
Abstract
Exosomes derived from mesenchymal stem cells (MSCs) show therapeutic potential despite limited yield. In contrast, the yield of exosome-mimetic vesicles (EMVs), which share a similar structure and size with exosomes is high. Previous studies have compared their proteomic profiles, and the microRNA (miRNA) expression signatures remain unexplored. EMVs from human bone marrow MSCs were isolated through continuous extrusion and exosomes were isolated from the supernatant via differential ultracentrifugation. miRNA sequencing was performed using high-throughput sequencing, and the miRNA expression profiles of MSC-EMVs, MSC-exosomes, and MSCs were compared. Following the comparison of differentially expressed miRNAs in MSC‐EMVs and MSC-exosomes, target gene prediction and functional enrichment analyses were performed. Furthermore, a trend analysis was conducted on the miRNA expression levels in the three groups to further explore the relationship between miRNA expression levels. Our study confirmed that EMVs could be stably produced and that their yield was approximately 100-fold higher than that of exosomes. A total of 763 known miRNAs were identified through comparison using the miRBase library. The miRNAs in EMVs and exosomes overlapped with those in MSCs; however, EMVs shared more miRNAs with the parent cells. Comparative analysis identified 21 upregulated and 17 downregulated miRNAs in EMVs versus exosomes, while trend analysis revealed 108 miRNAs preferentially expressed in MSCs and EMVs. Functional enrichment of differentially expressed miRNAs provides new insights for EMV-based therapies. Importantly, we demonstrated that both MSC-EMVs and MSC-exosomes significantly attenuated LPS-induced inflammation in THP-1 macrophages by modulating cytokine secretion (ELISA), suppressing iNOS expression (immunofluorescence), and inhibiting NF-κB activation (western blot). In an lipopolysaccharide (LPS)-induced acute kidney injury model, both vesicle types effectively reduced renal inflammation and tissue damage (histopathology and protein analysis). Our findings not only present the first comprehensive miRNA profiling comparison between MSC-derived EMVs and exosomes but also validate their comparable anti-inflammatory efficacy, supporting EMVs as a viable high-yield alternative for cell-free therapies.
Introduction
Mesenchymal stem cells (MSCs) are multipotent stromal cells derived from the mesoderm; they can differentiate into osteoblasts, chondrocytes, or adipocytes. Studies have demonstrated their enormous therapeutic potential in biomedical research. 1 –4 However, cell therapy has limitations such as immunogenic risk, leading to tumor development or tumor metastasis. 5 In addition, recent studies have shown that the beneficial effects of MSCs are associated with their secretory substances rather than from their integration into tissues or ability to differentiate into necessary cells of various types. 6 –8 Therefore, extensive research is being conducted on cell-free therapies, such as those utilizing MSC-derived exosomes (MSC-exosomes). MSC-exosomes are endogenous nanoparticles secreted autonomously by MSCs that carry a variety of genetic materials including microRNAs (miRNAs), mRNAs, and proteins. 9 MSC-exosomes have been demonstrated to play a therapeutic role in various disease models of different organ systems and in fields such as regenerative medicine. 10 –16 However, the main obstacles to the clinical application of exosomes include their limited production from donor cells and complex purification processes.
To address this issue, the concept of exosome mimetic vesicles (EMVs) has been proposed. EMVs are artificial vesicles that were first prepared by Jang et al. in 2013; 17 they mimic exosomes in morphology, size, and composition. 18 EMVs are produced by the continuous extrusion of cells in a mini-extruder with polycarbonate membrane filters of different sizes. The yield of EMVs and their RNAs and protein expression levels are several-fold higher than those of exosomes. 17,19 MSC-derived EMVs (MSC-EMVs) have been shown to play therapeutic roles in various diseases, such as the repair of spinal cord and heart injuries, inhibition of osteoblast apoptosis, and targeted delivery of chemotherapy drugs for malignancies. 20 –27 Therefore, EMVs are expected to become a promising substitute for exosomes in cell-free therapy and are worthy of in-depth study.
miRNAs are small noncoding RNAs that post-transcriptionally regulate gene expression via mRNA degradation or translation inhibition. miRNAs are considered key regulators of the self-renewal ability, pluripotency maintenance, proliferation, and differentiation potential of stem cells by directly targeting the pluripotency and transcriptional factors implicated in the above processes. 28 Exosomal miRNAs have been shown to have diagnostic and predictive effects in various diseases. 29,30 However, comparative analyses of the miRNA profile between MSC-EMVs and MSC-exosomes remain limited. Similarly, no studies have directly compared the miRNA expression profiles of MSC‐EMVs, MSC-exosomes, and MSCs. Therefore, we performed miRNA sequencing to compare the miRNA expression profiles of MSC-EMVs, MSC-exosomes, and MSCs. We hypothesize that MSC-EMVs, while artificially generated, contain genetic cargo profiles comparable with natural exosomes and exhibit at least 5-fold higher production yields, suggesting their potential as functionally equivalent yet more scalable alternatives for cell-free therapies.
Materials and Methods
Cell culture
Human bone marrow-derived mesenchymal stem cells (hBM-MSCs) purchased from ScienCell Research Laboratories (San Diego, California) were isolated from human bone marrow. The hBM-MSCs were validated by flow cytometry, showing positive expression of CD73, CD90, and CD105 (>95%), and negative for hematopoietic markers (e.g., CD34, CD45), confirming their identity as MSCs. The hBM-MSCs were cultured in mesenchymal stem cell medium (MSCM, Cat. #7501) containing 10% fetal bovine serum (FBS) and 5% penicillin/streptomycin at 37°C with 5% CO2. The cells were maintained in MSCM supplemented with 10% FBS and incubated at 37°C with 5% CO2 to generate EMVs. To generate exosomes, the cells were cultured in MSCM with 10% exosome-free FBS at 37°C with 5% CO2 for 48 h; subsequently, the supernatant was collected for further experiments.
EMV generation
EMVs were generated following the protocol reported in a previous study. 17 Briefly, MSCs were harvested and resuspended in phosphate-buffered saline (PBS) at a concentration of 1 × 106/mL. The cell suspension was serially extruded five times through 10-, 5-, and 1-μm polycarbonate membrane filters (Nuclepore, Whatman, Inc., Clifton, NJ) in a mini-extruder (Avanti Polar Lipids, Birmingham, AL). The EMVs were further purified and collected using OptiPrep density gradient ultracentrifugation. Briefly, 50% iodixanol (1 mL, Axis-Shield PoC AS, Oslo, Norway) and 10% iodixanol (2 mL) were added to these samples to perform the gradient, and then ultracentrifuged at 100,000g for 2 h at 4°C after adding the sample (7 mL) in the ultracentrifuge tube. The EMVs were obtained from the interface of the 50% and 10% iodixanol layers. After washing thrice with PBS, the EMVs were resuspended in PBS and filtered through a 0.22-μm filter.
Exosome isolation
Exosomes were isolated from the supernatant by differential ultracentrifugation as previously described. 31 Briefly, the supernatant was collected and centrifuged to sediment dead cells (300g, 10 min), remove cellular debris (2,000g, 10 min), and dispose of large microvesicles (10,000g, 30 min). Afterward, the resultant supernatant was ultracentrifuged at 100,000g for 70 min at 4°C to pellet exosomes. Subsequently, the exosome pellets were resuspended in PBS and centrifuged again at 100,000g for 70 min at 4°C. Finally, the exosome pellets were resuspended in PBS and then filtered (with 0.22-μm filters) to remove large particles and ultimately obtain exosomes.
Transmission electron microscopy
Exosomes and EMV particles were evaluated using transmission electron microscopy (TEM). First, 10 µL of each sample was added to 200 ultra-thin carbon-coated copper mesh for 2 min, which was then quickly dried on filter paper. Then, 1% uranyl acetate (filtered through a 0.22-μm filter) was used to perform negative coloring on the grid. After completely drying for 1 min, imaging was performed using a Hitachi HT-7700 TEM at 100 kV.
Nanoparticle tracking analysis
Nanoparticle tracking was performed using a Flow Nano Analyzer. This system was calibrated using 200-nm polystyrene beads with a defined concentration of 2.01 × 1010 particles/mL, which was also used as a reference for particle concentration. In addition, four different sizes of monodisperse silica beads (68, 91, 113, and 155 nm) were used as reference standards to calibrate the size of exosomes. Newly filtered (0.22 μm) PBS was analyzed as a background signal and subtracted from the other measured values. The exosome sample was diluted with filtered PBS, maintaining the particle count within the optimal range of 2,500–12,000, and sample data were collected at a sample pressure of 1.0 kPa for 1 min. The particle concentration and particle size distribution were calculated using the NanoFCM software NF professional v1.08.
