
Abstract
It was recognized that periodontal progenitor cells penetrate disintegrated Hertwig's epithelial root sheath, and contact with root dentin give rise to periodontium formation. Clinically, direct contact of the conditioned or denuded root surfaces with periodontal cells seems to be a prerequisite for periodontal regeneration. In this study, we investigated the biological effect of dentin noncollagenous proteins (DNCPs) on the human periodontal ligament stem cells (HPDLSCs) in vitro and in vivo. Chemical-conditioned root dentin (CCRD) was prepared by process of partly demineralization and deproteinization. Treated HPDLSCs with DNCPs showed increased proliferation and adhesion ability. Induced HPDLSCs presented several features of cementoblast differentiation, as indicated by morphologic changes, enhanced alkaline phosphatase (ALP) activity, increased matrix mineralization, and upregulated expression of mineralization-associated genes. Incubation of treated HPDLSC aggregate in vivo revealed that cementum-like tissues formed along the CCRD surface with fibrous tissue adjacent to or inserted into it, but untreated HPDLSCs cannot form similar structure. To our knowledge, this is the first study to apply active proteins derived from dentin with periodontal stem cells to construct periodontal structure, which may shed light on human periodontal tissue regeneration.
Introduction
For true periodontium regeneration, direct contact of the conditioned or denuded root surface with periodontal cells seems to be a prerequisite.7,8 Clinically, disinfection and modification of the contaminated root become necessary, which include physical and chemical treatment to restore its biocompatibility and to favor the attachment of periodontal structures.9,10 After the smear layer removal, the new dentin collagenous and noncollagenous proteins are exposed, and they are supposed to be a chemoattractant for periodontal fibroblasts. DNCPs, which compromise glycoproteins
As stem cells are capable of self-renewal and multilineage differentiation, which make them very promising in regenerating organs and tissues, these cells are of paramount importance in periodontal morphogenesis.14–16 The present study was designed to investigate the biological effects of DNCPs on human periodontal ligament stem cells (HPDLSCs) in vitro, and to determine the potential of reconstructing a periodontal complex with DNCP-treated HPDLSCs on chemical-conditioned root dentin (CCRD) in vivo. For the first time, active proteins derived from dentin with periodontal stem cells were combined to construct periodontal structure, so the strategy may throw light on human periodontal tissue regeneration.
Materials and Methods
Materials
MesenPRO RS basal medium, Growth Supplement, TrypLE™ Express, fetal calf serum (FCS), and Dulbecco's modified Eagle's medium (DMEM) were from Invitrogen Corporation (San Diego, CA). 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) is from Sigma Corporation (St. Louis, MO). All other reagents were of analytical grade.
Isolation of HPDLSCs
HPDLSCs were isolated from 10 healthy donors, cultured, and expanded as previously described.6,15 Procedures were performed according to the approval of the institutional review board and the informed consent of the patients. Wisdom and premolar teeth intended for extraction due to orthodontic reasons were used as the cell source. Periodontal ligament tissue was obtained from the middle third of the tooth root and cultured in MesenPRO RS medium containing 2% FCS, 100 units/mL penicillin, and 100 μg/mL streptomycin. The explants were maintained in standard conditions until proliferating cells became confluent. Then, the digested cells with trypsin (TrypLE Express) were cultured at a low-cell density. Multiple colony–derived HPDLSCs at passage two were used in experiments.
Adipogenic/osteogenic differentiation and immunohistochemistry of the stem cells
For differentiation investigation, 1 × 104 HPDLSCs/well were plated in a 12-well dish and cultured in DMEM supplemented with 2% FCS, 100 units/mL penicillin, 100 μg/mL streptomycin, and 50 μg/mL anti–ascorbic acid (medium A) for 1 day. Then, the cells were incubated with adipogenic or osteogenic medium for another 21 days, and detected by Oil red O solution or Alizarin red staining, respectively. 16 The Oil red O–positive cells were counted in multiple rectangular frame under a phase-contrast microscope (Olympus IX51, Tokyo, Japan), and the percentage to the total cell number was calculated.
Isolated putative stem cells were seeded on six-chamber slides and cultured in MesenPRO RS medium for 24 h. After being washed in phosphate-buffered saline (PBS, pH 7.4) and fixed in 4% paraformaldehyde for 15 min, the samples were incubated with STRO-1 antibody (1:200; R&D Systems Inc., Minneapolis, MN) for 2 h, and subsequently incubated with fluorescein isothiocyanate (FITC)-conjugated anti-mouse secondary antibody. The surface marker STRO-1–positive cells were also analyzed by flow cytometry.
Preparation of CCRD
The teeth for culturing HPDLSCs were prepared for CCRD. Under continuous water coolant, the roots were treated to eliminate all adhering soft tissues (pulp and periodontal ligament) and cementum with high-speed diamond burs. Then, roots were sliced in longitudinal section, and the coronal portion was discarded. Total of 24 specimens approximately measuring 0.5–1.0 mm × 4 mm × 5 mm were obtained. A series of physical and chemical procedures were performed according to reported protocols with minor modifications.7,17,18 In brief, dentinal materials were treated for 3 min with chloroform–methanol (1:1), 0.6 M hydrochloric acid for 5 min, 0.5 M EDTA for 3 min, 2 M calcium chloride for 3 min, lithium chloride for 5 min, and absolute ethanol for 10 min at room temperature. Then, samples were ultrasonically cleaned in sterile deionized water for three times (5 min for each time). Thereafter, the freeze-dried process was performed (ALPHA 2-4 LSC; CHRIST, Osterode, Germany). At last, all samples were incubated in PBS buffer containing 1000 units/mL penicillin and 1000 μg/mL streptomycin overnight.
