
Abstract
Premature ovarian failure (POF) is a significant reproductive disorder characterized by the loss of ovarian function, leading to infertility and endocrine disruption. Hormone replacement therapy (HRT) remains the most commonly used clinical treatment for POF. However, in patients with a history of ovarian or breast cancer, HRT poses significant risks, necessitating the development of alternative approaches. Stem cell-based therapy has emerged as a promising option for treating female infertility disorders such as POF. This study aimed to evaluate the therapeutic effects of ovarian granulosa-like cells (OGLCs) derived from Wharton’s jelly-mesenchymal stem cells (WJ-MSCs) in a POF mouse model. WJ-MSCs were successfully differentiated into OGLCs using combination with a growth factor cocktails, as confirmed by the significant upregulation of granulosa cell-specific markers (P < 0.01). To assess their therapeutic potential, POF was induced in female mice using cyclophosphamide and busulfan, and OGLCs were injected into the ovaries. After 3 weeks, vaginal smear analysis revealed restoration of estrus cycle in OGLC-treated mice. Enzyme-linked immunosorbent assay analysis demonstrated the recovery of serum 17β-estradiol and follicle-stimulating hormone levels (P < 0.05), while histological staining confirmed increased follicular development and restoration of ovarian structure. Furthermore, real-time quantitative polymerase chain reaction analysis showed a significant upregulation of genes related to follicular development and primordial follicle activation, including downstream molecules of the mTOR/PI3K pathway, following OGLCs treatment. These findings suggest that OGLCs possess a strong potential for restoring ovarian function in POF. This study provides evidence supporting the use of OGLCs as a novel cell-based therapeutic approach for female reproductive diseases.
Introduction
The ovary is organized by oocyte and somatic cells such as granulosa cells, theca cells, and ovarian stromal cells. Developing oocytes are influenced by bidirectional signaling with these somatic cells, which coordinate the formation and development of follicles, ovulation, and the corpus luteum. 1 Among ovarian somatic cells, granulosa cells play an important role. For autocrine, paracrine, and endocrine functions, these cells secrete a range of growth factors, sex hormones, and cytokines. Additionally, granulosa cells offer an ideal microenvironment for the oocytes 2 and differentiate into appropriate stages during follicular development. 3 Dysfunctions in somatic supporting cells can lead to various gonadal diseases. 4 For example, granulosa cell abnormalities are associated with disorders such as polycystic ovary syndrome, premature ovarian failure (POF), and granulosa cell tumors may result from granulosa cell malfunction. 5 –7 Among these, POF is one of the most notable conditions linked to granulosa cell dysfunction. Therefore, maintaining the proper function of granulosa cell is essential for ovarian health.
Amenorrhea and infertility are typically the symptoms of POF. The POF is characterized by a high concentration of serum follicle-stimulating hormone (FSH) and a low level of anti-Müllerian hormone (AMH) and estrogen. 8 The etiology of POF is complex, involving factors, including genetic mutations, autoimmune diseases, infections, and endocrine malfunction, all of which are closely related to a loss in ovarian reserve. 9 Hormone replacement therapy (HRT) is still the most used clinical treatment for POF. It helps reduce the risk of complications such as osteoporosis, cardiovascular disease, and urogenital atrophy, while also improving the quality of life for patients. 10 However, HRT is considered unsafe for patients with a history of ovarian or breast cancer, highlighting the need for safer alternatives. 11 Given these limitations of HRT, especially in high-risk patients, stem cell-based therapy has emerged as a promising and potentially safer option to alleviate POF-related symptoms and improve ovarian function.
Currently, cell therapy is regarded as a new option for the treatment of female infertility disorders. 12 In animal models of chemotherapy-induced ovarian dysfunction, several studies have demonstrated the beneficial effect of stem cell-based therapy. 13,14 In addition, the transplantation of differentiated iPSCs into granulosa-like cells recovered ovarian dysfunction mouse model. 15 Therefore, cell therapy for POF is one of the novel therapeutics and promising treatments. Among various stem cell types, mesenchymal stem cells (MSCs) have shown particularly promising results in treating ovarian dysfunction.
MSCs are multipotent adult stem cells derived from the mesodermal lineage. These nonhematopoietic stem cells can be isolated from various adult tissues, including bone marrow and adipose tissue. 16 MSCs possess a great ability for self-renewal and multilineage differentiation into many lineages, including ectodermal, endodermal, and mesodermal lineage. 17,18 They have been utilized in treating various diseases, including damaged endometrium and POF. 19,20 The therapeutic mechanism of MSCs is primarily attributed to their ability to differentiate into target tissue cells, modulate inflammatory and immune responses, and exert anti-apoptotic effects, thereby promoting tissue repair and regeneration. 21 Among MSC sources, Wharton’s jelly-MSCs (WJ-MSCs) are particularly noteworthy due to their minimal nonstem cell contamination, robust stemness characteristics, and high differentiation potential. These properties make WJ-MSCs promising candidates for treating POF as they contribute to ovarian damage repair through granulosa cell growth and upregulation of ovarian stem cell function. 22
In this study, WJ-MSCs were differentiated into ovarian granulosa-like cells (OGLCs), which were then injected into a POF mouse model. It is aimed to investigate the therapeutic effects of OGLCs derived from WJ-MSCs on POF at both the molecular and pathological levels.
Materials and Methods
Chemicals and media
All chemicals were purchased from Sigma (St. Louis, MO, USA) and media from Gibco (Gibco Life Technologies, Gaithersburg, MD, USA), unless otherwise specified.
Ethics approval and animals
All procedures in this study were performed in accordance with relevant guidelines and received approval from the Animal Ethics Committee of Gyeongsang National University (GNU-181210-M0063). For this study, animal experiments were performed using female C57BL6/J mice aged 7–8 weeks old and body weight 18–22 g, which were purchased from Jackson Laboratories (Bar Harbor, USA). Female C57BL6/J mice were maintained under standard laboratory conditions (12 h light-dark cycle, humidity of 30%–40% and 25°C ± 2°C).
Isolation of MSCs derived from Wharton’s jelly’s in umbilical cord
All volunteers for specimen donation were provided informed consent, and the Ethics Committee of the Gyeongsang National University Hospital approved the study (No. GNUH IRB‐2012-09‐004). The human umbilical cord from female was obtained from full-term births, undergoing either cesarean section or normal vaginal delivery. After obtaining the umbilical cord, the tissues were transported to the laboratory in sterilized phosphate-buffered saline (PBS) within 2 h. WJ-MSCs were isolated by previous protocol with minor modification. 23 In brief, the umbilical cord was cut into 2–3 cm lengths, and washed 3 times using Dulbecco’s phosphate-buffered saline (DPBS) containing 1% penicillin-streptomycin (10,000 IU and 10,000 µg/mL, respectively). Following the removal of two arteries and one vein, the slices of 1 mm pieces of Wharton’s jelly were attached to a culture media-coated dish. The culture media contained advanced Dulbecco’s modified Eagle’s medium (ADMEM) with 10% fetal bovine serum (FBS), 100 µg/mL streptomycin, and 100 IU/mL penicillin. After 10–14 days, the attached tissues were removed from the culture dish, and remained cells were cultured in fresh media. Change of the media was performed twice a week. All of the cells were cultured in an incubator at 37°C in a humidified atmosphere containing 5% CO2. Upon reaching 70%–80% confluence, the cells were harvested using 0.25% trypsin-ethylene-diamine-tetra-acetic acid (Trypsin-EDTA) solution with centrifugation of 300g for 5 min. The cells were cultured to passage 3 for each experiment.
