
Abstract
This paper explores online stiffness modulation within a single tail stroke for swimming soft robots. Despite advances in stiffening mechanisms, little attention has been given to dynamically adjusting stiffness in real-time, presenting a challenge in developing mechanisms with the requisite bandwidth to match tail actuation. Achieving an optimal balance between thrust and efficiency in swimming soft robots remains elusive, and the paper addresses this challenge by proposing a novel mechanism for independent stiffness control, leveraging fluid-driven stiffening within a patterned pouch. Inspired by fluidic-driven actuation, this approach exhibits high bandwidth and facilitates significant stiffness changes. We perform experiments to demonstrate how this mechanism enhances both thrust and swimming efficiency. The tail actuation and fluid-driven stiffening can be optimized for a specific combination of thrust and efficiency, tailored to the desired maneuver type. The paper further explores the complex interaction between the soft body and surrounding fluid and provides fluid dynamics insights gained from the vortices created during actuation. Through frequency modulation and online stiffening, the study extends the Pareto front of achievable thrust generation and swimming efficiency.
Introduction
Fish and other swimming animals demonstrate a wide range of swimming behaviors, 1 showing high thrust rapid swimming to escape predators or performing precise maneuvers, and also more energy efficient swimming for traveling extreme distances for migration.2,3 While this arises from many different aspects of their bio-mechanics including their tail morphology, fin structures, and muscle activation patterns, 4 their tail stiffness plays a large role.5,6 Many fish species (tuna notably being one 7 ) have the ability to adjust the stiffness in their tails, and this flexibility is crucial for their swimming performance and maneuverability. Fish can modulate the stiffness of their tails in response to various factors, including changes in swimming speed, environmental conditions, and the need for precise control during different activities. This adaptability allows fish to optimize their swimming efficiency and respond to dynamic aquatic environments. 8 Investigating and exploring mechanics and control approaches for stiffening could provide bio-inspired swimming fish robots,9,10 with a similar range of functionality and behavior. However, this requires mechanisms for rapidly modulating stiffness and also means of analyzing and optimizing stiffness change.
There has been a significant body of work related to stiffening mechanisms in robotic fish. 11 This includes a number of different mechanisms that seek to tune the stiffness of the tail. This includes those which are actuated through antagonistically controlled stiffness by tendons, 12 or antagonistic pairs of soft actuators. 13 Others exploit structurally controlled stiffness for online change, for example, by extending a rigid sleeve over a spring, 14 or an adjustable robotic tendon. 15 There are many mechanical approaches that have been implemented in fish-like robots, the most notable of which uses the pre-tensioning of internal springs connected to a peduncle to vary tail stiffness.16,17 These mechanisms can also be used together and decoupled for advantageous effects. 18 These mechanisms have been incorporated into robots and have shed light on the relationship between tail stiffness at different actuation frequencies, and the effect on swimming thrust, velocity, and efficiency. Compliance has been demonstrated to aid ‘high performance’ swimming, yet this focuses on maximizing only thrust or swimming speed. 19 The dynamic distribution of stiffening has also been shown to be effective for affecting a swimming robots performance. 20 Investigating the trade-off between thrust and efficiency, 21 it has been demonstrated how tunable stiffness can enable fast and efficient swimming by modulating the stiffness appropriately for the swimming frequency. 16 Despite progress in stiffening mechanisms and their exploitation, there has been limited exploration of online, temporal stiffness modulation within a single tail-beat, posing challenges in developing mechanisms with bandwidth-matching tail actuation. Table 1 compares several recent robotic fish platforms, showing their stiffening mechanisms, speed of stiffening, and dynamic stiffening capabilities, thereby motivating the need to investigate stiffening within a tail-beat. In addition, fully leveraging stiffening requires evaluating performance across a spectrum of mapping efficiency and thrust generation, presenting a formidable balance for swimming soft robots. The intricate interplay between a soft body and surrounding fluid poses another challenge, 26 as hydrodynamics simulations are resource-intensive and may lack precision in real-world settings. 27 The complexities of fluid-structure interactions in swimming complicate the design process, with traditional simulation approaches struggling to capture nuanced dynamics. This underscores the need for innovative methodologies. Understanding the intricate dynamics of soft robotic swimming calls for a comprehensive grasp of the complex relationship between morphology, stiffness modulation, and control strategies—an exploration pivotal in advancing the field.