Assessment of the effect of DNCPs on HPDLSCs
Cell growth rate assay, flow cytometry, and bromodeoxyuridine incorporation
To evaluate the proliferation potential and viability of the HPDLSCs treated or untreated with DNCPs, growth rate MTT assay, cell cycle analysis, and bromodeoxyuridine (BrdU) incorporation into DNA were conducted. In the present study, DNCPs (kindly provided by Professor A.J. Smith) were dissolved in 5 mM acetic acid solution to enhance solubility instead of conventional dissolution methods.11,12 Two groups were set: in the test group, 250 ng/mL DNCPs was added to medium A (medium B), and the control group contains only the medium A. Cells were plated at a density of 5 × 103 cells/well in 96-well plates and cultured in medium A or medium B, and the MTT assay was carried out for 5 days according to the cell proliferation kit protocol (Sigma). Absorbance was determined at 490 nm with a microplate reader (Bio.TEK Instruments, Winooski, VT). All reported values are the means of triplicate samples. This test was repeated twice.
About 2 × 105 single-cell suspensions were maintained in medium A or medium B and cultured in 75 cm2 flask; at day 5, flow cytometry analysis was respectively treated as previous report. 19 The fractions of cells in the G1, S, and G2 phases of cell cycle were analyzed.
DNA synthesis of the HPDLSCs in different medium was assessed by measuring BrdU incorporation. HPDLSCs were maintained in MesenPRO RS medium. Following 24 h of serum starvation, cells were exposed to medium A or medium B for 24 h. BrdU (10 mM) was added 4 h before the end of incubation. Immunodetection of the incorporated BrdU into cells was performed as described previously. 20 The BrdU-labeling index was determined as the percentage of BrdU-positive cells to the total cell number.
Clonogenic assays and cell morphology observation
To assess colony-forming efficiency, single-cell suspensions within medium A or medium B (1 × 103 cells) were seeded into six-well dishes. Day-7 cultures were fixed with 4% paraformaldehyde, and stained with 0.1% toluidine blue. Aggregates of over 50 cells were scored as colonies; results were recorded and compared between two groups. Cell morphology was examined under phase-contrast microscope and scanning electron microscope (SEM, S-3400N, 5.0 KV; Hitachi, Tokyo, Japan).
Cell adhesion assay
A cell adhesion assay was performed using the methods of Rodrigues et al. 20 Briefly, 24-well culture plates were incubated with 1 mL of PBS containing 250 ng/mL DNCPs for 16 h at 4°C, and were subsequently washed three times with PBS. Nonspecific binding sites were blocked with 3% bovine serum albumin (Sigma). Five-hundred-microliter cell suspension (5 × 104/mL) in medium A was plated into uncoated plates, meanwhile 500-μL cell suspension (5 ×104/mL) in medium B was plated into coated plates. Incubation was performed for 4 h at 37°C in 5% CO2. Following incubation, wells were gently rinsed with 1 mL PBS to remove unattached cells. Attached cells were harvested with trypsin, and counted in a hemocytometer. Cells suspended in serum-free medium A and seeded in uncoated plates were used as negative control. Data are expressed as the percentage of attached cells compared with negative control.
Analysis of alkaline phosphatase activity, quantitative real-time PCR, and mineralization
To investigate the potential of HPDLSCs treated with DNCPs to differentiate into mineralizing cementoblast-like lineages in vitro, the alkaline phosphatase (ALP) activity, real-time RT-PCR, and mineralization behavior were analyzed. HPDLSCs were seeded into 24-well plates (5 × 103 cells/well) and cultured under medium A or medium B. We have assessed the different concentrations of DNCPs on HPDLSC differentiation by determining the level of ALP activity according to the description in detail by Zhang et al. 21 The quantitative ALP activity was measured at 1, 7, and 14 days. Four samples were taken at each measurement. The amount of p-nitrophenol released by the enzyme reaction was measured at 405 nm using a Microplate Reader (Bio.TEK Instruments). ALP activity was always expressed relative to the amount of DNA with PicoGreen dsDNA quantitation kit (Molecular Probes, Eugene, OR).
The mRNA expressions of several osteoblastic markers, including type I collagen (col1), alkaline phosphatase (ALP), and osteocalcin (OCN), were evaluated by real-time quantitative RT-PCR analysis following stimulation by DNCPs. Total RNA was extracted from the HPDLSCs after incubation in medium A or medium B for 24 and 48 h using Trizol Reagent (Invitrogen, Paisley, UK). cDNA of 2.5 μL was amplified in 25 μL volume containing 12.5 μL of SYBR Green Realtime PCR Master Mix (Toyobo, Osaka, Japan) and 0.2 μM of each primer. Real-time PCR was carried out on a Chromo 4™ Continuous Fluorescence Detector and analyzed using Opticon Monitor™ software (MJ Research, Waltham, MA). The PCR primer sequences listed in Table 1 were designed according to Nagai et al. 22 and were found unique after “blastn” (http://www.ncbi.nlm.nih.gov/BLAST/). Moreover, the unique amplified products were confirmed on 3% agarose gel. The melting curve was constructed for each primer to ensure reaction specificity. Following PCR, the threshold cycle was obtained and relative quantities determined by comparison with glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Triplicate reaction and three separate experiments were performed.