Western blot analysis
Total proteins were extracted from cells, exosomes, and EMVs using RIPA lysis buffer (containing protease and phosphatase inhibitors) followed by centrifugation (12,000g, 10 min, 4°C). Protein concentrations were determined by the BCA assay, and equal amounts (40 μg/lane) were resolved by 10% SDS-PAGE. For THP-1 macrophage experiments, proteins were transferred to PVDF membranes, while nitrocellulose membranes were used for vesicle characterization. Membranes were blocked with 5% skim milk/TBST (1 h, RT) and probed overnight (4°C) with primary antibodies against: (1) exosomal markers (CD63, TSG101; 1:1,000) and Golgi control (GM130; 1:1,000) for vesicle validation; or (2) phospho-p65 (1:1,000), total p65 (1:1,000), and loading controls (β-actin 1:5,000 or GAPDH 1:3,000) for signaling analysis. After TBST washes, HRP-conjugated secondary antibodies (1:5,000) were applied (1 h, RT). Protein bands were visualized by enhanced chemiluminescence and quantified using Image Lab (Bio-Rad) or Quantity One software, with normalization to respective loading controls.
Library construction and miRNA sequencing
Total RNA was extracted using a TRIzol reagent kit (Invitrogen, Carlsbad, CA, USA), followed by enrichment of RNA molecules ranging in size from 18 to 30 nt using polyacrylamide gel electrophoresis. Subsequently, 3′ and 5′ adapters were ligated to the RNAs, and the ligation products were reverse transcribed by polymerase chain reaction (PCR) amplification. PCR products ranging from 140 to 160 bases were then enriched to generate a cDNA library. The library was sequenced using the Illumina Novaseq 6000 platform by Gene Denovo Biotechnology Co. (Guangzhou, China). All clean tags were aligned with small RNAs in the GeneBank (Release 209.0) and Rfam (Release 11.0) databases to identify and remove ribosomal RNAs, small conditional RNAs, small nucleolar RNAs, small nuclear RNAs, and transfer RNAs. Subsequently, the clean tags were aligned with the reference genome, and those mapping to exons, introns, and repeat sequences were excluded. The remaining clean tags were then searched against the miRBase database (Release 22) to identify known (Species studied) miRNAs (existing miRNAs). The expression level of miRNA was quantified and normalized to transcripts per million (TPM) using the following formula:
Target gene prediction and functional enrichment analysis
Target genes were predicted using Miranda (v3.3a) and TargetScan (v7.0). The Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses were performed using DAVID Bioinformatics Resources (http://david.abcc.ncifcrf.gov/, DAVID Bioinformatics Resources 6.8). The significantly enriched GO terms or pathways were identified based on the criterion of P < 0.05.
Trend analysis
The expression patterns of three groups (MSC-EMVs, MSC‐exosomes, and MSCs) of miRNAs were analyzed using the Short Time-series Expression Miner (STEM, v1.3.8). The union set of all the miRNAs was profiled into eight expression patterns. Among them, the clustered profiles with P < 0.05 were considered as significantly enriched. Then, GO and KEGG analyses were conducted on biologically significant trends to elucidate the biological relationships between miRNA target genes.
Reverse transcription-quantitative PCR
Total miRNA was reverse-transcribed to cDNA using the Qiagen miScript II RT kit (miRcute miRNA First-Strand cDNA Synthesis Kit; TianGen, China) following the manufacturer’s instructions. Quantitative PCR was performed using the Qiagen miScript SYBR Green PCR kit according to the manufacturer’s instructions. Reactions were performed using the StepOne Plus Real-Time PCR System (Applied Biosystems) and StepOne software v2.1. The miRNA primers used in this study were purchased from FulenGen (Guangdong, China). U6 was used as the internal reference. Gene expression was estimated using the 2−ΔΔCq (Livak) method. Each experiment was repeated thrice to test reproducibility.
Anti-inflammatory experiments in vitro and in vivo
Labeling and tracing of exosomes and EMVs in THP-1 macrophages
The EMVs and exosomes were labeled with the lipophilic fluorescent dye DiD according to the manufacturer’s protocol. THP-1 macrophages were seeded in 24-well plates at a density of 2 × 105 cells/well and differentiated with 100 nM PMA for 48 h. DiD-labeled EMVs and exosomes (100 μg/mL) were added to the cultured macrophages. Following incubation for 6 h, 12 h, and 24 h at 37°C with 5% CO2, cells were washed three times with PBS, fixed with 4% paraformaldehyde for 15 min, and counterstained with DAPI (1 μg/mL, 5 min) for nuclear visualization. Cellular uptake was examined using a fluorescence microscope (Nikon Eclipse Ti, Japan) with consistent exposure settings across samples.
LPS stimulation in THP-1 macrophages
THP-1 monocytes were cultured in RPMI-1640 medium supplemented with 10% FBS and differentiated into macrophages by treatment with 100 nM PMA for 48 h. The differentiated macrophages were then stimulated with 500 ng/mL lipopolysaccharide (LPS) (E. coli O111:B4) for 24 h to induce inflammatory responses. For treatment groups, MSC-EMVs and MSC-exosomes were added at a concentration of 100 μg/mL simultaneously with LPS stimulation. All cells were maintained at 37°C in a humidified 5% CO2 incubator.
ELISA analysis
Following 24 h of treatment, cell culture supernatants were collected and centrifuged at 300×g for 5 min to remove cellular debris. The levels of pro-inflammatory cytokines (TNF-α, IL-6) and anti-inflammatory cytokines (IL-10, TGF-β) in the supernatants were quantified using commercial ELISA kits (R&D Systems) according to the manufacturer’s protocols. Absorbance was measured at 450 nm using a microplate reader, and cytokine concentrations were determined by comparison with standard curves.
Immunofluorescence (iNOS)
For iNOS detection, THP-1 macrophages were fixed with 4% paraformaldehyde for 15 min at room temperature, followed by permeabilization with 0.1% Triton X-100 for 10 min. After blocking with 5% BSA for 1 h, the cells were incubated with anti-iNOS primary antibody (1:200 dilution) overnight at 4°C, followed by Alexa Fluor 488-conjugated secondary antibody for 1 h at room temperature. Nuclei were counterstained with DAPI (1 μg/mL) for 5 min. Fluorescence images were captured using a Nikon Eclipse Ti microscope with consistent exposure settings across all samples.
Animals and treatment
Male C57BL/6 mice (8 weeks old) were randomly divided into four experimental groups (n = 6 per group): (1) control group receiving PBS, (2) LPS group (5 mg/kg, intraperitoneal injection), (3) LPS + MSC-EMV group (1 × 1010 particles/mouse, intravenous injection), and (4) LPS + MSC-Exo group (1 × 1010 particles/mouse, intravenous injection). MSC-derived vesicles were administered 1 h after LPS injection. All animals were sacrificed 24 h post-LPS administration, and kidney tissues were collected for subsequent histopathological and molecular analyses.
Statistical analyses
Differential miRNA expression between two groups was analyzed using the edgeR software. miRNAs with a fold change of >2 and a P value of <0.05 were considered statistically significant in our comparisons. Each experiment was performed in triplicate for technical replicates. All data are presented as mean ± standard deviation. Statistical analyses were performed using the one-way analysis of variance. Differences were considered statistically significant at P < 0.05.
Ethics approval and consent to participate
Title of the approved project: Exosome mimetic vesicles derived from human bone marrow mesenchymal stem cells alleviate acute kidney injury in a cardiopulmonary bypass rat model. Name of the institutional approval committee: Laboratory Animal Welfare Ethics Committee of Beijing Tongren Hospital. Approval Number: TRLAWEC2022-03. Date of approval: May 17, 2022.
Results
Extraction of EMVs and exosomes
EMVs were extracted from MSCs and exosomes were isolated from the supernatant of MSC cultures (Fig. 1). Exosomes were successfully obtained from the cell suspensions via differential ultracentrifugation (Fig. 1A). The EMVs were generated by extruding the MSCs (1 × 106) five times through polycarbonate membranes with different pore diameters (10, 5, and 1 μm), and purified using two-step OptiPrep density-gradient ultracentrifugation (Fig. 1B).