Flow cytometry
WJ-MSCs were evaluated for the expression of surface markers by BD FACS flow cytometer (BD FACS Calibur; Becton Dickinson, Franklin Lake, NJ, US). Briefly, MSCs (1 × 105) were fixed in a 4% paraformaldehyde solution for 1 h at room temperature. The fixed cells were washed by DPBS 2–3 times and reacted with FITC-conjugated mouse antihuman CD34 (1:200; BD Biosciences, NJ, USA), FITC mouse antihuman CD45 (1:200; BD Biosciences), FITC mouse antihuman CD105 (1:200; BD Biosciences), FITC rat antihuman CD44 (1:200; BD Biosciences) and APC-conjugated mouse antihuman CD73 (1:200; BD Biosciences), APC mouse antihuman CD105 (1:200; BD Biosciences) for 1 h at room temperature. After the reaction, the cells were washed by DPBS 2–3 times to remove unbounded antibodies. The standard was established by FITC mouse IgG1 (1:200; BD Biosciences) and APC mouse IgG1 (1:200; BD Biosciences). Data were analyzed by FlowJo v10 software.
Mesenchymal lineage differentiation
WJ-MSCs were differentiated into mesenchymal lineages following the previous study. 23 Briefly, adipogenic differentiation was induced for 3 weeks with DMEM containing 10% FBS, 100 µM indomethacin, 10 µM insulin, and 1 µM dexamethasone. The adipogenesis was confirmed by observing the accumulation of lipid droplets after staining with Oil Red O staining. Osteogenesis was induced for 3 weeks with DMEM supplemented with 10% FBS, 200 µM ascorbic acid, 10 mM β-glycerophosphate, and 0.1 µM dexamethasone. The mineralization and calcium deposition were confirmed by Alizarin Red S staining. Differentiation into chondrocytes was achieved using a modified pellet culture method. For pellet culture, WJ-MSCs (1 × 106 cells) were suspended in 1 mL and formed in 15 mL centrifuge tubes. The MSCs were cultured for 3 weeks in STEMPRO chondrogenesis media (STEMPRO chondrogenesis differentiation kit, Invitrogen, CA, USA). The differentiation into chondrocyte lineage was evaluated by the accumulation of proteoglycans using 1% Alcian blue solution staining.
Differentiation into OGLCs
With a minor modification, WJ-MSCs were differentiated into OGLCs. 2 Briefly, WJ-MSCs at 3 passages were seeded to 1 × 104/cm2 density, then cultured with ADMEM culture media containing 10% FBS for 24 h. To differentiate, WJ-MSC were induced to OGLCs using Dulbecco’s Modified Eagle Medium/Nutrient Mixture F12 (1:1) (DMEM/F12) media supplemented with 10% FBS, 1% nonessential amino acids, 0.1 mM beta-mercaptoethanol, 25 ng/mL recombinant human follistatin (Peprotech, NJ, USA), 5 ng/mL recombinant human FGF-basic (Peprotech, NJ, USA), and 10 ng/mL recombinant human BMP-4 for 7 days. The media was changed every 2 days intervals.
Real-time quantitative polymerase chain reaction
After sample collection, the samples were rapidly frozen in liquid nitrogen and stored at −80°C until RNA extraction. Total RNA was extracted from WJ-MSCs, OGLCs, ovarian tissue, and uterine tissue using RNeasy Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. After quantifying total RNA using a Nanodrop 1000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA), complementary DNA (cDNA) was synthesized with 500 ng of RNA sample using Omniscript Reverse Transcription Kit (Qiagen, Hilden, Germany). Real-time quantitative polymerase chain reaction (RT-qPCR) was reacted with Rotor-Gene (Qiagen, Hilden, Germany), and the reaction mix contained Quantinova SYBR® Green PCR Master Mix (Qiagen, Hilden, Germany), 50 ng of cDNA, and 0.7 μM forward and reverse primers. Real-time quantitative polymerase chain reaction (RT-qPCR) was run in a Rotor-Gene Q qRT-PCR machine (Qiagen) following predenaturation, denaturation, and combined annealing/extension step. Predenaturation at 95°C for 2 min; 40 PCR cycles at 95°C for 10 s, 60°C for 6 s. The melting curve shows that temperature alters from 60°C to 95°C by 1°C/s and cooling at 40°C for 30 s. The specificity of RT-qPCR amplification was confirmed by melt curve analysis, ensuring the presence of a single peak melting curve. Nontemplate controls (NTCs) were included to verify the nonspecific amplification. Rotor-Gene Q series software (Qiagen, Hilden, Germany) was used for the analysis of cycle threshold values (Ct values) values for each sample. In addition, the relative levels of gene expression were calculated by using the 2−ΔΔCT method. The primers used in this study are listed in Table 1.
Lists of Primers Used in RT-qPCR Analysis
Immunofluorescent staining
The cells were fixed with 4% paraformaldehyde for 15 min and permeabilized in 0.1% Triton-X for 10 min at room temperature. After blocking with 3% BSA solution for 1 h, the cells were incubated with primary antibodies such as rabbit anti-aromatase (1:100; Invitrogen, MA, USA), goat anti-FOXL2 (1:200; Invitrogen, MA, USA), and rabbit anti-FSHR (1:200; LSBio, WA, USA) for overnight at 4°C. They were then incubated with FITC-conjugated mouse anti-rabbit IgG (1:200; Santa Cruz, TX, USA) or FITC-conjugated donkey anti-goat IgG (1:200; Santa Cruz, TX, USA) for 1 h at room temperature. After being washed 2–3 times with DPBS, nuclei of cells were counterstained with 1 µg/ml 4′, 6-diamidino‐2-phenylindole (DAPI) for 5 min. A fluorescence microscope (Leica, Wetzlar, Germany) was used to capture the images of the cells.