Density of 1 × 104 cells/well in 24-well dishes was maintained in medium A or medium B and supplemented with osteogenetic media (10 mM β-glycerophosphate and 10 nM dexamethasone) for 21 days. Mineralized formation was identified by Alizarin red staining. Amount of Alizarin red bound to the mineral in each dish was quantified according to Jo et al. 16
In vitro cell attachment to CCRD
Prepared and sterilized CCRD was soaked in medium A or medium B for 24 h. HPDLSCs maintained in medium A or medium B for 3 days were harvested by trypsinization, and cell density was adjusted to 1 × 105 cells/mL. Fifty microliters of cell suspension was seeded on each CCRD surface (cells cultured in medium A were seeded on medium A–treated CCRD, and cells cultured in medium B were seeded on medium B–treated CCRD, respectively). The combinations were located into 24-well dishes and incubated in standard conditions for 3 h, and then 500 μL respective medium was supplemented and incubated for another 24 h. The morphologies of the CCRD and CCRD–cell composite were examined under SEM.
Statistical analysis
Data were analyzed using SPSS version 10.0 (Chicago, IL). Statistical analysis of the data was performed by Student's t-test. For all tests, significance level was set at p < 0.05.
In vivo study
About 1 × 106 HPDLSCs in 75 cm2 flasks were cultured for 7 days in medium A or medium B, and have grown into multiple layers that contained plenty of extracellular matrix (ECM). High density of HPDLSCs and ECM were harvested by TrypLE Express and centrifuged at 150 g for 6 min in order to get cell aggregate. Thereafter, cell aggregate containing approximately 1 × 107 cells was seeded on each CCRD, which having infiltrated with medium A or medium B (cells were seeded by the same methods as described in “In vitro cell attachment to CCRD” section) in 24-well culture plates. Following in vitro incubation for 24 h in standard conditions, all the CCRD–cell composites (six DNCP-treated composites and six DNCP untreated composites) were