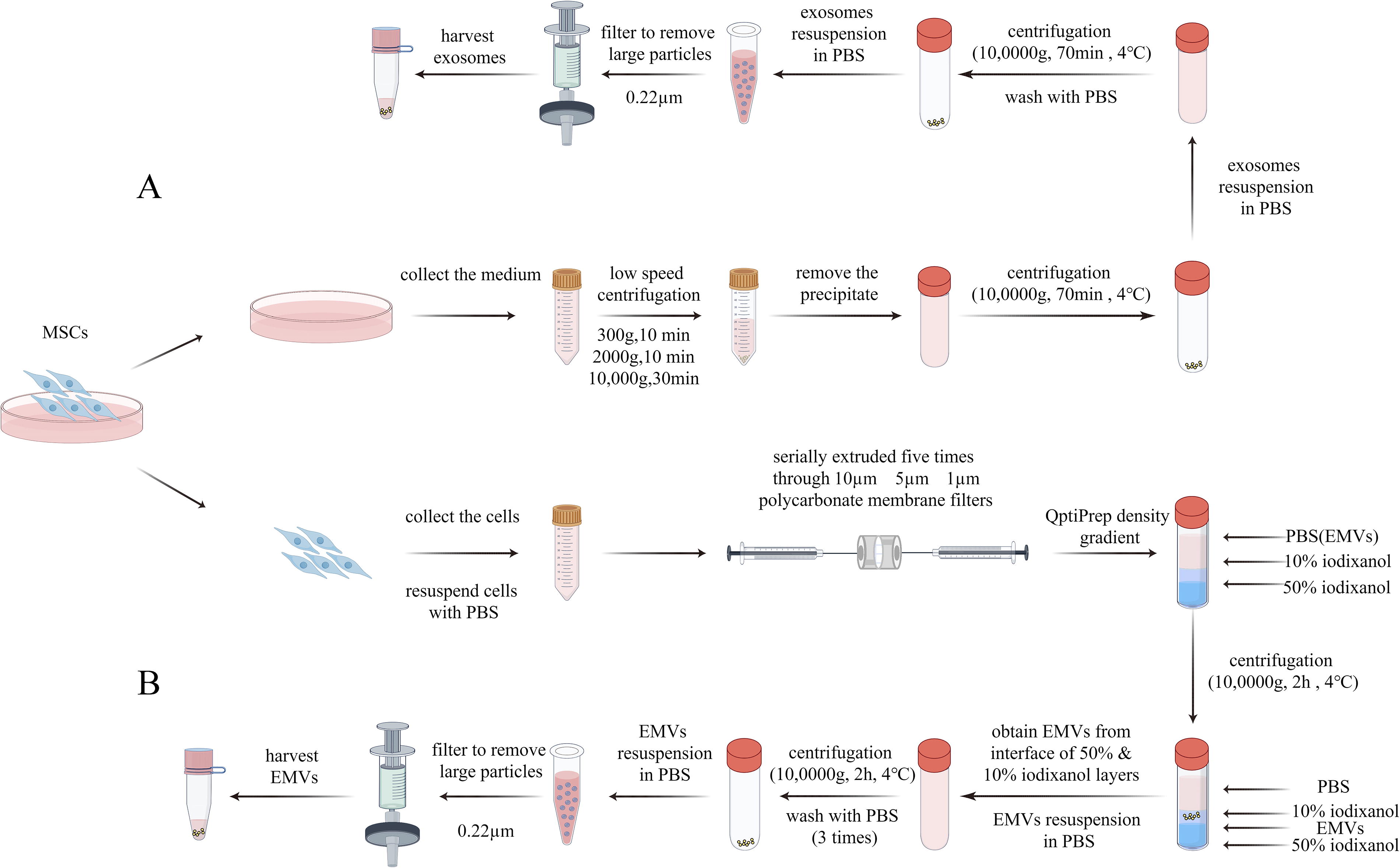
Schematic overview of MSC-exosomes and MSC–EMVs generation.
Characterization of MSC‐EMVs and MSC‐exosomes
The morphology of EMVs and exosomes was examined using TEM. The EMVs and exosomes were round or oval, enclosed in lipid membranes, and without the vesicles of the parent cells or cell fragments (Fig. 2A). Nanoparticle tracking analysis (NTA) revealed that the mean diameter of EMVs and exosomes was 76.65 ± 13.79 and 73.59 ± 13.26 nm, respectively (Fig. 2B). Western blot analyses showed that EMVs express common exosomal marker proteins such as CD63 and TSG101, as well as GM130, which is not present in exosomes (Fig. 2C). Furthermore, the total protein content in EMVs (310 μg) was more than 100-fold (average) higher than that in exosomes (3.0 μg) obtained from the same number of MSCs (1 × 106 cells) (Fig. 2D).
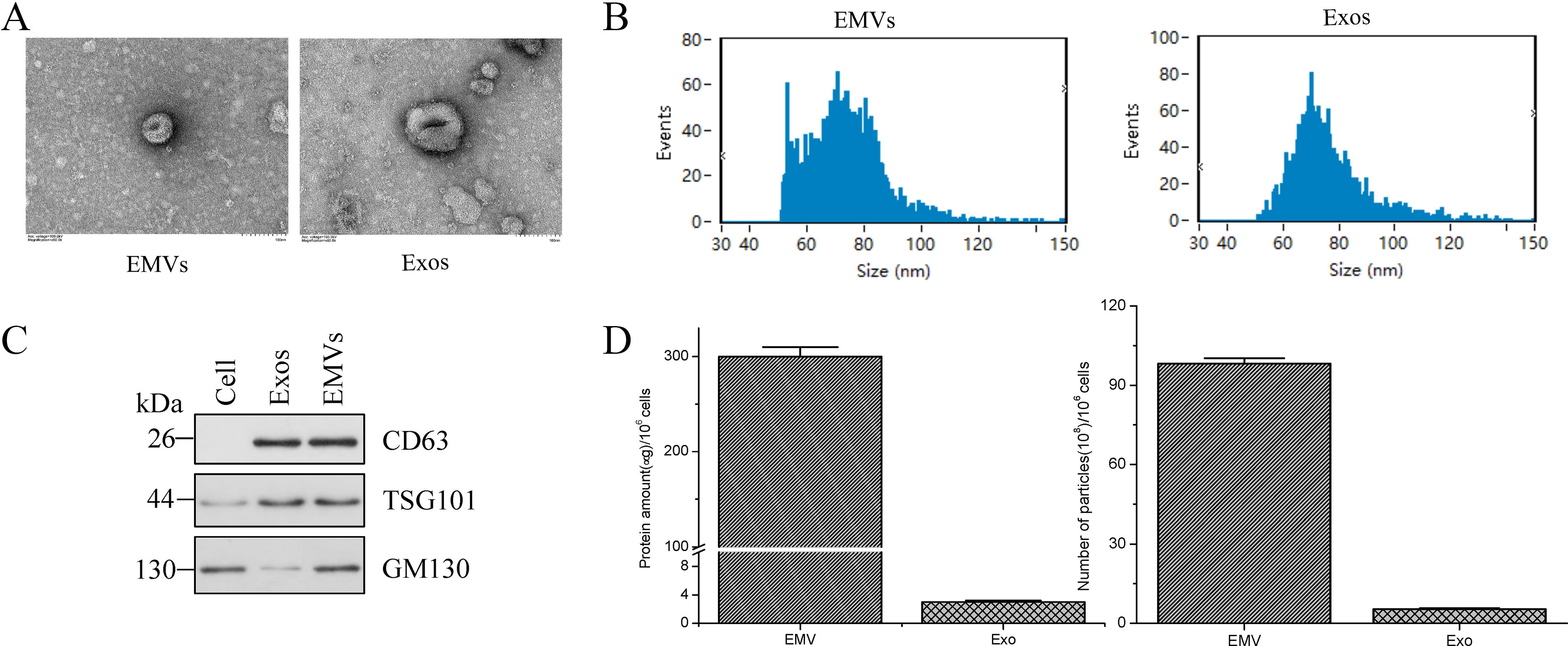
Characterization of MSC-EMVs and MSC-exosomes.
miRNA expression of MSC‐EMVs, MSC‐exosomes, and MSCs
We obtained the miRNA expression profiles of MSC-EMVs, MSC-exosomes, and MSCs using high-throughput sequencing. A total of 763 known miRNAs were identified through comparison with miRNAs in the miRBase library. miRNA profiling revealed 502 miRNAs in MSC-EMVs, 431 miRNAs in MSC-exosomes, and 706 miRNAs in parental cells. Using a Venn diagram to compare the miRNAs in the three groups, 359 miRNAs were commonly found in all groups, and 28, 22, and 198 unique miRNAs were identified in MSC-EMVs, MSC-exosomes, and MSCs, respectively (Fig. 3A). Principal component analysis showed variance within samples, indicating low variance between biological and technical replicates, and a distinct separation among MSC-EMVs, MSC-exosomes, and MSCs (Fig. 3B). The 359 shared miRNAs were hierarchically clustered into three groups, and the top 20 miRNAs with the highest expression levels and the lowest coefficients of variation were selected (Fig. 3C). The GO analysis of the shared miRNAs showed that the miRNAs in the biological process category were mainly concentrated in the cellular component organization, multicellular organismal processes, and the positive regulation of biological and developmental processes (Fig. 3D). The KEGG pathway analysis of the shared miRNAs revealed their enrichment in thermogenesis, NOD-like receptor signaling pathway, apoptosis, and lysosomes (Fig. 3E).
