
Abstract
Jumping locomotion is critical for microrobots to overcome obstacles. Among the microjumping robots, the development of an omnidirectional jumping mechanism is challenging. To avoid the complicated microfabrication process, we present an insect-computer hybrid robot by controlling the locomotions of an Oriental Migratory Locust (Locusta migratoria manilensis, Meyen 1835). The insect-computer hybrid robot achieves repetitive omnidirectional jumps of ∼100 mm high. A series of experiments on jumping control, turning control, and collaborative directional jumping control are carried out. We also demonstrate the implementation of a wireless stimulator backpack that provides remote locomotion control, which transforms the insect into a hybrid robot. Moreover, a feedback jump control system is subsequently presented. The results indicate that the hybrid robot could easily achieve an omnidirectional jump and maintain body righting after landing. This robot is well-suited for applications that require locomotion on uneven terrains, such as environmental surveillance and search and rescue.
Introduction
The animal structures have attracted tremendous research interest due to the soft materials in their body and the locomotion.1–3 Relying on the examples offered by animals, novel soft actuators and soft robots were fabricated in recent years, such as Octobot robots, 4 MagWorm, 5 and Dielectric Elastomer Actuators Robots. 6 Although these robots show advances in maneuvering, navigation in confined space, and motion with multiple degrees of freedom, the high performance in soft robots is still a challenge because of the intrinsic limitations of soft materials, especially in the high-speed locomotion, fast response, and high payload capability of movement. 7 In contrast, most soft microrobots can only perform single locomotion, such as walk, climb, and run, which limited the application in the complex space.
However, their natural examplers exhibit unimaginable performance on locomotion precision, speed, and response. Instead of fabricating the complex soft materials to be assembled as an artificial robot, controlling the natural insects by exogenous stimulations provide a novel solution to perform a complex task at higher speed and higher flexibility.
Insect–computer hybrid robots refer to cyborg insects that combine artificial electronic equipment with living insects or organs.8–12 Such hybrid robots can perform complex and unique tasks under extreme conditions because of their small size and innate excellent locomotion ability. Over the last two decades, researchers have developed various methods and protocols to electrically stimulate muscles,13,14 sensory receptors (such as antennae), 15 or neural tissue (such as ganglia) 16 and demonstrated the motor movements that they respond to, including the initiation and turn in insects' walking and flying.13,17,18 Several motor behaviors, such as detecting obstacles, avoiding dangers, and exchanging information, can be produced and regulated by antennae in insects. 19
The study of obstacle avoidance and escape behavior has progressed significantly. When insects turn their antennae to probe the space ahead, the antennae are deformed as they collide with obstacles. The mechanoreceptors in the antennae's base joints then detect the change in base angle and generate depolarization. These nerve impulses are then transmitted to the central nervous system (CNS), which collects information from the rest of the sense organs and encodes movement instructions into nerve pulses.20,21
Furthermore, the motor neurons receive these instructions and elicit the muscle to contract. Finally, the insect turns to avoid obstacles or dangers. Electrical stimulation of the antennae, for example, can elicit cockroaches or beetles to turn and walk.15,22 Insect cercus has a similar operational principle. 23 When an insect's cercus bends, it can cause various motor behaviors, such as walking forward in a cockroach 24 or adjusting the leg posture of a locust while flying. 25
However, although jumping is a widely used movement in insects, there are few successful demonstrations of jumping control on insects. Giampalmo elicited American grasshopper to jump by stimulating the metathoracic T3 ganglion. 26 Most studies have focused on the walking or flying movements of beetles or cockroaches. Locust can not only fly long distances on their wings but also walk and jump on the ground. Meanwhile, the locusts reveal excellent surface adaptability for takeoff, and they can jump and land on various spatial surfaces using their unique claws and foot pads to overcome obstacles for hunting and avoiding predators.27,28
Moreover, the locusts show extremely strong explosive power, jumping several times the height and length of their body in a short time. This is due to a combination working of the flexor, the extensor, a lump of cuticle on the inner surface of the distal end of the femur, and a semilunar process (SLP) at the femorotibial joint.29–33 Before the jump, the extensor and flexor muscles of the hind legs of the locust will contract together for 200 to 500 ms, which is the cocontraction stage. 34 At this stage, the lump of cuticle clamps the flexor apodeme and stops the hind legs from extending. 35
At the same time, the SLP in the knee joint of the hind leg is deformed to store the energy under cocontraction, to overcome the limitation that the maximum force and speed cannot be produced at the same time and hence realize the stretch for high force and high speed needed to propel the jump.29–31 By transforming a live locust into a hybrid robot, the dexterous jump movement can be retained. Thus, the insect–computer hybrid technique can improve the robot's ability to overcome large obstacles by controlling the locust's jump movement. Moreover, precise jumping control cannot be achieved without directional control. A control model containing a gradient relationship between stimulus parameters and angular velocity is on-demand for precise turning control.