Comparison of Stiffening Mechanisms and their Dynamic Capabilities in Various Robotic Fish Platforms
By modulating the stiffness of the tail within the tail beat period, we predict that the tail can help the robot cover a broader performance spectrum in terms of thrust and swimming efficiency compared to static stiffness configurations. Three key contributions summarize this work. First, our novel mechanism for independent control of the stiffness with respect to the tail morphology, which utilizes fluid-driven stiffening, leveraging the rapid change of fluid pressure within a patterned pouch. Inspired by fluid-driven actuation, 28 this approach offers high bandwidth and enables significant stiffness changes without the need for a closed-loop system, as it directly uses environmental fluid for stiffening. Secondly, by analyzing the vortices created during actuation, we can experimentally demonstrate not only the importance of stiffness but also the critical timing of stiffening at different points in the stroke of the fish’s tail. The findings from our approach are the final contributions to this work. We demonstrate how through frequency modulation and online stiffening during the actuation period, we can access and extend the Pareto front, with advantages to both the achievable speed (39% increase) and achievable swimming efficiency (34% increase). While biological fish alter the stiffness of their entire tail, including muscles and skeletal structures, our study focuses specifically on the caudal fin. Although the caudal fin is typically a passive structure, our design incorporates active stiffness modulation to explore its impact on swimming performance. This approach provides initial insights into the role of temporally modulated stiffness in bio-inspired robotic swimming, with future work aimed at integrating additional tail components.
In the remainder of the paper, we first present the stiffness-tunable fish tail design, and methods used to explore this design and co-optimize the pump sequence, followed by experimental results and fluid-tail interaction analysis through particle image velocimetry (PIV). We will discuss the results and conclude with suggestions for future work.
Methods
To achieve efficient swimming through stiffness change, we wish to quickly vary the stiffness in the fish tail, as well as find the ideal stiffening sequence for each swimming regime. The tradeoff between the swimming thrust and efficiency is generated through the tail-fluid interactions, which are dependent on the morphology of the tail actuated in a periodic motion at the root. The fabrication method of the fish tail is first shown, followed by a stiffness characterization of the tail between soft and hard. The experimental setup for data acquisition, actuation optimization, and analysis using PIV is shown, as well as finally the robotic hardware for free-swimming experiments.
Fabrication of the fish tail
To create a soft fish tail with adjustable stiffness for enhanced swimming performance, we utilize a technique involving a vacuum-sealed pouch enveloped in a thin layer of silicone (Fig. 1A), similar to printable pouch motors made from heat bonding. 29 The modulation of fish tail stiffness is accomplished through fluid-driven actuation (Fig. 1B).

The silicone casing that covers the inner vacuum-seal pouch optimally disperses water pressure, ensuring comparability among various inner pouch designs. Several inner pouch patterns are tested, which exhibited similar deformation properties when inflated; the final pattern is selected for its ease of fabrication and optimal water flow dynamics. Using CAD software, we draw the tail design with a maximum line thickness of 0.4 mm and extrude it by 0.2 mm. Subsequently, the STL file is produced and used with the Creality CR-10 Pro 3D printer. The vacuum pouch is affixed to the printer bed, with cooking paper secured above it. The heated printer nozzle is directly used to trace the tail design onto the cooking paper, effectively sealing the top and bottom layers of the vacuum pouch underneath. For this specific printer, the settings encompass a layer height of 0.2 mm, a printing temperature of 260°C, a build plate temperature of 0°C, a print speed of 5 mm/s, with the fan and cooling both turned off. In order to modulate the stiffness of the fish through fluid-driven actuation, the inner pouch, fabricated from the sealed vacuum sheet features two outlets. Water is pumped in through a silicone tube connected to a water pump, causing inflation and deflation of the inner pouch, thereby controlling the tail’s stiffness.
Stiffness characterization of the fish tail
To assess the efficacy of the fluid-driven actuation in modulating the stiffness of the fish tail, both stationary and actuated conditions of the tail are investigated in water inside a glass tank. The tail’s stiffness is evaluated in stationary conditions (Fig. 2) and during sinusoidal motion actuation at the root using a motor (Fig. 3). With the pump turned off, the tail exhibits a softer state, while activation of the pump makes it stiffer.
