
Abstract
Periodontal tissue regeneration is important for preserving teeth. Periodontal ligament stem cells (PDLSCs) are useful in periodontal tissue regeneration; however, tooth extraction is required to obtain these cells. Therefore, we focused on induced pluripotent stem (iPS) cells and established a method to obtain PDLSC-like cells from iPS cells. Specifically, we first differentiated iPS cells into neural crest–like cells (iNCs). Next, we obtained PDLSC-like cells (iPDLSCs) by culturing iNCs on extracellular matrix (ECM) derived from human primary periodontal ligament cells (HPDLCs). This differentiation method suggested that ECM derived from HPDLCs is important for iPDLSC differentiation. Thus, we aimed to identify the PDLSC-inducing factor present in HPDLC-derived ECM in this study. We first performed comprehensive analyses of HPDLC genes and identified fibrillin-2 (FBN2), an ECM-related factor. Furthermore, to clarify the effect of FBN2 on iPDLSC differentiation, we cultured iNCs using ECM derived from HPDLCs with FBN2 knocked down. As a result, expression of PDL-related markers was reduced in iNCs cultured on ECM derived from HPDLCs transfected with FBN2 siRNA (iNC-siFBN2) compared with iPDLSCs. Furthermore, the expression of CD105 (a mesenchymal stem cell marker), proliferation ability, and multipotency of iNC-siFBN2 were lower compared with iPDLSCs. Next, we cultured iNCs on FBN2 recombinant protein; however, expression of PDL-related markers did not increase compared with iPDLSC. The present results suggest the critical involvement of FBN2 in inducing iPDLSCs from iNCs when in fact it does not promote iPDLSC differantiation. Therefore, we need to elucidate the entire HPDLC-ECMs, responsible for iPDLSCs induction.
Introduction
Severe periodontal disease and caries often cause severe damage to periodontal (PDL) tissues, leading to tooth extraction. 1 –3 Tooth loss reportedly causes oral frailty and ultimately leads to a decline in quality of life. 4 –6 Therefore, various methods to regenerate periodontal tissue have been developed to preserve teeth, such as stem cell transplantation, use of recombinant growth factors, and improvement of scaffolds to replace bone. 7 –10 However, regenerative therapy that can be applied to severely damaged periodontal tissue has not yet been established.
Periodontal ligament stem cells (PDLSCs) are derived from cranial neural crest cells (NCs) 11 and comprise ∼0.07% of cells in PDL tissues. 12 PDLSCs have some unique characteristics including reported expression of mesenchymal stem cell (MSC) markers (eg, CD90, CD105, and CD146) and multipotency to differentiate into osteoblasts, adipocytes, and chondrocytes in vitro. 13 –15 Several reports have revealed that PDLSC clones express PDL-related markers such as α-smooth muscle actin (α-SMA), collagen I (COL1), osteoprotegerin (OPG), periodontal ligament-associated protein 1 (PLAP1), and periostin (POSTN). 16,17 Furthermore, Feng et al. showed that PDLSCs could be autologously transplanted into patients with periodontal disease and confirmed periodontal tissue regeneration. 18 As described above, although PDLSCs appear to be useful for periodontal tissue regeneration, tooth extraction is required to obtain these cells. In other words, it is difficult to apply periodontal tissue regeneration therapy using PDLSCs to patients who do not have extra teeth, such as wisdom teeth.
Induced pluripotent stem (iPS) cells can be established from somatic cells and have high multipotency. 19,20 In a previous report, we focused on iPS cells and succeeded in differentiating them into PDLSC-like cells (iPDLSCs) using a unique method of culturing iPS cell-derived neural crest cells (iNCs) on extracellular matrix (ECM) derived from human PDL cells (HPDLCs). 21 However, our results revealed that iNCs cultured on ECM derived from skin fibroblasts (SFs) did not have the characteristics of PDLSCs. These results suggest that there is a factor in ECM derived from HPDLCs that induces iNCs to differentiate into iPDLSCs.
ECM is reportedly important for the differentiation and maturation of stem cells. Wijelath et al. revealed that culturing endothelial progenitor cells on fibronectin, a well-known ECM, promoted their differentiation into endothelial cells in vitro. 22 Laminin activated integrin/phosphoinositide-3K/focal adhesion kinase signaling to promote cardiomyocyte differentiation in rat embryonic stem cells. 23 However, the ECM involved in PDLSC differentiation has not yet been clarified.
In this study, we performed a comprehensive analysis of ECM derived from HPDLCs and aimed to identify the factors involved in PDLSC induction.
Materials and Methods
Cell culture
Human iPS cells derived from SFs and SFs were purchased from RIKEN (Saitama, Japan, HPS No.0063). Human iPS cells were cultured on mouse embryonic fibroblasts (ReproCELL, Kanagawa, Japan) in Primate ES Cell Medium (ReproCELL) containing 5 ng/mL human recombinant basic fibroblast growth factor (ReproCELL). HPDLCs were isolated from the teeth of two healthy patients who visited Kyushu University: a third molar from a 23-year-old male patient (3S) and a third molar from a 23-year-old female patient (3U). HPDLCs and SFs were maintained in α-Minimum Essential Medium (α-MEM; Thermo Fisher Scientific, Waltham, MA, USA) containing 10% fetal bovine serum (FBS; Sigma-Aldrich, St. Louis, MO, USA), 50 μg/mL streptomycin, and 50 U/mL penicillin (10% FBS/α-MEM). All procedures were performed in compliance with the Research Ethics Committee, Faculty of Dentistry, Kyushu University.
Microarray analysis
Total RNA was isolated from HPDLC-3U and SFs using TRI Reagent (Molecular Research Center, Cincinnati, OH, USA) and subsequently purified. Microarray analysis using an Agilent Technologies System (Agilent Technologies, Santa Clara, CA, USA) was performed by Cell Innovator, Inc. of the Venture Business Laboratory of Kyushu University (Fukuoka, Japan).