This study presents a novel microjumping robot based on the Oriental Migratory Locust (Locusta migratoria manilensis, Meyen 1835), which is the first-ever demonstration of the omnidirectional jumping control of insect-compute hybrid robots (Fig. 1 and Supplementary Movie S1). The steering and jumping of the locusts were achieved through electrical stimulation of the antennae and cercus, respectively. The locust's antennae and cercus are multichannel sensory organs that consist of various sensory receptors, including mechanical, chemical, vibration, and thermal receptors. The electrically induced jumping performance approximates the locust's natural jump in terms of distance, height, and force.

Overview of a cyborg locust.
Following other insects, the electrical stimulation of the antennae induced body rotation promptly. Meanwhile, a wireless microstimulator backpack is designed to address the remote control of a locust. Because the turning velocity reveals gradient effect as a function of stimulation frequency, a feedback proportional-derivative control system was built to rotate the heading direction of the hybrid robot to a predetermined angle and then jump away under remote control. The whole hybrid system weighs ∼2.7 g in total (inclusive of insect, backpack, and battery) and can turn, jump, and land on uneven ground with a short body length of 5 cm. Given its superior maneuverability, we would like to use this novel jumping robot in search and rescue missions shortly. The use of this animal and the methodology have been approved by the Animal Ethical and Welfare Committee (IACUC-2020026).
Results and Discussion
Kicking under the mechanical and electrical stimulation of the cercus
The cercus of a locust can detect external mechanical stimuli quickly and respond with powerful kicking movements. Under mechanical and electrical stimulation of the cercus, the time sequence of bioelectrical signals in the terminal ganglion, the rapid extensor nerve (N5) in the metathorax motor ganglion, and the extensor muscle in the hind leg were measured (Fig. 2A, B and Supplementary Fig. S1). In a mechanical stimulation experiment on the cercus, a probe in slight contact with the locust's cercus was swung 4° with the motor, causing the cerci to bend ∼3 mm toward the head (Fig. 2C and Supplementary Data). Figure 2E shows a trial in which a locust kicked in response to a mechanical stimulus. After the cerci was bent, the terminal ganglion of the locust began to produce nerve signals in 36.3 ms, and the nerve impulses were generated by the N5 in 87.3 ms.

Comparison of attached kicking and electrical stimulated kicking.
In addition, the first spike of electromyography (EMG) signals was generated by the extensor muscle of the hind leg in 104.1 ms. At this point, the cocontraction stage began. Then the hind leg tibia began to extend at 243 ms. After 256 ms, the Femur-Tibia (F-T) angle reached its maximum angle of 158.9° (Fig. 2E). Typically, a larger F-T angle corresponds to a stronger angular momentum and torque.
Based on the responses to mechanical stimulation, electrodes were implanted to perform electrical stimulation on the cercus, and electrical stimulation on the cercus can successfully induce ganglion response and trigger jump. In the electrical stimulation experiment, the locust's cercus was electrically stimulated with 400- and 200-ms pulse signals (Fig. 2D).
Figure 2F and G shows a trial in which a locust kicked in response to an electrical stimulus under 400- and 200-ms, respectively. The F-T joint of this locust could reach a maximum angle of 182.8° under 400-ms simulation, which was bigger than mechanical stimulation (Fig. 2F). Under 200-ms pulse, it was only 90° (Fig. 2G). Furthermore, the time of extension under 400-ms pulse was only 10 ms from the start of the tibial joint extension to the maximum angle, whereas the time of extension under 200-ms pulse time was 122 ms. These significant differences imply that, under the approximate conditions, a 400-ms pulse can induce larger angular momentum and torque and ultimately affect the jump distance and height.