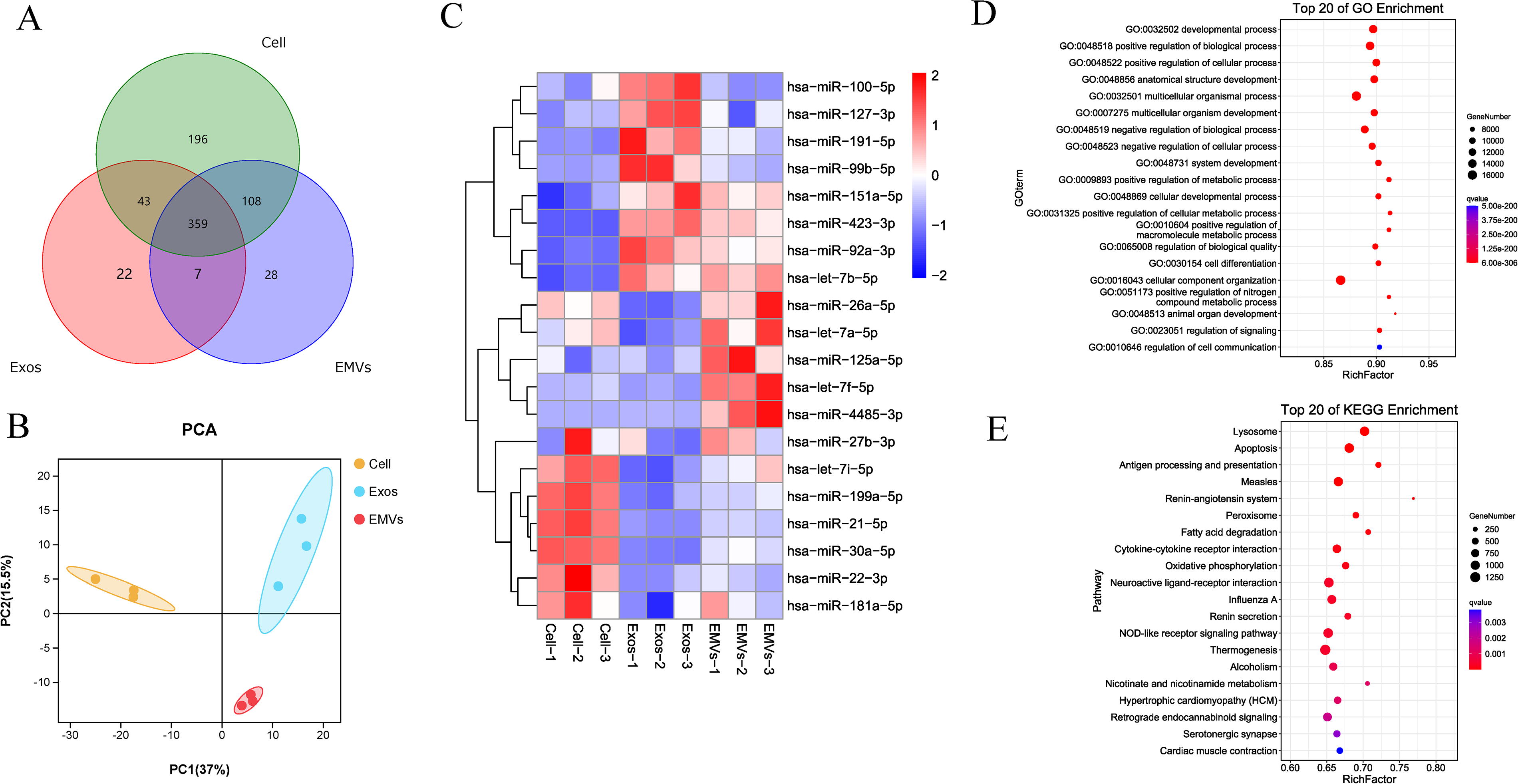
MicroRNA profiling of MSC-EMVs, MSC-exosomes, and MSCs.
Differentially expressed miRNAs in MSC‐EMVs and MSC‐exosomes
A comparison of the miRNAs in EMVs and exosomes using a volcano plot revealed 38 differentially expressed miRNAs. Compared with those in exosomes, 21 miRNAs were upregulated and 17 were downregulated in EMVs (Fig. 4A). The top 10 upregulated and downregulated miRNAs were listed as a table (Fig. 4B). GO analysis on these upregulated and downregulated miRNAs showed that most upregulated miRNAs were concentrated in the regulation of biological processes, regulation of cellular processes, regulation of response to stimulus, and multicellular organismal processes in the biological process category (Fig. 4C). However, most downregulated miRNAs in the biological process category were concentrated in the regulation of biological processes, regulation of cellular processes, multicellular organismal processes, regulation of cell communication, and nervous system development (Fig. 4E). KEGG analysis revealed the enrichment of the upregulated miRNAs in the mitogen-activated protein kinase signaling pathway, animal autophagy, the NOD-like receptor signaling pathway, and apoptosis (Fig. 4D). The downregulated miRNAs were enriched in the Ras signaling pathway, nucleotide-binding oligomerization domain (NOD)-like receptor signaling pathway, apoptosis, and the Forkhead box O signaling pathway (Fig. 4F).
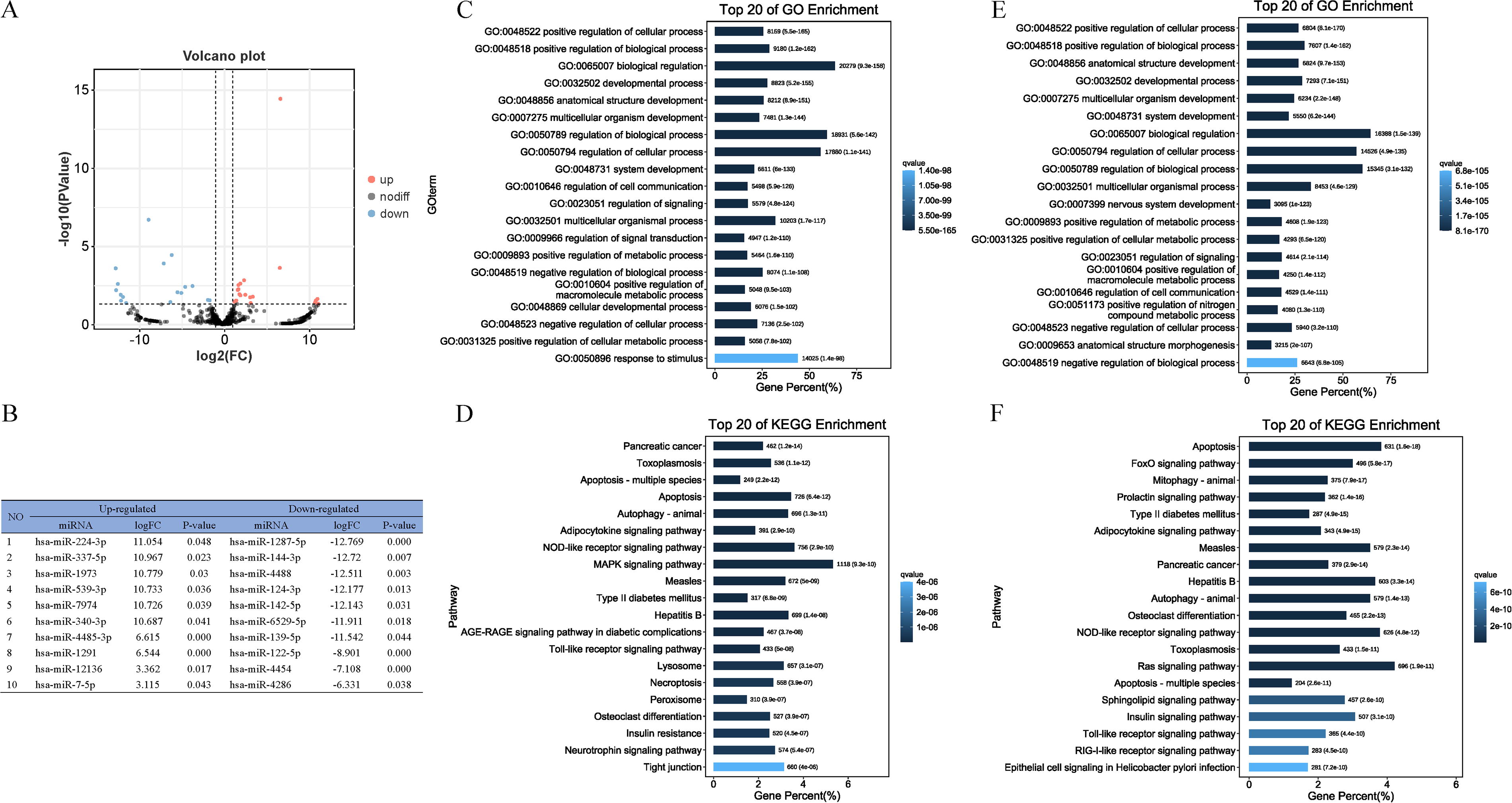
Differentially expressed miRNAs in MSC‐EMVs and MSC‐exosomes.