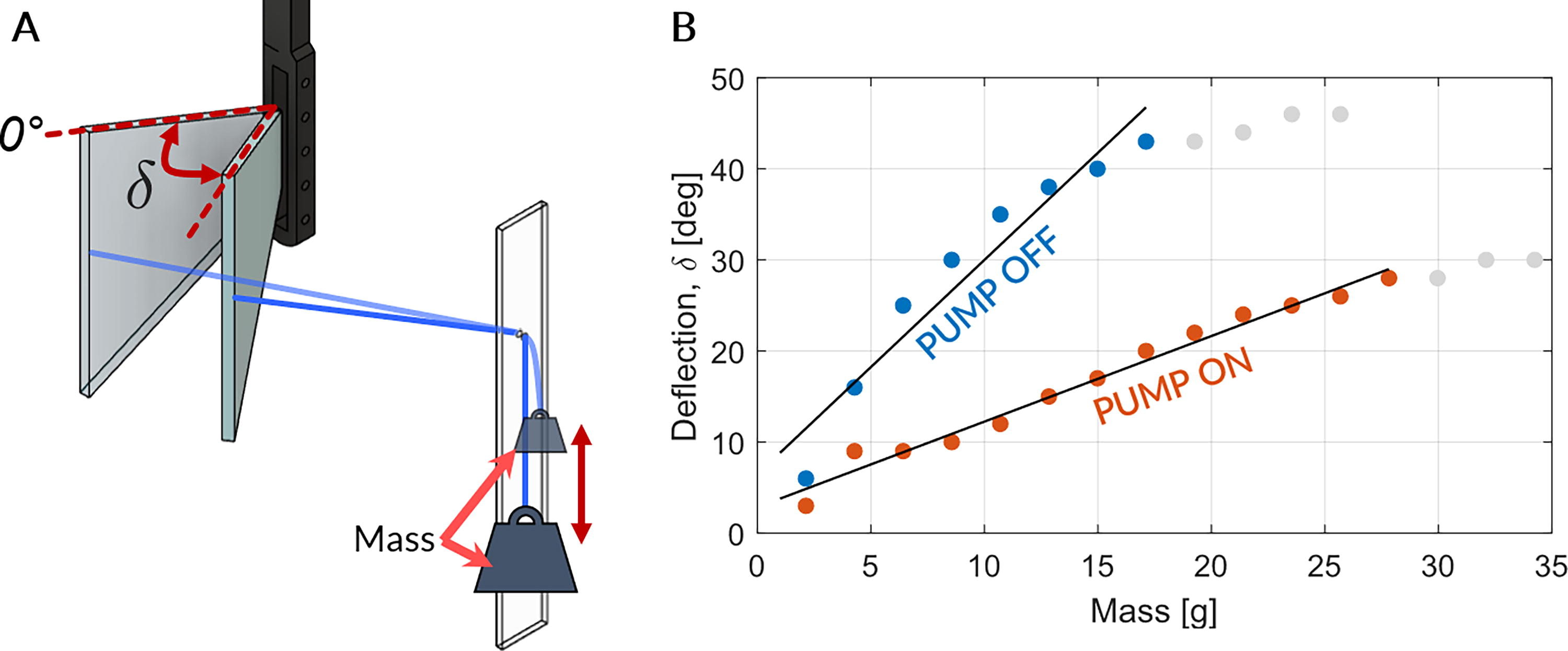

In stationary tests, we measure tail deflection by attaching weights of varying masses to the tip (Fig. 2A). Larger masses results in greater deflection, though the slope of the deflection-to-mass curve differs between pump-on and pump-off scenarios, as anticipated (Fig. 2B). The deflection curve exhibits a lower slope when the pump is activated.
We also investigate how stiffness influences tail deflection and movement during motorized sinusoidal actuation at the root of the tail. The tail angle, θ in relation to the motor arm, α and the tail phase offset, ϕ in relation to the motor position are investigated (Fig. 3A), revealing discernible differences between pump-on and pump-off states (Fig. 3B). The maximum tail angle that is observed for varying actuation frequency when the pump is off and on is shown in Figure 3C, and the tail angle phase offset for the same is shown in Figure 3D.
Quantifying and characterizing these parameters is important, as the interplay of stiffness and associated lag influences the physics of thrust generation during swimming. These insights are crucial for optimizing the tail’s efficiency in propelling through the fluid, akin to the flicking motion observed in natural fish swimming.