implanted into dorsal subcutaneous area of athymic mice (BALB/c-nu; FMMU Medical Laboratory Animal Center, Xi'an, China). CCRD containing no cells treated or untreated with DNCPs was also implanted into dorsal subcutaneous area of athymic mice as negative control. All animal experimental procedures were carried out in accordance with the Guidelines and Regulations for the Use and Care of Animals of the Review Board of Fourth Military Medical University Medical Laboratory Animal Center. The specimens were harvested at 4 weeks posttransplantation and stained with hematoxylin and eosin.
Results
Characteristics of HPDLSCs
We isolated postnatal stem cells from periodontium, and the isolated cells formed single-cell-derived colonies and most of the cells retained their fibroblastic spindle shape. Ex vivo–expanded HPDLSCs expressed the cell surface molecules STRO-1 by immunohistochemical staining, which is early mesenchymal stem cell marker (Fig. 1A). A total of 29.01% of the periodontal cell population was positive for STRO-1 expression by flow cytometric analysis (Fig. 1B). After 3 weeks of culture with an adipogenic inductive cocktail, HPDLSCs developed into Oil red O–positive lipid fat cells (Fig. 1C). A total of 8.3% of the PDL cell population was positive for Oil red O staining. Small round Alizarin red–positive nodules formed in the HPDLSC cultures after 3 weeks of osteoblastic induction, indicating calcium accumulation in vitro (Fig. 1D).
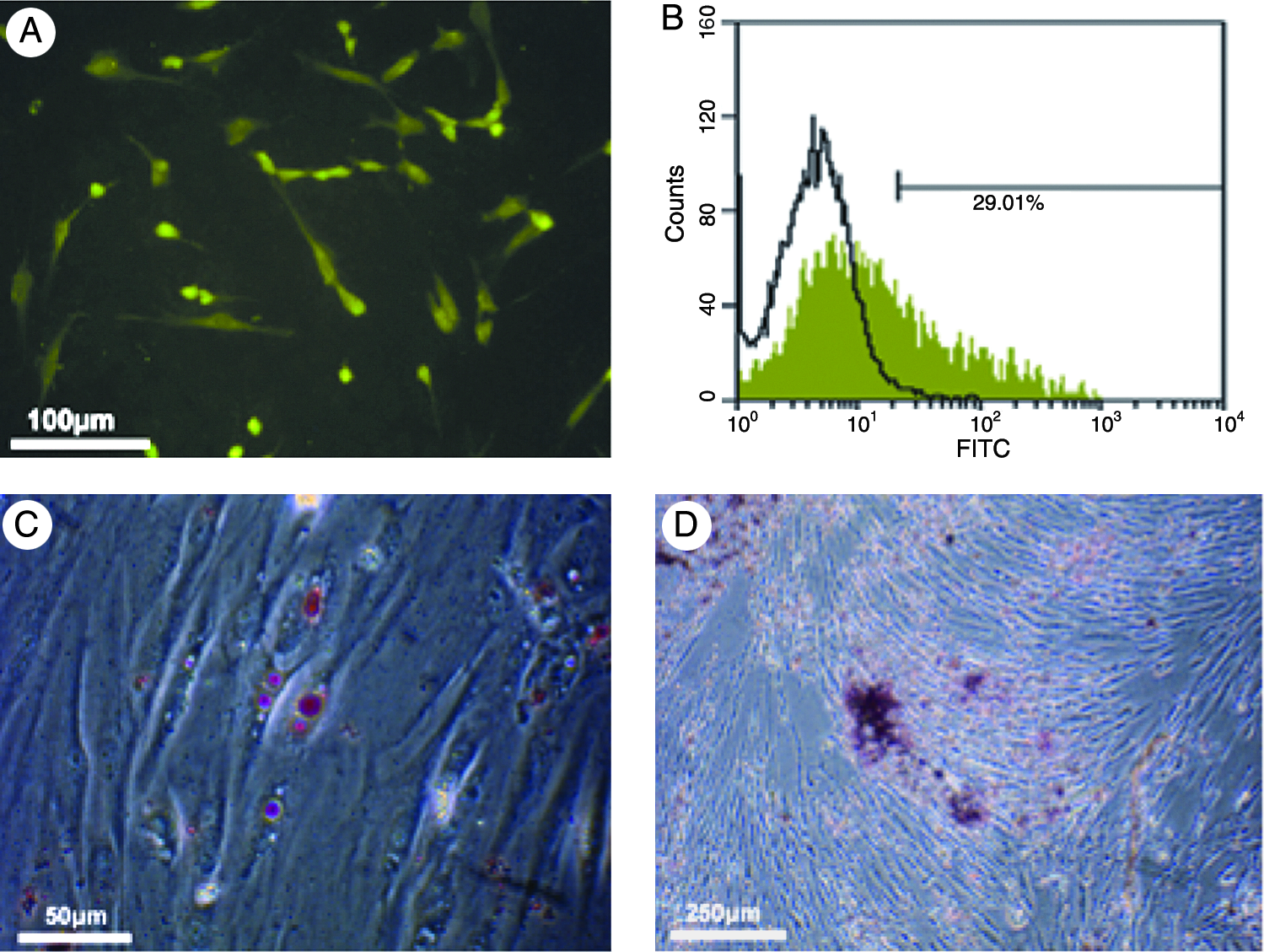
Characteristics and multilineage differentiation of HPDLSCs. (
SEM observation of CCRD
When the specimens were examined under SEM, CCRD demonstrated clean surface, which was eliminated of smear layer and had opened and widened dentin tubules (Fig. 2A, B). Cells showing well growth were seen on all specimens. In dense cell area, cell sheets were formed and covered dentin surface (Fig. 2C). In sparse cell area, DNCP treated cells showed polygonal or cubical shape instead of spindle shape (Fig. 2D).