Preparation of POF mice model and transplantation of OGLCs into ovaries
To induce POF, the mice were treated with alkylating agents in minor modification. 24 In brief, the mice were randomly classified into the following three groups: (1) Normal control (Normal; n = 8), which were healthy mice with no treatment. (2) PBS-only injected POF mice (POF; n = 8), which were induced POF using cyclophosphamide and busulfan. (3) OGLCs-injected POF mice (OGLC-POF; n = 10), which were injected with OGLCs into ovaries. The initial sample size was determined based on experimental feasibility and ethical considerations. A slightly larger number of mice were included in the POF-OGLC group (n = 10) to ensure sufficient statistical power for evaluating the effects of OGLC treatment. The POF and Normal groups (n = 8 each) were designed to provide an appropriate comparison while minimizing the use of animals in accordance with ethical guidelines. Animals were randomly assigned to groups based on body weight, ensuring even distribution and minimizing bias. To induce POF, recipients were injected intraperitoneally with cyclophosphamide 120 mg/kg in normal saline and busulfan 12 mg/kg in Dimethyl Sulfoxide (DMSO). After 2 weeks from POF induction, the mice were anesthetized with Avertin (0.75 mg/g), and injected with only 10 μl PBS in the POF group or with 2 × 106 OGLCs in 10 μl PBS in the OGLC-POF group into both ovaries. To evaluate the recovery of ovarian function from OGLCs injection, the final experiment was conducted 3 weeks postinjection.
Vaginal smear of the POF mice model
With minor modification, estrus cycle of the mice was observed by daily vaginal smear at 9–10 am from 1 week after cell injection. 25 The vaginal smear was conducted for 12 days as the estrus cycle of mice lasts approximately 4–5 days. To ensure reliable cyclic patterns analysis, the vaginal smear was conducted for 12 days allowing for observation of at least three complete cycles. The mice were then sacrificed by cervical dislocation 3 weeks after cell injection to collect the uterus and ovaries.
Enzyme-linked immunosorbent assay
Three weeks after injection, blood samples were collected from the mice via direct cardiac puncture using a 26-gauge syringe. The levels of 17β-estradiol and FSH were measured using an enzyme immunoassay kit. Briefly, the blood samples were incubated at room temperature for 20 min. After allowing clot formation, blood samples were centrifuged at 2200g for 10 min. The serum was stored at −80°C. 17β-estradiol (Abcam, Cambridge, UK) and FSH (MyBioSource, CA, USA) levels in serum were analyzed with an enzyme immunoassay kit according to the manufacturer’s protocol, respectively.
Clinical and histological assessment of the POF mice model
After being measured the weight of the uterus and ovary, the middle of uterine horns on both sides were collected for the evaluation of uterus thickness and gene expression. The ovaries from each mouse were classified respectively for isolation of mRNA and histological assessment. To estimate histological assessment, the parts of the uterus and ovary were fixed with 4% paraformaldehyde for overnight to evaluate the formation of follicles and uterus thickness, respectively, and followed by rinsed with DPBS 3 times for 5 min and dehydrated overnight with 30% sucrose. The samples were processed optical cutting temperature compounds (Tissue-Tek®, USA) embedding, cut into 5 μm sections, and mounted on glass slides. The slide was stained with counterstaining for cell tracking with 1 μg/ml DAPI for 5 min. The slides were stained with hematoxylin and eosin (H&E) to examine pathological morphology. Specifically, hematoxylin was applied for 10 min, followed by differentiation in 1% hydrochloric acid in 70% ethanol. Subsequently, eosin Y staining was performed for 20 s. After staining, all sections underwent dehydration, clearing, and mounting.
Tracking of OGLCs in the POF mice ovary
Human-specific cells in ovarian tissues of POF mice were observed by immunohistochemistry using the immunohistochemical kit (Abcam, Cambridge, UK) containing rabbit-specific horse radish peroxidase-diaminobenzidine (HRP-DAB). In brief, tissue sections were performed endogenous peroxidase blocking with 3% hydrogen peroxide solution for 10 min at room temperature. Then, the nonspecific protein was blocked with 3% goat serum solution for 30 min. After being washed with DPBS 3 times, the sections were reacted with primary antibody such as rabbit antihuman Ku80 (1:500; Cell Signaling, MA, USA) at 4°C overnight. Subsequently, the sections were incubated with secondary antibody such as biotinylated goat anti-rabbit IgG (H + L) for 10 min. To develop positive signals, the sections reacted with 0.05% diaminobenzidine tetrahydrochloride (DAB, Sigma-Aldrich). All slides were mounted, stained with hematoxylin as a counterstain, and examined under a microscope.
Statistical analysis
Data analysis was performed using GraphPad Prism version 8. Independent experiments were conducted at least 3 times for each set of data, with data representing the mean ± standard error of the mean (SEM) of all repeats. The statistical differences among experimental groups were estimated by Student’s t-test for WJ-MSCs differentiation into OGLCs and one-way analysis of variance (ANOVA) with Newman–Keuls post hoc for mice experiments in this study. Results were considered statistically significant at P < 0.05.
Results
Isolation and characterization of MSCs derived from Wharton’s jelly
WJ-MSCs were successfully isolated and cultured from umbilical cords of female donors. To confirm MSC-specific characteristics, WJ-MSCs were analyzed for surface markers. They were positive for MSC-specific markers, including CD44, CD73, CD90, and CD105, and negative for hematopoietic markers, such as CD34 and CD45 (Fig. 1A). The differentiation potential of WJ-MSCs into mesenchymal lineages was assessed by specific cytochemical staining. WJ-MSCs successfully differentiated into adipocytes, osteocytes, and chondrocytes over a period of 21 days. Adipogenic differentiation was verified by Oil Red O staining, revealing the accumulation of intracellular lipid droplets. Osteogenic differentiation was validated by Alizarin Red S staining, which indicates the formation of mineralized nodules. Chondrogenic differentiation was confirmed by Alcian blue staining, showing the accumulation of proteoglycans (Fig. 1B).