Cap analysis gene expression and gene ontology enrichment analysis
Total RNA was isolated from HPDLC-3U, HPDLC-3S, and SFs using TRI REAGENT and subsequently purified. Cap analysis of gene expression (CAGE) allows genome-wide analysis of gene transcription start sites and quantitative study of the RNA transcribed from them by DNAFORM (Yokohama, Kanagawa, Japan). 24
Preparation of coating plates
HPDLCs were seeded at a density of 1 × 104 cells/well into a 24-well plate. After 4 days, HPDLCs were detached using 2% EDTA (Nacalai Tesque, Kyoto, Japan) treatment to obtain plates coated with ECM derived from HPDLCs.
In addition, human FBN2-recombinant protein-coated plates were also prepared. FBN2 protein solutions with different concentrations were put into a 24-well plate and incubated for 2 h at 37°C.
Knockdown of FBN2 in HPDLCs
Small interfering RNA (siRNA) for human FBN2 (MISSION siRNA, Sigma-Aldrich) or human control siRNA (MISSION siRNA Universal Negative Control, Sigma-Aldrich) were transfected with HPDLC-3U using Lipofectamine RNAiMAX (Thermo Fisher Scientific) according to the manufacturer’s protocol. Briefly, HPDLC-3U cells were seeded in 24-well plates at a density of 1 × 104 cells per well. After 2 days, siRNAs were transduced. The siRNA-lipid complex was prepared by mixing 5 pmol of siRNA and 0.75 µL of Lipofectamine RNAiMAX in 50 µL of α-MEM. The complex was incubated for 5 min at room temperature, then added to cells, and the mixture was incubated for 48 h.
Development of iPDLSCs, iNC-cont, and iNC-siFBN2 cells
Human iPS cells were first differentiated into iNCs, according to a previous report. 25 Briefly, colonies of iPS cells were separated into single cells and cultured on Matrigel-coated dishes at a density of 1 × 105 cells/dish. When the cells reached 50% to 70% confluence, they were cultured in Knockout Serum Replacement Medium and Neurobasal Medium for 12 or 13 days. After culturing, iNCs were isolated by magnetic-activated cell sorting (Miltenyi Biotec, Bergisch Gladbach, Germany) using a bead-conjugated antibody for p75NTR, a neural crest marker. The resulting iNCs were cultured further for 2 weeks on ECM produced by cultured HPDLC-3U (designated as iPDLSCs), HPDLC-3U + control siRNA (designated as iNC-cont cells), or HPDLC-3U + FBN2 siRNA (designated as iNC-FBN2 cells).
Proliferation assay
iPDLSC, iNC-siCont, and iNC-siFBN2 cells were seeded at a density of 5 × 103 cells/well into the wells of 48-well plates for up to 3 days. On days 0, 1, 2, and 3, proliferation was examined using a WST-1 Cell Proliferation Assay Kit (Takara Bio, Shiga, Japan) according to the manufacturer’s protocol. The supernatant density was measured by an iMark microplate reader (Bio-RAD Laboratories, CA, USA) at an absorbance of 450 nm.
Flow cytometric analysis
iPDLSC, iNC-siCont, and iNC-siFBN2 cells were detached with Accutase (ReproCELL) and the number of cells was adjusted to 5 × 105 cells/tube. Cells were then incubated with PE-conjugated mouse anti-human CD34, CD45, CD90, CD105, or CD146 antibodies (Sony Biotechnology, San Jose, CA, USA) for 1 h at 4°C. Cells were washed with flow cytometry buffer (R&D Systems) and the percentage of positive cells was measured by flow cytometry (EC800 Cell Analyzer; Sony, Tokyo, Japan). The data were analyzed using Eclipse software (Sony).
Quantitative Reverse Transcription Polymerase Chain Reaction
Total cellular RNA was isolated by TRI Reagent and reverse‐transcribed using a PrimeScript RT Reagent Kit (Takara Bio). Quantitative polymerase chain reaction (PCR) assays were carried out with a KAPA SYBR Fast qPCR kit (Nippon Genetics, Tokyo, Japan) using a Thermal Cycler Dice Real‐Time System (Takara Bio) according to the manufacturer's protocol. Primer sequences, annealing temperatures, and product sizes for human α-SMA, bone morphogenetic protein-2 (BMP2), BMP4, COL1, FBN2, lipoprotein lipase (LPL), osteocalcin (OCN), OPG, PLAP1, POSTN, Peroxisome Proliferator-Activated Receptor γ (PPARγ), and β-actin are listed in Table 1. Primer sequences were designed using the GenBank database (National Center for Biotechnology Information), and a BLAST search of GenBank was performed on the primer sequences to ensure specificity.
Primer Sequence, Product Size, Annealing Temperature, for Quantitative Reverse Transcription Polymerase Chain Reaction
Western blotting
Cell lysate was isolated by using RIPA buffer (Thermo Fisher Scientific) including a protease inhibitor cocktail (Sigma‐Aldrich). An antihuman β‐actin mouse polyclonal antibody (1:1000; Santa Cruz Biotechnology) or an antihuman FBN2 mouse polyclonal antibody (1:100; Santa Cruz Biotechnology) were used as primary antibodies. Biotinylated anti‐mouse IgG (Nichirei Bioscience Inc) was used as secondary antibody. Membranes were then incubated with streptavidin‐HRP (Sigma‐Aldrich). Positive reactions were visualized by using ECL Select (Thermo Fisher Scientific).
