Ethics Committee of the Institute of Basic Medical Sciences, Chinese Academy of Medical Sciences (Project No. 022-2015) [15]. MSCs were isolated from discarded fat tissues of patients undergoing liposuction in the Department of Plastic Surgery, Peking Union Medical College Hospital [16]. Informed consent was obtained from all patients.
The human gastric cancer cell line, AGS, and human colorectal cancer cell line, HCT116, were purchased from the Cell Resource Center, Institute of Basic Medical Sciences, Chinese Academy of Medical Sciences. AGS cells were cultured in Dulbecco's modified Eagle's medium (DMEM)/F12 containing 10% fetal bovine serum (FBS). HCT116 cells were cultured in high-glucose DMEM containing 10% FBS.
Exosome extraction
Exosome extraction was performed as described previously [17]. Briefly, tumor cells were incubated in serum-free DMEM for 36 h. The supernatant was collected and centrifuged at 3,000 rpm for 30 min to remove dead cells. The supernatant was transferred to 100,000-Mw ultrafiltration tubes and exosomes were extracted by an ultrafiltration method. When the volume of the supernatant was reduced to 2 mL, the supernatant was washed with PBS until the liquid became colorless. Finally, the supernatant was filtered with a 0.22-μm filter and stored at −80°C.
Exosome observation by transmission electron microscopy
The purified exosomes were diluted and dropped on a copper net for 5 min. They were dried and redyed with 3% aqueous phosphotungstic acid for 2 min. Exosomes were observed under a transmission electron microscope.
Western blot analysis
The primary antibodies used were as follows: α-SMA (Cell Signaling Technology, Beverly, MA), FAPA (Cell Signaling Technology), GAPDH (Cell Signaling Technology), HSP70 (Abcam, Waltham, MA), HSP90 (Abcam), CD63 (Cell Signaling Technology), Bax (Cell Signaling Technology), Bcl-2 (Cell Signaling Technology), AKT (Cell Signaling Technology), and p-AKT (Cell Signaling Technology). The secondary antibodies were purchased from NeoBioscience (Shenzhen, China).
Cell viability assay
The cells were cultured in 96-well plates (cell density, 5 × 103 cells per well) containing DMEM plus 10% FBS. After adherence, the cells were treated with different concentrations of regorafenib for 1–5 days. The cells were washed with PBS, and MTS reagent (Promega) was applied. Following incubation at 37°C for 1–2 h, absorbance at 490 nm was measured every 24 h to establish the growth curve.
Cell cycle and apoptosis assays
For the cell cycle assay, CAFs were treated with regorafenib at the indicated concentrations for 24 h. The PI/RNase staining buffer (BD Biosciences, Heidelberg, Germany) was used to analyze the cell cycle distribution. For the apoptosis assay, CAFs were treated with regorafenib at the indicated concentrations for 24 h. After staining with Annexin V-FITC and propidium iodide, the cells were detected using a BD Accuri C6 flow cytometer.
Animal models
All nude mice were purchased from the Sibf Biotechnology Co., Ltd., (Beijing, China) and maintained under specific pathogen-free conditions. Both animal usage and experimental procedure were approved by the Institutional Animal Care and Use Committee of MDL (approval ID: MDL2021-11-16-01). Tumor models were generated by subcutaneously implanting HCT116 cells (5 × 106; HCT116 tumors) and HCT116 cells with MSCs (HCT116 cells: 5 × 106, MSCs: 1 × 106; HCT116+MSC tumors) into nude mice.
Mice with tumor volumes of 100–200 mm3 were divided into the following four groups: (i) mice with HCT116 tumors that were treated daily with PBS through oral gavage (n = 10); (ii) mice with HCT116+MSC tumors that were treated daily with PBS through oral gavage (n = 10); (iii) mice with HCT116 tumors that were treated daily with 7.5 mg/kg regorafenib through oral gavage (n = 10); and (iv) mice with HCT116+MSC tumors that were treated daily with 7.5 mg/kg regorafenib through oral gavage (n = 10).
The tumor volume was recorded every 4 days, which was evaluated by caliper and calculated by the following formula: tumor volume = length × width2 × 0.5. After 20 days of drug treatment, the nude mice were sacrificed and tumors were separated for staining analysis.
Hematoxylin and eosin, immunohistochemistry, and TUNEL assays
Some of the tumor sections were stained with hematoxylin and eosin and others were stained with α-SMA, FAP, F4/80, Ki67, and CD31 for immunohistochemical analyses, as described previously [18]. Images were taken using a microscope (Olympus Optical Co., Ltd., Tokyo, Japan). Apoptosis cells in tissue sections were detected by TUNEL assay according to the manufacturer's instructions (apoptosis detection kit).
Finally, ImageJ software (National Institutes of Health, Bethesda, MD) was used for quantitative analysis.
Immunofluorescence assay
The sections were stained with α-SMA, β-catenin, and Ki67, and apoptotic cells were identified for immunofluorescence analyses, as described previously [19]. Fluorescent images were observed under a fluorescence microscope (Olympus Optical Co., Ltd.).
Statistical analysis
Data are presented as mean ± standard deviation (SD). Each experiment was repeated at least three times. Two-tailed Student's t-tests were used to assess the significance of differences between the two groups. Statistical analysis was performed using GraphPad Prism 7.0 (GraphPad Software, Inc., San Diego, CA). A value of P < 0.05 was considered to be statistically significant.
Results
Regorafenib inhibits tumor growth in nude mice
To study the therapeutic effect of regorafenib on stromal cells, we established HCT116 and HCT116+MSC-inoculated animal models (Fig. 1A). The growth curve showed that tumor size in the two tumor models increased over time before regorafenib treatment. The tumor growth rate was significantly higher in the HCT116+MSC group than in the HCT116 group (P < 0.05; Fig. 1B). Treatment was started when the tumor volume reached 100–200 mm3.