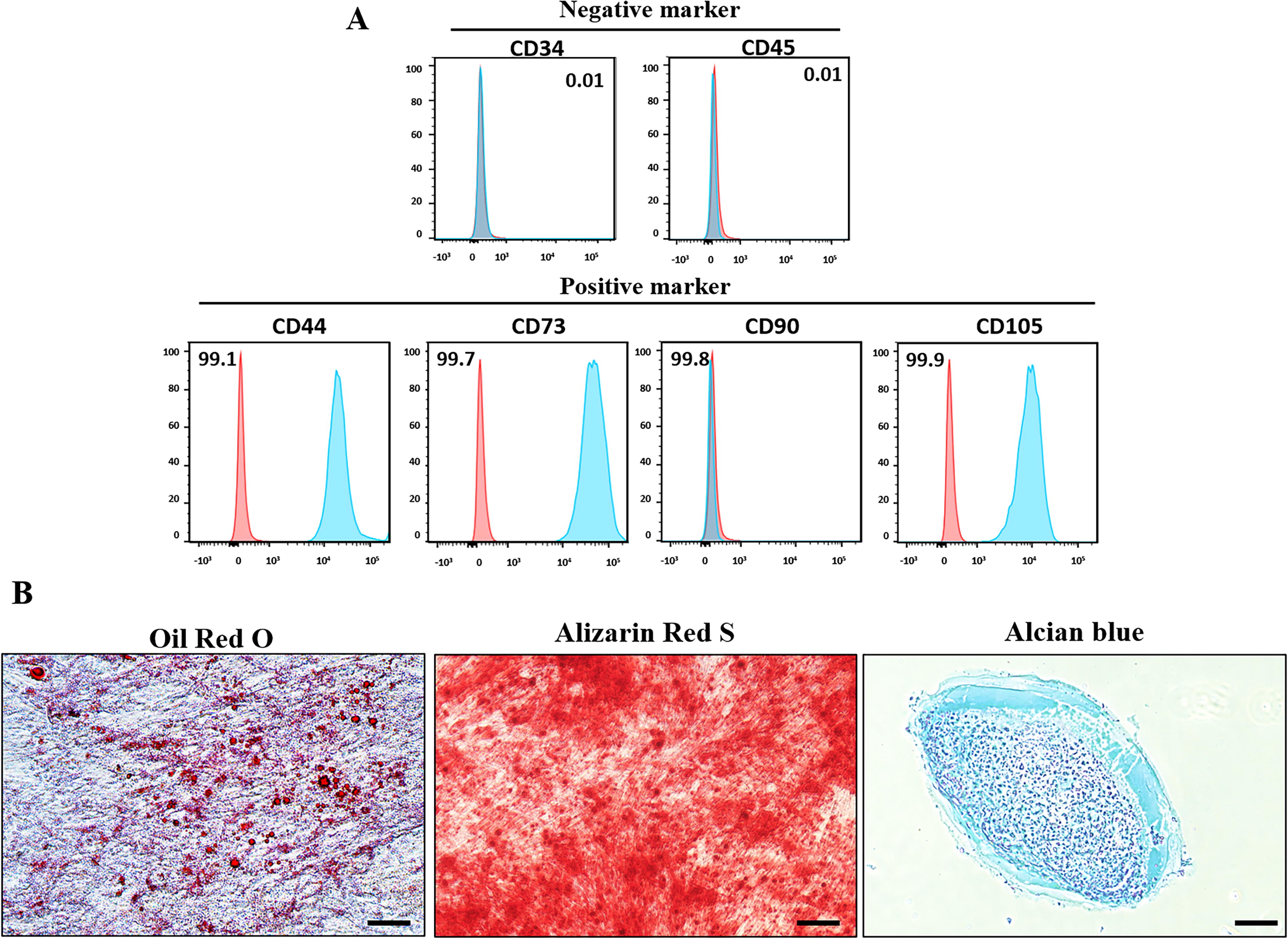
Characterization of WJ-MSCs.
Differentiation potential of WJ-MSCs into OGLCs
The differentiation potential of WJ-MSCs into OGLCs was assessed following a previously established protocol with minor modification. 2 After 7 days of induction, WJ-MSCs exhibited slight morphological changes indicative of granulosa cells (Fig. 2A). Differentiated cells showed a significant upregulation of granulosa cell-specific markers, including follicle-stimulating hormone receptor (FSHR, P < 0.001), forkhead box protein L2 (FOXL2, P < 0.01), aromatase (CYP19A1, P < 0.01), and anti-Müllerian hormone receptor type 2 (AMHR2, P < 0.001) compared to undifferentiated MSCs (Fig. 2B). Furthermore, immunostaining confirmed the expression of granulosa cell-specific proteins such as FSHR, CYP19A1, and FOXL2 in differentiated cells (Fig. 2C).
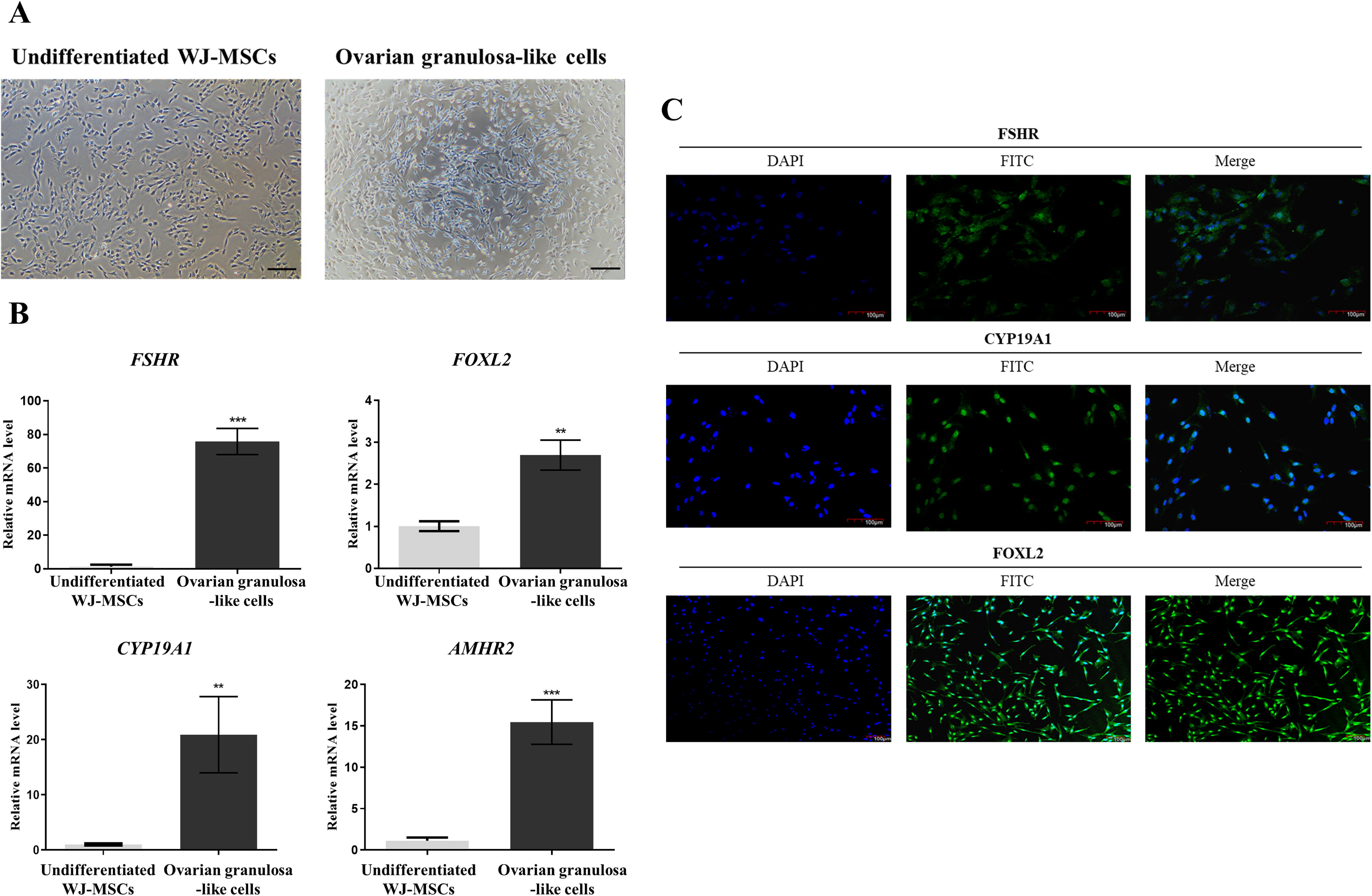
Differentiation into OGLCs derived from WJ-MSCs.