Static experiment for the fish tail
Experimental setup
To gather experimental data of the generated thrust and torque required to actuate the fish tail in water, an experimental setup (Fig. 4) is created. A DYNAMIXEL XL430-W250-T servo powers the mechanism to actuate the root of the fish tail in a sinusoidal sweep and a 0–3 kg load cell is used to measure the forward thrust as well as the torque (Fig. 4B). The water tank size is 60 × 30 × 30 cm. The tank size has been chosen to be larger than the tail to minimize tail effects.

The same experimental setup can be used to perform a PIV analysis on the fluid-tail interactions, as shown in Figure 4A. This setup enables accurate measuring and visualization of the fluid flow velocity fields by tracking the motion tracer particles suspended in the fluid. A laser diode and a glass rod are used to create a sheet of laser where the particles are visible and the particles are recorded using a camera at 60 FPS from the bottom of the water tank. Using MATLAB PIVlab, 30 velocity vectors are extracted from the flow field around the fish tail by correlating the displacement of seeded particles in sequential image frames through cross-correlation. The vorticity obtained is used to quantitatively show the performance benefits of temporal modulations in stiffness.
Furthermore, the pumps, each with a flow rate of 240 L/h, can inflate the tail pouch with a volume of 20.25 cm3, in approximately 0.15 s, and deflate it instantaneously when hydraulic pressure is no longer supplied. Four chosen pump actuation sequences as shown in Figure 5A, are plotted along with a single cycle of motor position. The motor position varies from –20º to 20º from the center position (0º).
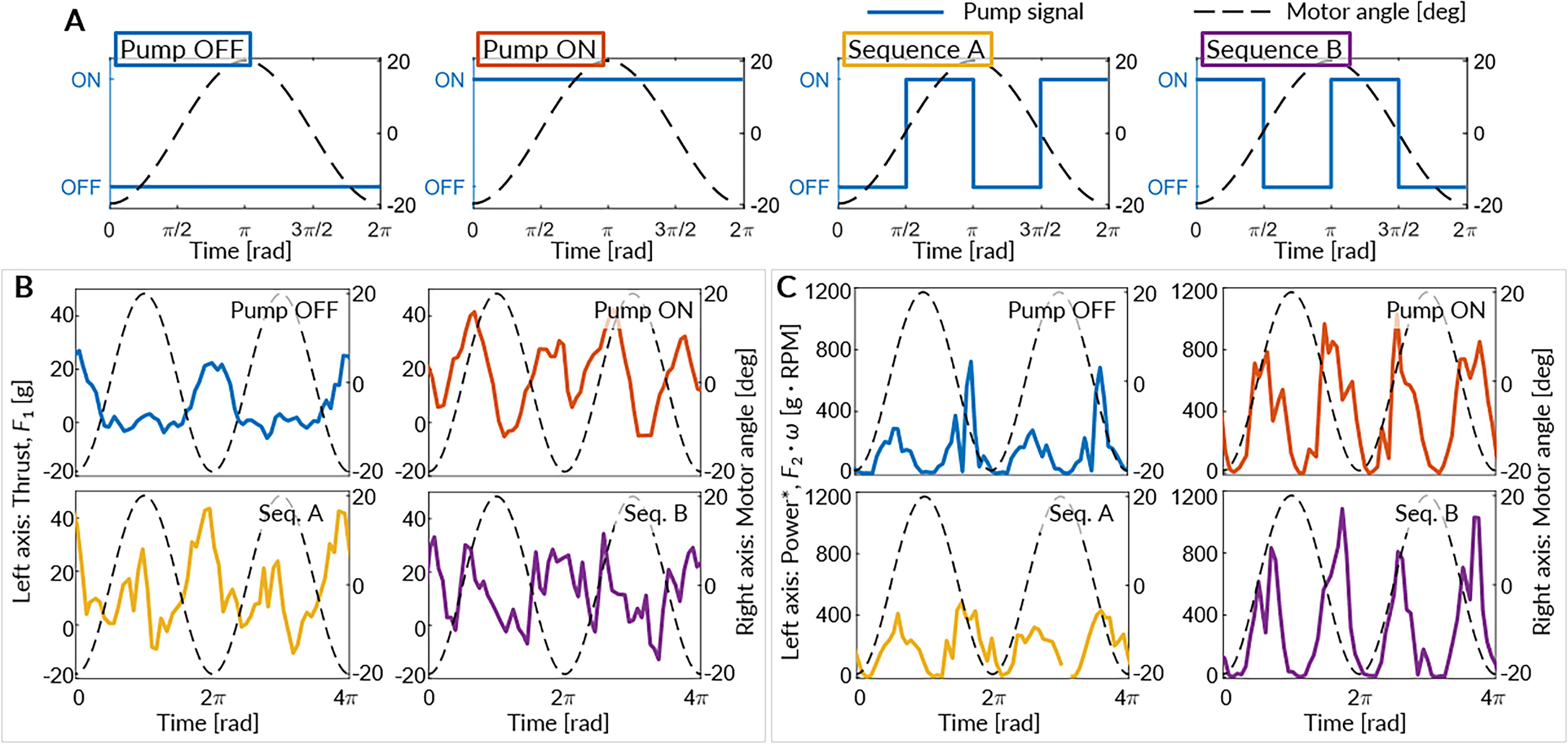
Pump OFF and Pump ON are chosen as standard examples of the tail because they remain either stiff or soft throughout the tail strokes. Sequence A and Sequence B are chosen while considering the hydrodynamic propulsion observed in a biological fish’s swimming motion.