When the electrical stimulation signal of 400-ms was applied to the cercus, the EMG signal of the hindleg extensor muscle lasted for 200-ms before the tibial joint was kicked out. Under the 200-ms stimulation, the EMG signal only lasted for 50 ms. The duration of the extensor muscles is related to the cocontraction time of the locusts, and the duration of the cocontraction will affect the kick strength. 31 The extension time under 400-ms pulse is only 10 ms from the beginning of tibial joint extension to the maximum angle, and the average angular velocity is as high as 21.6°/ms, while the extension time under 200-ms pulse time is 122 ms, and the average angular velocity is less than 1°/ms.
These significant differences mean that the EMG signal generated by electrical stimulation for 200-ms is short and cannot effectively induce the cocontraction process, while the electrical stimulation for 400-ms can effectively achieve the cocontraction process, the duration of which is related to the strength of the kick. This means that under similar conditions, the 400-ms electrical stimulation signal may induce greater angular momentum and torque in the hind legs and ultimately affect the jump distance and height. Therefore, the duration of 400-ms is suitable for electrical stimulation.
The functional sequence for the locust has been revealed in detail, from bending of filiform hairs on the abdominal cerci, stimulation of mechanosensory cells, conduction using giant fibers in the abdominal nerve cord to thoracic ganglia, and the CNS.36,37 However, few data on locust escape jumping are available, although the cercus and giant interneuron systems of locusts are anatomically and physiologically well understood and have been used in studies of flying attitude control.25,38
Although mechanical and electrical stimulation of the locusts' cercus produced kicking movements, the action was not similar. Electroneurography produced by the terminal ganglia during mechanical stimulation lasted ∼200-ms, whereas electrical stimulation of 200-ms did not induce the same kick effect. Under mechanical stimulation, physical touching on the cercus can cause the filiform hairs to bend, and the filiform hairs directly affect the mechanosensory, and the giant interneurons generate the nerve pulse, while some nongiant interneurons also produce neural signals. Compared with exogenous electrical stimulation signals, these integrated neural signals were transmitted together to the terminal ganglion and contain more complex coded information, 39 resulting in a fast kick.
Jumping through the electrical stimulation of the cercus
Stimulation of the cercus induces the locust to jump away from stimuli (Fig. 3A, N = 10 locusts, n = 48 jumps). Following the study of electrically stimulated kicking, a 400-ms electric pulse signal was applied to the cercus of a loosely tethered locust, and the locust performed the jumping response accordingly. In the experiment, the locust remained stationary before stimulation, and the hind legs began to flex 73 ± 10 ms after the cercus stimulation. The locust jumped away from the platform 545.15 ± 129.72 ms (Supplementary Table S1) after responding to the stimulation. Each locust was stimulated six times consecutively, and the overall success rate was 80.0%. The average jump height of the jumping trajectories is 111.59 ± 41.26 mm, and the average distance is 369.31 ± 94.69 mm.

Externally driven jumping and endogenous jumping.
Meanwhile, the maximum vertical acceleration was 83.00 ± 20.91 mN/g, and the maximum horizontal acceleration was 78.00 ± 22.08 mN/g (Fig. 3B). As a comparison, the natural jumps revealed a height of 81.66 ± 27.06 mm and a distance of 418.92 ± 99.24 mm on average. The maximum vertical acceleration was 71.67 ± 14.59 mN/g, and the maximum horizontal acceleration was 79.50 ± 17.29 mN/g (Fig. 3B, N = 7 locusts, n = 32 jumps). In terms of jumping height and distance, the locusts under electrical stimulation on cercus could achieve approximate jumping targets as those under the natural jumping, and the maximum horizontal and vertical acceleration during electrical stimulation jumping was also close to those of the spontaneous jumping, indicating that electrical stimulation on cercus could achieve approximate jumping ability.
