
Abstract
Neural progenitor cells are self-renewable, proliferative, and multipotent cell populations that generate diverse types of neurons and glia to build the nervous system. Transcription factors play critical roles in regulating various cellular processes; however, the transcription factors that regulate the development of neural progenitors are yet to be identified. In the present study, we demonstrated that zebrafish etv5a is expressed in the neural progenitor cells of the neuroectoderm. Downregulation of endogenous Etv5a function by etv5a morpholino or an etv5a dominant-negative variant increased the proliferation of sox2-positive neural progenitor cells, accompanied by inhibition of neurogenesis and gliogenesis. These phenotypes in Etv5a-depleted embryos could be rescued by a co-injection with etv5a cRNA. Etv5a overexpression reduced sox2 expression. Direct binding of Etv5a to the regulatory elements of sox2 was affirmed by chromatin immunoprecipitation. These data revealed that Etv5a directly suppressed sox2 expression to reduce the proliferation of neural progenitor cells. In addition, the expression of foxm1, a putative target gene of Etv5a and a direct upstream transcription factor of sox2, was upregulated in Etv5a-deficient embryos. Moreover, the suppression of Foxm1 function by the foxm1 dominant-negative construct nullified the phenotype of upregulated sox2 expression caused by Etv5a deficiency. Overall, our results indicated that Etv5a regulates the expression of sox2 via direct binding to the sox2 promoter and indirect regulation by inhibiting foxm1 expression. Hence, we revealed the role of Etv5a in the transcriptional hierarchy that regulates the proliferation of neural progenitor cells.
Introduction
Neural progenitor cells are self-renewing, proliferative, and multipotent cells that give rise to neuronal and glial derivatives of the nervous system. These cells are generated from the neuroectoderm, which is derived from the ectoderm during early embryogenesis. The maintenance, proliferation, and differentiation of neural progenitor cells must be tightly controlled by environmental signals and intrinsic regulators.
The dysregulated function of these factors results in the defective development of neural progenitors, leading to neurological disorders and tumors of the nervous system. Signaling pathways, such as bone morphogenetic protein (BMP), Wnt, fibroblast growth factor (FGF), Sonic hedgehog (SHH), and Notch, are major players in regulating neural progenitor development. Further, their downstream transcription factors serve as convergence points of cell signaling by directly binding to the promoters or enhancers of target genes and activating or repressing DNA transcription.
SOX2 is a core transcription factor that is involved in the induction, maintenance, and differentiation of pluripotent embryonic stem cells [1 –3]. SOX2 also regulates neuroectoderm specification and governs the maintenance, self-renewal, and proliferation of neural progenitor cells [reviewed in Ref. (4)]. Neural progenitors differentiate into neuronal precursors that are regulated by transcription factors, such as hairy and enhancer of split-1 (HES1) and HES5.
These transcription factors regulate self-renewal and inhibit neuronal differentiation by repressing the expression of proneural genes such as achaete-scute family bHLH transcription factor 1 (ASCL1), neurogenin 1 (NEUROG1), and NEUROG2. In contrast, oligodendrocyte differentiation is regulated by oligodendrocyte transcription factor 1 (OLIG1) and OLIG2 [5,6].
The E26 transformation-specific (ETS) transcription factor family plays multiple roles in cellular processes, such as proliferation, differentiation, migration, and apoptosis, and is implicated in many disorders and cancers [7 –9]. The ETS family members contain a conserved winged helix–turn–helix ETS domain that commonly binds the GGA(A/T) sequence in the ETS-binding site (EBS).
Further, 28 human ETS proteins have been identified and are divided into 12 subfamilies according to their sequence similarity [10]. The ETS variant transcription factor 5 (ETV5; also known as ERM) belongs to the PEA3 subfamily, which includes ETV1, ETV4, and ETV5. ETV5 has attracted attention because of its role in various cancers, including thyroid cancer [11], colorectal cancer [12], synovial sarcoma [13], and neuroblastoma [14,15].
ETV5 regulates the proliferation of mouse embryonic stem cells [16], and it is required for the proliferation of neural stem cells in the ventricular zone of mouse embryos [17]. Further, ETV5 regulates the differentiation of Schwann cells from neural crest progenitor cells [18] and represses NEUROG2 expression to regulate the differentiation of glutamatergic neurons from neural progenitor cells [19].
Zebrafish contain two paralogs of mammalian ETV5, namely Etv5a and Etv5b (also known as Erm in zebrafish) [20]. Previous studies have demonstrated that Etv5a regulates hematopoiesis [20] and kidney development [21], whereas Etv5b mediates the development of hypothalamic serotoninergic neurons [22]. In the present study, we discovered that zebrafish Etv5a regulates the proliferation of neural precursor cells in the neuroectoderm by repressing sox2 and foxm1 expression. This study provides further insights into the role of transcription factors in neural progenitor processes.
Materials and Methods
Ethics statement and zebrafish maintenance
The experiments performed in this study followed standard guidelines for zebrafish research and were approved by the Institutional Animal Care and Use Committee of Chang Gung University (IACUC approval no.: CGU11-118).
Tü (wild-type) zebrafish were purchased from the Zebrafish International Resource Center (Oregon). Adult fish were raised, maintained, and paired, and embryos were collected according to standard guidelines. Embryos were staged according to the hours post-fertilization (hpf) and days post-fertilization [23].
cRNA and morpholino for injection
Zebrafish etv5a, etv5aΔacidic , and etv5b constructs were used to create capped RNA for injection, and they were prepared as previously described [20]. etv5a morpholino (MO) (5′-TCACCTGGGTCTTCAAAGAGGCTCC-3′) that overlaps the ATG start codon (−26 to −2) was purchased from Gene Tools, LLC (Oregon), and the concentration for injection was determined as described by Chen et al. [20].
Zebrafish foxM1 ΔC was amplified with TOOLS Ultra High Fidelity DNA Polymerase (BIOTOOLS, Taiwan). The sox2 promoter region (sox2p , −2.2 to −1.2 kb) and constructs containing the different EBS sequences (EBS-i, EBS-ii, and both EBS-i and EBS-ii deletions) were amplified by polymerase chain reaction (PCR) and cloned into the pEGFP-N3 vector (see Supplementary Table S1 for a full list of primers). We injected 0.15 ng sox2p:eGFP variants and 2.512 ng etv5a cRNA. Embryos were harvested at 8 hpf and subjected to quantitative real-time PCR (qRT-PCR) analysis.
Histological analysis
For in situ hybridization, digoxigenin-uridine triphosphate-labeled riboprobes were synthesized, and in situ hybridization was performed as previously described [24]. mRNA expression was observed using the nitro blue tetrazolium (NBT)/5-Bromo-4-chloro-3-indolyl phosphate (BCIP) substrate (Roche). For double in situ hybridization, digoxigenin- and fluorescein-labeled riboprobes were used and detected using anti-digoxigenin or anti-fluorescein antibodies. These antibodies were conjugated with alkaline phosphatase.
The color reaction protocol involved the following two steps: First, ImmPACT Vector Red (Vector Laboratories, Inc.) was used. Once the initial signals were developed, alkaline phosphatase was inactivated using 0.1 M glycine-HCl at a pH of 2.2. Second, an alkaline phosphatase-conjugated anti-digoxigenin or anti-fluorescein antibody was introduced, followed by the addition of BCIP/NBT for the second color reaction.