Osteoblastic differentiation
iPDLSC, iNC-siCont, and iNC-siFBN2 cells were seeded at a density of 2 × 104 cells/well into the wells of a 24-well plate and cultured in 10% FBS/α-MEM. After reaching confluence, the culture medium was replaced with osteoblastic differentiation medium (OM) composed of 10% FBS/α-MEM supplemented with 2 µM β-glycerophosphate (Sigma Aldrich) and 50 μg/mL ascorbic acid (Nacalai Tesque). After 3 weeks, cells were fixed with 10% formalin (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) and stained with 0.3% Alizarin Red S (Thermo Fisher Scientific) for calcium detection.
Adipocytic differentiation
iPDLSC, iNC-siCont, and iNC-siFBN2 cells were seeded at a density of 2 × 104 cells/well into the wells of a 24-well plate and cultured in 10% FBS/α-MEM. After reaching confluence, the culture medium was replaced with adipocytic differentiation medium (AM) composed of 10% FBS/α-MEM supplemented with 1%
Statistical analysis
All values are expressed as the mean ± standard deviation. Statistical analyses were performed using appropriate tests (unpaired doubled-tailed t-test or one-way ANOVA). A P-value of <0.05 was considered statistically significant.
Results
Comprehensive analysis of extracellular matrix secreted by HPDLCs
First, we performed two comprehensive analyses to select the factors involved in iPDLSC induction from ECM-derived HPDLCs. In our previous report, we showed that SF-derived ECM did not have the ability to induce iPDLSCs. We also revealed that the efficiency of iPDLSC induction differed depending on the ECM of HPDLCs. In brief, HPDLC-3U-derived ECM had a higher ability to differentiate iNCs into iPDLSCs than HPDLC-3S-derived ECM, which had a higher ability than SF-derived ECM. Based on these results, we performed microarray analysis of SFs and HPDLC-3U and CAGE analysis of SFs, HPDLC-3S, and HPDLC-3U.
CAGE analysis detected a total of 13,718 genes (Fig. 1A), 94 of which were expressed at a level five times higher in HPDLC-3U than in SFs and twice the level in HPDLC-3U than in HPDLC-3S. Microarray analysis detected a total of 1,255 genes (Fig. 1B), 413 of which were expressed at a level five times higher in HPDLC-3U than SFs. Among the 413 genes selected by microarray analysis and the 94 genes selected by CAGE analysis, 28 genes matched. Among these 28 genes, two genes matched the Gene Set Enrichment Analysis ECM gene set: BMP4 and FBN2 (Fig. 1C).
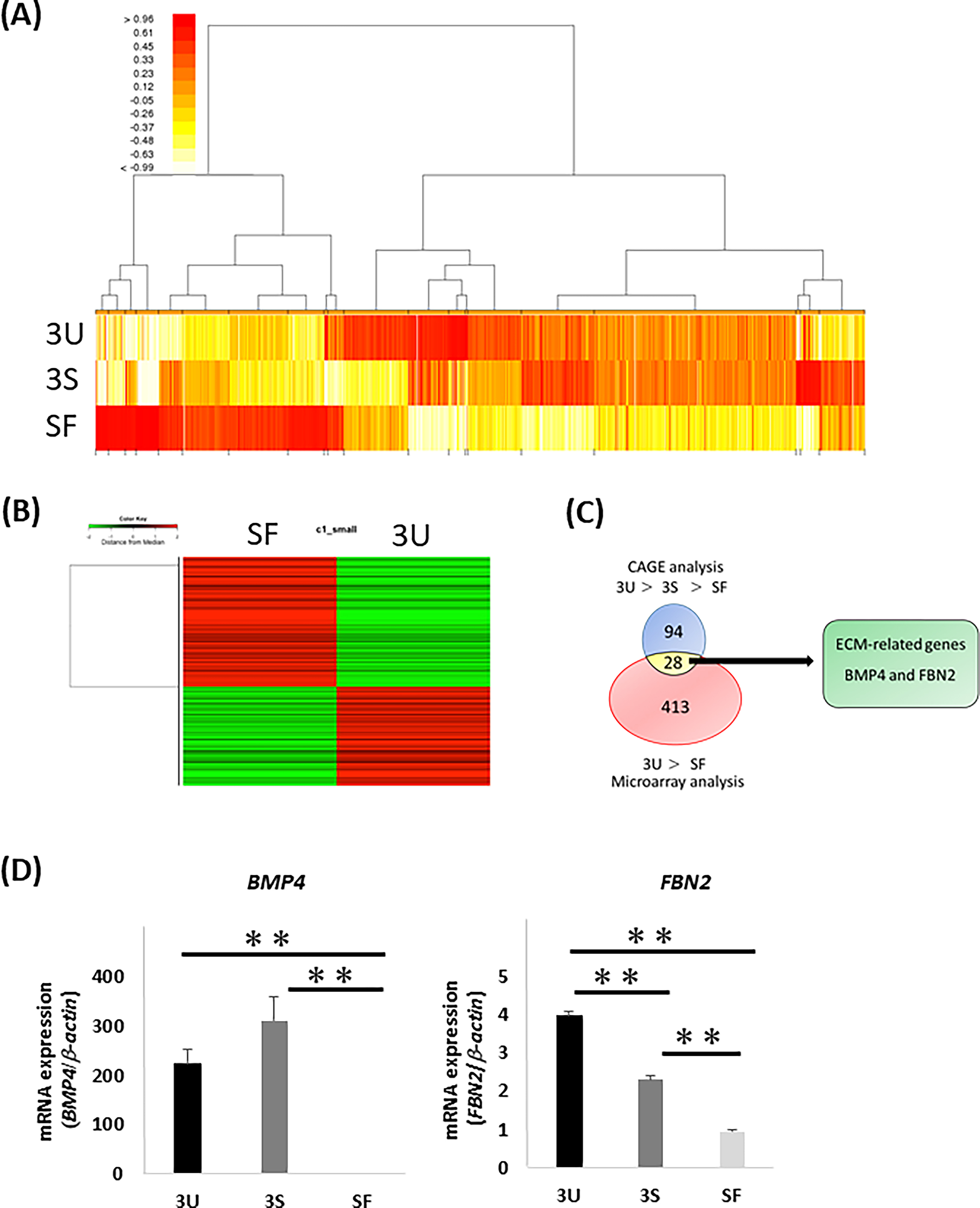
Identification of ECM with ability to induce iPDLSCs.