Scanning electron micrographs of CCRD and cell spreading. (
Effect of DNCPs on HPDLSCs
MTT assays showed that cells cultured with DNCPs demonstrated a statistically significant increase in proliferation at days 3 and 5 as compared to control group (Fig. 3A) (p < 0.05). In addition, the former showed a significantly higher rate of BrdU uptake than the later (Fig. 3B). The higher proliferation activity of induced HPDLSCs was further confirmed by flow cytometry. A higher percentage of cells in S (17.0%) and G2 (9.2%) phases and a lower percentage in G1 phase (73.8%) were detected in DNCP-stimulated cells (Fig. 3C) compared with noninduced cells (Fig. 3D) (p < 0.05).
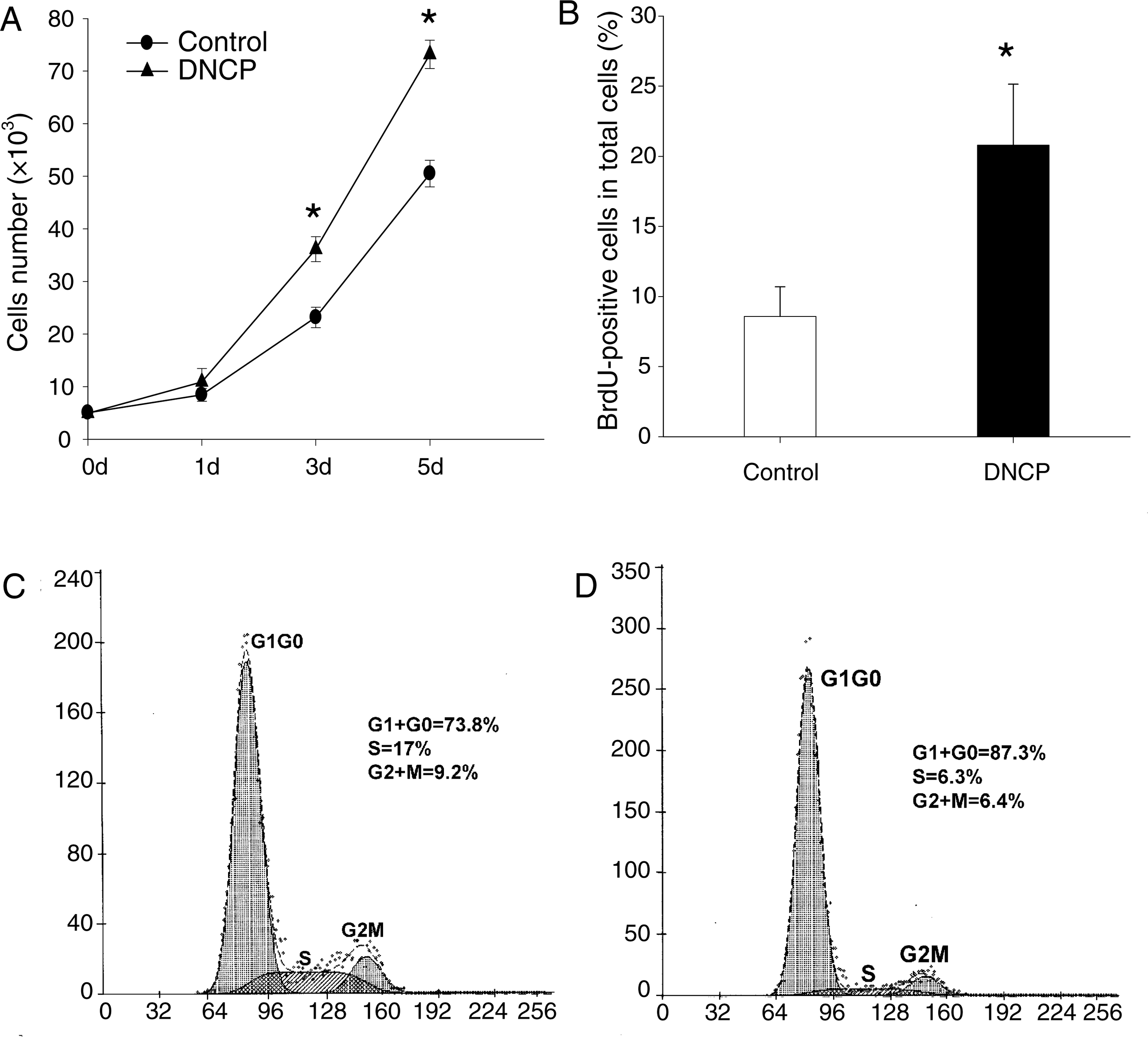
Effect of DNCPs on proliferation of HPDLSCs. (
Both groups of HPDLSCs showed the ability to form adherent clonogenic cell clusters (Fig. 4A, B). HPDLSCs treated with DNCPs formed 51 colonies, whereas untreated cells generated 49 colonies. These results showed no statistically significant difference between the two groups (Fig. 4G). When observed under phase-contrast microscope, untreated cells were fibroblastic and bipolar in shape (Fig. 4A). While treated cells became flatter and most of them were cuboidal or polygonal after the induction of DNCPs for 7 days (Fig. 4B). The SEM results were coincident with the above light microscope findings, showing that most of the untreated HPDLSCs were elongated, spindle shaped, and displayed abundant microvilli on the cell surface (Fig. 4C), while the treated cells closely adherent to the substrate appeared cuboidal or polygonal shaped with multiprocesses (Fig. 4D). In addition, compared with the control, mineralized secretary matrix granules can be found on the surface of the treated cells.
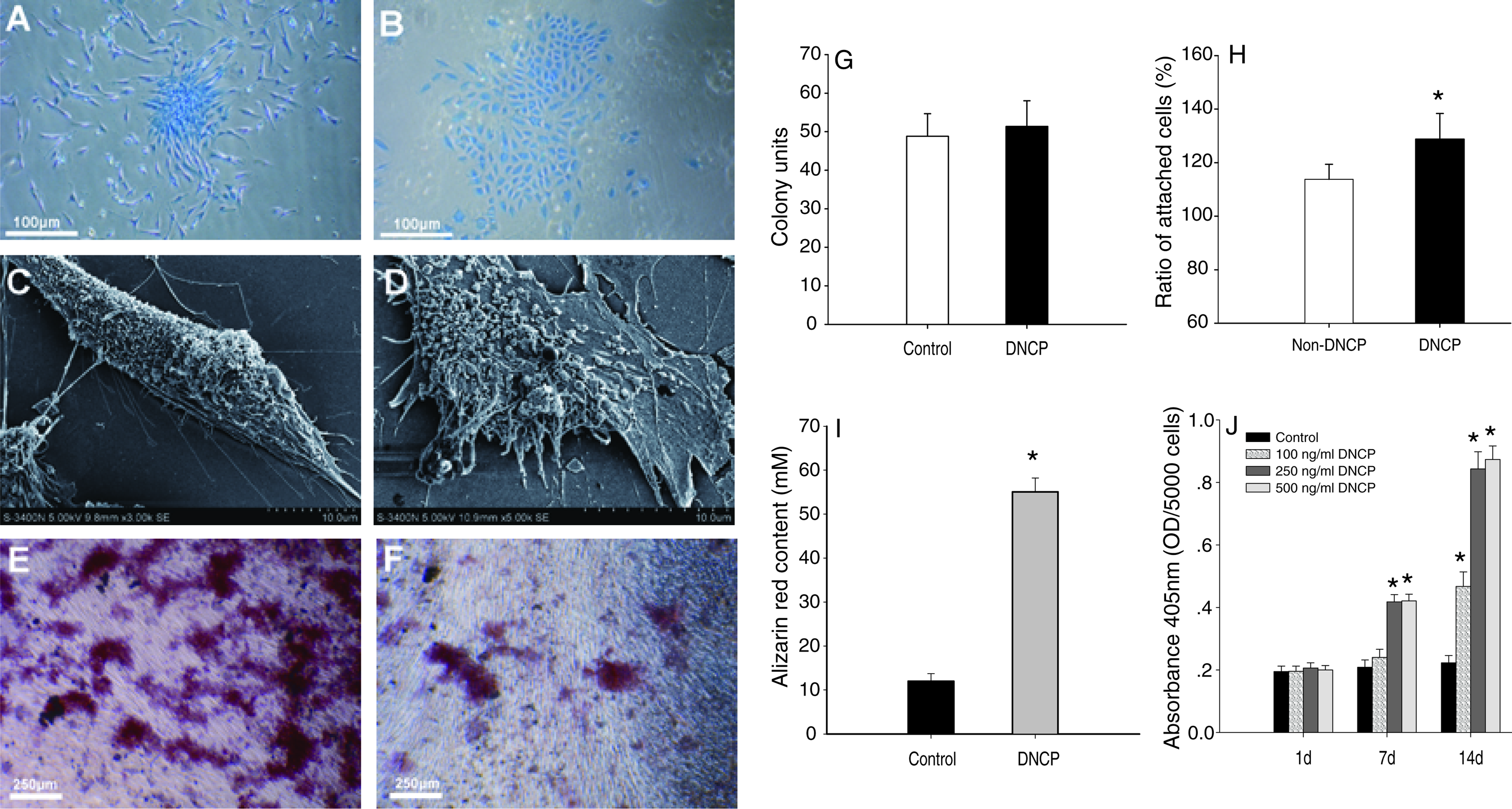
Effect of DNCPs on differentiation of HPDLSCs. (
Cell adhesion assay showed that both groups have proper adhesive ability. But increased cell attachment was presented statistically in the group whose plates and cells were incubated with DNCPs (Fig. 4H; p < 0.05).