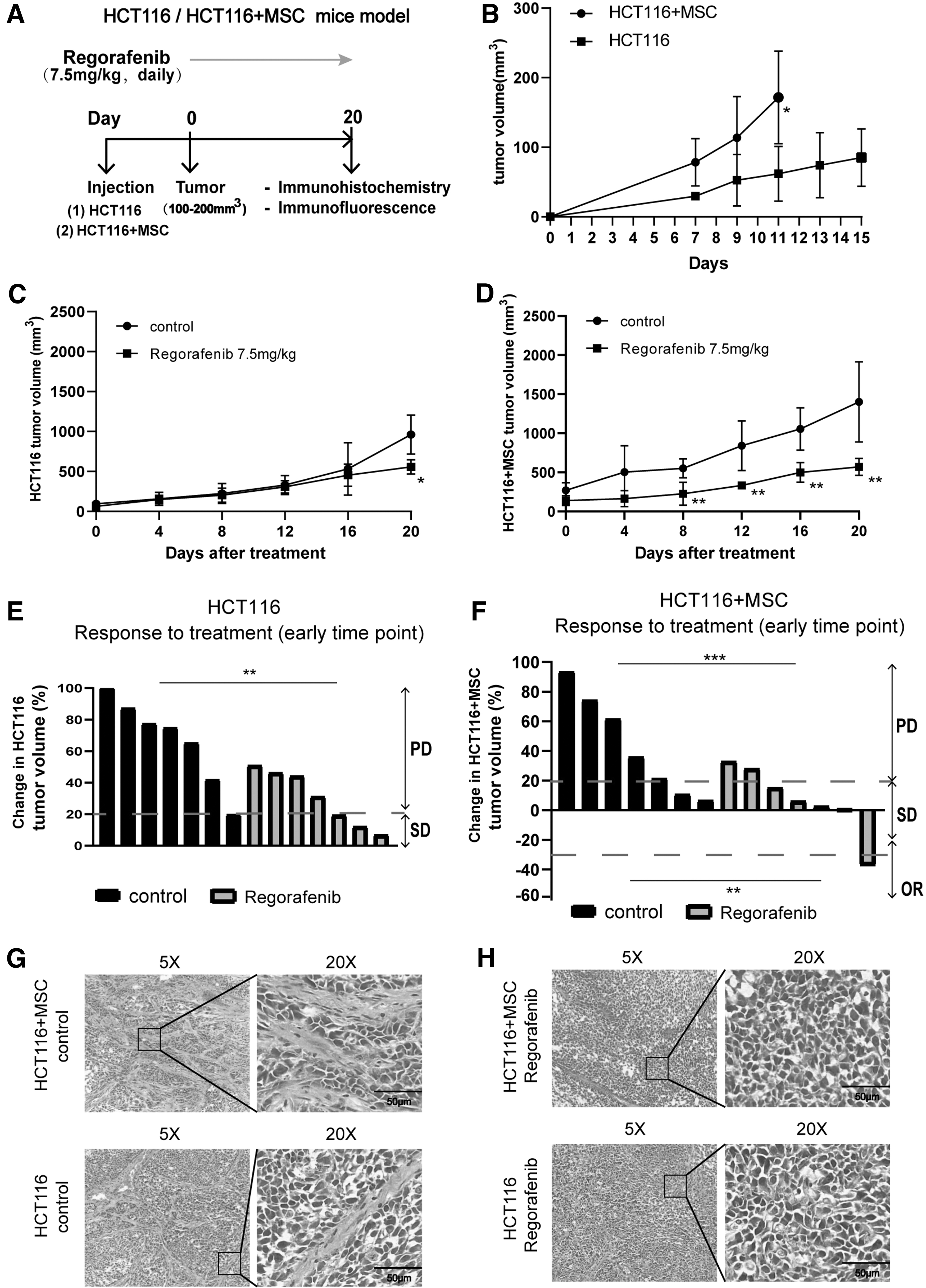
Regorafenib inhibits tumor growth in nude mice.
After 20 days of regorafenib treatment, compared with their respective control groups, the tumor size in the HCT116 treatment group decreased by 42.0% (P < 0.05, Fig. 1C) and that in the HCT116+MSC treatment group decreased by 59.3% (P < 0.01; Fig. 1D). The objective response time was the time point at which there was a significant difference in the change in tumor volume in the regorafenib treatment groups compared with their respective control groups.
Compared with their respective control groups, the HCT116 treatment group did not show significant tumor volume reduction until day 20 (P < 0.05, Fig. 1C), while the HCT116+MSC treatment group began to show significant reduction as early as day 8 (P < 0.01, Fig. 1D). Tumor volume growth of 20% or more was defined as progressive disease (PD). A reduction of 30% or more was defined as an objective response. Any change less than 20% increase and 30% reduction was classified as stable disease [20]. Compared with their respective control groups, both treatment groups exhibited a reduced PD rate at the early time point (Fig. 1E, F, and Supplementary Table S1).
Notably, the early objective response rate (ORR) in the HCT116+MSC treatment group was higher than that in the HCT116 treatment group (P < 0.05; Supplementary Table S1). Meanwhile, median tumor volume was equal at randomization (Supplementary Fig. S1C, D), and body weight of mice was maintained at stable levels (18–22 g) (Supplementary Fig. S1A, B). Overall, regorafenib exhibited antitumor activity in both groups of mice.
However, the HCT116+MSC treatment group required shorter treatment response time and improved ORR relative to the HCT116 treatment group at the early time point. In addition, H&E staining showed that the HCT116+MSC control group had more active stromal cells than the HCT116 control group (Fig. 1G). Taken together, these results indicated that regorafenib was more effective in tumors with a high stromal ratio. It is reasonable to speculate that regorafenib suppresses tumor growth by acting on CAFs.