Gene expression of BMP4 in HPDLCs was significantly higher than in SFs, whereas there was no difference between HPDLC-3U and HPDLC-3S. Gene expression of FBN2 in HPDLCs was significantly higher than in SFs, and FBN2 was more highly expressed in HPDLC-3U than in HPDLC-3S. These results suggest that FBN2 is involved in the induction of iPDLSCs.
Expression of PDL-related markers in iPDLSCs, iNC-Cont, and iNC-siFBN2 cells
To evaluate the effect of FBN2 in HPDLCs on differentiation of iNCs into iPDLSCs, iNCs were cultured in wells coated with ECM derived from HPDLCs transfected with FBN2 siRNA (Fig. 2A). Gene and protein expression of FBN2 were suppressed in HPDLCs transfected with FBN2 siRNA (Supplementary Fig. S1A and B). Quantitative RT-PCR was used to analyze the expression of PDL-related genes (i.e., α-SMA, COL1, OPG, PLAP1, and POSTN) in iPDLSCs, iNC-Cont, and iNC-siFBN2 cells. We found that the expression of each of these PDL-related genes in iNC-siFBN2 cells was decreased compared with iPDLSCs and iNC-Cont cells (Fig. 2B).
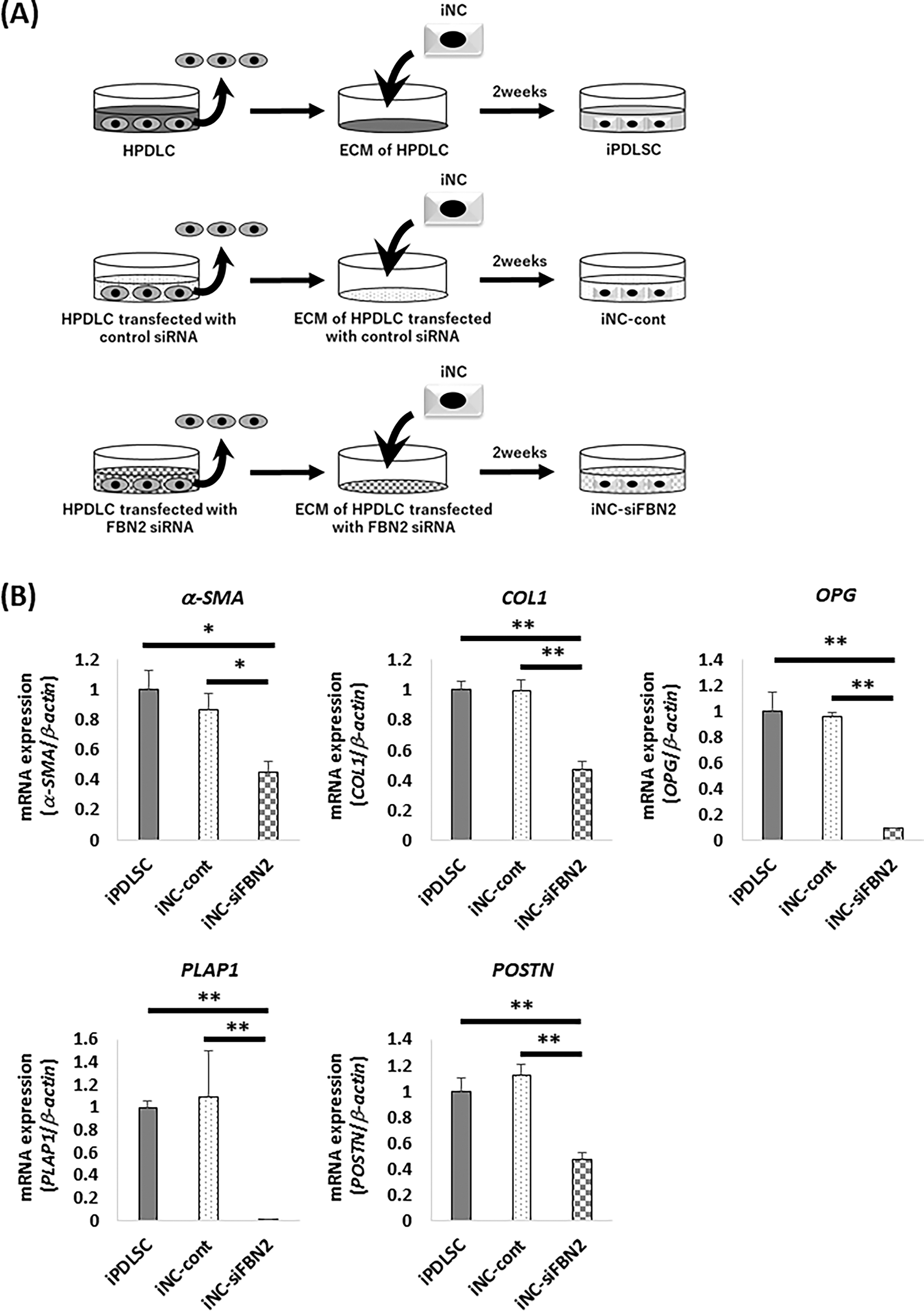
Expression of PDL-related markers in iPDLSCs, iNC-cont, and iNC-siFBN2 cells.
Overall, we observed that FBN2 plays an important role in the acquisition of periodontal ligament properties of iNCs.