In the takeoff stage, the locust produces jumping force through the joint action of the flexor and extensor muscles of the hind leg, and the acceleration generated is directly related to the final jumping height and distance. This hypothesis was verified by comparing the locust's jumping distance with the maximum acceleration during the takeoff. The linear relationship between jumping distance and maximum horizontal acceleration was verified in stimulated and spontaneous jump (stimulated jump: y = 0.20x + 4.85, R2 = 0.7218; natural jump: y = 0.15x + 16.56, R2 = 0.7438). Similarly, the linear relationship between the jumping height and maximum vertical acceleration was verified (stimulated jump: y = 0.47x + 30.50, R2 = 0.8619; natural jump: y = 0.51x + 30.23, R2 = 0.8860) (Fig. 3C).
The height and distance of a locust's jump are determined by various factors, including the locust's characteristics, 40 kicking strength of the hind leg muscle,41,42 and the angle at which the locust takes off. 43 The cocontraction time was the main factor in determining the kicking strength of the hind leg muscle. 31 The cocontraction time was instructed by the CNS. The distributions of height and distance in natural jumps show that the locusts can alter jumping distances and heights based on information gathered around them.
In addition, we severed the nerve fiber between prothorax and mesothorax, as well as the nerve fiber between mesothorax and metathorax. These locusts failed to jump under identical electrical stimulations. Therefore, instead of acting directly on the hind leg muscles, nerve impulses generated by electrical signals at the terminal ganglia were transmitted to the CNS. Then the brain synthesizes global information and sends motor commands to the corresponding motor neurons, affecting the cocontraction time and producing different jumping effects. Furthermore, this method of eliciting jumping causes no damage to the locust's nerves and muscles. The locust retains its ability to gather information about its surroundings, changing its jumping decision to overcome or avoid obstacles.
Finally, locusts can sense the airflow changes caused by the movement of natural enemies in their cercus. Due to the survival instinct of living creatures, locusts can quickly avoid the stimulus source of the cercus by producing jumping motions. Taking advantage of this, electrical stimulation of the cercus can induce the locust robot to jump.
Turning response to antenna stimulation