For immunohistochemistry, rabbit phospho-histone H3 antibody or rabbit monoclonal anti-active caspase-3 antibody (Abcam) was applied to the samples. The signal was detected using fluorochrome-conjugated Alexa Fluor 488 goat anti-rabbit IgG antibodies (Invitrogen). Goat anti-rabbit IgG horseradish peroxidase (HRP) (Invitrogen) was used to detect the primary antibodies, and 3,3′-diaminobenzidine was used as a substrate for secondary antibody-conjugated HRP (Amresco) (see Supplementary Table S2 for the full list of antibodies).
The embryos were mounted using Vectashield mounting medium and stained with 4′,6-diamidino-2-phenylindole (Vector Laboratories, Inc.) to visualize nuclear DNA and preserve fluorescence. Phospho-histone H3- and caspase-3-positive cells were manually counted.
Quantitative real-time polymerase chain reaction
Total RNA was extracted from the embryos and subjected to cDNA synthesis using the TOOLS Easy Fast RT Kit (BIOTOOLS, Taiwan). RT-qPCR was performed using the TOOLS 2X SYBR qPCR Mix (BIOTOOLS, Taiwan). Gene expression levels were normalized to that of gapdh and measured using the comparative Ct method (40 cycles) according to the manufacturer's instructions (Applied Biosystems).
Cell maintenance and transfection
The 293T (human embryonic kidney) and U87 (human gliobastoma) cell lines were obtained from American Type Culture Collection and cultured in Dulbecco's modified Eagle's medium (Gibco) supplemented with 10% fetal bovine serum (Avantor) and 1% Antibiotics-Antimycotics (GeneTeks). For plasmid transfection, the plasmid DNA was diluted and incubated with DreamFect Gold reagents (OZ biosciences) for 15 min. Subsequently, the mixture was added to the culture medium, which was used to incubate the cells.
Chromatin immunoprecipitation
Chromatin immunoprecipitation (ChIP) was performed as described by Lin et al. [25]. In brief, etv5a was cloned into a pCS2-MT vector containing a Myc tag, transcribed into cRNA in vitro, and injected into zebrafish embryos. The injected embryos were fixed with formaldehyde to crosslink the proteins and DNA. Chromatin was isolated using a Magna ChIP A/G ChIP Kit (EMD Millipore) and fragmented to an average size of ∼300 bp using sonication.
Protein A- and G-conjugated magnetic beads and anti-Myc antibodies were added to the chromatin solution. Protein A- and G-conjugated beads—antibody and chromatin complexes—were collected using a magnetic separator, and the protein/DNA complexes were isolated. The immunoprecipitated DNA was subjected to PCR using primers that frame the EBS-i and EBS-ii regions of the 5′-untranslated region of sox2.
Human ETV5 was cloned into a pCS2-MT vector containing a Myc tag and transfected into U87 cells. The transfected cells were subjected to a ChIP assay using a Magna ChIP A/G ChIP Kit (EMD Millipore). Thereafter, the immunoprecipitated DNA was amplified using primers designed to amplify the EBS sequence in the SOX2 promoter.
Statistical analysis
Quantitative data are presented as mean ± standard deviation. Statistical analyses were performed using Student's t test or one-way analysis of variance (ANOVA). All experiments were performed in triplicate for each sample, and P < 0.05 was considered significant.
Results
etv5a is expressed in the neuroectoderm and developing central nervous system
We found that zebrafish etv5a was expressed in the neuroectoderm, which mainly contains neural progenitors (Fig. 1A). Accordingly, we validated the characteristics of these etv5a-positive cells at 8 h post-fertilization (hpf) by counter-staining for the neural progenitor marker sox2 using double in situ hybridization. sox2 expression was displayed as a rhombus representing the location of neural progenitors (Fig. 1A), and etv5a expression was co-localized with sox2 in the areas shown in Fig. 1B and C.
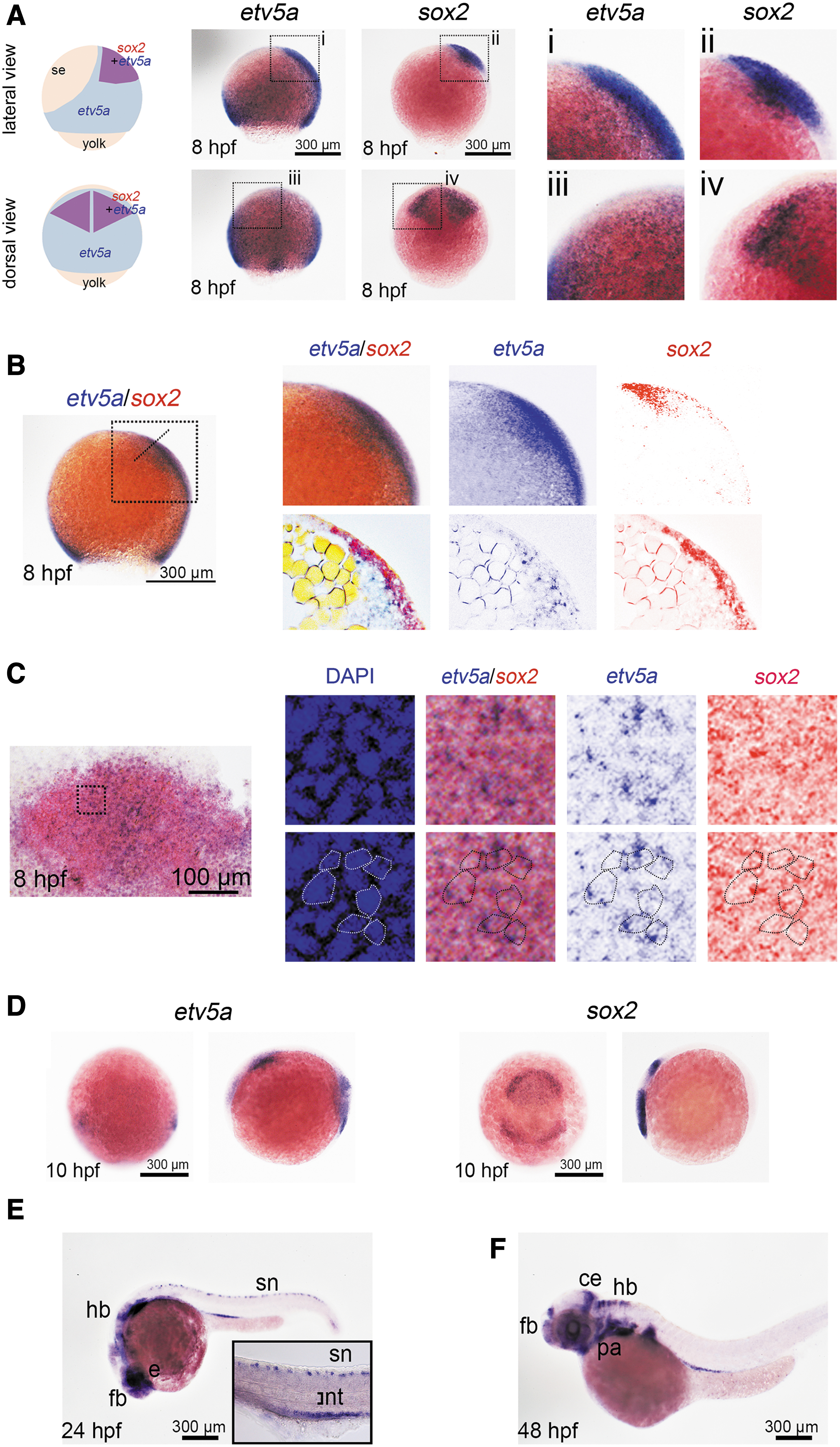
etv5a expression in neural progenitors and neural derivatives. In situ hybridization was performed to determine the type of etv5a-expressing cells in the zebrafish embryos.