Expression of MSC-related markers by iPDLSCs, iNC-cont, and iNC-siFBN2 cells
Next, we assessed the expression of MSC-related markers CD90, CD105, and CD146 in iPDLSC, iNC-cont, and iNC-siFBN2 cells using flow cytometric analysis (Fig. 3). There was no change in the expression of CD90 or CD146 in any cells. However, we found that iNC-siFBN2 cells displayed decreased expression of CD105 compared with iPDLSCs and iNC-cont cells (Fig. 3A). Expression of hematopoietic stem cell markers CD34 and CD45 was negative in all cells (Fig. 3B).
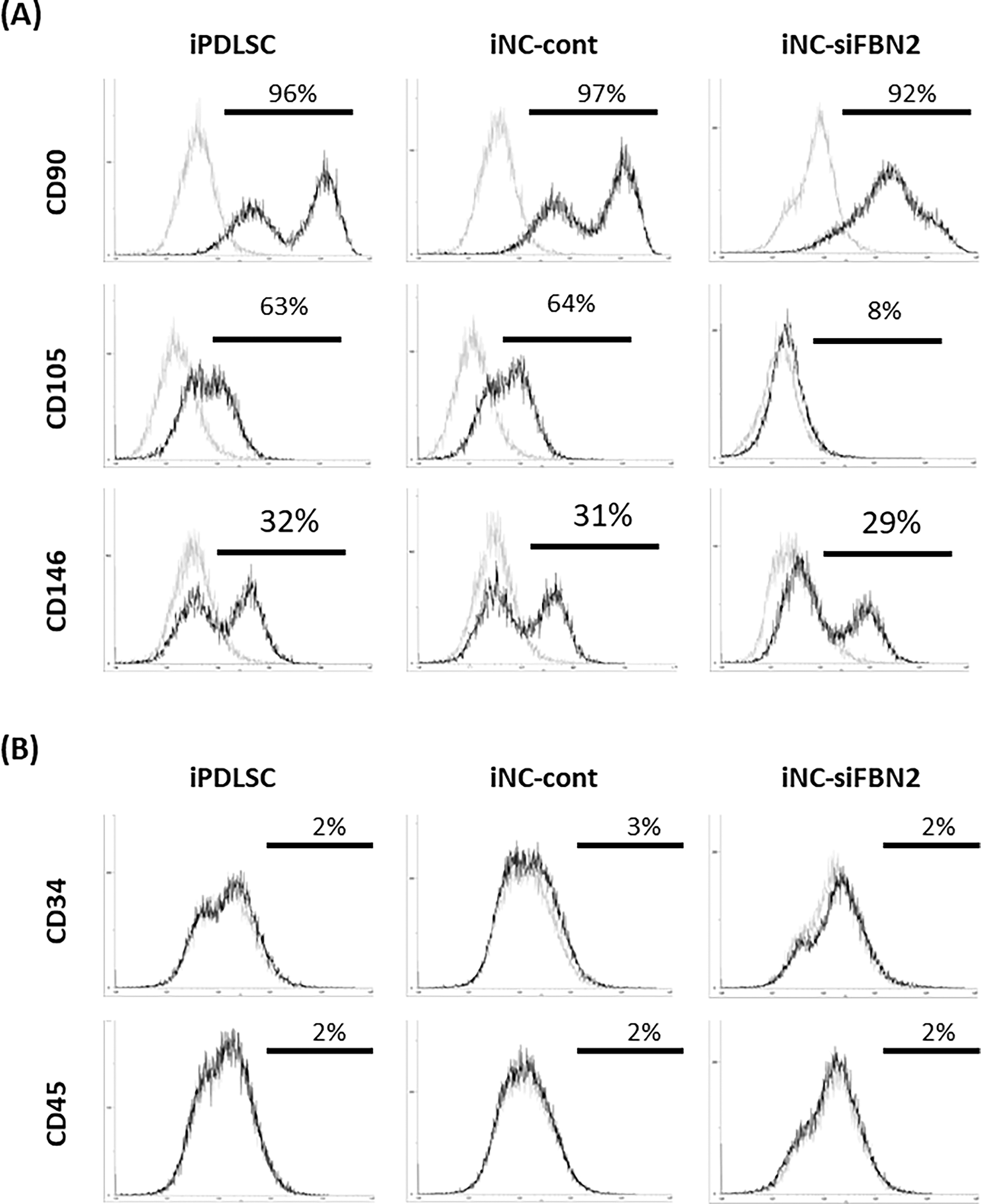
Expression of MSC-related markers in iPDLSCs, iNC-cont, and iNC-siFBN2 cells.
Proliferation assay of iPDLSCs, iNC-Cont, and iNC-siFBN2 cells
To evaluate the stem cell properties of iNC-siFBN2, we measured their proliferative ability using a WST-1 assay. Briefly, we cultured iPDLSCs, iNC-Cont, and iNC-siFBN2 cells for 3 days and measured their proliferative ability. We found that iNC-siFBN2 cells exhibited lower proliferation than the other cells (Fig. 4).