Trend analysis of miRNAs in MSC‐EMVs, MSC‐exosomes, and MSCs
We further analyzed the expression patterns of miRNAs using STEM. All miRNAs were categorized into eight expression trends, including two significant trends, which are displayed in color (Fig. 5A). The significant trend (profile 2) (P < 0.05) showed that the expression levels of these 108 miRNAs sharply decreased from cells to exosomes and then sharply increased in EMVs. GO analysis of the miRNAs in profile 2 showed that the miRNAs in the biological process category were mainly concentrated in cellular processes, metabolic processes, and biological regulation (Fig. 5B). KEGG pathway analysis of the miRNAs in profile 2 showed that they were enriched in many systems and diseases (Fig. 5C).
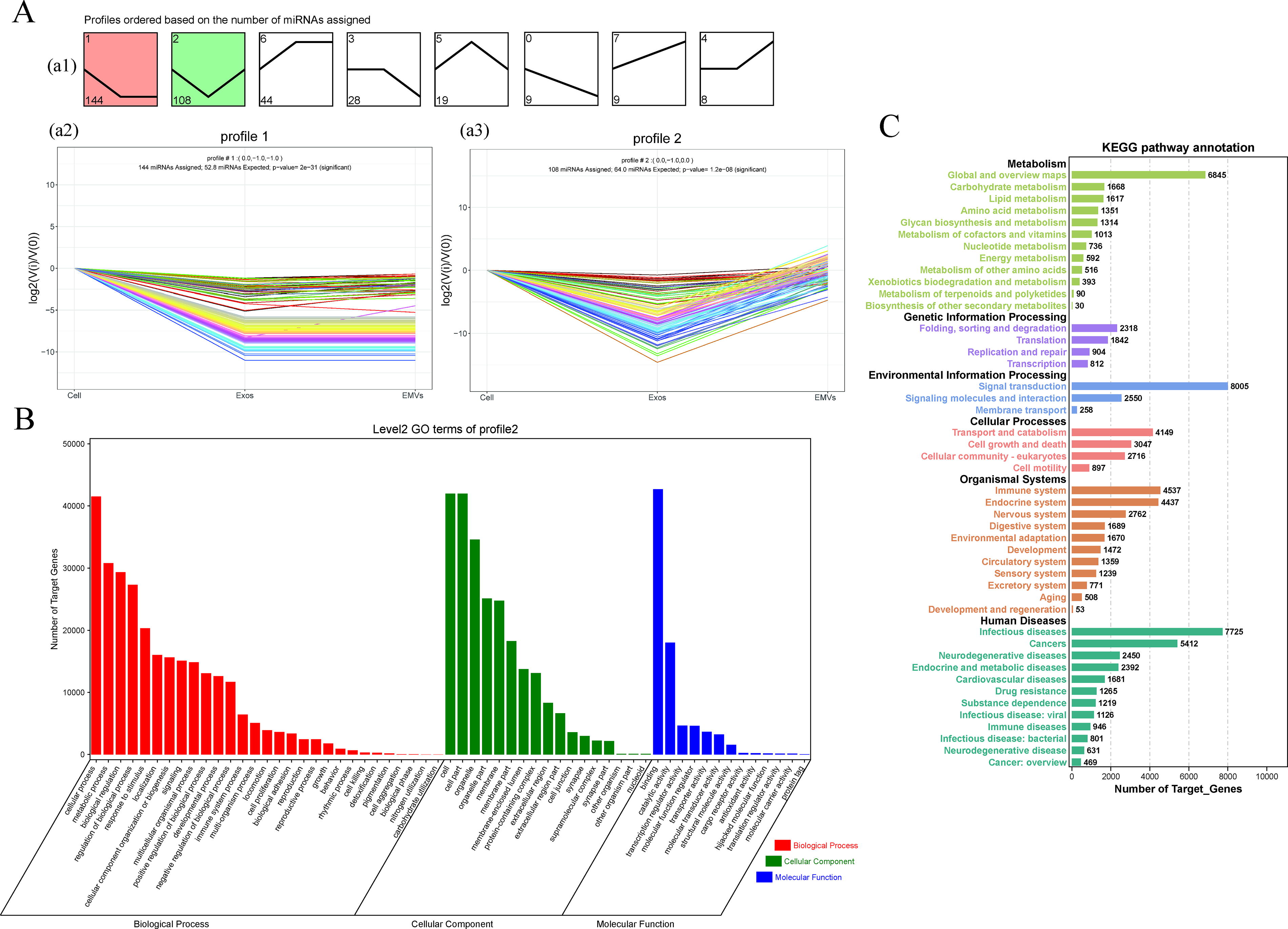
Analysis of miRNA expression trends in MSCs, MSC–EMVs, and MSC-exosomes.
Validation of differentially expressed miRNAs using RT-qPCR
RT-qPCR verification of 10 differentially expressed miRNAs was performed among MSC-EMVs, MSC-exosomes, and MSCs (Fig. 6). Compared with that in the MSC and EMV groups, the expression of miR-100-5p, miR-127-3p, miR-191-5p, and miR-99b-5p was significantly upregulated in the MSC-exosome group. In the EMV group, the expression of let-7f-5p, miR-125a-5p, and miR-4485-3p was significantly higher than that in the other two groups. However, miR-199a-5p, miR-21-5p, and miR-30a-5p expression was significantly upregulated only in the MSC group. This result was similar to the expression trends detected using miRNA-seq.
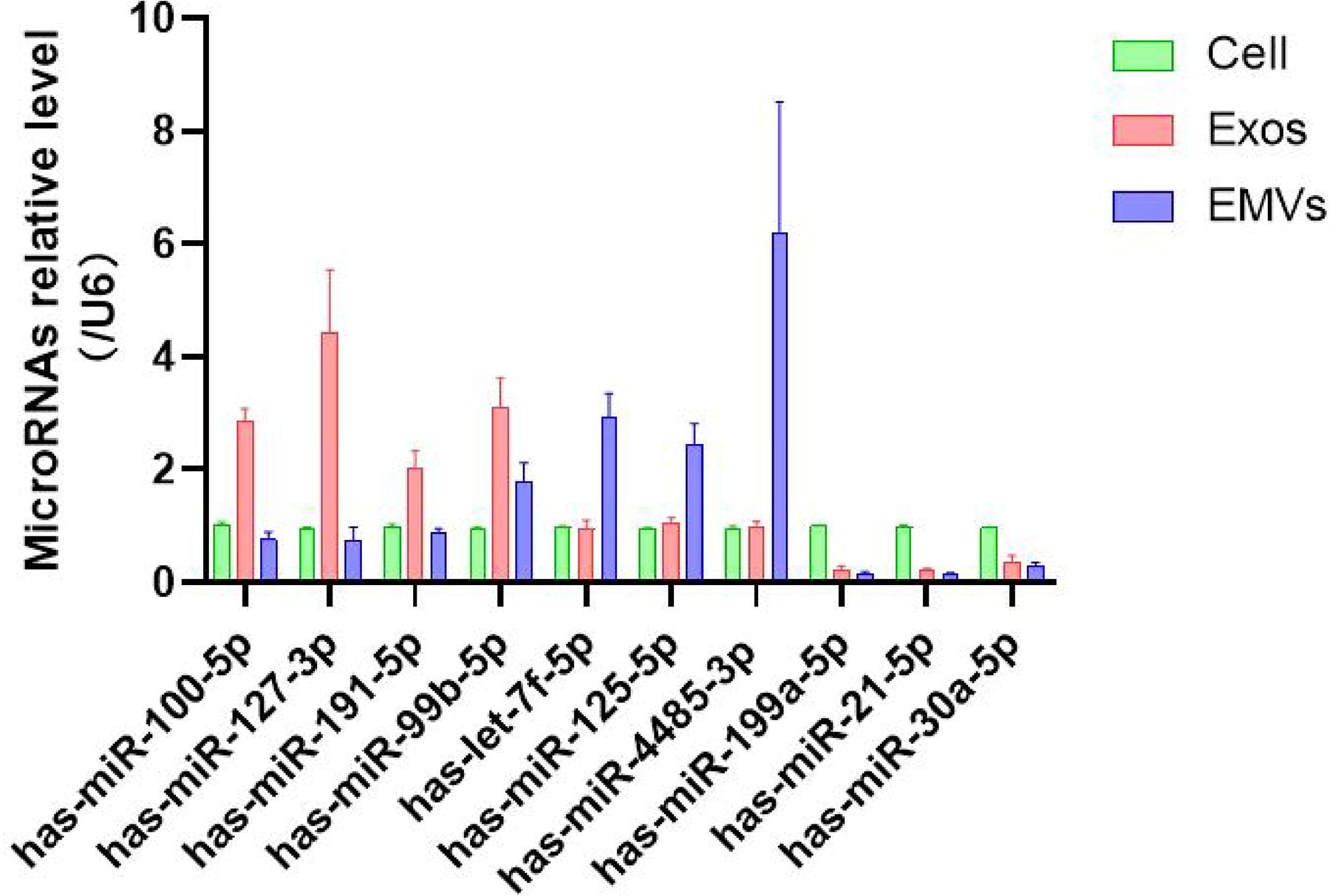
qRT-PCR validation of selected miRNAs. Data presented as mean ± SEM; miRNA, microRNA; qRT-PCR, quantitative reverse-transcription polymerase chain reaction.