Outside of the sox2-positive area, ubiquitous weak expression of etv5a was observed in the entire neural ectoderm and mesoderm, whereas relatively strong expression was observed in the dorsal neuroectoderm and ventral mesoderm, as previously described [20] (Fig. 1A, B). This result confirmed that etv5a is expressed in neural progenitors and may regulate the development of these cells. Further, etv5a was expressed in the brain and spinal cord during the segmentation and organogenesis stages (Fig. 1D–F), indicating that Etv5a may also regulate the formation and differentiation of neural derivatives.
Etv5a deficiency induces the proliferation of neural progenitor cells
To study the role of Etv5a in zebrafish neural development, we used etv5a MO and an etv5a dominant-negative variant to perform loss-of-function analysis. The specificity and effectiveness of this etv5a MO knockdown have been demonstrated in hematopoiesis, and it does not cause off-target p53 activation [20].
Moreover, the effectiveness of etv5 MO was confirmed by a deletion variant that lacks the acidic transcription regulatory domain (etv5aΔacidic ) and causes a dominant-negative effect [20]. Injection of etv5 MO or etv5aΔacidic cRNA induced expression of the neural progenitor marker sox2 (Fig. 2A, C). The phenotypes caused by etv5a MO were rescued by a co-injection with etv5a cRNA (Fig. 2A, C).
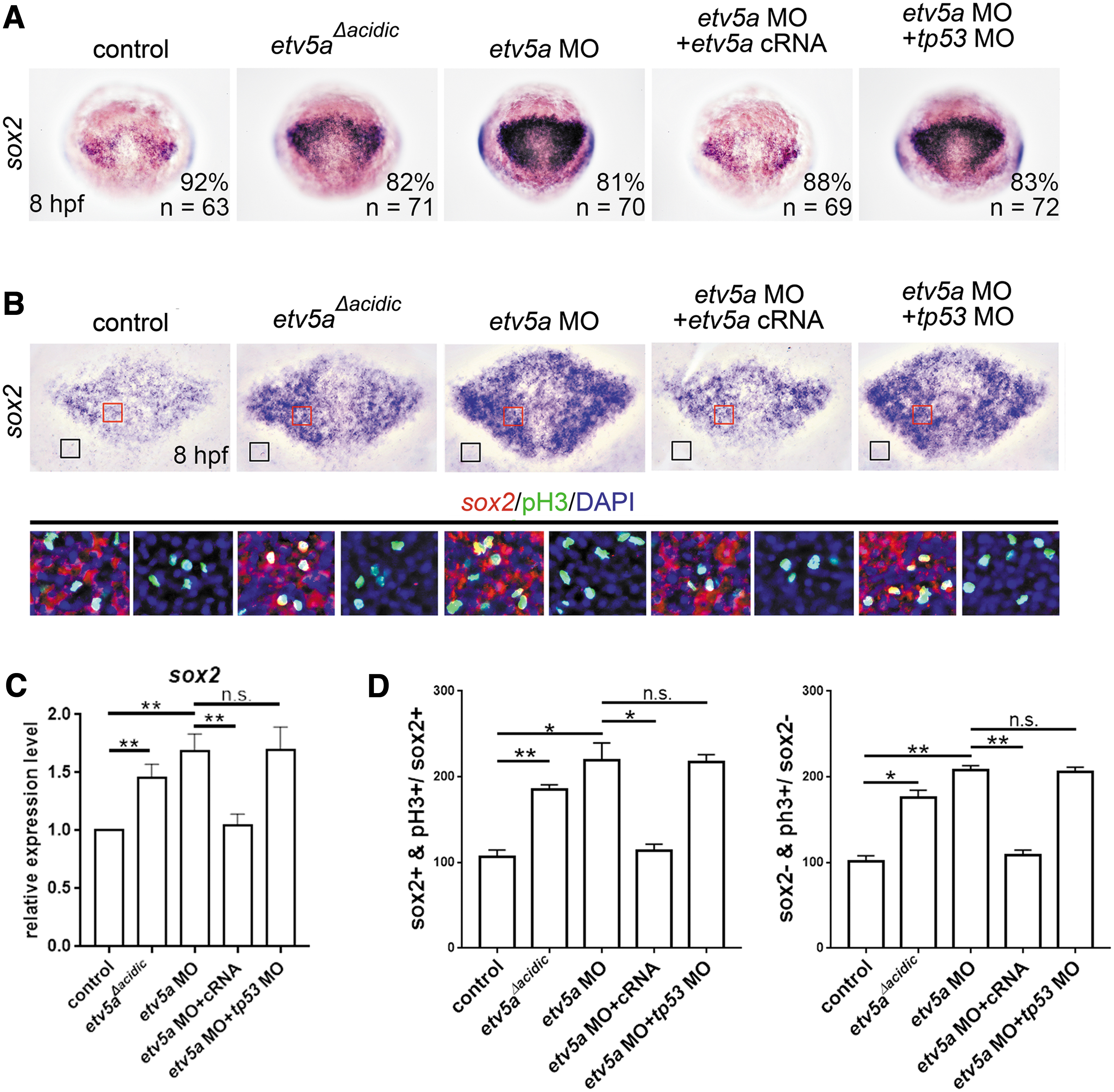
Etv5a deficiency induces the proliferation of neural progenitor cells.
These results demonstrate the specificity of etv5a MO. Next, we analyzed neural progenitor proliferation by counter-staining for sox2 with the cell proliferation marker phosphorylated histone-3 (pH3). Our results revealed that the number of sox2 and pH3 double-positive cells had increased by etv5 MO or etv5aΔacidic injection (Fig. 2B, D), indicating that Etv5a deficiency is sufficient to induce the proliferation of neural progenitors.
Notably, Etv5a deficiency also induced the proliferation of sox2-negative neuroectodermal cells (Fig. 2B, D). This result suggests that Etv5a also regulates the proliferation of neural progenitor cells that give rise to the posterior central nervous system, because sox2 is expressed strongly in the presumptive forebrain and correlates with future subdivisions of the brain [26].
To examine the effect of Etv5a on the apoptosis of neural progenitor cells, we counter-stained for sox2 and apoptotic marker activated caspase-3. Our results revealed that injection of etv5 MO or etv5aΔacidic did not affect the number of sox2 and caspase-3 double-positive cells (Supplementary Fig. S1), indicating that Etv5a deficiency does not regulate the apoptosis of neural progenitors. To further confirm this, we knocked down p53 expression in Etv5a-deficient embryos by co-injecting etv5a MO with tp53 MO and found that tp53 MO could not restore the phenotype of increased sox2 expression caused by etv5a MO (Fig. 2B, D, and Supplementary Fig. S1).
Etv5a deficiency reduces the formation of neural derivatives
Next, we evaluated the effect of Etv5a deficiency on neural derivatives. Injection of etv5a MO or etv5a Δacidic downregulated the expression of the neuronal precursor marker neurog1 at 10 hpf (Fig. 3). In contrast, Etv5a deficiency did not alter the number of caspase-3 positive apoptotic cells in the neurog1-positive areas (Supplementary Fig. S1).