ALP activity of HPDLSCs in response to different DNCP concentrations was increased as indicated in Figure 4J. The ALP activity of day 1 referred only to basal level in HPDLSCs. ALP activity of HPDLSCs was obviously higher under the induction of DNCPs with respect to noninduced group at 7 and 14 days (p < 0.05), but there was no significant difference between 250 and 500 ng/mL groups (Fig. 4J).
After incubation in mineralized culture for 21 days, both groups showed calcium deposits as visualized by Alizarin red staining (Fig. 4E, F). HPDLSCs cultured in the presence of DNCPs produced extensive sheets of calcified deposits (Fig. 4E), whereas deposits of the HPDLSCs in the absence of DNCPs were sparsely scattered (Fig. 4F). Quantification of the amount of Alizarin red also showed significant difference (Fig. 4I).
The stimulatory effect of DNCPs on ALP, type I collagen, and OCN mRNA expression was detected. Relative intensities compared to GAPDH are shown in Figure 5. The mRNA expressions of type I collagen, ALP, and OCN in the presence of DNCPs caused a 1.7-, 2.2-, and 1.2-fold increase compared with those in the absence of DNCPs, respectively, for 2-day incubation. type I collagen and ALP caused 1.2-fold and 1.4-fold increase in mRNA levels after incubated for 1 day. There was no significant difference in the expression level of OCN between two groups during the 1-day culture period (Fig. 5A, B, and C).
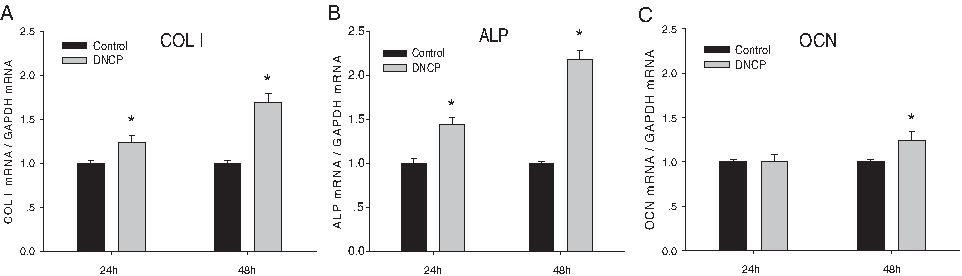
Effect of DNCPs on gene expressions of HPDLSCs by quantitative real-time RT-PCR analysis. Stimulatory effect of DNCPs on COL I, ALP, and OCN mRNA expression was detected. The graphs show the ratio of target genes to GAPDH mRNA. Values are expressed as mean ± SD (*p < 0.05).
Historical observations of tissue samples in vivo
All mice appeared to be in good health throughout the test period, and there were no symptoms of severe inflammation at the implantation sites. In two out of six DNCP-treated specimens for 4-week transplantation, new cementum-like tissue formed along the CCRD surface with cell-rich fibrous tissue adjacent to or inserted into it (Fig. 6A, B). In three out of six DNCP-treated specimens, there are obvious separations between neoformed tissues and dentin surface, only showing monostratified or stratified cubical cementoblast-like cells regularly aligned on dentin surface (Fig. 6C, D). While in all the DNCP untreated group and one DNCP-treated specimen for 4 weeks posttransplantation, no obvious cementum-like structure or periodontal fibrous tissue formed along the CCRD surface, and there were broad gaps between hard dentin surfaces and soft tissues (Fig. 6E, F). In the negative control of naked CCRD transplantation, there was no host cell attachment or tissue regeneration on all CCRD surfaces (Fig. 6G, H).