Effects of regorafenib on cell proliferation and apoptosis in vivo
We further explored the effects of regorafenib in vivo using the previously constructed HCT116 and HCT116+MSC tumor models. The HCT116+MSC control group had a higher proliferation rate (Fig. 2A, B) and a lower apoptosis rate (Fig. 2C, D) than the HCT116 control group. After regorafenib treatment, proliferation ratios of both HCT116 and HCT116+MSC groups decreased (P < 0.05; Fig. 2A, B), whereas apoptosis ratios of both HCT116 and HCT116+MSC groups increased (P < 0.05; Fig. 2C, D).
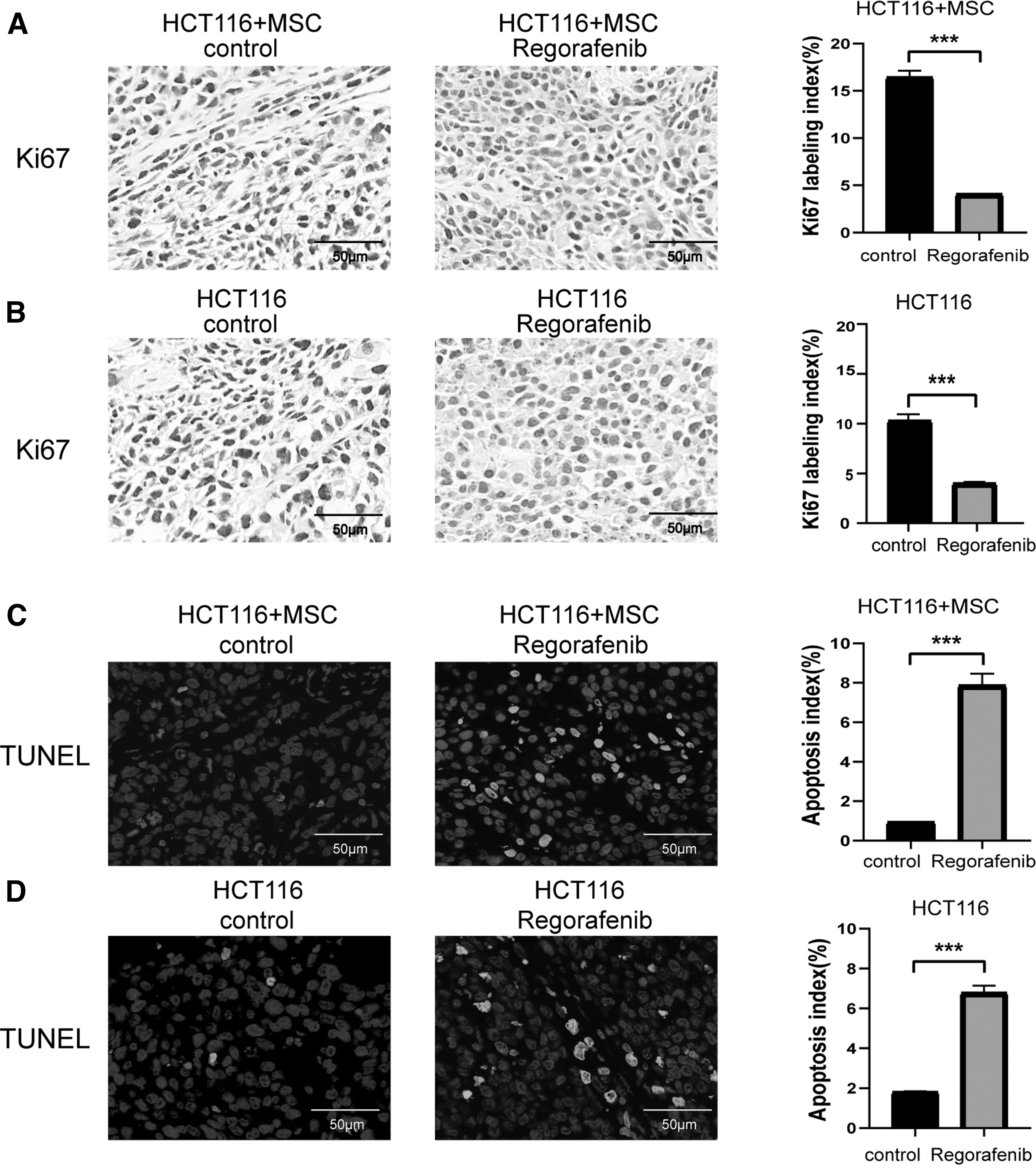
Effects of regorafenib on cell proliferation and apoptosis in vivo. Tumors extracted from mice on day 20 of treatment.
We next investigated the effects of regorafenib on proliferation and apoptosis of stromal cells and tumor epithelial cells by double immunofluorescence staining for α-SMA/Ki67, β-catenin/Ki67, α-SMA/TUNEL, and β-catenin/TUNEL (Fig. 3A–D). The results showed that regorafenib treatment decreased the number of proliferating stromal cells and tumor epithelial cells and increased the number of apoptotic stromal cells and tumor epithelial cells (P < 0.05, Fig. 3E–H).