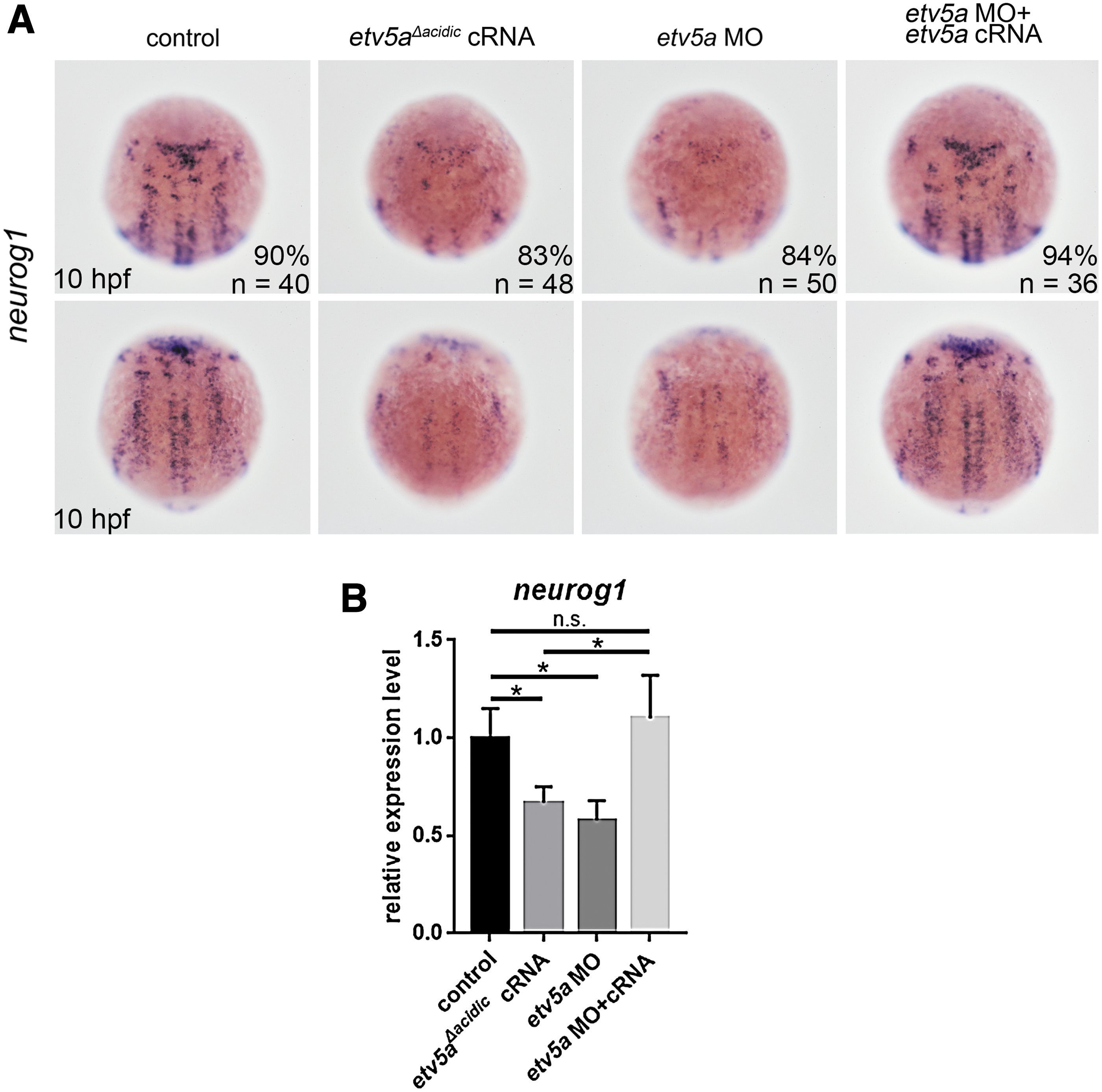
Etv5a deficiency inhibits the development of neuronal derivatives.
This result suggests that Etv5a deficiency causes the proliferation of neural progenitors and maintains progenitor cells, consequently inhibiting neuronal differentiation, as demonstrated by the decreased number of neuronal precursors.
To address the role of Etv5a in gliogenesis, we first analyzed the radial glial marker glial fibrillary acidic protein (Gfap) in Etv5a-depleted embryos at 24 hpf. Injection of etv5a MO or etv5a Δacidic induced gfap expression (Fig. 4). Next, we examined the effect of Etv5a deficiency in glial derivatives using the oligodendrocyte marker olig1 and mag (myelin-associated glycoprotein).
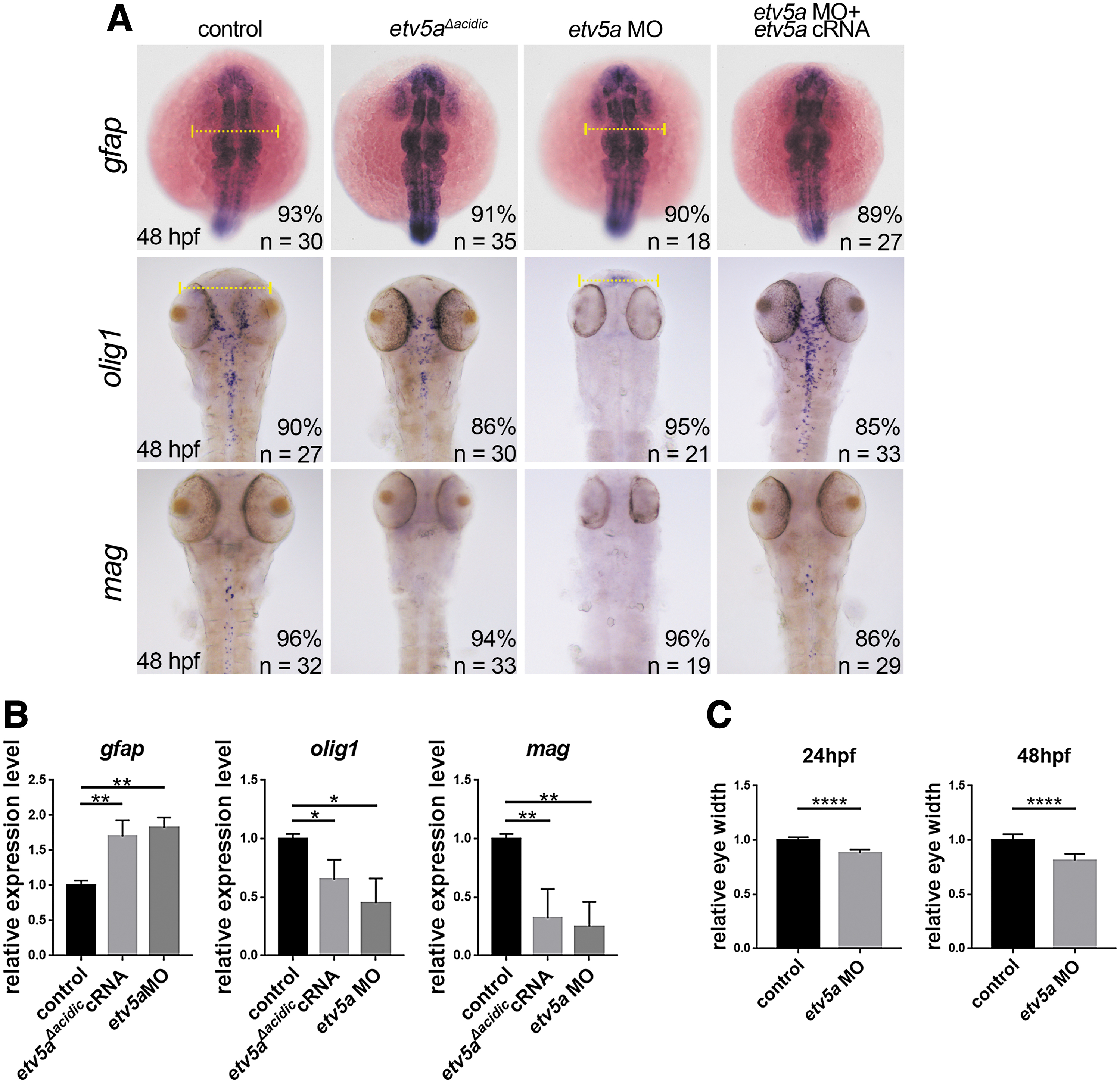
Etv5a deficiency induces radial cell formation and inhibits oligodendrocyte formation.