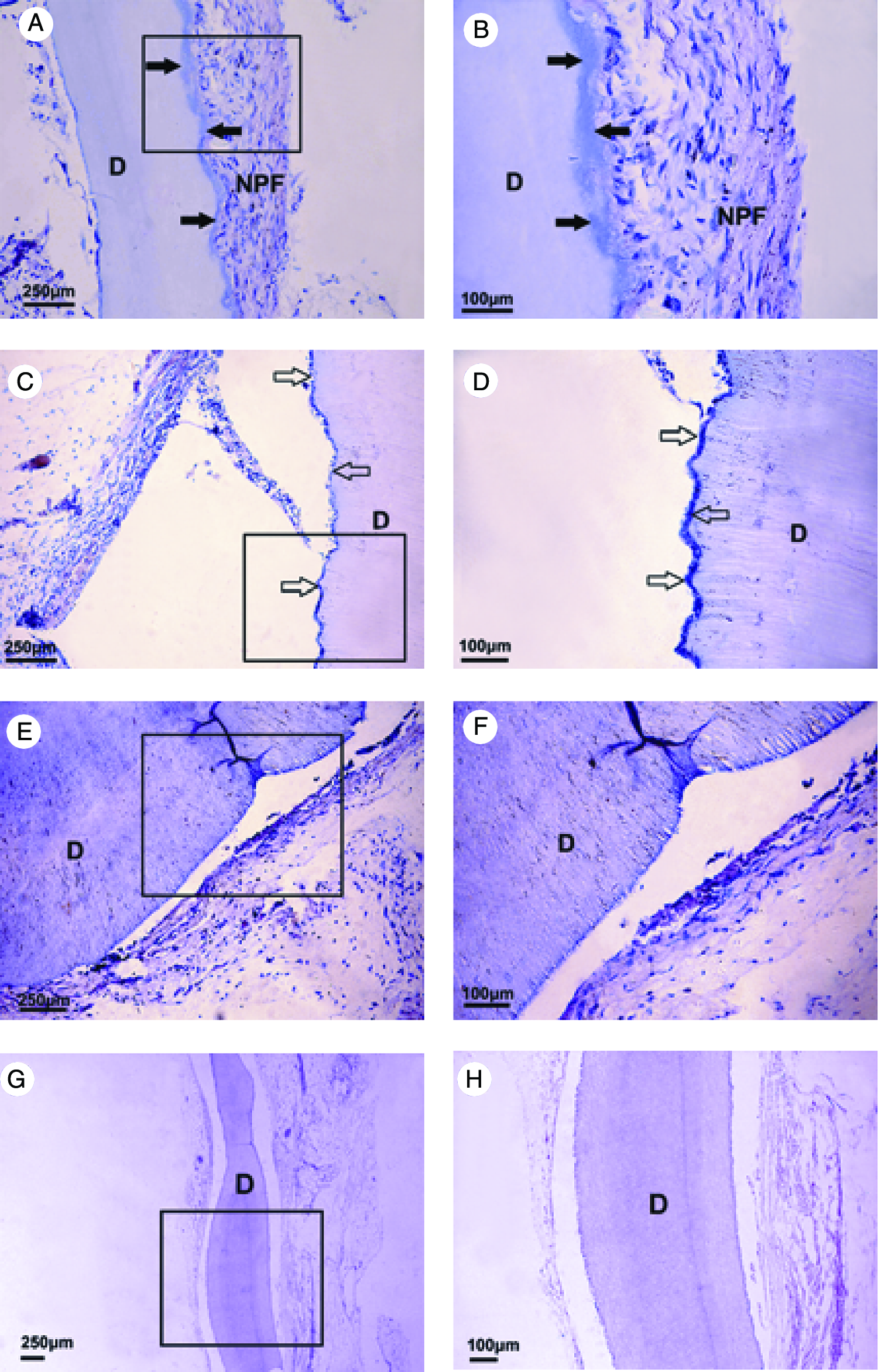
Histological observation 4 weeks posttransplantation (hematoxylin-eosin staining). (
Discussion
Periodontal regeneration is dependent on a sequence of associated events, including cellular proliferation, migration, differentiation, and attachment to components of the ECM as well as organic matrix synthesis and mineralization.23,24 Natural dentin contains numerous collagenous protein and noncollagenous signaling molecular protein, sequestered in mineralized matrix, which includes bone morphogenetic proteins (BMPs), transforming growth factor beta (TGF-β), dentin sialoprotein (DSP), dentin phosphoprotein (DPP), dentin matrix protein 1 (DMP-1), and others.11,25 Accordingly, we hypothesized that dentin molecules bound to matrix components or to hydroxyapatite crystals may be exposed or released as a consequence of injury to the periodontal ligament. It has been suggested that these numerous signals can not only get involved in the nucleation process and hydroxyapatite growth during dentin formation, but also induce greater amounts of mineralized bone and superior cementogenesis in periodontal regeneration and embryonic development.26,27
Previous experimental studies have highlighted the interactions between dentin and surrounding cells. And meanwhile these studies revealed that surface of dentin is a substratum for cell adhesion, dentin molecules might function as regulatory signals for various mesenchymal and inflammatory cells to enhance the healing of periodontal tissues.12,13,28,29 It has been generally accepted that the disruption of HERS is the key event at the onset of root development.1,2 Thus, periodontal progenitor cells have the access to root dentin surface, and they have great possibility to undergo cementoblastic differentiation and cementum formation by dentinal induction. 4 We hypothesized that dentin matrix produced by odontoblasts could migrate through and diffuse to periodontal tissues, which is similar to the well-known phenomenon of diffusion of enamel matrix proteins through the premineralizing mantle dentin into the odontoblast layer. 30 Therefore, these molecules can be expected to act in conjunction with other factors to promote cementogenesis by affecting periodontal cells just in the same way as they are in enamel matrix derivatives (EMD). Based on previous studies and theory of root onset, we managed to determine whether the DNCPs derived from dentin provided additional advantages for the cellular events associated with periodontal regeneration. The present study indicated that DNCPs can promote proliferation potential and viability of HPDLSCs. In addition, induced HPDLSCs presented several features of cementoblast differentiation, as indicated by morphologic changes, enhanced ALP activity, increased matrix mineralization, and upregulated expressions of mineralization-associated genes, such as type I collagen, OCN, and ALP. Further, our adhesion assay showed that DNCPs can slightly promote HPDLSC attachment ability, compared with control treatment. It is suggested that although the underlying mechanisms are still not well understood, cemento/osteoblastic differentiation of HPDLSCs may be affected by the dose of DNCPs according to our results.