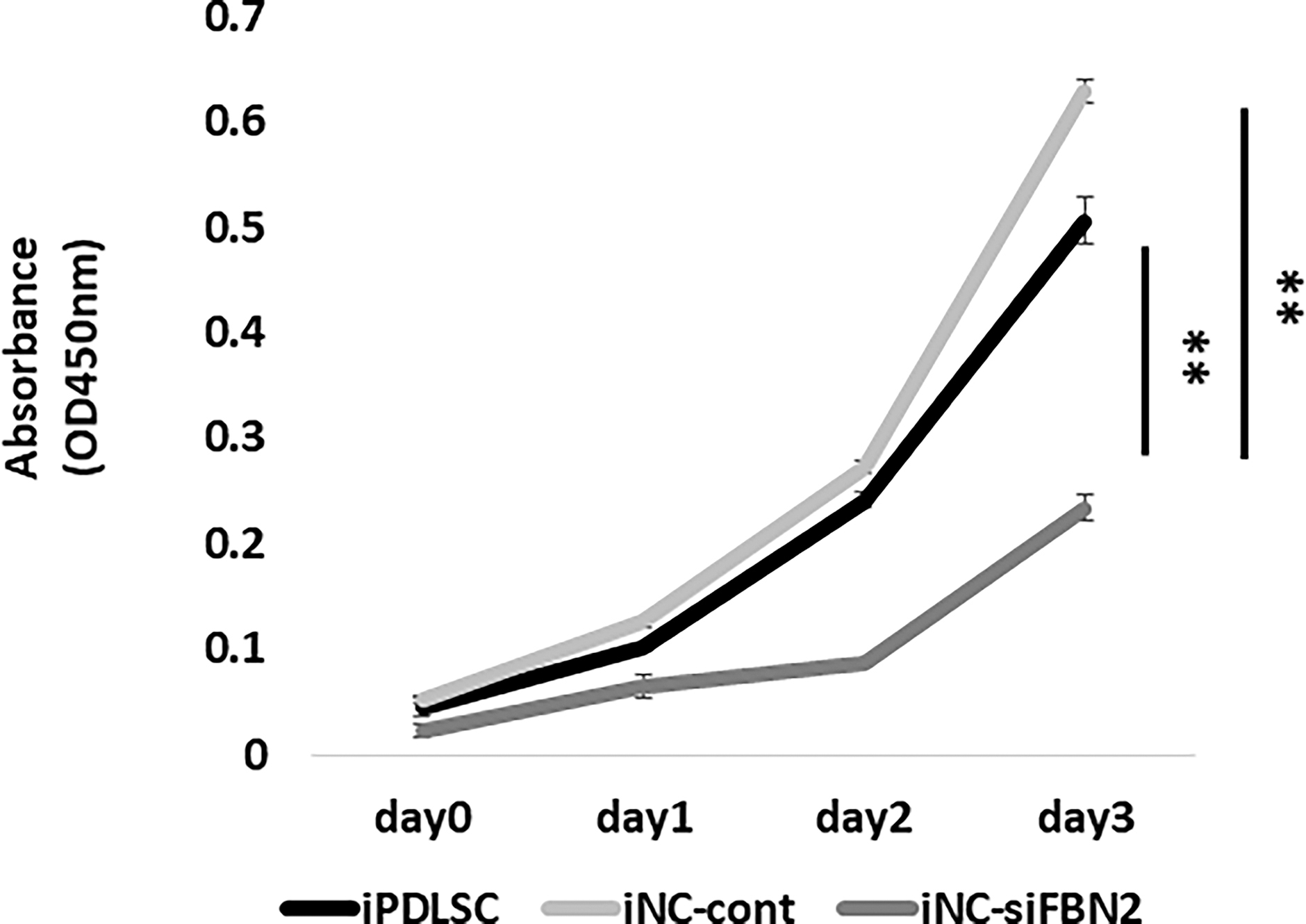
Proliferation of iPDLSCs, iNC-cont, and iNC-siFBN2. Proliferation of iPDLSCs, iNC-cont, and iNC-siFBN2 cells was examined by WST-1 assay. Data are portrayed as the mean ± standard deviation (n = 4). **P < 0.01.
Osteoblastic and adipocytic differentiation of iPDLSCs, iNC-Cont, and iNC-siFBN2 cells
To evaluate the multipotency of cells cultured in osteoblastic or adipocytic differentiation medium, we first cultured iPDLSCs, iNC-Cont, and iNC-siFBN2 cells in control medium (CM) or OM. After 3 weeks, we stained the cells using Alizarin Red S (Fig. 5A and B). iPDLSCs and iNC-Cont cells cultured in OM showed strong staining with Alizarin Red S, whereas iNC-siFBN2 cells cultured in OM had significantly fewer Alizarin Red S-positive areas (Fig. 5A and B). Likewise, expression of osteoblast-related genes BMP2 and OCN was lower in iNC-siFBN2 cells than in the other cell types (Fig. 5C).