The injection of etv5a MO or etv5a Δacidic inhibited the expression of olig1 and mag at 48 hpf (Fig. 4), indicating that Etv5a is essential for oligodendrocyte formation. Co-injection with etv5a cRNA rescued these glial phenotypes in Etv5a-depleted embryos.
Etv5a directly binds to the upstream region of sox2 and inhibits its transcription
Etv5a deficiency induced sox2 expression (Fig. 2), whereas overexpression of the full coding sequence of etv5a cRNA reduced sox2 expression, suggesting that Etv5a may bind to sox2 (Fig. 5A, B). Mammalian ETV5 recognizes the consensus sequence ACCGGAAGT(G/A) of the 5′ untranslated region of target genes [27]. We identified two putative Etv5a binding sites (namely EBS-i and EBS-ii) at −2 kb upstream from the start codon of sox2 (Fig. 5C) and discovered the direct binding of Etv5a to the sox2 promoter using ChIP analysis.
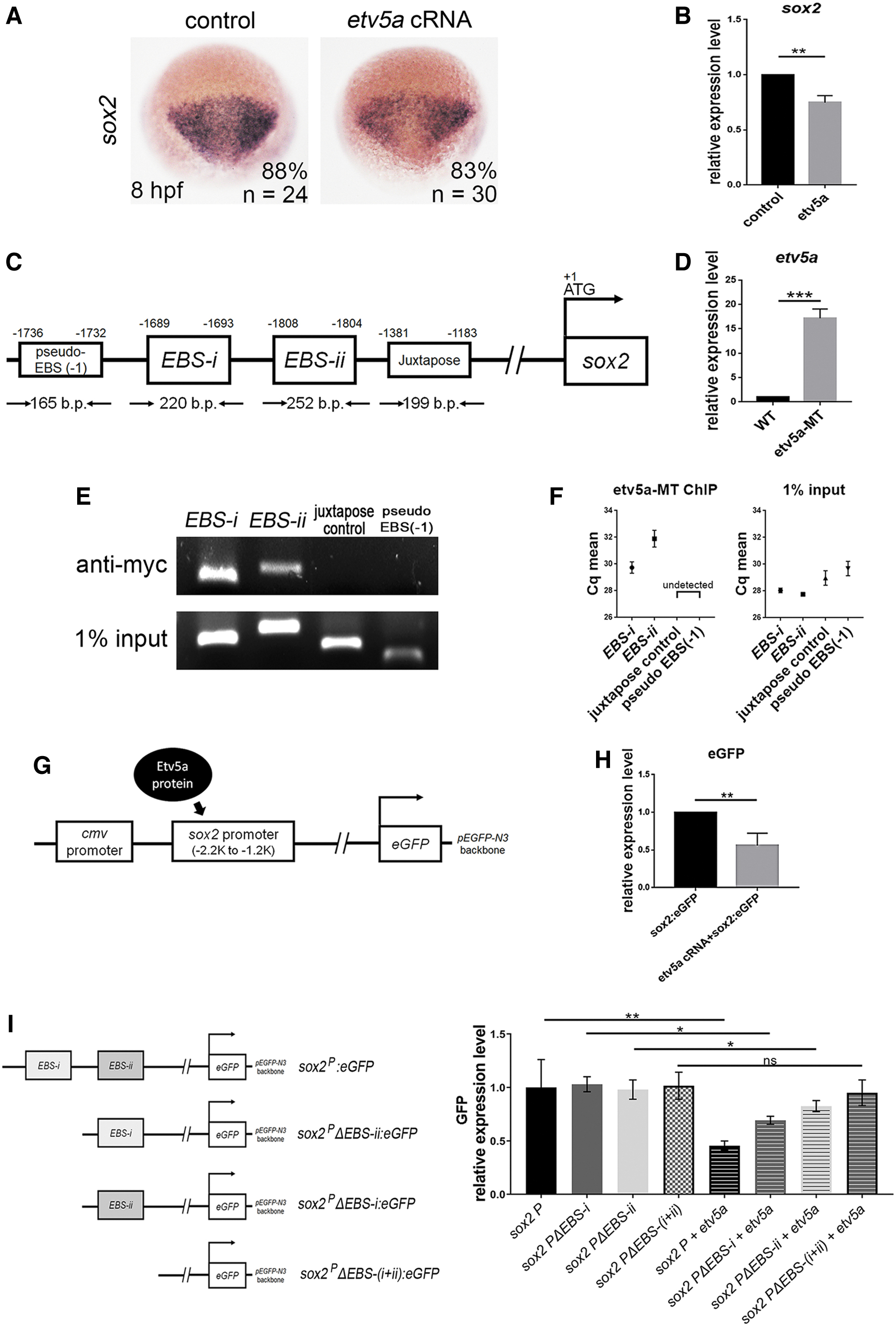
Etv5a represses sox2 expression by directly binding to the promoter region.
Myc-tagged etv5a cRNA was injected into fertilized eggs (Fig. 5D), and chromatin fragments isolated from these embryos were immunoprecipitated with an antibody against the Myc tag. In addition to the two putative binding fragments, we designed two non-binding sequence controls, namely, a fragment adjacent to the putative binding sites and a fragment at the anti-sense strand.
qPCR analysis was performed, using primers designed for the putative binding or non-binding sequences. The putative binding fragments were selectively amplified (Fig. 5E, F). To confirm the results of the ChIP-qPCR assay, we performed an Etv5a-sox2 promoter-reporter assay. The zebrafish sox2 promoter region (−2.2 kb to −1.2 kb), which contains the Etv5a-binding fragments, was cloned into a pEGFP-N3 vector, named sox2p:eGFP.
This sox2p:eGFP construct was injected with or without etv5a cRNA into zebrafish embryos. A concomitant injection of etv5a and sox2p:eGFP inhibited eGFP expression (Fig. 5G, H), demonstrating that Etv5a binds to the sox2 promoter to inhibit eGFP expression. To confirm whether EBS-i and/or EBS-ii are essential for Etv5a to inhibit sox2 expression, we generated GFP reporter constructs driven by the sox2 promoter and containing different EBS-i and EBS-ii sequences.
In these constructs, either one of or both EBS-i and EBS-ii were deleted [sox2pΔEBS-i:eGFP (EBS-i deletion), sox2pΔEBS-ii:eGFP (EBS-ii deletion), and sox2pΔEBS-(i+ii):eGFP (EBS-i and EBS-ii deletion)]. Co-injection of etv5a with sox2p:eGFP inhibited GFP expression, whereas etv5a did not repress GFP expression when co-injected with sox2pΔEBS-(i+ii):eGFP (Fig. 5I).
In contrast, Etv5a inhibited GFP expression when co-injected with sox2pΔEBS-i:eGFP or sox2pΔEBS-ii:eGFP, demonstrating that Etv5a binding to either EBS-i or EBS-ii inhibited GFP expression (Fig. 5I). However, the inhibitory level was less effective than that of Etv5a co-injected with sox2p:eGFP (with intact EBS-i and EBS-ii) (Fig. 5I).