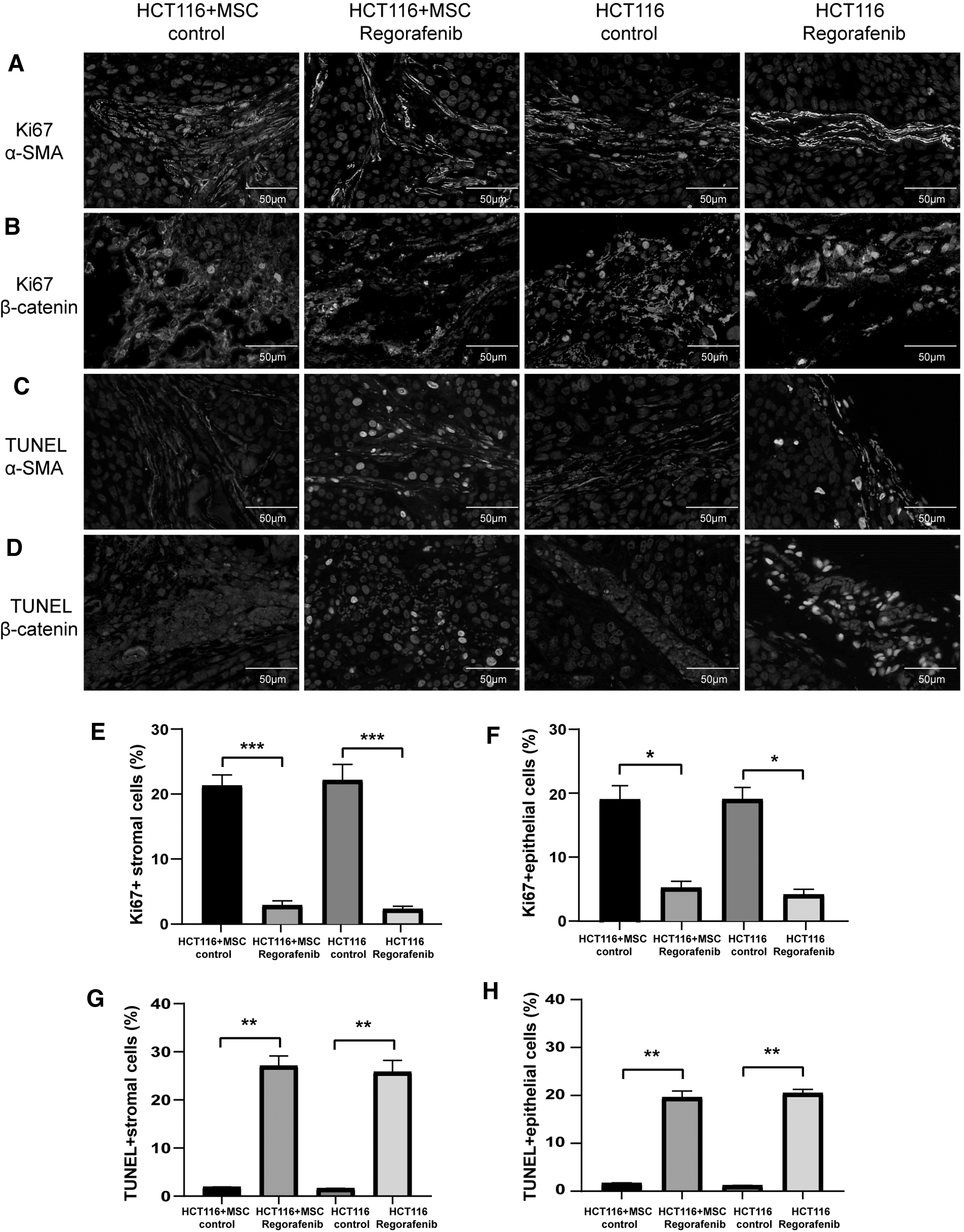
Effects of regorafenib on proliferation and apoptosis of stromal cells and tumor epithelial cells in vivo. Expression of
These results indicated that regorafenib inhibited CAF proliferation and induced CAF apoptosis in vivo, and regorafenib inhibited tumor growth by targeting both tumor cells and stromal cells.
Regorafenib inhibits proliferation of gastrointestinal CAFs in vitro
We further studied the effects of regorafenib on CAFs in vitro. Previously, we have reported that exosomes secreted by tumor cells promoted differentiation of MSCs into CAFs [21]. Because MSCs were more accessible than primary CAFs from tumors, CAFs differentiated from MSCs by tumor exosomes were used in this study as in vitro models. Identification of hAMSCs and differentiation of MSCs into CAFs by tumor exosomes are shown in Supplementary Figs. S2 and S3.
First, AGS and HCT116 cells were treated with a series of concentrations of regorafenib (1, 5, 10, and 20 μM) for 1–5 days. Next, the MTS assay was used to assess the cell viability of gastrointestinal tumors. We selected the most effective concentration of regorafenib to treat tumor cells (Fig. 4A, B).