Stimulation of one side of antenna induces the locust to rotate in the opposite direction. During the stimulation, the locusts kept turning and their heading angles changed from 0° to 76.60° ± 36.76° for the right antenna stimulation at 60 Hz (Fig. 4A, N = 10 locusts, n = 10 trials). After the stimulation was terminated, the locusts stopped and kept stationary after turning another ∼8° because of the moment inertia. The locust's response time before turning was ∼0.14 s. During this period, locusts sensed the antenna stimulation and activated motor neurons for turning movements.44,45 The angular velocity of locusts toward the desired direction increased linearly during the first ∼0.4 s and then slowly increased to the maximum turning rate of 94.40 ± 31.29°/s (Fig. 4B).
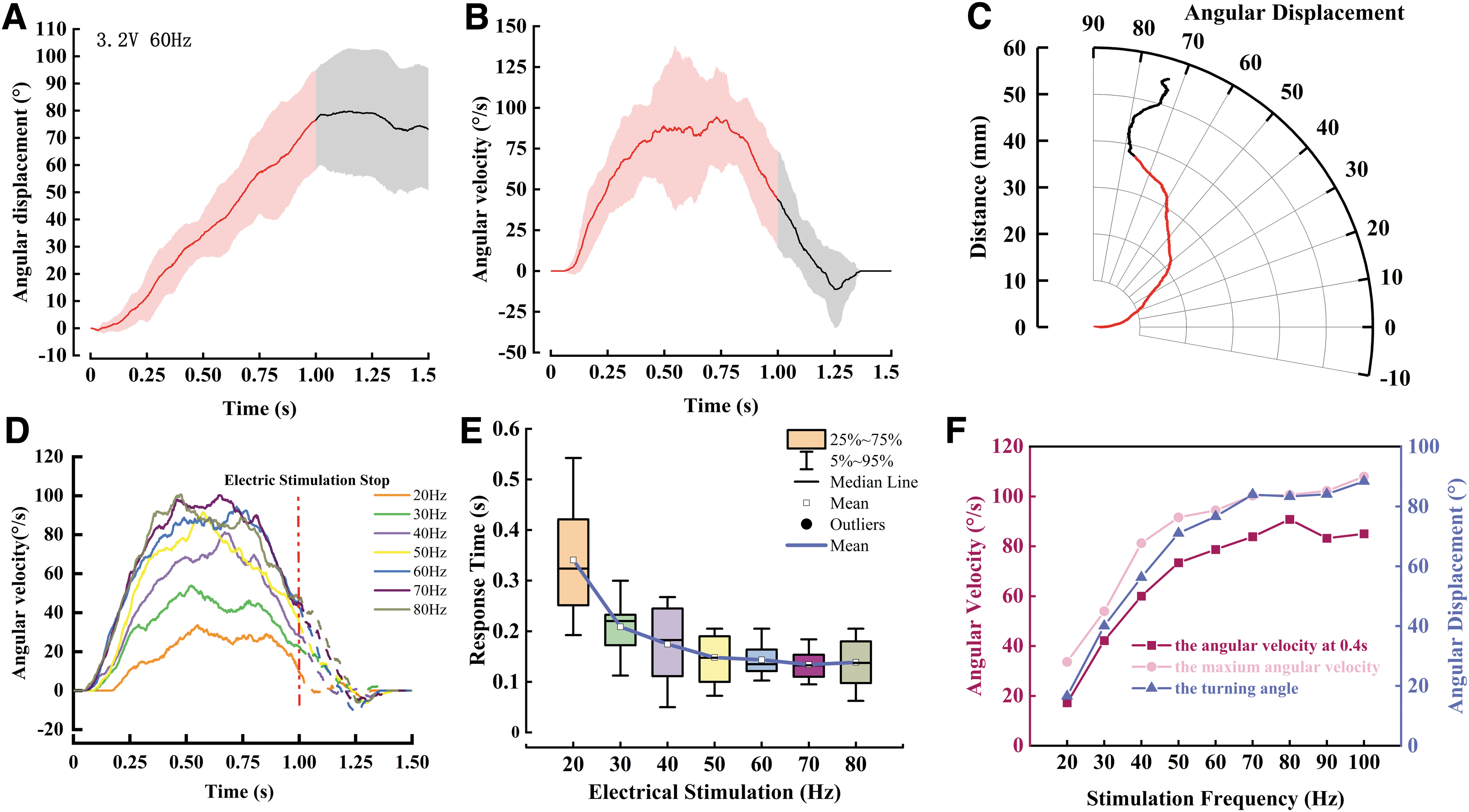
The locust's responses to the electrical stimulation on the antennae.
After the peak, the angular rate fluctuates and gradually dropped down to 43.85°/s when the stimulation was terminated. After electrical stimulation, the turning rate took another ∼0.3 s to return to 0°/s because of the turning inertia. It is also found that the locust did not reveal apparent sideways walking while rotating under electrical stimulation (Fig. 4C).
The graded reaction on turning as a function of stimulation frequency was observed (Fig. 4D and Supplementary Movies S2 and S3). The rotation angles (Spearman's correlation test, ρ = 0.68, p < 0.01, df = 51; Supplementary Table S3) and angular velocities (Spearman's correlation test, ρ = 0.63, p < 0.01, df = 51; Supplementary Table S3) increased monotonically as the stimulation frequencies increased from 20 to 70 Hz. In the electric stimulation period of 1 s, the locusts showed a turning rate of ∼17°/s under 20 Hz, which is smaller than the turning velocity under higher frequencies. As the stimulation frequency increased to 70 Hz, the locusts showed a stronger response, with a turning rate of ∼80°/s (Fig. 4F and Supplementary Figs. S4 and S5).
These turning responses could be due to the activation of sensory neurons distributed on the insect's antennae, which could trigger the escape or avoidance behavior under a mechanical touch.22,46 We believe that the electrical signals simulated the neural signals of mechanical touch, thus triggering the insect's escape reaction and producing the contralateral steering action. The locust with antennae stimulation, similar to other insects, had an inherited avoidance strategy, which could lead to a rapid acceleration when identifying danger and then maintaining a high speed to avoid danger.