These results affirmed that both EBS-i and EBS-ii are essential for Etv5a binding and that interaction with either inhibits sox2 expression. Taken together, these results indicate that Etv5a inhibits sox2 transcription by directly binding to the EBSs in the promoter region.
Etv5a suppresses neural progenitor proliferation via inhibiting Foxm1 function
In Xenopus embryos, Foxm1 is expressed in the neuroectoderm and is required for the proliferation of neural plate cells [28]. In primary glioblastoma cells, FOXM1 directly binds to the SOX2 promoter, and the overexpression of FOXM1 upregulates SOX2 expression and induces the growth of glioblastoma cells [29]. In zebrafish gastrula, foxm1 was expressed in the neuroectoderm and mesoderm, a pattern very similar to that of etv5a expression (Fig. 6A).
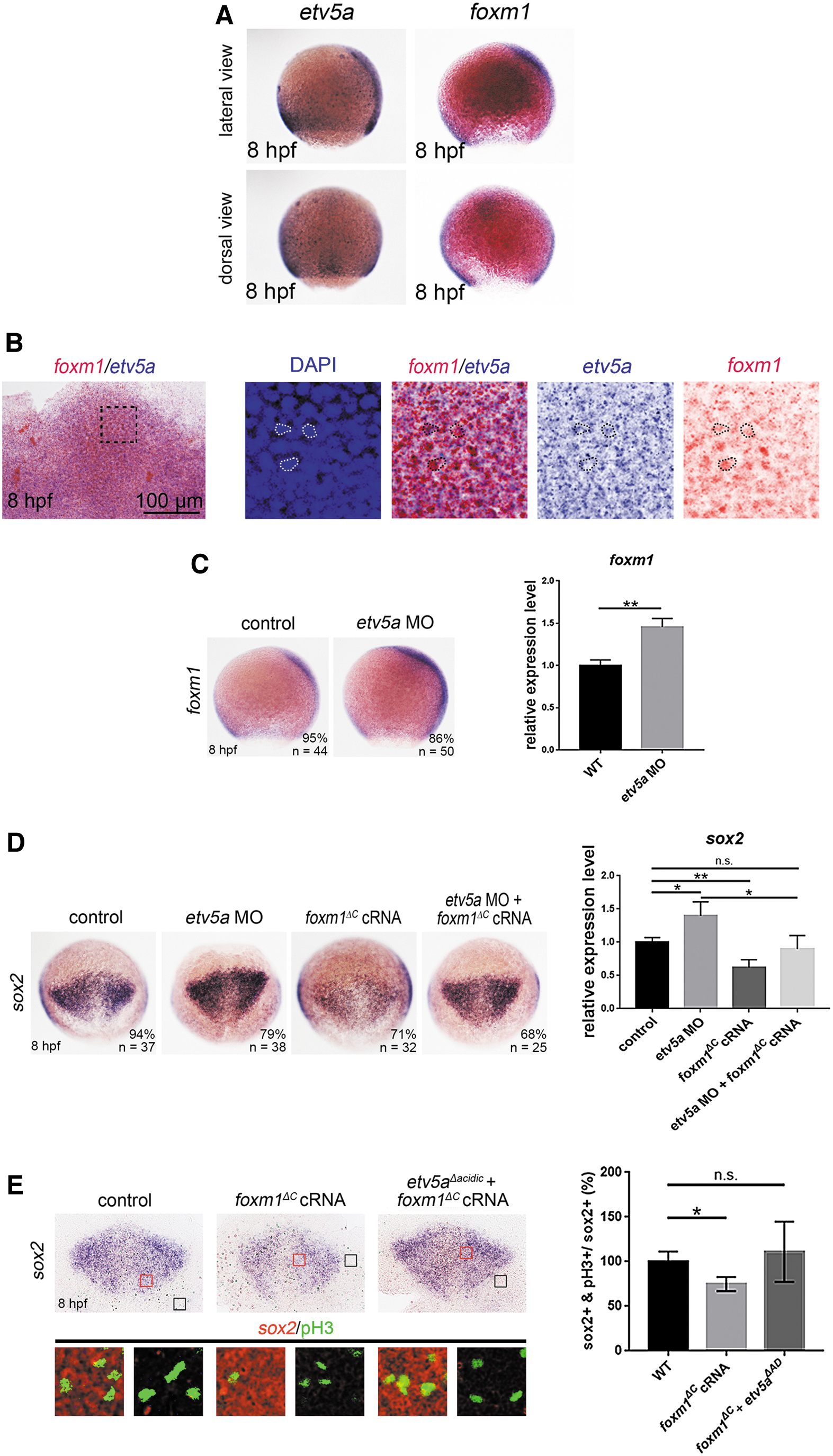
Etv5a deficiency induces sox2 expression via upregulating foxm1 expression.
Double in situ hybridization results demonstrated that the expression of etv5a and foxm1 was co-localized in the neuroectoderm (Fig. 6B), suggesting that Etv5a also regulates Foxm1 in the zebrafish neuroectoderm. To gain insights into the regulatory mechanism of Etv5a in controlling neural progenitor proliferation, we examined the role of Foxm1 in Etv5a-Sox2 mediated neural progenitor proliferation. Injection of etv5a MO significantly upregulated foxm1 expression, as analyzed using in situ hybridization and qRT-PCR (Fig. 6C), suggesting negative regulation of Etv5a on Foxm1.
To confirm that foxm1 upregulation is critical for neural progenitor development in Etv5a-deficient embryos, we used a dominant-negative Foxm1 variant to reduce the effect of foxm1 upregulation and to inhibit Foxm1 function. The design of this variant was based on a previous study demonstrating that deletion of the C-terminal domain of FOXM1 (FOXM1 ΔC ) abolishes the transcriptional activity of endogenous FOXM1 in cancer cell lines [30].
In the present study, the overexpression of zebrafish foxm1 ΔC cRNA reduced sox2 expression and restored the upregulated sox2 expression caused by Etv5a deficiency (Fig. 6D). To verify the role of FoxM1 in neural progenitor proliferation, we overexpressed foxm1 ΔC and examined the proliferation of neural progenitors by counter-staining for sox2 with the proliferation marker, pH3.
The foxm1 ΔC injection reduced the proliferation of neural progenitors, whereas foxm1 ΔC overexpression in Etv5a-deficient embryos restored the number of proliferating neural progenitors, caused by Etv5a deficiency (Fig. 6E). This demonstrated that foxm1 ΔC disrupted the function of Foxm1 and thus nullified the function of the upregulated foxm1 caused by Etv5a deficiency, thereby confirming that Foxm1 is a downstream factor of Etv5a. Thus, Etv5a indirectly regulates sox2 expression and neural progenitor proliferation by suppressing Foxm1 function.
Etv5a, Etv5b, and human ETV5 are functionally diverse in regulating neural progenitors
Zebrafish Etv5a and Etv5b are paralogues to the mammalian ETV5. To examine whether Etv5b is functionally conserved with or diverted from Etv5a, we first examined etv5b expression. In the late gastrula stage, etv5b expression was very similar to that of etv5a, which was expressed in the entire neuroectoderm. At later stages, etv5b expression resembled that of etv5a at 8, 24, and 48 hpf in the developing nervous system (Supplementary Fig. S2).