Restoration of hormonal balance by OGLCs in POF mice
After inducing POF, OGLCs were injected into both ovaries of the mice. Then, vaginal smear was performed to estimate the estrus cycle for 12 days after 1 week of cell injection (Fig. 3). When compared to Normal mice, POF mice exhibited prolonged diestrus with no progression through the estrous cycle stages. Whereas, OGLC-POF mice showed an improved estrus cycle, displaying all stages of estrus cycle as proestrus, estrus, metestrus, and diestrus (Fig. 3A). The duration of the estrus cycle in OGLC mice was prolonged 1.5 times compared with Normal estrus cycle (Fig. 3B). To evaluate the therapeutic effect on sex hormone, serum levels of 17β-estradiol and FSH were measured using enzyme-linked immunosorbent assay assay. POF mice had significantly lower levels of 17β-estradiol compared to Normal mice (P < 0.01), whereas OGLC-POF mice showed a significant increase (P < 0.01) in 17β-estradiol levels, which were comparable to those of Normal mice. Conversely, FSH level in POF mice was significantly (P < 0.05) higher than in Normal mice. However, the FSH level in OGLC-POF mice was significantly (P < 0.01) decreased, which were similar to Normal mice (Fig. 3C).
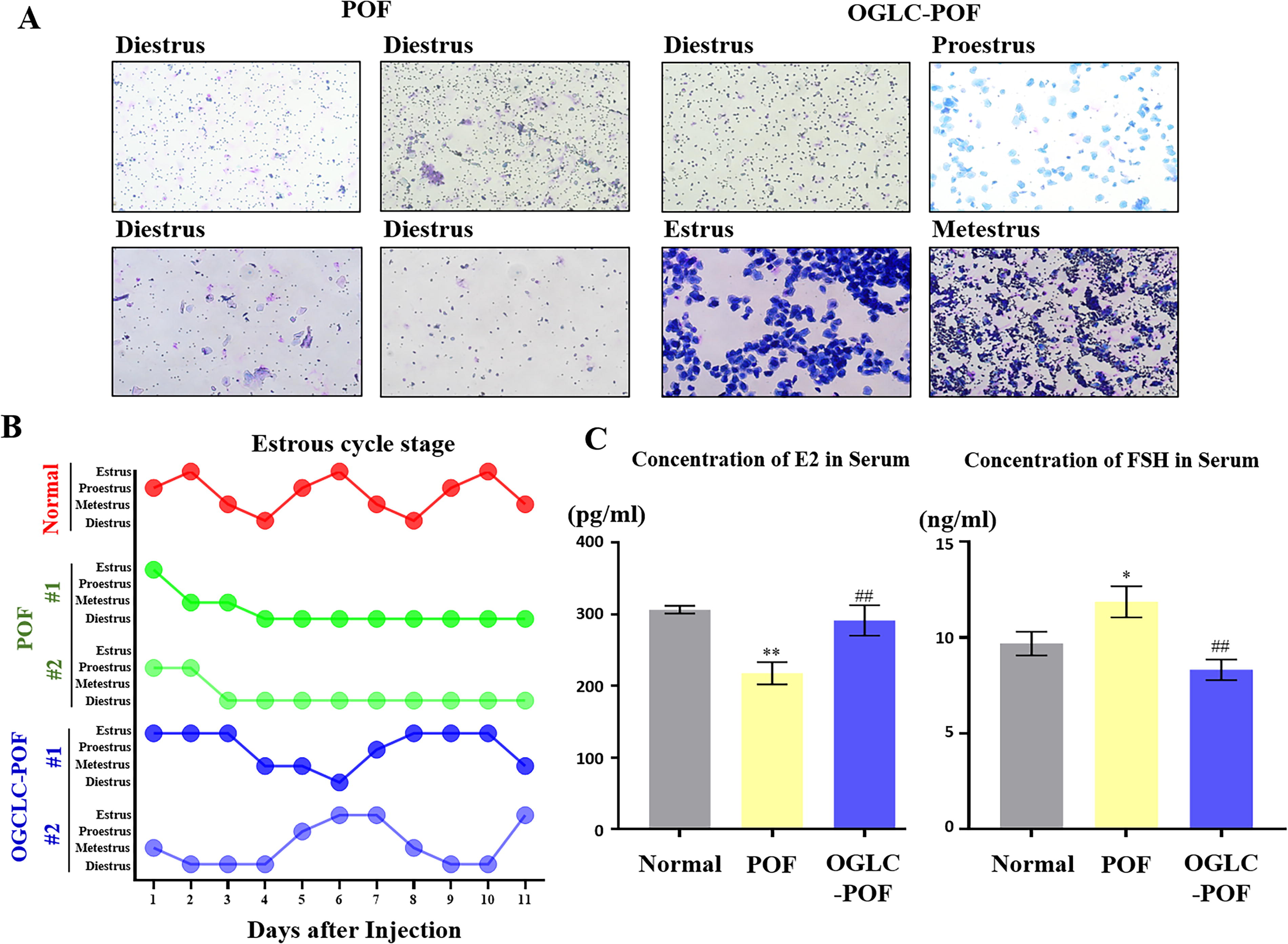
Restoration of sex hormone balance by OGLCs.
Therapeutic effect of OGLCs in POF mice reproductive organ
Three weeks postinjection, all mice were sacrificed. The uterus and ovaries were observed macroscopically morphology. In POF mice, the uterus and ovaries appeared pale and atrophic, along with a significantly (P < 0.001) lower weight compared to Normal mice. In contrast, OGLC-POF mice displayed uterine and ovarian morphology and coloration similar to those of Normal mice (Fig. 4A). The uterine weight in OGLC-POF mice was comparable to that of Normal mice whereas significantly higher compared with POF mice (P < 0.001). Uterine thickness in Normal mice was significantly increased in POF mice (P < 0.05). Moreover, OGLC-POF mice significantly increased uterine thickness compared with the POF mice (P < 0.05) (Fig. 4B).