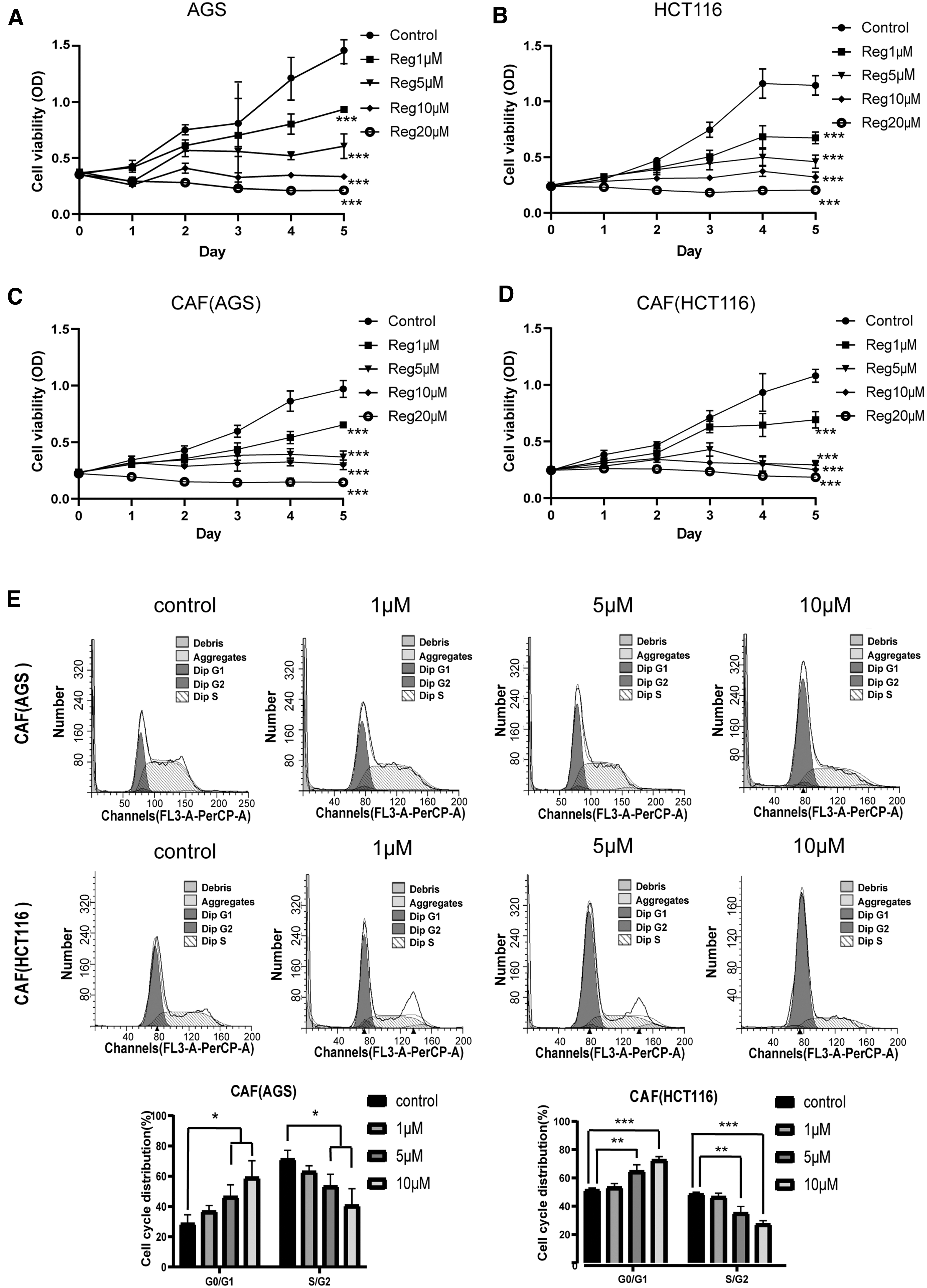
Regorafenib inhibits proliferation of gastrointestinal CAFs in vitro. MTS detection of
Following the treatment, we examined the effect of regorafenib on proliferation of gastrointestinal CAFs with reference to the concentration of regorafenib acting on tumor cells. Similar to the effect of regorafenib on tumor cells, regorafenib significantly inhibited proliferation of gastrointestinal CAFs in a dose- and time-dependent manner (P < 0.001, Fig. 4C, D).
Next, we explored the effect of regorafenib on the cell cycle of CAFs. Flow cytometry results showed that regorafenib administration into gastrointestinal CAFs induced G0/G1 phase arrest that was accompanied by a decreased number of cells in the S/G2 phase (Fig. 4E).
Compared with the control group, the cell cycle progression of HCT116-related CAFs was significantly inhibited by 5 μM regorafenib (P < 0.01). The inhibition was extremely significant when regorafenib concentration was 10 μM (P < 0.001). Similarly, cell cycle progression of AGS-related CAFs was also inhibited by 5 μM or 10 μM regorafenib (P < 0.05).
Collectively, these results confirmed that regorafenib inhibited proliferation of gastrointestinal CAFs.
Regorafenib induces apoptosis of gastrointestinal CAFs in vitro
Gastrointestinal CAFs were treated with different concentrations of regorafenib (1, 5, and 10 μM) for 24 h. Subsequently, we used flow cytometry to examine the effect of regorafenib on gastrointestinal CAF apoptosis (Fig. 5A, B). With the increase of regorafenib concentration, the apoptosis rates of gastrointestinal CAFs gradually increased.