However, etv5b was not as highly expressed as etv5a in the tail region at 10 hpf. These observations are similar to those previously published [31]. etv5b overexpression did not alter the expression of sox2 (neural progenitor marker) or neurog1 (neural precursor marker) (Fig. 7A), which suggests that etv5b alone did not affect the formation of neural progenitor cells or neural precursors.
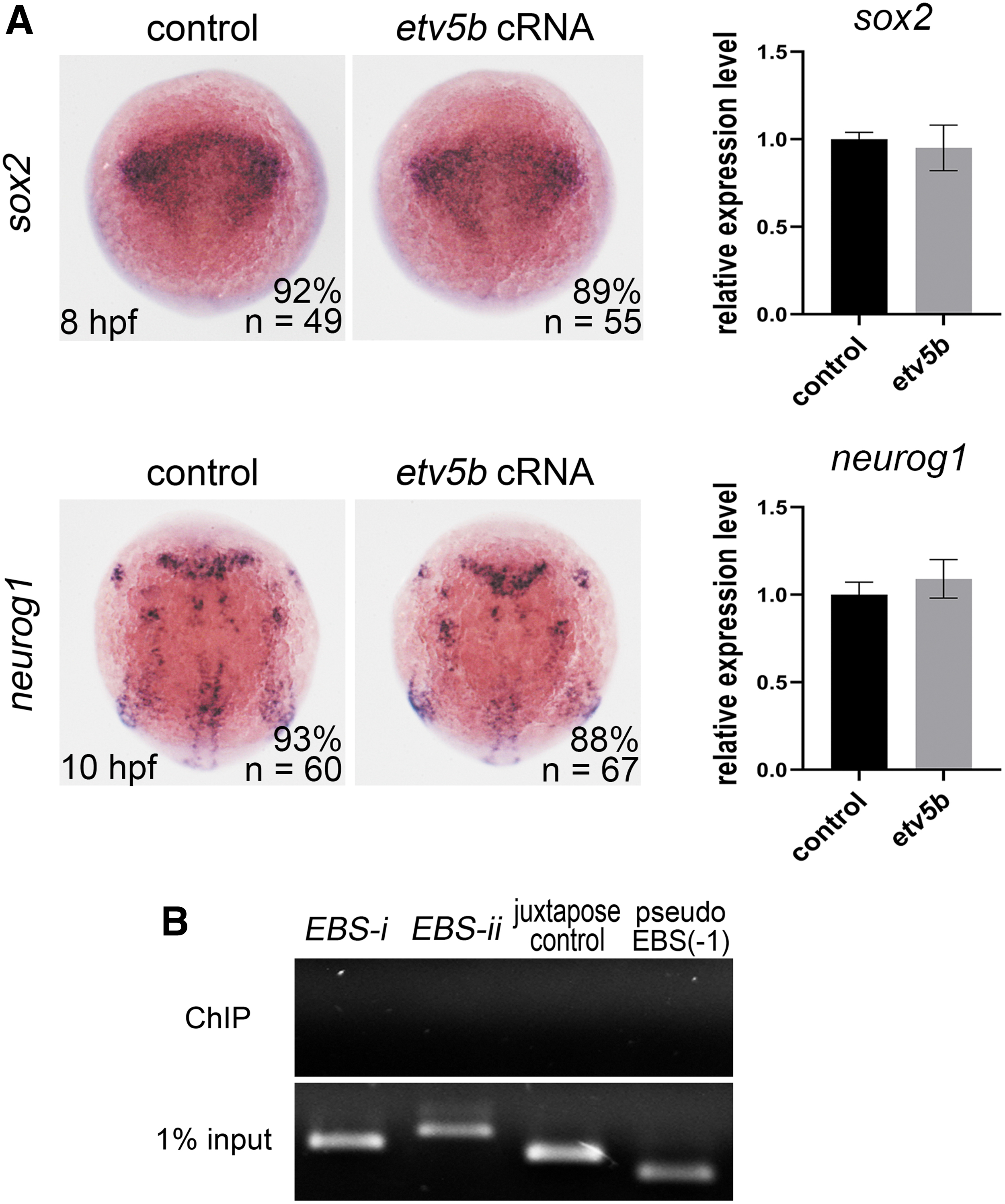
Etv5b has no effect on sox2 or neurog1 expression and does not bind to the sox2 promoter.
In addition, we performed a ChIP-PCR analysis to determine whether Etv5b binds to the sox2 promoter and found that Etv5b did not bind to EBS-i or EBS-ii, the region where Etv5a binds (Fig. 7B), demonstrating that Etv5b could not bind to the sox2 promoter. Overall, these results demonstrated that Etv5a and Etv5b are functionally different in regulating neural progenitors.
Next, we performed a ChIP analysis to investigate the potential interaction between human ETV5 and the SOX2 promoter using the U87 cell line. Myc-tagged ETV5 was transfected into U87 cells, and ETV5-bound chromatin fragments were immunoprecipitated using anti-Myc antibodies. The EBS in the SOX2 promoter was described by Cooper et al. [27], and we designed the primers accordingly to amplify the EBS sequences.
ChIP-PCR and ChIP-qPCR results demonstrated that the EBS was selectively amplified, indicating that ETV5 binds to the EBS in the SOX2 promoter (Fig. 8A–C). In addition, we examined whether zebrafish Etv5a is functionally conserved with human ETV5 by transfecting zebrafish etv5a cRNA into human 293T embryonic kidney cells and U87 glioblastoma cells to examine its effect on human SOX2 expression. qRT-PCR results demonstrated that the transfection of zebrafish etv5a did not alter the expression of human SOX2 (Fig. 8D).
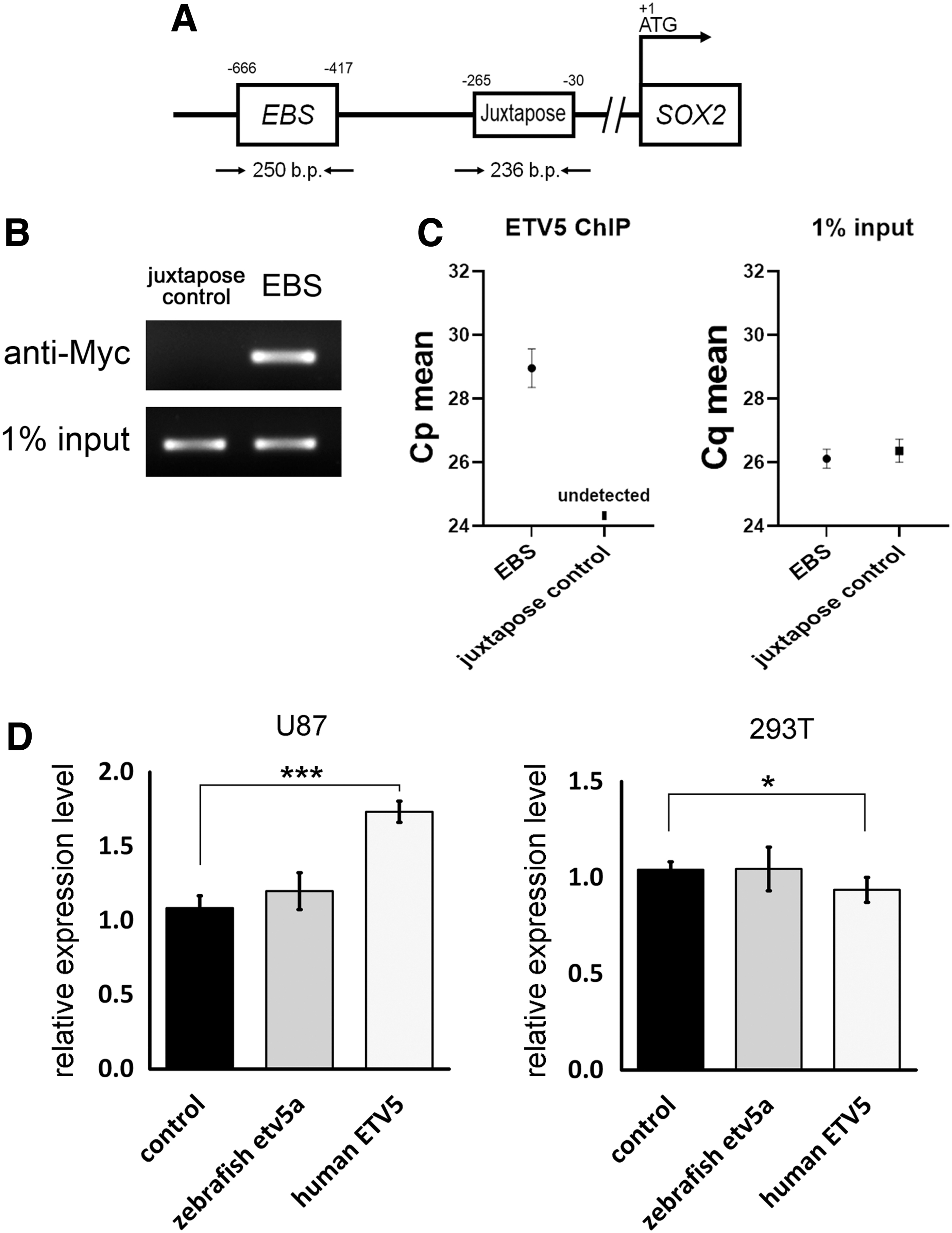
Human ETV5 binds to the EBS in the SOX2 promoter and regulates SOX2 expression.