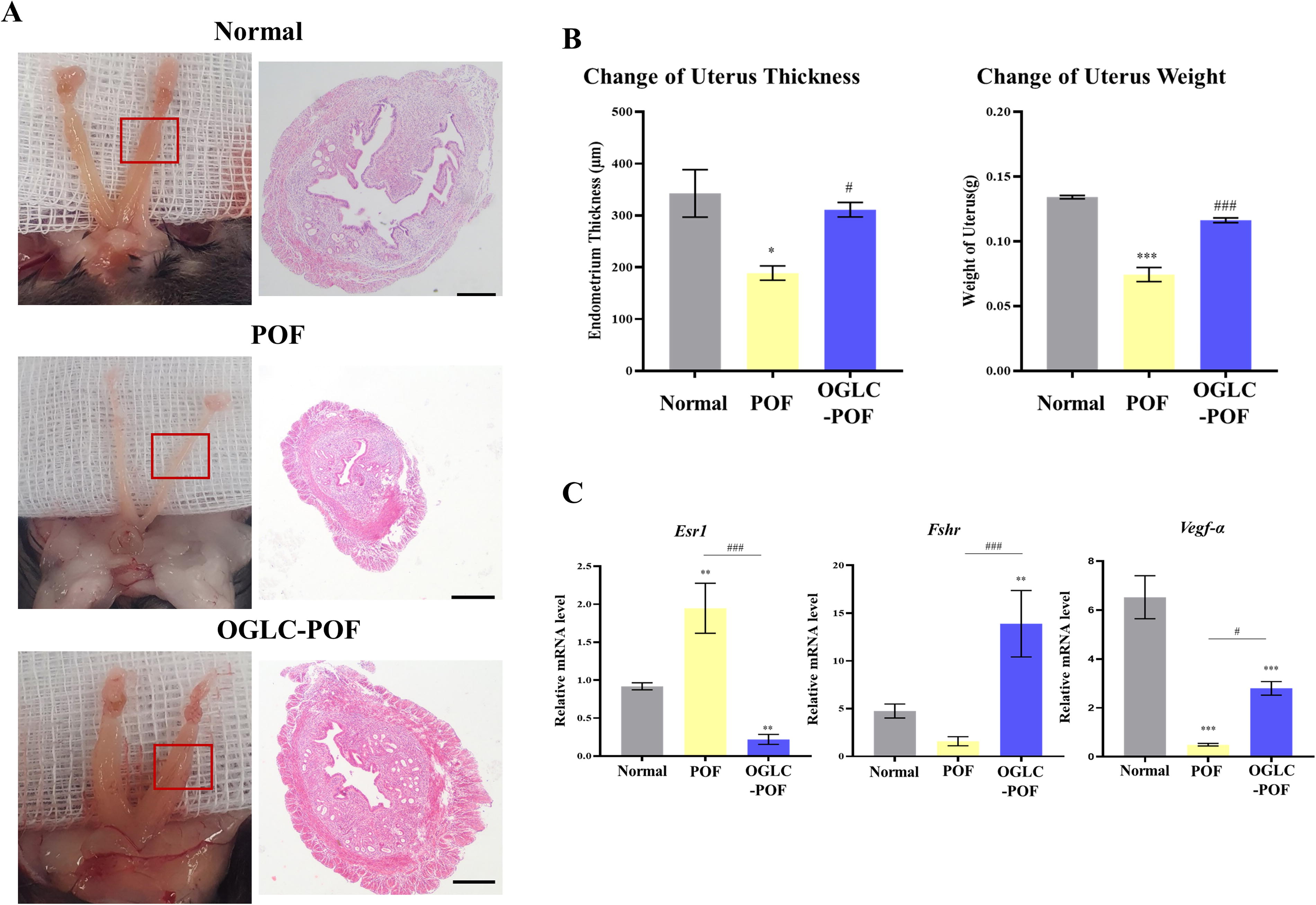
Therapeutic effect of OGLC on uterus of POF mice.
Given expression analysis of hormone receptor and angiogenesis marker in uterus was performed by qRT-PCR. The expression of Esr1 was significantly increased (P < 0.01) in POF mice compared to Normal mice, but significantly decreased in OGLC-POF mice (P < 0.01). The mRNA expression level of Fshr in POF was lowest and showed no significant difference in Normal mice (P = 0.46). However, in OGLC-treated POF mice, Fshr expression was significantly elevated compared to both Normal (P < 0.01) and POF mice (P < 0.001). Regarding angiogenesis, Vegf-α expression was significantly higher in Normal mice compared to both POF and OGLC-POF groups (P < 0.001). Nonetheless, Vegf-α expression in OGLC-POF mice was significantly increased compared to POF mice (P < 0.05) (Fig. 4C).
In Normal group ovary, various stage follicles and healthy oocytes were clearly observed. However, in the POF group, healthy follicles were rarely detected in the ovary. Histological analysis of POF mice ovaries observed disrupted folliculogenesis, along with extensive destruction of ovarian stromal tissue and germ cells, and severe follicular atresia. In contrast, ovarian histological assessment in OGLC-POF mice clearly showed a marked reduction in atretic follicles and the presence of healthy follicles (Fig. 5A). Additionally, there was a notable improvement in the destruction of ovarian stromal tissue and germ cells. To estimate the therapeutic effect on folliculogenesis, the number of follicles was counted on H&E staining. In OGLC-POF mice, the number of follicles at different stages—primordial, primary, secondary, and antral—was significantly increased compared to the POF group (Fig. 5B). Furthermore, the injected OGLCs were traced using the human-specific antibody Ku80, and their localization was identified near oocytes within the follicles (Fig. 5C).
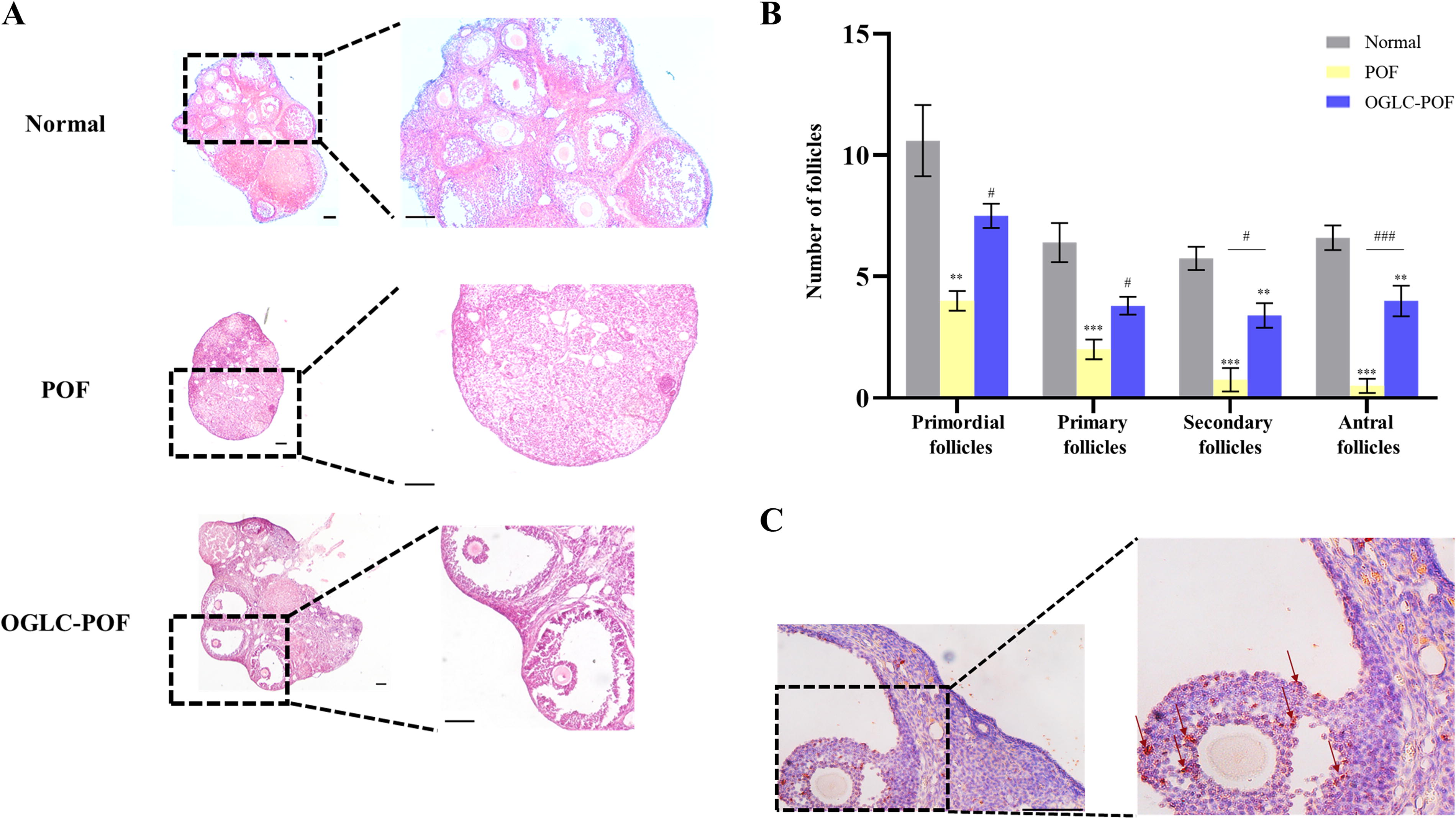
Therapeutic effect of OGLC on ovaries of POF mice.