THP-1 macrophages internalize MSC‐exosomes and MSC‐EMVs
We examined whether THP-1 macrophages could internalize MSC-EMVs and MSC-exosomes. The results showed that THP-1 macrophages efficiently internalized both MSC-EMVs and MSC-exosomes. At 6 h, both EMVs and exosomes were internalized by the cells, with the uptake increasing over time. Colocalization with DAPI-stained cell nuclei (blue) confirmed the internalization of EMVs and exosomes into the cells (Fig. 7A, B).
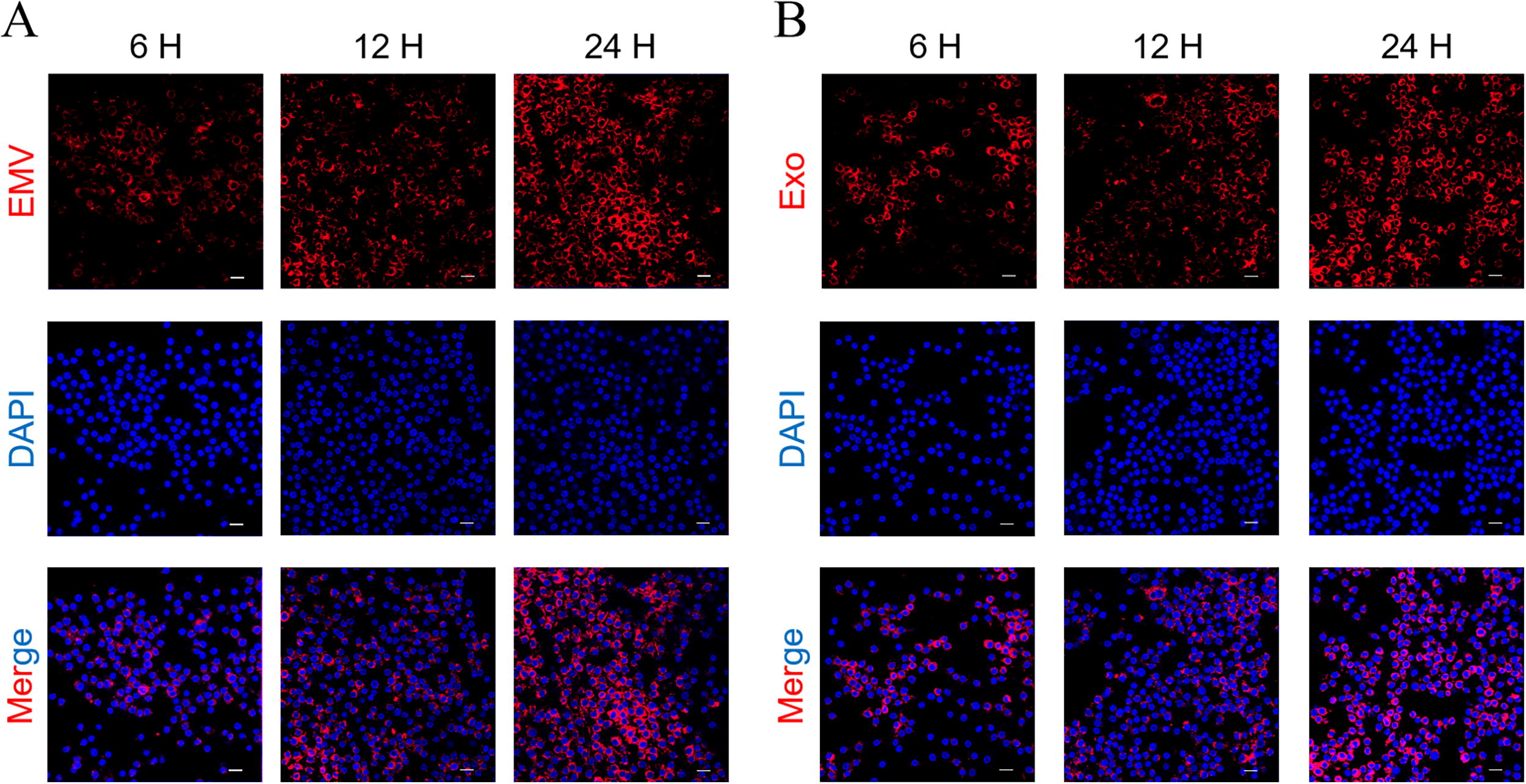
Cellular uptake of MSC-derived vesicles by THP-1 macrophages. Fluorescence microscopy images showing internalization of
MSC-EMVs and MSC-exosomes attenuate LPS-Induced inflammatory responses in THP-1 macrophages
ELISA analysis of THP-1 supernatants revealed that LPS stimulation (500 ng/mL, 24 h) significantly increased the secretion of pro-inflammatory cytokines TNF-α and IL-6 compared with the control (P < 0.0001). Treatment with either MSC-EMVs or MSC-exosomes significantly attenuated these LPS-induced elevations (P < 0.0001 vs. LPS group). Conversely, while LPS alone did not affect anti-inflammatory cytokine levels (TGF-β, P > 0.05 vs. control), both MSC-EMV and MSC-exosome treatments significantly enhanced IL-10 and TGF-β production (P < 0.0001 vs. LPS) (Fig. 8A–D). Immunofluorescence analysis demonstrated that LPS stimulation significantly upregulated iNOS expression compared with the control group (P < 0.0005). Both MSC-EMVs and MSC-exosomes treatment effectively reduced LPS-induced iNOS expression (P < 0.0001 vs. LPS group) (Fig. 8E, F). Western blot analysis revealed that LPS treatment significantly increased phosphorylated p65 (p-p65) levels compared with the controls (P < 0.0001), while total p65 (t-p65) expression remained unchanged (P > 0.05). Both MSC-EMV and MSC-exosome treatments markedly attenuated the LPS-induced p-p65 (P < 0.0001 vs. LPS group) without affecting t-p65 expression (Fig. 8G–I). These results collectively demonstrate that MSC-EMVs and MSC-exosomes effectively suppress LPS-induced inflammation through cytokine modulation and NF-κB inhibition, while iNOS expression requires alternative regulatory mechanisms.