In contrast, the transfection of human ETV5 induced SOX2 expression in U87 cells but not in 293T cells (Fig. 8D). These findings demonstrated that although both ETV5 and Etv5a are capable of binding to EBS in the SOX2 and sox2 promoters, respectively, they have distinct functions within their respective species. Further, human ETV5 induced SOX2 expression in U87 cells but not in 293T cells, which also indicated that the function of ETV5 varies depending on the cell type.
Discussion
The self-renewal and multipotency characteristics of neural progenitor cells make them attractive for studies on developmental biology, tissue regeneration, and stem cell therapy. However, the molecular regulation of the development of neural progenitor cells is unclear, and data obtained from in vivo and in vitro analyses are inconsistent.
In this study, we discovered that zebrafish Etv5a inhibited the proliferation of neural progenitor cells in the embryonic neuroectoderm by directly binding to sox2 regulatory elements and inhibiting sox2 transcription. Further, Etv5a indirectly inhibits sox2 expression by inhibiting the expression of foxm1, a sox2 upstream gene. Our results reveal a hierarchy of transcriptional regulation in the proliferation of neural progenitor cells.
Etv5a and Etv5b exhibit high similarity in terms of their amino acid sequences; however, their redundant roles have not been fully determined. Simultaneous knock-down of Etv5a, Etv5b, and Etv4 (previously named Pea3) causes defects in cardiac progenitors and isthmic organizers, demonstrating the redundant role in regulating cell development [32].
However, they also perform non-redundant roles in several tissues. Etv5a regulates hemato-vascular derivatives [20] and nephron epithelial cells [21], whereas Etv5b governs hypothalamic-serotoninergic neurons [22], retina [33], and rhombomere segmentation [34,35]. Our findings demonstrated that only Etv5a (and not Etv5b) regulates sox2 expression and the formation of neural progenitors.
In our previous study, we found that Etv5a downregulation increases the proliferation of ventral mesoderm cells and causes defects in hemato-vascular derivatives [20]. Therefore, Etv5a may regulate the proliferation of many cell types with stemness or progenitor characteristics. Cancer stem cells share many characteristics with normal ones, such as high proliferation, self-renewal, and multipotency.
Therefore, Etv5a deficiency induced the proliferation of neural progenitor cells, suggesting that ETV5 may act in the same manner in cancer cells as in the nervous system. However, studies have demonstrated an oncogenic role of ETV5 in tumors of the nervous system. In neuroblastoma cell lines, anaplastic lymphoma kinase activation induces ETV5 expression, and ETV5 binds to the RET promoter to drive neuroblastoma oncogenesis [14].
Hence, ETV5 is required for proliferation, migration, and colony formation in neuroblastoma cells [15]. Moreover, ETV5 is essential for glioma growth in a mouse model with a constitutively active Ras pathway [36]. FOXM1 is also oncogenic in nervous system tumors. FOXM1 binds to SOX2 to promote stemness and radiation resistance in glioblastoma cells [29].
Further, FOXM1 induces SOX2 expression and inhibits neuroblastoma cell differentiation [37]. SOX2 is considered to be an oncogene in many tumors. In tumors of the nervous system, SOX2 is highly expressed in glioblastomas [38] and maintains the stemness of glioma stem cells [39] and medulloblastoma cells [40]. According to these studies, ETV5, SOX2, and FOXM1 are oncogenic in nervous system tumors. This conclusion is inconsistent with our observation that Etv5a inhibits the proliferation of neural progenitors by inhibiting sox2 expression and foxm1-sox2 signaling.
Most studies involving mammalian ETV5 have been performed in immortalized tumor cell cultures whose tissue complexity, heterogeneity, and molecular regulation could be very different from those of animal models, namely the zebrafish model used in the present study. Overall, these results indicate that the role of Etv5a in embryonic neural progenitors is different from that of ETV5 in tumor cells of the nervous system.
Although we discovered that Etv5a is upstream of the transcriptional hierarchy that regulates the proliferation of neural progenitor cells, the upstream regulator of Etv5a is unknown in this cell population. The major signaling pathways that regulate neural progenitor development are BMP, Wnt, FGF, SHH, and Notch. By searching the published literature listed in PubMed, we did not find evidence describing the regulation of BMP, Wnt, SHH, and Notch on ETV5/Etv5a.
In contrast, FGF signaling induces the expression of ETV5/etv5. In zebrafish embryos, Fgf3 and Fgf8 induce etv5b expression in various regions of the developing nervous system [41,42]. During neural induction, FGF inhibits BMP signaling in neuroectoderm specification. As Etv5a is expressed in the neuroectoderm and inhibits sox2 expression, it is required for inhibiting neural progenitor cells.
One possible explanation is that FGF-Etv5a signaling induces neural induction, temporarily inhibits sox2 expression, and downregulation of Etv5a expression relieves this inhibition and favors Sox2 function in the formation of neural progenitor cells. However, this hypothesis and the upstream signaling that regulates Etv5a in the neuroectoderm remain to be confirmed.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from Chang Gung Medical Foundation and Chang Gung Memorial Hospital (CMRPD1I0093, CMRPD1M0091, and BMRP857 for Y.-C.C.; CORPG3J0261, CORPG 3J0262, and CMRPG3H0322 for Y.-C.H.), the Ministry of Science and Technology, Taiwan (MOST 106-2311-B-182-001-MY3 and 110-2320-B-182-008-MY3 for Y.-C.C.; NMRPG3J6141 and NMRPG3J6142 for Y.-C.H.), and Taipei Medical University (DP2-107-21121-01-N-10, DP2-108-21121-01-N-10-01, TMU106-AE1-B20 for T.-H.Y.).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.