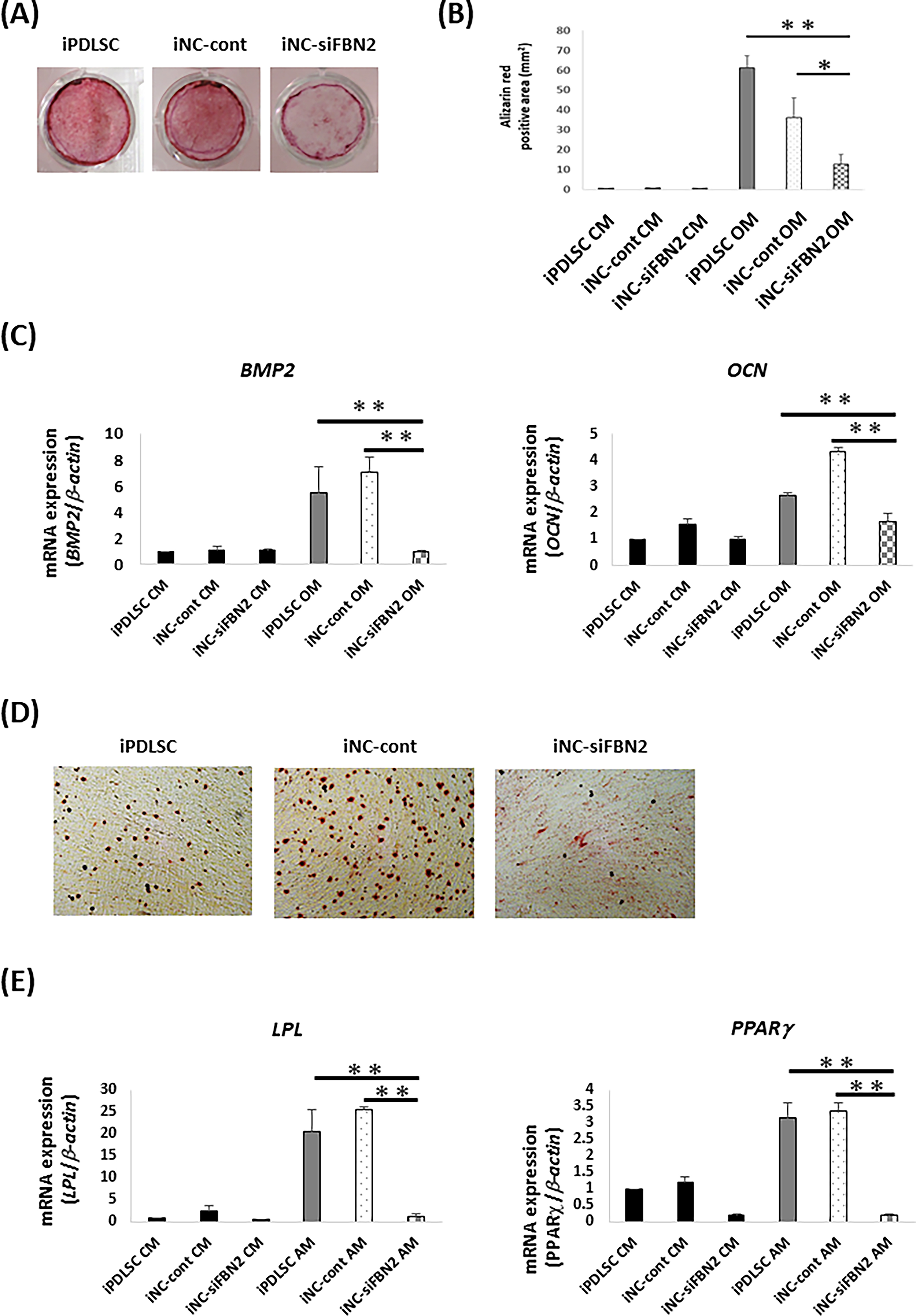
Osteoblastic and adipocytic differentiation in iPDLSCs, iNC-cont, and iNC-siFBN2 cells.
iPDLSCs, iNC-cont, and iNC-siFBN2 cells were also cultured in CM or AM. After 4 weeks, we stained the cells with Oil Red O to test for the production of fatty lipids (Fig. 5D). We found that iPDLSCs and iNC-cont cells cultured in AM produced many fatty lipids, whereas there was no production of Oil Red O-positive lipids in iNC-siFBN2 cells cultured in AM. Likewise, expression of adipocyte-related genes LPL and PPARγ was lower in iNC-siFBN2 cells than in the other cells (Fig. 5E).
Expression of PDL-related markers in iNC-FBN2 cells
Finally, we sought to investigate the ability of recombinant FBN2 protein to induce iPDLSCs from iNCs by culturing iNCs on FBN2 recombinant protein for 2 weeks (Fig. 6A). We found that expression of PDL-related genes in iNCs cultured on recombinant FBN2 protein at all concentrations was significantly lower than in iPDLSCs (Fig. 6B). Overall, FBN2 protein alone could not differentiate iNCs into iPDLSCs.
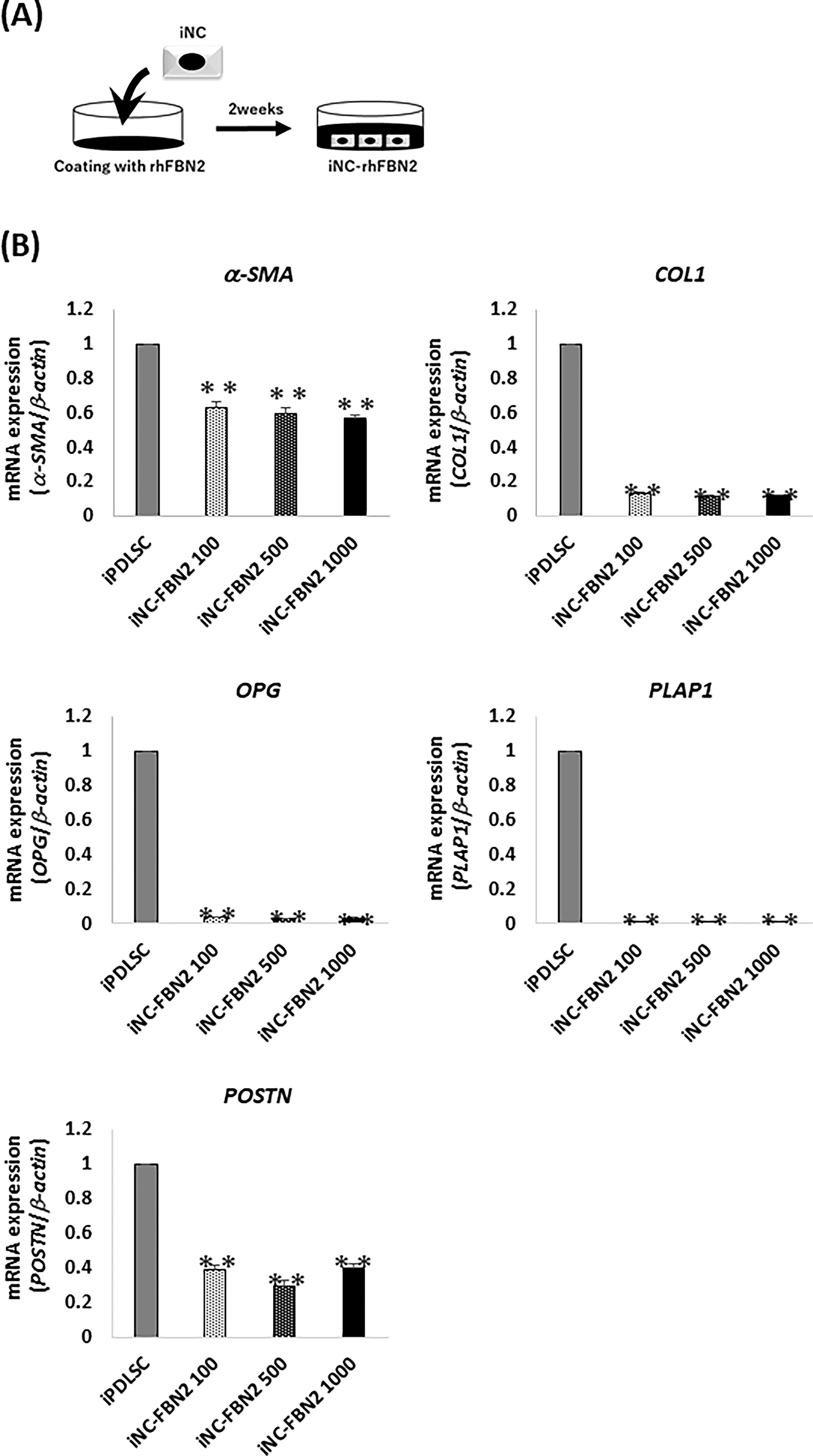
Expression of PDL-related markers in iPDLSCs and iNC-rhFBN2 cells.