Meanwhile, under different intensities of electrical stimulation, a gradient relationship between turning velocity and response time was observed. In the antennae of the cockroach, a single spike is elicited under a slight touch of either the ipsilateral or contralateral antenna, whereas about 10 action potentials are elicited by a stronger touch.47,48 The steering speed is considered to be related to the number of nerve emissions. Thus, the graded responses to stimulation frequency imply that the locust's antennae can trigger a graded willingness to escape.
The response time to the antenna electrical stimulation was defined as the time interval between the start of the stimulation, the timing of turning rate >13°/s, and the maximum swing speed in the free walking. Under low frequency, the locusts could be elicited to turn at a faster response time through increasing frequency. In addition, the response time cannot be reduced once it has reached 0.14 s, the least time to initiate the turning (Fig. 4E). The response time under different frequencies reflects a response time limit in the whole process, from the antenna stimulation to the start of steering action, including neural signal production in antennal-mechanosensory neuropils, transmission to the thoracic ganglion, and the contraction of leg muscles. 20
The left/right turning control of the locust robot can be achieved by electric stimulation on the right/left antenna. The locust can detect obstacles using the antennae. After being pressed and bent, the locust changes the current direction of movement to avoid the obstacle, so a contralateral turning movement occurs.
Collaborative turning and jumping control
The accurate jump control of a locust can be achieved by the collaborative electrical stimulation of antennae and cercus. To assemble a biohybrid jumping robot, we installed a custom-made wireless electronic backpack on the locust's tergum. The backpack can generate multichannel electrical stimulation signals for steering and jumping control, based on the commands of the feedback control system (Fig. 5A–D and Supplementary Figs. S2 and S3). First, the locust was steered using antenna stimulation. After reaching the preset direction, the locusts were elicited to complete the jump action under electrical stimulation to the cercus.

Setup of the feedback control experiment.
A successful feedback-controlled trial in which the locust jumped after turning 100° was shown in Figure 5E. The cyborg locust turned to the objective direction under seven stimulations on the antenna. After the last electric pulse on antenna, the locust jumped under the trigger of cercus stimulation (Supplementary Movie S4).