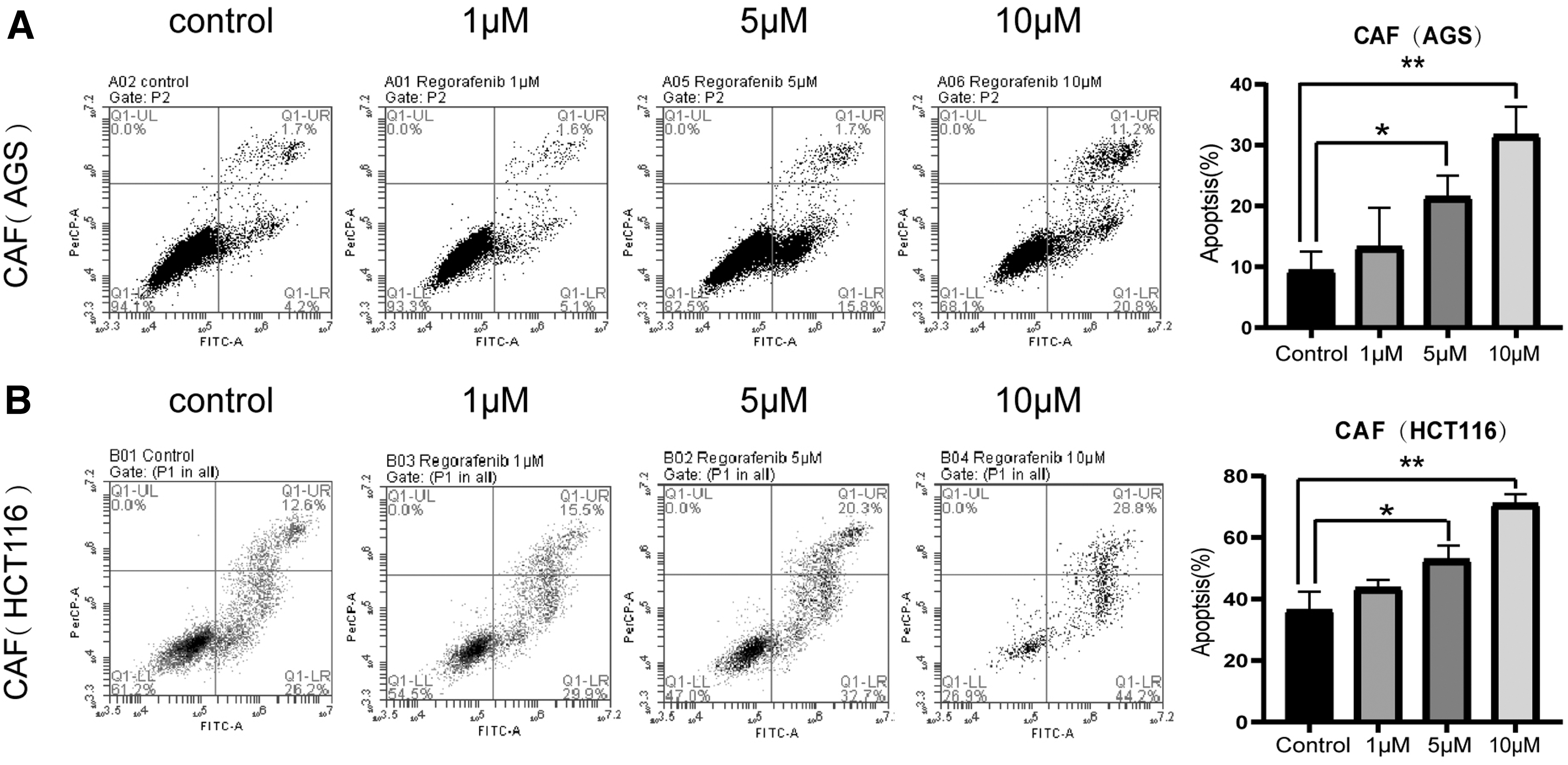
Regorafenib induces apoptosis of gastrointestinal CAFs in vitro. The ratio of apoptotic cells was measured in
When regorafenib concentration was 10 μM, apoptosis was significantly induced (P < 0.01). The results showed that regorafenib induced apoptosis of gastrointestinal CAFs in a dose-dependent manner.
Regorafenib induces apoptosis of gastrointestinal CAFs by inhibiting the AKT phosphorylation signaling pathway
Having demonstrated that regorafenib effectively induced the apoptosis of gastrointestinal CAFs, we set out to explore the molecular mechanism underlying this effect. First, we treated gastrointestinal CAFs with different concentrations of regorafenib. Next, we used western blot to detect the expression of apoptosis-related proteins and found that as the regorafenib concentration increased, the protein level of Bcl-2 decreased and the protein level of Bax was upregulated (P < 0.05, Fig. 6A).
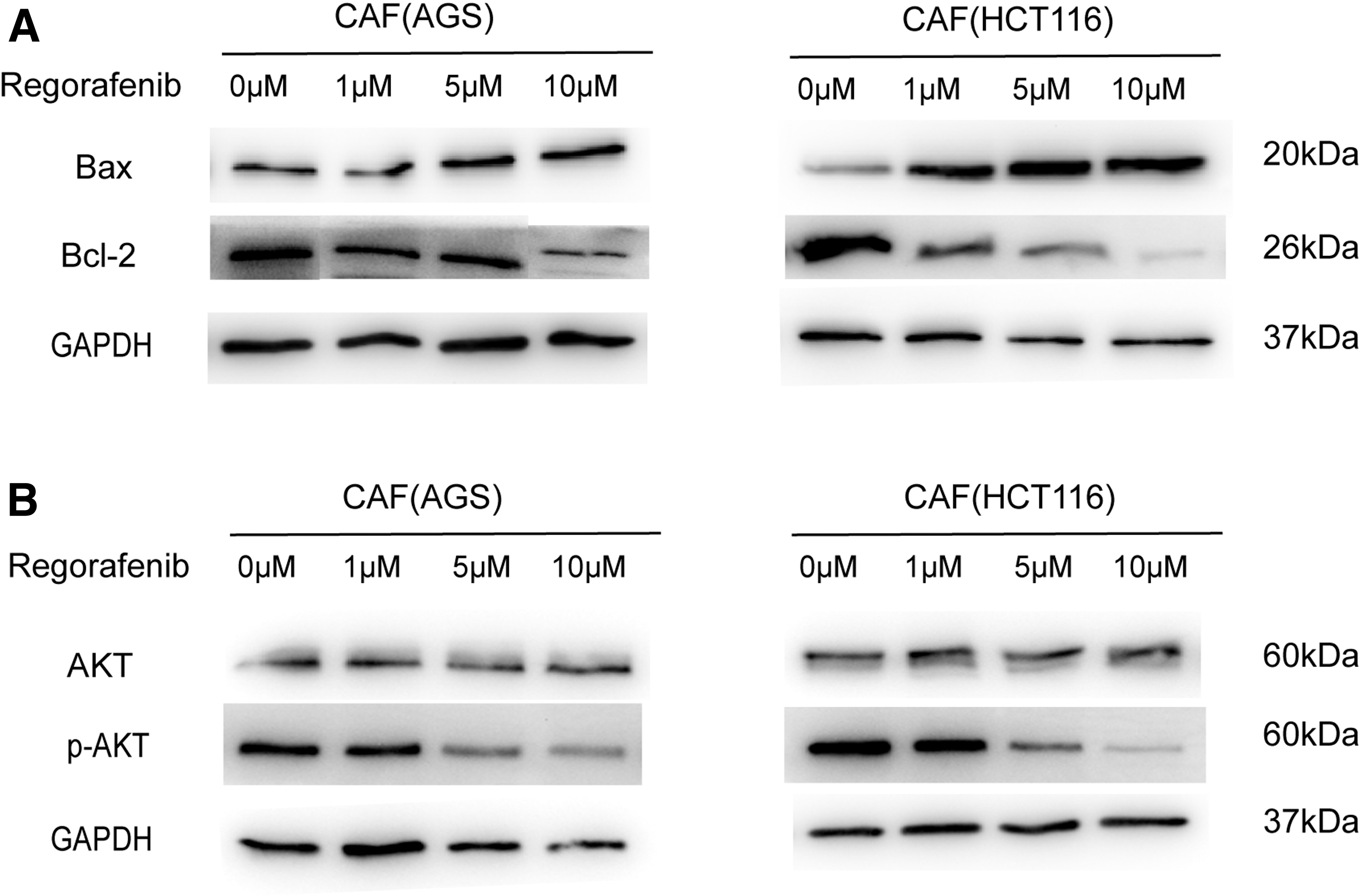
Regorafenib affects the AKT phosphorylation signaling pathway.