31
While a fish’s hydrodynamics is inherently complex, Sequence A is tailored to modify the tail during the phase that generates forward thrust, aligning with the forward-facing vector of water resistance force. Specifically, the pump is inactive as the tail moves from the outer to the central region (motor positions –20º to 0º or 20º to 0º), and active as the tail moves from the central to the outer region (motor positions 0º to –20º or 0º to 20º). Sequence B is the reciprocal counterpart to Sequence A, so we can observe the dynamic interplay in the hydrodynamic propulsion process. The period of the tail actuation is chosen such that the transition time between soft and rigid states is much smaller, rendering it negligible for the practical implementation of the proposed mechanism in continuous, dynamic scenarios. A representative raw time series obtained from the load cells for two motor position cycles is contrasted for the four pump actuation sequences. The thrust, F1 is shown in Figure 5B, and a parameter representative of power (thus Power*),
Performance metrics of the fish tail
Several parameters are analyzed to quantify the performance of the fish tail, including the thrust produced and swimming efficiency. For the fish tail, the performance is evaluated in a glass tank. The setup is shown in Figure 4 and explained in the following sections. One of the important performance metrics is the output thrust, F1, produced by the tail when actuated, and is obtained from a load cell. Another metric is efficiency,
Free-swimming experiment of the robotic hardware
Robotic hardware
In Figure 6, the robotic hardware floats in a paddling pool measuring 200 × 300 cm. Similar to the static setup, a servo motor imparts a sinusoidal motion to the fish tail at the root. The input power driving the robot’s fin is measured using a load cell. The robot swims free of tethers due to its on-board battery and Raspberry Pi. The integrated pumps can be activated or disabled, allowing control over the inflation and deflation of the tail pouch. This dynamic control mechanism enables precise management of tail stiffness throughout the entire swim or within each stroke. The velocity of the robot is extracted from video recordings captured during each swimming experiment.

Robotic hardware with on-board controls and the integrated stiffness-tunable fish tail.
Performance metrics for the robot
For the robotic hardware with the integrated stiffness changing fish tail, the performance is evaluated in a paddling pool (shown in Fig. 6 and Fig. 10E). Similar to the fish tail setup, two performance metrics are considered. One of them is the swimming velocity, U, a metric similar to the thrust metric of the tail. The velocity is obtained by post-processing of videos taken during the pool experiments. Another metric is the efficiency,
Results
Performance of fish tail with various stiffness sequences
We initially assess the fish tail’s performance in a static experimental setup, manipulating pump sequences to induce varied stiffness patterns within each tail stroke. Our mechanism enables not only the maintenance of constant stiffness (either soft or stiff) throughout experiments but also rapid adjustments in stiffness sequences through fluid-driven pumps.
Averaged thrust (Fig. 7A) and efficiency (Fig. 7B) metrics are presented for the pump sequences across diverse motor actuation frequencies, spanning a sinusoidal motion range of −20º to 20º. Notably, when the pump is off (soft tail), thrust remains constant and low across all frequencies, while the steepest slope occurs with increasing frequency when the pump is on (stiff tail). When the pump sequence is active within a stroke, the slope is lower than Pump ON but still exhibits a positive relationship with increasing frequency.

Results for the static experiment setup for fish tail with the four sequences of pump actuation and a sequence chosen from Bayesian optimization.
Efficiency peaks at lower frequencies when the pump is off and dips when the pump is on, aligning with expectations that stiffer tails demand more power to navigate through water due to increased fluid resistance. Sequence B closely parallels Pump OFF in efficiency, whereas Sequence A demonstrates high efficiency across most frequency ranges.