The feedback-controlled jumps of the locust toward 50° and 100° targets were conducted (Fig. 6, N = 14, n = 51 trials). Toward the 50° target, the locust was stimulated 4.7 ± 2.05 times while rotating 49.45° ± 2.75° in 1.92 ± 0.81 s (Supplementary Fig. S8). Toward the 100° target, the locust was stimulated 7.51 ± 2.62 times while rotating 99.54° ± 2.45° in 3.13 ± 1.1 s (Fig. 6A, B). The feedback control system achieved a jumping distance of 334.18 ± 122.64 mm and a jumping height of 94.12 ± 55.8 mm (Fig. 6A, C). The success rate of turning to the expected direction was 94.3% in 84 trials, and the overall success rate with jumping control was 60.71%.
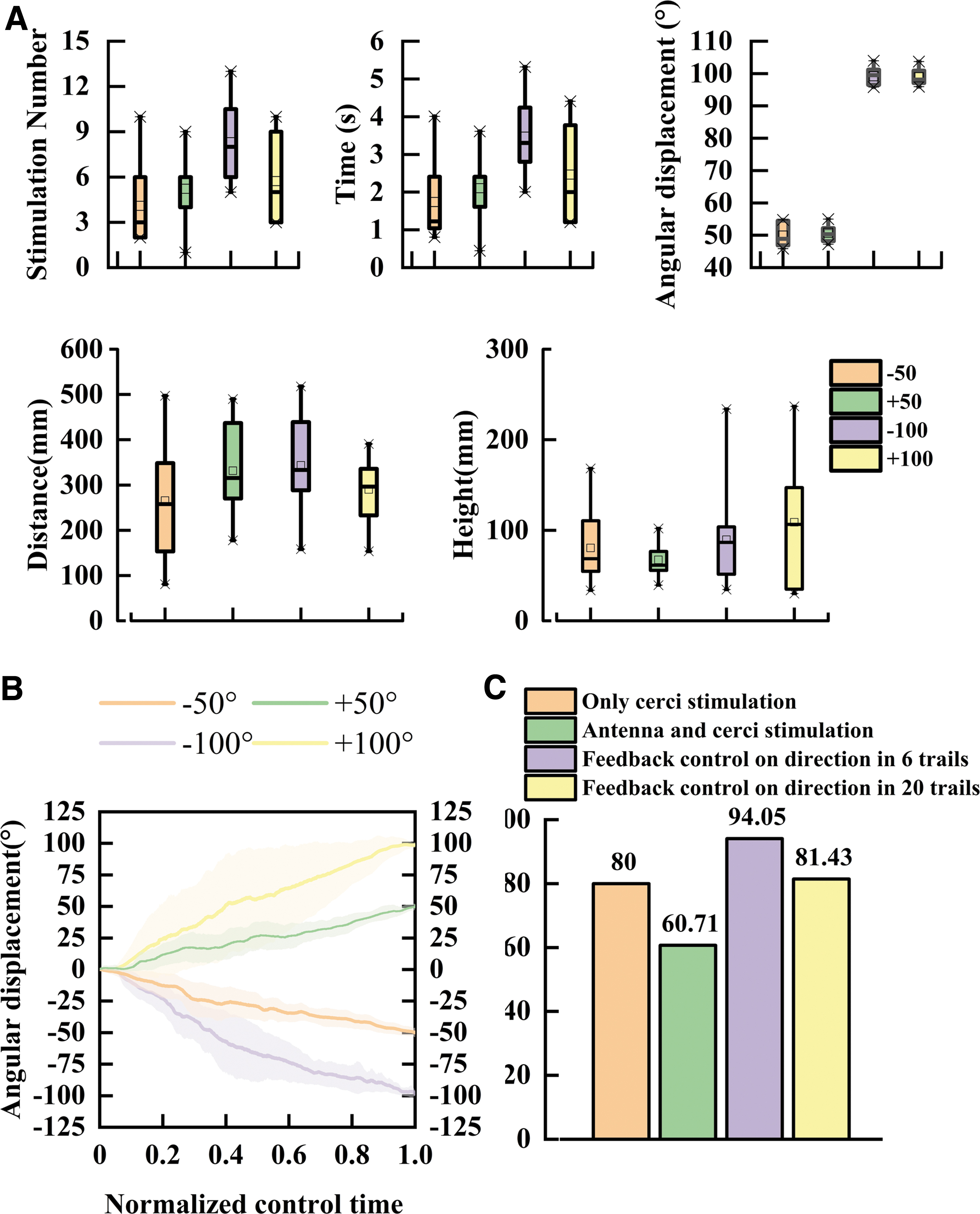
The performance of cyborg locusts under different targets.
A failed stimulation is typically caused by two conditions. One condition is that the locust performs a powerless jump after stimulation. Although the locust kicks the legs, the jump height is lower than the body length, which is not an effective jump. It happens after repeated jumps, so it might be caused by fatigue. In the other condition, there is no jumping response after stimulation. It usually happens right after a successful jump. Thus, it might be caused by the failure of implanted electrodes. In the landing process, the locust's abdomen hits the ground, causing a potential drift of the electrode. Specifically, in clockwise rotations, 28 out of 42 trials were successful, while in counterclockwise rotations, 23 of 42 trials were successful. There was no significant difference found between clockwise and counterclockwise rotations.
The steering control revealed better durability than jumping control. After repeating the cercus stimulation six times consecutively, the locust's response to the stimulus decreased sharply and they jumped seldomly. However, the antenna stimulus for turning did not attenuate apparently and can be used for much more trials. The pure steering feedback control experiments were performed under 100° rotation (Supplementary Fig. S9, N = 7, n = 140 trials).
The overall success rate was still 81.42%. Considering that the number of stimuli required for 100° rotation was 7.51 times, and thus, the total number of electrical stimuli on antenna was ∼150 times, there is no significant decrease in turning velocity with the increase of stimulus. It is different from the cockroach's antennae which showed adaptability under 10 electrical stimuli. 49 This feature implies that the electrical stimulation does not damage the nervous system, and the locust does not generate obvious habituation, which is an ideal platform to make the insect-computer hybrid robots. 50 Thus, the control of turn-and-jump in locust reveals latent application values.