Since the PI3K/AKT signaling pathway plays an important role in regulating cell proliferation, apoptosis, and cell cycle [22,23], we detected whether regorafenib treatment could regulate the activation of PI3K/AKT. We found that AKT phosphorylation was significantly reduced after regorafenib treatment (P < 0.05, Fig. 6B).
Therefore, we concluded that regorafenib induced CAF apoptosis by inhibiting AKT phosphorylation and regulating the expression of apoptosis-related proteins.
Effects of regorafenib on stromal cells and macrophage infiltration in the tumor models
Since stromal cells and macrophage infiltration are closely related to tumorigenesis, we assessed the effects of regorafenib on these two modalities by immunohistochemistry analysis. Stromal cells were assessed by α-SMA staining of tumor tissues. The staining showed that stromal cells were more abundant in the HCT116+MSC control group compared with the HCT116 control group (Figs. 1G and 7A, B).
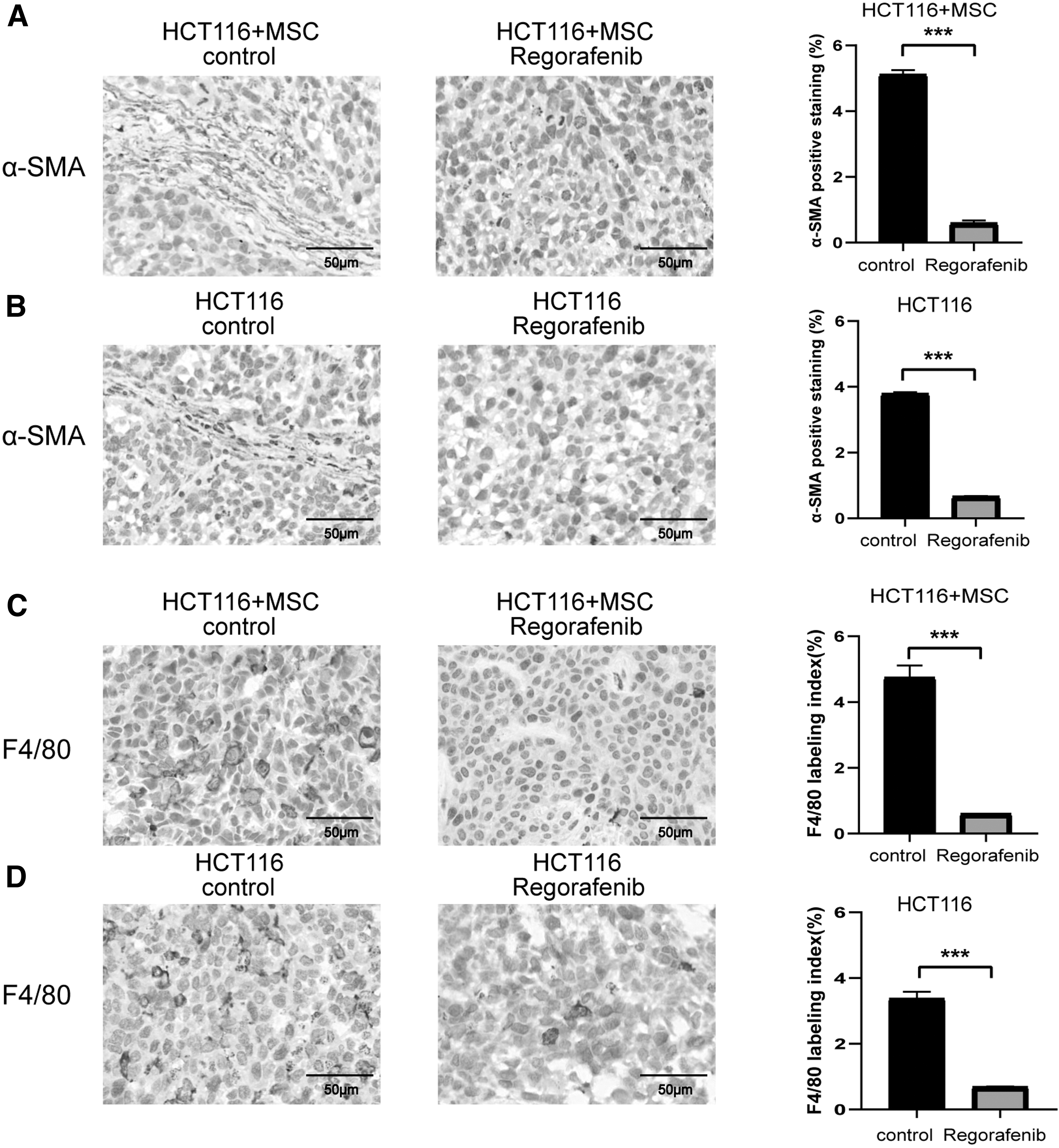
Effects of regorafenib on stromal cells and macrophage infiltration in vivo.