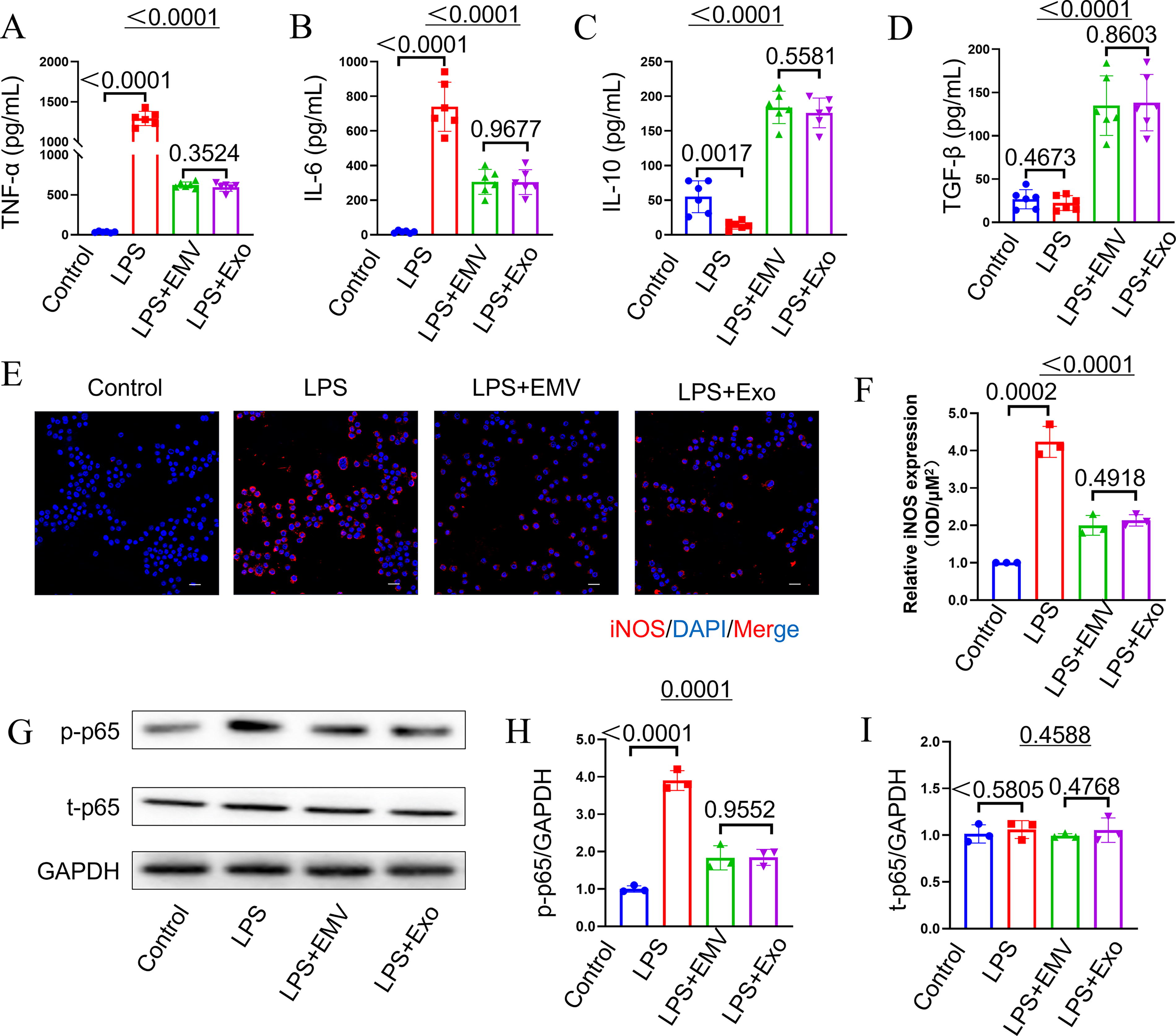
MSC-EMVs and MSC-exosomes attenuate LPS-induced inflammatory responses in THP-1 macrophages. ELISA quantification of
MSC-EMVs and MSC-exosomes attenuate LPS-induced acute lung injury in mice
In the LPS-induced acute kidney injury (AKI) model, treatment with MSC-EMVs and MSC-exosomes significantly ameliorated renal inflammation and tissue damage. Hematoxylin and eosin staining revealed that LPS challenge caused severe tubular epithelial cell necrosis, luminal cast formation, and extensive inflammatory cell infiltration in the renal interstitium, whereas MSC-EMV and MSC-exosome administration markedly reduced these pathological changes (Fig. 9A). These observations were corroborated by semi-quantitative kidney injury scoring, showing significantly lower pathological scores in treatment groups compared with the LPS controls (P < 0.05, Fig. 9B). Western blot analysis demonstrated that LPS stimulation significantly upregulated renal TNF-α and IL-6 protein expression compared with the controls, while both MSC-EMV and MSC-exosome treatments substantially attenuated these increases (P < 0.05 vs. LPS group) (Fig. 9C–E). These results suggest that MSC-EMVs and MSC-exosomes attenuate AKI by mitigating renal inflammation at the protein expression level, mirroring the therapeutic effects of intact MSCs.
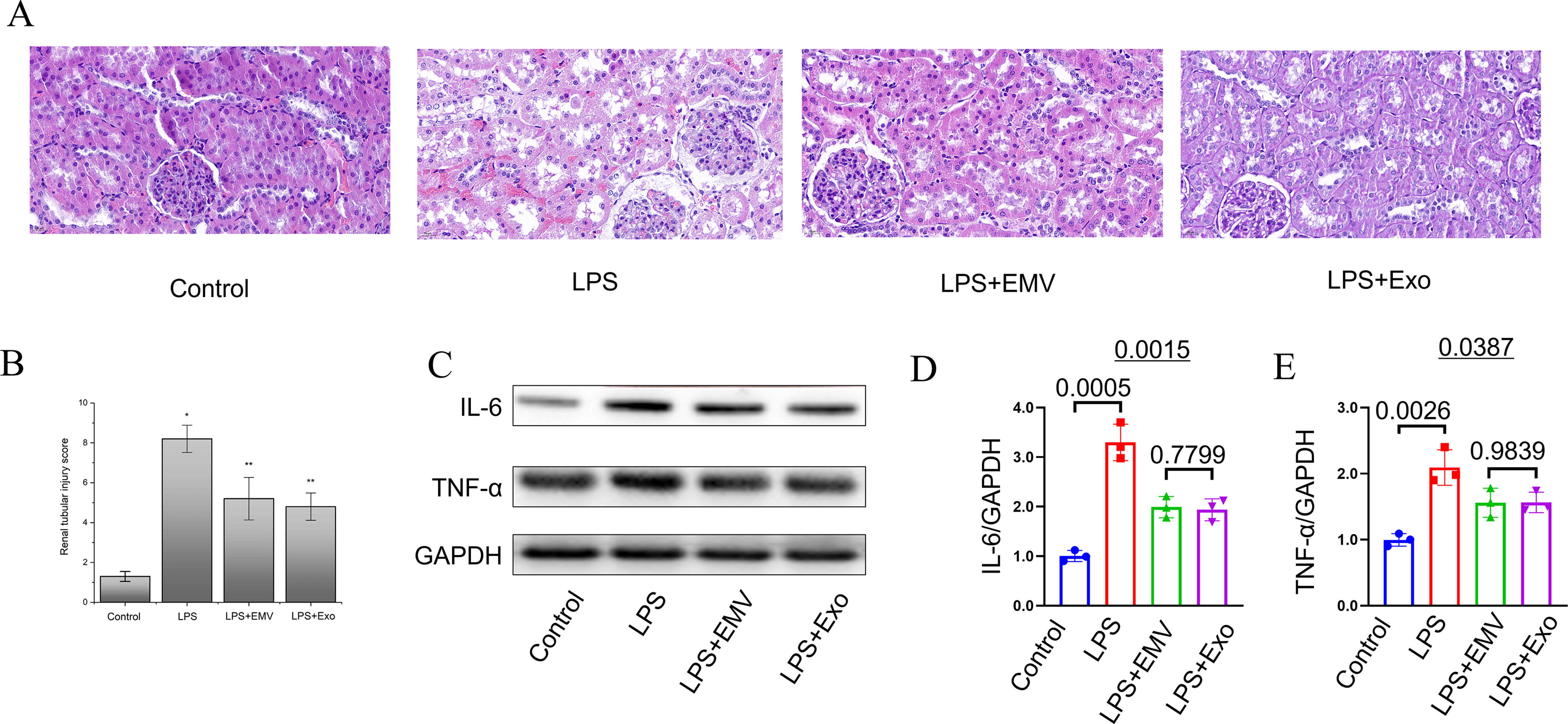
MSC–EMVs and MSC-exosomes attenuate LPS-induced acute kidney injury in mice.
Discussion
Despite the advantages of using MSC-derived exosomes for cell-free therapies in a variety of clinical and animal experiments, their complex production processes and low yield limit their applications. 32 To overcome this limitation, the concept of artificial vesicles has been proposed, which preserve the natural characteristics of cells, have a structure and size similar to those of exosomes, and have a higher production yield. 33 Therefore, EMVs represent a more physiologically relevant option. 34
miRNAs perform various functions, and research has focused on the miRNA expression profiles of exosomes derived from MSCs. 35 Currently, a few studies have conducted miRNAome analysis of EMVs; 36 however, no studies have directly compared the miRNA expression profiles of MSC‐EMVs, MSC-exosomes, and MSCs. A recent study focusing on the proteomics of EMVs and exosomes derived from human umbilical cord MSCs has shown that EMVs and exosomes shared most proteins; however, proteins unique to EMVs and exosomes were also found. 37 It also suggested that most shared proteins would lead to the replacement of exosomes with EMVs in clinical applications. 37 miRNAs also have multiple biological functions; therefore, the miRNAs of EMVs derived from MSCs are also worthy of further exploration.
In this study, we successfully generated EMVs from MSCs, which are similar to MSC-exosomes in structure and size. The proteins expressed by these MSC-EMVs include highly enriched exosomal markers, such as CD63 and TSG101. However, GM130 was identified in MSC-EMVs but not in MSC-exosomes, which may be related to their different production methods. In addition, the production of these EMVs was higher than that of exosomes, which is consistent with the results of a previous study. 37
In the present study, we identified both shared and unique miRNAs by comparing the miRNA expression profiles of MSC‐EMVs, MSC‐exosomes, and MSCs. Our Venn diagrams and heatmap analyses of the top 20 highly expressed miRNAs showed that MSC-EMVs share more miRNAs with parental MSCs than MSC-exosomes. This observation aligns with their distinct biogenesis pathways: EMVs are generated through plasma membrane extrusion, which nonselectively captures cytoplasmic contents, while exosomes form via ESCRT-dependent endosomal sorting that enables more selective miRNA packaging. 38 The functional enrichment analysis showed that these shared miRNAs participate in regulating cell apoptosis, lysosomes, cytokine–cytokine receptor interactions, neuroactive ligand–receptor interactions, the NOD-like receptor signaling pathway, thermogenesis, and other aspects. Notably, exosomal miRNAs are selectively sorted through sequence-specific motifs recognized by RNA-binding proteins such as hnRNPA2B1, whereas EMVs retain a broader miRNA repertoire that reflects their direct derivation from the cytoplasmic pool. These fundamental differences in miRNA composition and sorting mechanisms may underlie their distinct therapeutic potentials in cell-free regenerative medicine.