To elucidate a performance Pareto front for the fish tail, we compare efficiency and output thrust on the same plot (Fig. 7C). The optimal points emerge from a combination of pump sequences at different frequencies: Pump OFF performs well with lower frequencies, Sequence A excels at medium frequencies, and Pump ON at higher frequencies. This outcome implies that by strategically leveraging the interplay between actuation frequencies and pump sequences, the fish tail can consistently attain optimal performance tailored to specific maneuvers.
Bayesian optimization of pump sequence control
As previously noted, we select pump sequences inspired by biological fish. However, to confirm the effectiveness of these pump sequences in mimicking fish behavior, we employ Bayesian optimization to find a pump sequence that maximizes both efficiency

The optimization process uses equation 3 as its objective function, considering the maximum and minimum thrust values and efficiencies from previous experiments, scaled from 0 to 1. While we aim to balance the optimization for both thrust and efficiency, initial trials reveal a tendency to get stuck in a local minimum where efficiency is high but the thrust is low. To address this, we slightly prioritize thrust in our scaling. We optimize two parameters: Para 1, the duration the pump stays on during a tail cycle, and Para 2, the timing of the pump activation within a cycle. The optimization is carried out for a 3-second actuation period (p = 3 s), as constrained by equation 4. Para 1 and Para 2 both have optimizable ranges of 0 to
With an exploration ratio set at 0.5, the optimizer explores a broad range of values until convergence at iteration 10. The maximum objective function is achieved at iteration 12, with Para 1 set at 0.5 s and Para 2 at 0.8 s. Interestingly, when compared to pump Sequence A, they appear quite similar at a cycle period of 3 s (Fig. 8B). Examining Figure 7, the Bayesian optimization line closely aligns with Sequence A but shows slight improvement. This suggests that the chosen sequence is indeed a robust and effective choice.
Fluid and fish tail interactions
To gain insights into why Pump Sequence A exhibits superior performance for a large range of actuation frequencies, we compare it with a chosen baseline, namely Pump ON, where the tail remains consistently stiffened. Figure 9A shows an example flow field series of snapshots from a PIV experiment for one period of motion, whereas Figure 9B shows the vorticity at various points while the fish tail undergoes a 3-second actuation period. Key points in time are highlighted for analysis.
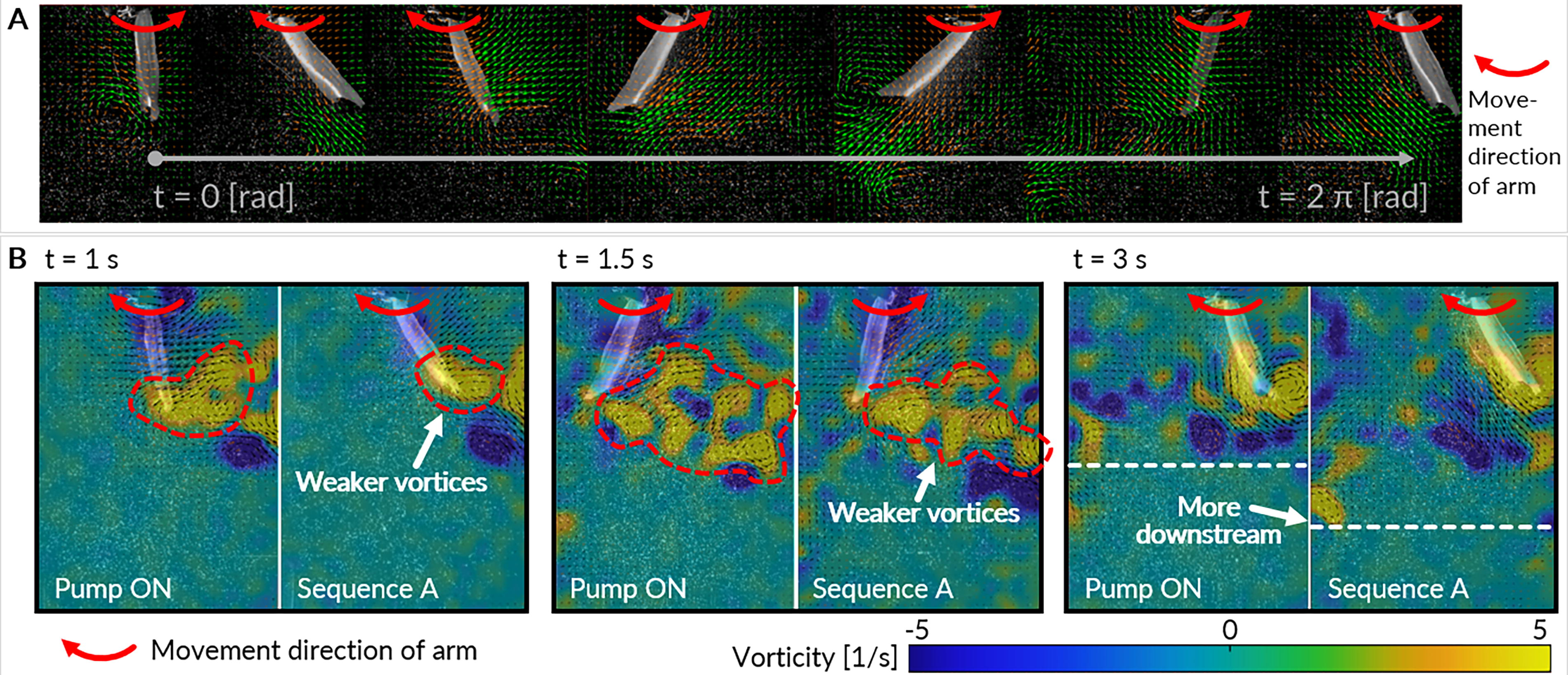
At t = 1 s, when the tail is sweeping to the left, we observe that the trailing vortices are considerably smaller for Sequence A compared to Pump ON. This observation potentially contributes to the higher average efficiency observed across different actuation frequencies. At t = 1.5 s, during a momentary pause in the tail’s motion, before it begins moving back to the right, a similar pattern emerges with smaller trailing vortices for Sequence A.