After regorafenib treatment, α-SMA-positive areas were reduced in both HCT116 and HCT116+MSC groups (Figs. 1H and 7A, B). We further verified by FAP staining of tumor tissues that the results were consistent with that of α-SMA staining (Supplementary Fig. S5). Next, by measuring f4/80-positive areas, we observed that the HCT116+MSC control group had more infiltrated macrophages than the HCT116 control group (Fig. 7C, D).
After regorafenib treatment, the staining showed that the infiltrated macrophage proportions in both groups were significantly reduced (Fig. 7C, D). In sum, the above results together with the report that stromal cells could promote macrophage infiltration in tumors [24] demonstrated that regorafenib reduced the proportion of stromal cells and thus affected macrophage infiltration in the tumor models.
Discussion
The tumor microenvironment is one of the major factors affecting tumor progression [25]. It mainly consists of CAFs, smooth muscle cells, endothelial cells, inflammatory cells, and ECM [26]. CAFs are the most prominent cell type within the tumor stroma and can originate from normal fibroblasts, MSCs, and perivascular cells near the tumor site. CAFs play an active role in the progression of cancer through promoting tumor proliferation, regulating tumor angiogenesis, and contributing to tumor chemotherapy resistance [27 –29].
In addition, CAFs have been implicated in tumor immunity. CAFs express a series of immunomodulatory molecules and recruit immunosuppressive cells to mediate tumor immune escape. For example, CAFs are involved in the recruitment and activation of regulatory T cells (Tregs) by secreting regulatory agents such as CXCL12, CD40, and CD73 [30].
In addition, CAFs express complement C5, TGF-β, and other molecules to recruit TAMs and myeloid inhibitory cells (MDSCs), thereby inhibiting T cell activation and promoting tumor progression [31,32]. Therefore, development of CAF inhibitors is key to improving the efficacy of tumor therapies.
Regorafenib is a novel multitarget inhibitor [33 –35]. Through phase I, II, and III clinical trials, the antitumor effects of regorafenib have been demonstrated in a variety of cancers [36 –38]. However, inhibition of CAFs by regorafenib has not been substantiated. In this light, we investigated the effects of regorafenib on gastrointestinal CAFs.
In HCT116 and HCT116+MSC tumor models, regorafenib was demonstrated to have good antitumor activity. Immunofluorescence results showed that regorafenib reduced the number of proliferating stromal cells and tumor epithelial cells, but increased the number of apoptotic stromal cells and tumor epithelial cells. In addition, regorafenib also inhibited tumor angiogenesis (Supplement Fig. S4A, B).
Importantly, despite the existence of the same cell line, regorafenib had different inhibitory effects on HCT116 and HCT116+MSC tumors. Regorafenib was more effective on HCT116+MSC tumors that have a high proportion of stroma than HCT116 tumors. Echoing the effects of regorafenib in vivo, regorafenib had significant inhibitory effects on gastrointestinal CAFs in vitro. Regorafenib inhibited CAF proliferation and induced CAF apoptosis.
Taken together, these results unambiguously demonstrate that regorafenib suppresses tumor growth by inhibiting proliferation of tumor cells and CAFs and concurrently inducing apoptosis.
We further explored the molecular mechanism of regorafenib inducing gastrointestinal CAF apoptosis. We found that regorafenib induced the apoptosis of gastrointestinal CAFs by inhibiting AKT phosphorylation, which decreased the expression of Bcl-2, but increased the expression of Bax.
Compared with conventional therapies, immune checkpoint inhibitors are more effective in improving patient prognosis, but the clinical response rate is relatively low [39]. At present, clinical trials involving mainstream immune checkpoint therapy, including anti-PD-L1, show that the efficacy of immunotherapy alone is relatively limited in many tumor patients [40]. Previous studies have shown that the combination of immune checkpoint inhibitors with regorafenib exhibited more robust antitumor efficacy [41].
These results suggest that regorafenib may influence the tumor microenvironment by inhibiting CAFs, thereby enhancing the efficacy of immune checkpoint therapy. This study showed that regorafenib reduced the proportion of CAFs in tumor tissues and thus affected macrophage infiltration. Therefore, the combination of immune checkpoint inhibitors with regorafenib holds promise for clinical application in tumor treatment.
Conclusions
In conclusion, our study demonstrates that regorafenib is a potent inhibitor for gastrointestinal CAFs through inhibiting CAF proliferation and inducing CAF apoptosis. Moreover, we show that the proapoptotic action of regorafenib on CAFs results from inhibition of AKT phosphorylation. In addition, we find that regorafenib reduces the proportion of CAFs in tumor tissues, thereby affecting macrophage infiltration.
Collectively, this study provides a new experimental basis for the clinical application of regorafenib as a treatment strategy for gastrointestinal tumors.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the National Key R&D Program of China (2018YFA0109800); National Natural Science Foundation of China (81971324, 81902848); National Key R&D Program of China (2016YFA0101000, 2016YFA0101003, 2020YFA0113000, and 2020YFA0113001); Key R&D Project of Guangdong Province (2021B0909060001); 2021 General Research Institute for Nonferrous Metals “Unveiling and Commanding” Project (2021JC0103); and a 111 Project (B18007).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.