Assessment of follicular development-related gene expression in the ovary
The follicular development relative genes such as Vegf-α and Igf-1 were estimated using qRT-PCR in experimental mice ovaries. The expression of Vegf-α in OGLC-POF mice was significantly higher compared to both Normal (P < 0.01) and POF mice (P < 0.01). The gene level of Igf-1 in POF revealed significant decrease (P < 0.05) compared with Normal. However, the expression of Igf-1 in OGLC-POF was significantly increased compared with POF mice (P < 0.05) with no significant difference observed between Normal mice (P = 0.96). The expression of Fshr in POF was significantly decreased compared with Normal mice (P < 0.001). In OGLC-POF mice, Fshr expression was significantly upregulated compared to POF mice (P < 0.001), but no significant difference was observed compared with Normal mice (P = 0.13) (Fig. 6A).
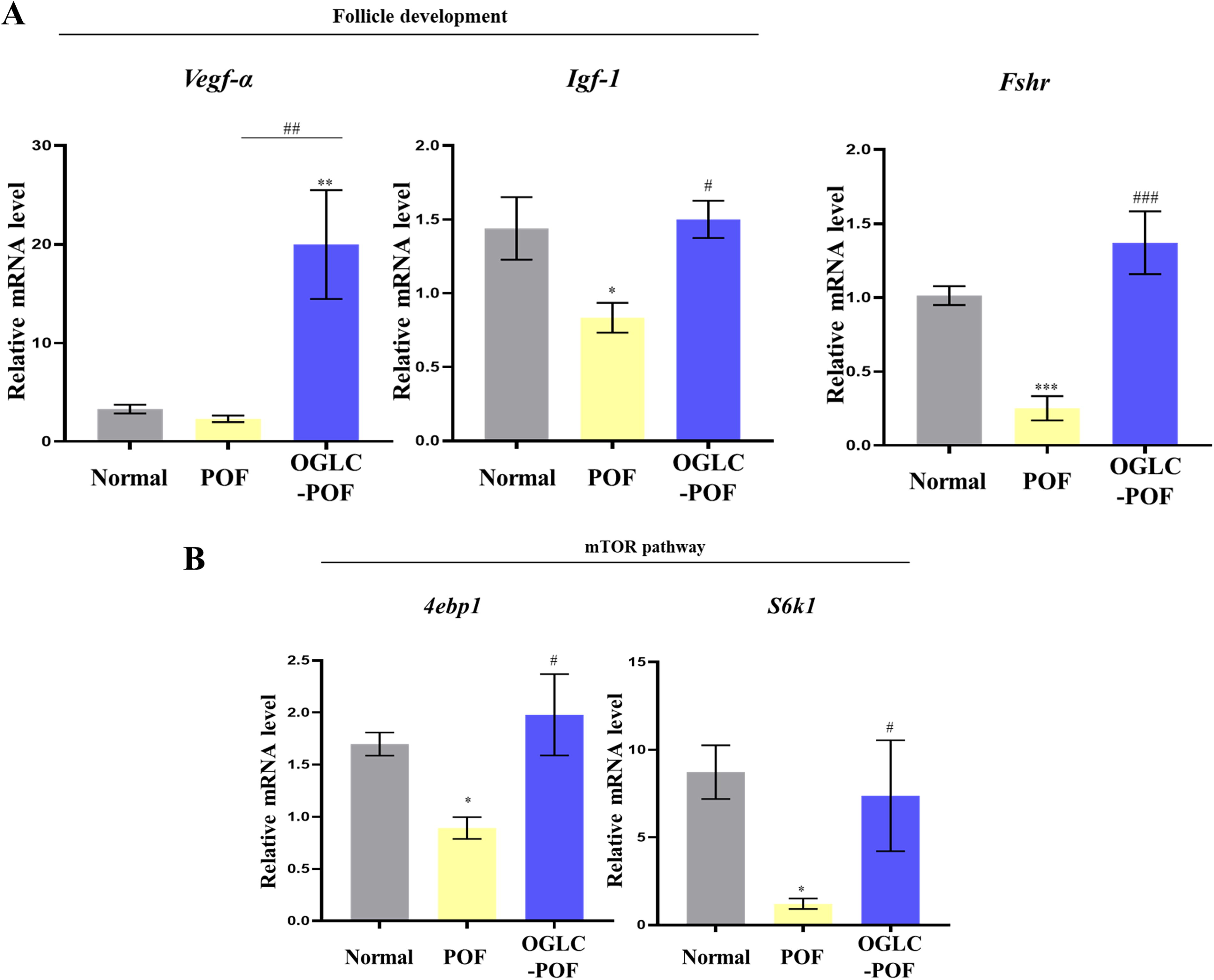
Gene expression in POF mice ovary following OGLC injection. The ovaries collected from the mice were analyzed for gene expression using RT-qPCR.
Additionally, the activation of primordial follicle relative genes, 4ebp1 and S6k1 genes, which are downstream molecules of the mTOR/PI3K pathway, were examined with qRT-PCR in mice ovary. When compared with Normal mice, mRNA expressions of downstream molecules of the mTOR/PI3K in POF group were significantly decreased (P < 0.05). However, in OGLC-POF mice, the expression of 4ebp1 and S6k1 was significantly increased compared to POF mice (P < 0.05), while remaining comparable to Normal mice (Fig. 6B).