Finally, at t = 3 s, we note that for Sequence A, the vortices travel more downstream compared to Pump ON. This difference may explain why, for the amount of thrust produced to move the fin, less power is consumed. Given that vortices in swimming fish play a crucial role in propelling the fish forward, this downstream travel effect in Sequence A aligns with more efficient utilization of power. These fluid dynamics insights suggest that the reduced size and downstream travel of vortices in Sequence A contribute to its higher performance, showcasing its effectiveness in optimizing thrust and efficiency during tail actuation. Stiffness tuning within a tail-beat induces a phase offset and alters the temporal evolution of the tail shape, which can optimize force orientation to boost thrust and reduce drag. 11
Performance of robot in free-swimming experiments
Having previously evaluated the fish tail’s performance independently in a water tank, we proceed to integrate it into untethered robotic hardware for assessment in free-swimming experiments. Similar to the fish tail experiments, we evaluate performance for four pump sequences across various motor actuation frequencies. In Figure 10A, the average velocity of the robot is depicted, Figure 10B illustrates efficiency, and Figure 10C presents a combined plot of velocity and efficiency, allowing for a comparative analysis of performance with that of the tail. Results reveal that the success observed in fish tail experiments translates effectively to robotic hardware. The performance front indicates Pump OFF as optimal for lower velocities, Sequence A for middle range velocities, and Pump ON for the highest velocities. This causes a Pareto front shift (Fig. 10D) when tail stiffness is modulated within the tail-beat period compared to a binary (ON/OFF) stiffness state. At the indicated point, there is a simultaneous increase of approximately 39% in velocity and 34% in efficiency compared to the Pareto front from the binary stiffness state.

Results for the free-swimming experiment for the robotic hardware with the four sequences of pump actuation.
To validate our hypothesis about the advantage of rapidly modulating between actuation frequency and pump sequences for different robotic maneuvers, we conduct experiments (Fig. 10) in the paddling pool with the robotic hardware.
Three experiments are performed, where the lettered labels match those in Figure 10F, G:
Experiment 1: Modulating stiffness (pump sequences) starting at (a) Pump ON to (b) Sequence B to (c) Sequence A at an actuation frequency of 0.33 Hz. Experiment 2: Modulating actuation frequency starting at (d) 0.2 Hz to (e) 0.33 Hz to (f) 0.66 Hz with pump Sequence A. Experiment 3: Simultaneously modulating stiffness and frequency starting at (g) Pump OFF, 0.2 Hz to (h) Sequence A, 0.33 Hz to (i) Pump ON, 0.66 Hz.