Among the 20 miRNAs, miR-151a-5p, miR-423-3p, miR-92a-3p, let-7b-5p, miR-26a-5p, let-7a-5p, miR-125a-5p, let-7f-5p, and miR-4485-3p showed higher expression in MSC-EMVs than in the other two groups. Gao et al. found that miR-125a-5p, enriched in extracellular vesicles derived from MSCs can exert a significant cardioprotective effect by delivering its modified oligonucleotides (agomir) to the mouse ischemia/reperfusion myocardium by increasing cardiac function and limiting adverse remodeling. 39 In another study, miR-125a-5p agomir treatment increased the polarization of M2 macrophages, promoted angiogenesis, and weakened the proliferation and activation of fibroblasts, which subsequently helped improve the apoptosis and inflammation of myocardial cells. 40 As a mimic of exosomes, and with enriched miR-125a-5p expression levels higher than that in exosomes, these findings suggest that MSC-EMVs can be used as a targeted miRNA delivery therapy that may be superior to exosomes. Therefore, we speculated that the role of MSC-EMVs could be stronger than that of MSC-exosomes in the relevant directions where these nine miRNAs play regulatory roles. The growing recognition of EMVs and exosomes across biomedical fields—from oncology to immunology and neurology—highlights the need to understand these distinct vesicle populations. While exosomes remain the gold standard for targeted drug delivery due to their refined tropism and lower immunogenicity, EMVs offer distinct advantages, including higher yield and broader cargo diversity, making them ideal for multi-target therapies such as sepsis or acute organ injury. EMVs share key translational frameworks with exosomes, but their scalability could reduce costs for large-scale applications. Challenges include standardizing isolation methods and defining niche applications, but ongoing trials pave the way for EMV clinical adaptation. With optimized engineering, EMVs may soon complement exosomes in treating inflammatory and degenerative diseases.
We compared the miRNA expression profiles between MSC-EMVs and MSC-exosomes; using a Venn diagram, intersections and unique miRNAs were identified between the groups. Furthermore, the analysis of differentially expressed miRNAs between MSC-EMVs and MSC-exosomes revealed 21 upregulated and 17 downregulated miRNAs in the MSC-EMVs group. These 38 miRNAs warrant further exploration to determine their similarities and differences in the functions of MSC-EMVs and MSC-exosomes. For example, the significantly upregulated miR-224-3p has been shown to exert anti-inflammatory effects. A recent study confirmed that miR-224-3p inhibits endothelial cell apoptosis, reduces inflammatory response, and lowers intracellular reactive oxygen species levels by targeting Fos-like antigen 2, leading to anti-atherosclerosis effects and endothelial protection. 41 However, understanding the precise effects of MSC-EMVs compared with those of MSC-exosomes for the prevention and treatment of arteriosclerosis warrants further exploration.
The trend analysis of the miRNA expression levels of the three groups revealed two significant trends, with 144 miRNAs in profile 1 and 108 in profile 2. The miRNAs in profile 1 showed higher expression levels in MSCs than in the other two groups, indicating the richness of genetic material in the parental cells. In contrast, the miRNAs in profile 2 showed relatively high expression in MSCs and MSC-EMVs and relatively low expression in MSC-exosomes. This finding is consistent with the result that the genetic material content in EMVs is higher and they cross more with parental cells, 42 which indicates that MSC-EMVs are superior to MSC-exosomes in transferring genetic material. In the field of functional regulation involving these 108 miRNAs, MSC-EMVs are superior to MSC-exosomes. For example, mir-26a-5p, an miRNA identified in profile 2, has been shown to regulate RNA polymerase III subunit G activity, suggesting that it is a promising therapeutic target for cancer drugs and chemotherapy sensitivity. 43 This reinforces the idea that EMVs, by delivering high-quality miRNAs, might hold promise in cancer therapy. Another study found that mir-26a-5p is a potential therapeutic target in Alzheimer’s disease. 44 It has been found that mir-26a-5p expression is associated with coronavirus disease 2019. 45 In addition, miR-30a-5p carried by extracellular vesicles secreted by the maternal endometrium is considered a major regulatory factor for embryo implantation and development. 46 Elena et al. reported that miR-30a-5p is associated with preeclampsia and miscarriage 47 and suggested further in-depth research to identify useful diagnostic and therapeutic targets for successful embryonic implantation. Rui et al. reported that prenatal exposure to bisphenol S may lead to a decrease in the production of miR-29a-3p in adipocyte-derived exosomes and a decrease in the content of miR-29a-3c delivered to liver cells, further negatively regulating collagen type IV alpha 1 chain expression and promoting liver lipid deposition. 48 These findings provide clues regarding liver metabolic diseases caused by maternal environmental exposure, and as an exogenous miR-29a-3c supplement, EMVs could potentially exert better therapeutic effects than exosomes. Taken together, these findings suggest that MSC-EMVs, with their high yield and rich miRNA content, show promise in therapeutic applications, particularly in cardiovascular diseases, neurodegenerative disorders, cancer, and reproductive health.
The therapeutic potential of MSC-derived extracellular vesicles (EMVs and exosomes) has gained significant attention for their ability to modulate inflammation. Our findings show that both MSC-EMVs and MSC-exosomes effectively attenuate inflammatory responses by suppressing pro-inflammatory cytokines (TNF-α, IL-6) and upregulating anti-inflammatory mediators (IL-10, TGF-β). These vesicles exert immunomodulatory effects through the internalization by macrophages, initiating regulatory cascades, including the inhibition of NF-κB signaling. Our miRNA sequencing data revealed significant enrichment of anti-inflammatory miRNAs in both vesicle types, such as miR-125a-3p, which has been experimentally shown to inactivate the Wnt/β-catenin and NF-κB pathways to reduce the proliferation and inflammation response of RA-FLS via targeting MAST3. 49 This molecular mechanism is consistent with our observed inhibition of NF-κB activity in LPS-stimulated macrophages, suggesting that miR-125a-3p and related miRNAs may mediate these therapeutic effects. In vivo, both vesicle types ameliorated LPS-induced AKI, highlighting their potential as cell-free therapeutics. Despite differences in biogenesis, their similar performance suggests shared immunomodulatory components, reinforcing their therapeutic promise. While MSC-derived vesicles offer advantages over whole-cell therapies, such as reduced immune rejection and easier handling, challenges remain in optimizing dosing, targeting, and large-scale production. Ongoing clinical trials (NCT04313647 and NCT04602104) are addressing these issues, with future research focusing on engineering vesicles to enhance their anti-inflammatory effects and investigating their biodistribution for broader therapeutic applications.
This study has several limitations that warrant discussion. First, while we identified differential miRNA profiles between MSC-EMVs and MSC-exosomes, the functional roles of candidate miRNAs (e.g., miR-125a-5p, miR-224-3p) require experimental validation through gain/loss-of-function studies. Second, our analysis focused on known miRNAs, leaving potentially novel miRNAs unexplored. Third, although we standardized isolation protocols, potential variability in EMV/exosome yield and purity due to technical differences warrants further multi-method validation. Fourth, the single-donor MSC source limits the assessment of inter-individual variability in vesicle miRNA profiles—future multi-donor studies will be essential for clinical translation (all sequencing data are publicly available in GSA-Human: HRA007601 for community validation). Finally, while EMVs show therapeutic promise, their clinical adoption requires further validation of long-term safety and standardized production methods.
Conclusions
To our knowledge, this is the first study to perform a comparative miRNA analysis of EMVs, exosomes, and the originating MSCs. Our research confirmed that EMVs can be stably produced and that they share more miRNAs with parental cells, suggesting their potential as replacements for MSCs or exosomes in novel cell-free therapies. Furthermore, through functional inflammation assays, we preliminarily demonstrated the immunomodulatory and anti-inflammatory roles of both EMVs and exosomes. Comparative analysis also revealed shared and unique miRNA profiles among these vesicles and parental MSCs, enabling trend predictions of their biological functions. While these findings provide mechanistic insights, further studies are needed to fully elucidate the functional significance of specific miRNAs.
Footnotes
Acknowledgments
Figure 1 was created using Figdraw. The authors declare that artificial intelligence was not used in this study.
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
This research was funded by the National Natural Science Foundation of China (No. 82270411).
Availability of Data and Materials
The raw sequence data reported in this paper have been deposited in the Genome Sequence Archive (Genomics, Proteomics & Bioinformatics 2021) in the National Genomics Data Center (Nucleic Acids Res 2022), China National Center for Bioinformation/Beijing Institute of Genomics, Chinese Academy of Sciences (GSA-Human: HRA007601), which are publicly accessible at ![]() .
.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.