Discussion
Granulosa cells within developing ovarian follicles are primarily responsible for secreting endogenous estrogen and play a crucial role in female reproductive health, particularly ovarian reserve. The loss or dysfunction of ovarian reserve is strongly associated with onset of POF. 9,26 One of the critical factors contributing to the depletion of ovarian reserve is apoptosis of granulosa cells, as the quantity and viability of ovarian granulosa cells are directly linked to POF. 15,27 Stem cell-based therapies for POF have therefore focused on modulating the ovarian microenvironment, supporting follicular development, and enhancing granulosa cell function. Previous studies have demonstrated that MSCs treatment on POF can maintain follicles at various developmental stages of development and support granulosa cell viability. 28 In addition, MSCs injected into POF mouse ovaries have shown the potential to restore normal follicle numbers and differentiate into granulosa cells. 9 Given the potential of stem cell-derived granulosa cells to restore ovarian function, this study aimed to evaluate the therapeutic effects of OGLCs derived from WJ-MSCs in a POF mouse model.
Human WJ-MSCs, isolated from full-term umbilical cords, demonstrated in vitro culture adaptability and the potential to differentiate into mesenchymal lineages. This study confirmed that WJ-MSCs exhibit distinct characteristics, including the expression of specific cell surface markers and the ability to differentiate into mesenchymal lineages under defined culture conditions. These findings align with previous research. 23 Furthermore, WJ-MSCs successfully differentiated into mature granulosa cells, expressing granulosa cell-specific markers such as FSHR, CYP19A1, FOXL2, and AMHR2, and exhibited granulosa cell functionality. It is demonstrated that granulosa cells have typical spindle-shaped morphology. 29 Granulosa-like cells showed only subtle morphological changes between undifferentiated and differentiated cells. The differentiated cells expressed both granulosa cell-specific genes and proteins, consistent with earlier studies. 2,30 Taken together, WJ-MSCs, in combination with a growth factor cocktail, were effectively differentiated into functional granulosa cells.
POF mouse models are widely utilized in reproductive research to study female infertility. Cyclophosphamide and busulfan, which exhibit relatively low systemic toxicity, 31 are commonly used to induce POF by causing ovarian damage. 32 In this study, POF mice induced by cyclophosphamide and busulfan exhibit elevated serum FSH levels and reduced estradiol concentrations compared to normal mice. Additionally, these mice display irregular estrous cycles, characterized by prolonged periods of diestrus. This observation aligns with previous findings in POF mouse models. 33 In the present study, the injection of OGLCs into the ovaries of POF mice restored serum levels of 17β-estradiol and FSH to near-normal levels. While the estrous cycle in OGLC-POF mice remained approximately 1.5 times longer than in Normal mice, these mice exhibited all phases of the estrous cycle, including proestrus, estrus, metestrus, and diestrus. These findings are consistent with previous studies in which stem cell-derived granulosa cells were shown to improve ovarian dysfunction and restore normal ovarian function. 15
Uterine hypotrophy is a common characteristic of endocrine dysfunction, as observed in mice. 34 Similar to ovariectomized models, POF mice exhibit hypotrophy in reproductive organs, including the ovaries and uterus. 35 In this study, the reproductive organs of OGLC-POF mice appeared healthier, with notable improvements in size and weight, suggesting the restoration of endocrine function can reverse atrophy in these organs. 36 At the molecular level, healthy uterine tissue typically exhibited low mRNA expression of Esr1 and high expression of Fshr, but these patterns were inverted in POF mice. Following OGLC injection into the ovaries of POF mice, the expression levels of Esr1 and Fshr in the uterine tissue were restored to those of healthy controls, indicating that OGLCs may modulate the microenvironment of reproductive organs through hormone receptor expression. Similarly, the expression of the angiogenesis-related gene Vegf-α in the uterus of OGLC-POF mice mirrored the patterns observed in sex hormone-related genes such as Esr1 and Fshr. These findings are consistent with previous studies showing that vascular endothelial growth factor (VEGF) expression in the uterine endometrium of premature ovarian insufficiency models increased following MSC treatment. 37
The POF mice injected OGLCs showed a reduction in atretic follicles and an increase in developing follicles compared to POF mice. This suggested that OGLCs injection into both ovaries may be capable of restoring folliculogenesis in POF mice. In addition to their presence within follicles and ovarian tissue near the oocytes, OGLCs contributed to ovarian regeneration by acting similarly to native granulosa cells in ovarian follicles. A previous study showed that presumptive granulosa cells from foxl2 transfected Embryonic stem cells (ESCs) injected into mice ovaries were found only in the granulosa cell layer of growing follicles, and participated in folliculogenesis in vivo. 38 Although the results are not exactly matched with the present study, differentiated granulosa cells had an effect on folliculogenesis in the ovary.
FSH exerts its essential role in promoting granulosa cell growth through its interaction with FSHR. POF is closely associated with impaired follicle formation, primarily due to reduced FSHR sensitivity. 39 Follicular development is further supported by ovarian angiogenesis, which provides essential nutrients, hormones, and oxygen. VEGF is a well-established angiogenic factor critical for this process. 40 Similarly, IGF-1 is necessary for granulosa cell growth during the early stages of folliculogenesis. 41 Intraovarian factors such as VEGF, BMPs, and IGF-1 play a pivotal role in regulating follicular development, 42,43 while the PI3K/mTOR signaling pathway is instrumental in the activation and maintenance of primordial follicles. 44,45 In this study, the injection of OGLCs into the ovaries of POF mice significantly restored Fshr expression and increased the mRNA expression levels of Vegf-α and Igf-1 in ovarian tissue. These findings suggest that intraovarian injection of OGLCs has a therapeutic effect on key factors for follicular development, including Vegf-α, Igf-1, and Fshr. Additionally, OGLCs injection significantly enhanced the expression of mTOR/PI3K pathway-related genes, such as 4ebp1 and S6k1, in the ovaries of POF mice. These results align with previous studies demonstrating similar therapeutic effects in models of ovarian aging or POF. 46 –48 In summary, the injection of OGLCs into both ovaries markedly improved the condition of reproductive organs and ovarian function by enhancing follicular development and supporting sex hormone secretion.
Conclusion
POF mice induced by alkylating agents, including cyclophosphamide and busulfan exhibited ovarian dysfunction, characterized by irregular estrous cycles, imbalanced sex steroid hormone levels, and atrophied reproductive organs. The injection of OGLCs demonstrated therapeutic potential in alleviating these conditions. These findings suggest that OGLCs differentiated from WJ-MSCs could serve as a promising alternative therapy for POF.
Footnotes
Acknowledgments
The authors are highly thankful to Prof. B.W. Park, Dr. Ji-Eun Park, and Dr. Ji-Kwon Park for providing valuable human umbilical cord samples.
Author Disclosure Statement
No potential conflicts of interest were reported by the authors.
Funding Information
This study was supported by a grant from the National Research Foundation (NRF) of Korea, funded by the government of the Republic of Korea (grant no. NRF-2021R1A2C1007054), and Stem Centric Co. Ltd., Republic of Korea.