The path of the robot and the instantaneous velocities of the three experiments in the paddling pool are shown in Figure 10F. The bias in the +X-direction results from fabrication imperfections, particularly in the buoyancy foam, but it does not affect the analysis of the tail’s performance. Each of the segments of the experiment are labeled with letters that also correspond with the data points in Figure 10G. In Experiment 1, we demonstrate the impact of altering the pump sequence on the robot’s efficiency and velocity. While a sequential pump change yields noticeable increases in both metrics, the improvements are modest. In Experiment 2, elevating tail actuation speed clearly boosts the robot’s velocity, but at the expense of reduced efficiency. Experiment 3 combines rapid stiffness and actuation frequency changes, achieving a more pronounced performance enhancement depicted as a front in Figure 10G. By being able to perform at this front of velocity and efficiency, these rapid switches in pump and actuation control can be strategically leveraged based on specific swimming objectives, whether prioritizing energy conservation or swift maneuvers. These experiments demonstrate the capability of the robot to exploit switching of both tail actuation frequency and stiffness change for enhanced performance depending on the objectives. This underscores the utility of the tail mechanism in navigating the intricate dynamics of soft robotic swimming.
Discussion
The ability to dynamically tune stiffness within a single stroke is crucial for optimizing the swimming performance of fish-like robots. Fish in nature exhibit various swimming behaviors, ranging from high-thrust rapid swimming for evasive maneuvers to energy-efficient swimming for long-distance migration. Tail stiffness plays a significant role in these behaviors, and many fish species possess the capability to adjust their tail stiffness. This adaptability allows them to respond to dynamic environmental conditions, optimize swimming efficiency, and achieve precise control during different activities. In the context of robotics, replicating such adaptability is essential for creating bio-inspired swimming robots with a broad range of functionalities. Investigating mechanisms that enable rapid stiffness modulation provides valuable insights into achieving optimal performance in underwater propulsion systems.
Our approach utilizes fluid-driven stiffening to achieve rapid and independent control of tail stiffness. This mechanism leverages the rapid change in fluid pressure within a patterned pouch, allowing for high-bandwidth adjustments in stiffness. Through experimental analysis and optimization, we demonstrate that fluid-driven stiffening significantly enhances both thrust and swimming efficiency. The success of this approach lies in its ability to mimic the natural dynamics of fish tails, enabling effective tuning of morphology for diverse swimming conditions. The utilization of fluidic-driven actuation ensures a high bandwidth, allowing substantial stiffness changes within each tail stroke.
Insights gained from our experiments include the observation of smaller trailing vortices and downstream travel of vortices in sequences with adjusted stiffness. These fluid dynamics insights contribute to the understanding of how stiffness influences thrust and efficiency at different points in the tail stroke. While our proposed mechanism shows promise, challenges remain in intuitively designing the optimal combination of stiffness, shape, and controller. Complex fluid-structure interactions and environmental considerations add to the design complexity. Hydrodynamics simulation and learning processes demand extensive data and experimentation to navigate the complex dynamics of soft robotic swimming. Future work should focus on addressing these challenges, exploring additional tail components to fully capture the complexity of fish locomotion, hardware improvements such as waterproofing the electronics and adding buoyancy control, and further optimizing stiffness modulation for improved robotic swimming performance.
The success of our fluid-driven stiffening mechanism opens avenues for extending its applicability to wider scenarios beyond robotic fish tails. The ability to dynamically adjust stiffness without altering shape offers a key feature for effective tuning of morphology in diverse swimming conditions. While the focus has been on fish-inspired robotics, the principles learned from this approach could find application in other soft robotic systems. In addition, the insights gained from fluid dynamics interactions provide a foundation for designing more efficient underwater propulsion systems. As we move forward, the challenge lies in striking an optimal balance between thrust and efficiency for swimming soft robots, and our work contributes to addressing this challenge by presenting a novel approach to tail morphology with broad potential applications.
Footnotes
Data Availability Statement
Associated data are available upon request. Please contact:
Authors’ Contributions
N.O. and J.H. conceived the study. N.O., K.J., and P.S. designed the experimental set-up. N.O. and P.S. ran the experiments. N.O. analyzed the data. N.O., K.J., P.S., and J.H. wrote the manuscript.
Author Disclosure Statement
All the authors have no conflict of interest.
Funding Information
This project was partially funded by the European Union’s Horizon 2020 research and innovation program under the Marie Skłodowska Curie grant agreement N
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.