Discussion
iPDLSCs had similar characteristics to primary PDLSCs in vitro 21 and the ability to form periodontal tissue-like structures in vivo (data not shown), similar to previous reports. 16,26,27 Since ECM derived from HPDLCs was required to differentiate iPDLSCs, we considered that ECM derived from HPDLC contained PDLSC-inducing factor. Therefore, the purpose of this study was to identify PDLSC-inducing factor contained in ECM derived from HPDLCs.
ECM is important in the differentiation of various cell types, such as skeletal muscle cells, hematopoietic stem cells, and dental pulp stem cells. 28 –30 In a previous study, we reported that ECM derived from HPDLC-3U has a higher ability to differentiate iPDLSCs compared with ECM derived from SFs and HPDLC-3S. Although HPDLCs and SFs are both fibroblasts, they are derived from different tissues. 21 In this study, we show that HPDLCs and SFs produce different ECMs. Individual differences in the ECM produced by HPDLCs were previously reported. Indeed, several studies have demonstrated that the ECM produced by cells derived from PDL tissues differs depending on age and mechanical force. 31 –33 Therefore, it was suggested that differences in ECM produced by HPDLC-3U, HPDLC-3S, and SFs affect the differentiation ability of iPDLSCs.
FBN2, one of the ECMs that constitutes the microfibrils of connective tissue, is reportedly involved in the formation of elastic fiber structures. 34 Moreover, FBN2 has been implicated in the maturation of fibroblasts, such as ovarian fibroblasts and dermal fibroblasts. 35,36 Tatsuoka et al. reported that FBN2 was expressed in PDL tissues during the formation of mouse tooth germs. 37 However, the effect of FBN2 on NC differentiation was unclear. In this study, we demonstrate for the first time that FBN2 is required for iNCs to acquire PDL-like characteristics.
Although FBN2 is also involved in the maturation and differentiation of various stem cells other than fibroblast lineages, 38,39 no previous report indicates that FBN2 is directly involved in CD105 expression. However, a previous report demonstrated that FBN2 suppressed transforming growth factor (TGF)-β signals during bone remodeling. 40 Moreover, Sánchez-Elsner et al. clarified that TGF-β stimulation increased CD105 expression in human vascular endothelial cells. 41 Therefore, these results suggest that FBN2 might regulate CD105 expression by controlling TGF-β signaling during the differentiation of iNCs into iPDLSCs.
CD105 is a classic surface marker expressed on various MSCs, such as bone marrow stem cells, umbilical cord stem cells, and PDLSCs, 42 –44 that reportedly decreases with cell differentiation. 45 Lee et al. revealed that, among dermal fibroblasts, CD105-positive cells had higher multipotency and proliferation ability than CD105-negative cells. 46 These reports suggest that iNC-siFBN2 cells have lower multipotency and proliferation ability compared with iPDLSCs because of lower expression of CD105. Therefore, FBN2 in HPDLCs may support the differentiation of iNCs into iPDLSCs by controlling the expression of CD105.
In this study, we could not differentiate iNCs into iPDLSCs using FBN2 coating alone. Indeed, the function of ECM is reportedly enhanced not by itself but by the combination of multiple proteins. 47 Wang et al. revealed that neural stem cells cultured on collagen scaffolds containing hyaluronic acid had higher proliferation and neural differentiation abilities compared with collagen scaffolds without hyaluronic acid. 48 Therefore, it was suggested that differentiating iNCs into iPDLSCs using only FBN2 is difficult and requires other ECMs. In the future, we should clarify all ECMs required for iPDLSC differentiation and establish a xeno-free iPDLSC differentiation protocol without using HPDLCs.
Conclusion
iNCs cultured on HPDLC-derived ECM in which FBN2 was knocked down exhibited decreased PDLSC properties. Our results show that FBN2 in the ECM derived from HPDLCs is an important factor in iPDLSC differentiation.
Footnotes
Acknowledgments
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was financially supported by Grants-in-Aid for Scientific Research (Project Nos. JP21K09876, JP22H03270, JP22K17061, JP22K21018, JP23K09171) from the Japan Society for the Promotion of Science.
Supplementary Material
Supplementary Figure S1