Low power consumption and ease of use
Under single electrical stimulation, the average power consumption of antenna was 186 ± 27.1 μW and that of cercus was 124.8 ± 35.2 μW (Supplementary Fig. S7). The power consumption of this biological actuator was remarkably low. The maximum power consumption for a 100° turn-and-jump would be 1.427 mW, which is mainly for the microcontroller, compared with artificial miniature legged robots, whose consumption was 100–1000 mW. 51 Meanwhile, compared with the mechanical microjumping robot, the locust has a strong load capacity with a maximum payload of 2 g. Excellent load capacity offers the possibility of adding tiny devices such as cameras, chemical odor sensors, and ultrasonic transmitters that can be used in rescue missions.
Recently, many jumping robots have been developed, including JumpRoach, 52 Tribot, 53 Surveillance Robot, 54 MSU tailbot, 55 Salto-1p, 56 and Flea. 57 With advancements in manufacturing technology, microactuators, and biomimetic technology, existing miniature jumping robots can achieve longer jumping distances and higher jumping heights while remaining as small and light as insects. 58
Noh et al. 57 designed a flea-inspired jumping robot with a 1.1 g weight and 2 cm body size through using the shape memory alloy (SMA) spring actuators. Zaitsev59–61 developed a miniature jumping robot that can complete a single jump using the locust bionic research. Following that, the robot, which has a propeller-shaped tail, can adjust its aerial posture to achieve a smooth landing. Consequently, the added tail lengthens the body to 25 cm, five times the length of a locust. Zhakypov et al. 53 developed a 10 g origami robot Tribot by studying trap-jaw ants. The Tribot can adjust the angle of its three legs to change its mass center and thus improve postlanding stability, but it cannot change the direction of takeoff.
Overall, the microjumping robots still have many shortcomings in jumping controllability, such as fixed takeoff direction, uncontrollable jump speed, lack of landing stability, and inconsecutive jumps. 62 Especially, the controlling of jumping direction and attitude recovery after landing are the keys in designing jumping robotics.
However, our insect robot can better solve these two problems. The locust robot is designed to be less than 3 g (Supplementary Table S2) and reveals a good height/mass ratio (Supplementary Fig. S6). Because of its lightweight, the robot consumes less energy at the same jumping height; in addition, the robot is less vulnerable to landing impacts. Because of its soft tissue, the locust robot can well buffer the impact of landing, to ensure a smooth landing and avoid body overturning. Meanwhile, even if the body rolls over, the cyborg locust can recover its body automatically from the landing position without human intervention. Finally, the cyborg locust can jump continuously in any direction without requiring institutional preparation, such as heating SMA or compressing spring.
For robot applications, the stability of movement is a priority. In the jump control, the locust's abdomen may collide with the ground after the jump, resulting in the offset of the implanted electrode, thus affecting the implementation of the jump control. Simultaneously, the current electrode cannot meet the requirements for precise location, and there is a risk of nerve fiber damage.
In the future, the corresponding materials of electrodes, implantation methodology, and improved wireless electrode fixation to the implant site 63 can be used, just as the “StimDust,” 64 which is only 1.8 mm2. Above all, the results achieved through the system in this experiment were encouraging and instructive. The current system's architecture had the potential to enable the development of multimode control cyborgs (such as visual control with a camera and the brain-computer interface control), 65 and increasing the control modes could contribute to improvements in independent or online control, which would be designed and validated in further study.
Conclusion
In this study, we demonstrated the first feedback jump control of a steerable insect-computer hybrid robot through electrical stimulation of the antenna and cercus. The locust can be induced to jump with the height of ∼111 mm and the distance of ∼369 mm through the electrical stimulation on the cercus. Meanwhile, steering control induced by antennal stimuli showed gradient characteristics. The turning rate of locusts increases as the frequency of electrical pulses increase. The graded protocol appears to be a safe and efficient method of modulating locusts' heading direction, which is important for a precise jump.
Meanwhile, we found that the insect had the least response time of 0.14 s to initiate turning after stimulation. Finally, combining antennae and cercus electrical stimulation resulted in an omnidirectional biohybrid jumper. With the help of a wireless stimulator backpack, the feedback control on turning and jumping movements was demonstrated. With the ongoing improvements on electrode implantation and fixation, we believe the cyborg locust can become more reliable and stable and will be applicable in search and rescue missions in the future.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.