As to CCRD, several in vitro studies have suggested that root surface modification by different ways could influence the behavior of periodontal fibroblasts, improving their attachment, proliferation, and development.7,10,31,32 Clinically, organic or inorganic acid is frequently used as root conditioning in order to demineralize and deproteinize. Because of their acid pH, they may partly demineralize the planed root surfaces, minimize detrimental antigenicity, eliminate the smear layer, and expose some components of the ECM of dentin or cementum, such as proteoglycans, fibronectin, and signaling growth factors.10,33 In accordance with previous studies, our SEM observations of CCRD from partly demineralization and deproteinization demonstrated elimination of smear layer, and opening and widening of the tubules. Cell spreading was already evident, and the penetration of cytoplasmic process into dentinal tubules was frequently observed. HPDLSCs were attached to dentin surfaces, and formed cell junctions. The results indicated that conditioned dentin by this treatment presents prospective substratum for HPDLSC colonization, growth, and spread.
In order to fabricate new periodontal tissues to replace damaged tissues derived from periodontitis or reconstruct periodontium relationship between bioimplant root and adjacent alveolar, a cell-based tissue engineering technique has been proved to be one of the most promising alternative therapies for periodontal tissue regeneration. So far, many studies have demonstrated that stem cells exhibit astonishing capacities for regenerative therapy.14,16,34 Consistent with previous studies, from tissue of periodontium, we isolated and characterized stem cells that have the potential to differentiate into multiple cell lineages, such as adipocytes or cemento/osteoblasts. Further, in the present study, we found enhanced proliferation and differentiation ability of HPDLSCs induced by DNCPs and sustained by CCRD.
To validate whether induced HPDLSCs had the potential to regenerate the cementum or periodontium complex, 12 samples of ex vivo–expanded HPDLSC aggregates treated or untreated by DNCPs combining with CCRD were transplanted into immunocompromised mice. Four weeks posttransplantation, histological findings demonstrated that new cementum-like tissue formed on CCRD surfaces after the application with aggregate of induced HPDLSCs. In light of these in vivo evidence, we testified again that DNCPs have provided additional advantages for the cellular events associated with periodontal regeneration. We speculated that CCRD acted as physical-supporting environment, and DNCPs play an important role of tissue formation resulted from cemento/osteoblastic differentiation of undifferentiated mesenchymal cells.
In our study, some experimental samples from posttransplantation showed neoformed tissues separated from CCRD surfaces. The phenomenon may be illustrated by the following two reasons. The first is comparatively physical smooth feature of the CCRD surface. Although treated by the process of partly demineralization and deproteinization, CCRD still presents large-area smooth surface and only provides few dentinal tubules to permit penetration of cytoplasmic processes. Therefore, the smooth surface characteristics may depress the capability of adhesion, growth, and tissue neoformation arising from the stem cell. Just as surface roughness modification of present titanium-implant body to benefit osteogenetic cell contact and growth, 35 we also need to modify dentin surface and promote roughness to facilitate colonization and proliferation of HPDLSCs. The second reason may lie in the present limited understanding of process about periodontal occurrence and regeneration, which results in a situation where we cannot completely reproduce physiological periodontium only using monotonous growth factors or scaffolds with available methods now. As is well known, process of periodontium occurrence and regeneration includes interaction between mesenchymal and epithelial cells, special microenvironment including effect of ECM, and regulation of local or remote chemotactic growth factors as temporally and spatially connected events. For example, EMD produced by HERS are known to play an important role in cementogenesis and the periodontal attachment apparatus.3,4 In addition, platelet-rich plasma (PRP) that was derived from peripheral blood and involved in wound healing and repair of mineralized tissue was also confirmed to participate in periodontal development and regeneration. 36 Previous studies and our present research demonstrated that dentin-derived matrix can also affect the biological behaviors of periodontal cells. Therefore, the cocktail effect and combination of different growth factors with versatile scaffolds simulating ECM may help to regenerate preferable periodontium. Although the effective components of DNCPs should be analyzed in the future, this study affirmatively provides a better understanding of the underlying mechanisms of cementogenesis, which may be helpful for regenerative periodontal therapies. Next, we are going to identify the key factors in DNCPs and further explore the network of these factors on the differentiation of HPDLSCs.
In conclusion, our study has demonstrated that DNCP is an effective stimulator of HPDLSC proliferation, morphological changes, attachment, and cemento/osteoblastic differentiation. DNCPs facilitate HPDLSCs in forming cementum-like adjacent fibrous tissue on CCRD. These findings therefore provide convincing evidence and useful data for DNCPs as a potent tool to facilitate periodontal regeneration. Besides, DNCPs have rich source, for example, from human wisdom and premolar teeth intended for extraction due to orthodontic reasons or animal developmental tooth dentin. Compared with other matrix proteins available in market (e.g., EMD or PRP), DNCPs may have advantage in supply of source. But, the respective effect manner and relationship of different matrix proteins should be compared and investigated in our future studies.
Footnotes
Acknowledgments
We are grateful to Prof. Anthony J. Smith (Oral Biology, School of Dentistry, University of Birmingham, Birmingham, UK) for kindly providing DNCPs. This work was supported by grants from the Nature Science Foundation of China (Project No. 30572046 and No. 30725042).
Disclosure Statement
No competing financial interests exist.