
Abstract
We previously demonstrated the ability to create engineered arteries by carefully controlling the mechanical environment of intact arteries perfused ex vivo, yielding engineered arteries with native appearance and vasoactive response. Increased axial strain was sufficient to increase length up to 20% in 9 days through a growth and remodeling response. The amount of the achievable length increase, however, was highly dependent on the hemodynamic conditions acting through unknown mechanisms. Because matrix metalloproteinase-2 (MMP-2) and matrix metalloproteinase-9 (MMP-9) activity is increased, and often required, in mechanically induced remodeling in vivo, MMP-2 and MMP-9 expression was investigated to elucidate the hemodynamic mediation of artery length. Carotid arteries from 30 kg pigs were perfused for 9 days ex vivo at either in situ axial strain or with a gradual 50% increase in axial strain, under either arterial or reduced hemodynamics (∼10% of arterial hemodynamics). MMP-2 protein expression increased roughly twofold, while MMP-9 expression increased threefold under either reduced hemodynamics or increased axial strain (p < 0.05). The combination of reduced hemodynamics with increased axial strain demonstrated an additive increase in MMP-9 protein (p < 0.05) with no further change in MMP-2 expression. To investigate the mechanism by which axial strain and hemodynamics could additively increase MMP-9 expression, the expression of nuclear factor kappa B (NF-κB) subunits p50 and p65 was evaluated. Axial strain stimulated p65 expression and localization, while hemodynamics increased p50 expression, with both molecules being expressed only when both mechanical stimuli were applied. These data suggest that MMP-9 expression can be simultaneously stimulated by separate mechanical stimuli mediated by p50 and p65 expression, and that by using conditions that maximize MMP-9 expression, we can create an optimal remodeling environment to better direct the growth of engineered arteries and other tissues.
Introduction
An alternative approach explored by our group and others is to harvest a relatively small artery and direct its remodeling ex vivo as an intact vessel.15–18 The distinct advantage of this technique is the maintenance of native arterial structure, mechanical properties, and vasoactive function, avoiding many of the most difficult aspects of traditional vascular tissue engineering. However, this technique is not without substantial challenges, as current techniques have achieved only modest growth in relatively short time periods. We previously demonstrated that axial remodeling to increase length could be achieved in this system by gradually increasing the axial strain by 50% over 9 days.15,17 However, the achievable length increase was significantly increased when reduced hemodynamics was combined with increased axial strain, demonstrating for the first time that hemodynamics have a modulatory effect on axial remodeling. By elucidating the underlying mechanisms by which hemodynamic conditions modulate axial remodeling, we could potentially exploit these mechanisms to increase the efficacy of using directed remodeling to create engineered arteries.
Matrix metalloproteinases (MMPs) are produced and activated to remodel the existing ECM and incorporate new components in arterial remodeling. 19 Specifically, increased MMP-2 and MMP-9 expression has been demonstrated in response to altered blood flow20,21 and hypertension, 22 while altered axial strain caused sustained MMP-2 expression but only transient MMP-9 expression23,24 in arteries in vivo. Altered shear stress also produces soluble factors such as nitric oxide (NO) 25 that lead to MMP production and release in an NF-kB (p50/p65 subunit)–dependent response. 26 The response to altered shear stress is abolished when MMPs are blocked using pharmacological intervention,27,28 or the endothelial cells are removed. 29 However, endothelial denudation does not attenuate remodeling due to increased axial strain, 23 suggesting that there may be multiple and independent pathways to induce MMP expression in arteries. Mechanical strain has been shown to directly stimulate smooth muscle cells to produce MMP-2 and MMP-9 in vitro in monolayer studies 30 independent of functional endothelium. This demonstrates that there are at least two stimuli for MMP-mediated remodeling in arteries—direct mechanical stimulus of smooth muscle cells and endothelial signaling to smooth muscle cells—that could be simultaneously exploited to increase remodeling in engineered arteries.
We hypothesized that the additive remodeling seen in engineered arteries caused by concurrent alteration of axial strain and hemodynamics could be due to independent stimulation of MMP-2 and MMP-9 expression via direct and indirect pathways in smooth muscle cells. The data presented give insight into the mechanisms regulating remodeling in engineered arteries, and suggest that controlling MMP activity through mechanical or other stimulation could better control remodeling in engineered arteries and other tissues.
Materials and Methods
Ex vivo perfusion and elongation
The investigation conforms with the Guide for the Care and Use of Laboratory Animals published by the U.S. National Institutes of Health (NIH Publication No. 85-23, revised 1996), and all protocols were approved by IACUC. The ex vivo perfusion system and isolation procedures have been described in detail elsewhere.15,17 Briefly, carotid arteries were harvested from anesthetized 30 kg pigs, cleaned of excess connective tissue, and briefly pressurized to inspect for leaks or unligated branches. Arteries were cannulated at their in situ length onto 10 gauge stainless steel rods via silk sutures within a custom-built acrylic artery chamber. The artery chamber was then connected to a self-contained perfusion loop with a flow rate of 15 mL/min and 10 mmHg, and either the chamber was maintained at this level (reduced hemodynamics) or the flow rate and pressure were increased over 2–3 h until arterial hemodynamics were achieved (150 mL/min, 80/120 mmHg, arterial hemodynamics). For each hemodynamic condition, five to seven arteries were either maintained at their in situ length for 9 days or elongated 50% of their in vivo length in 9 days, by increasing the total length by 8.3% per day on days 2–7. We previously reported on the growth, remodeling, cellular proliferation, vasoactivity, and ECM composition for arteries cultured in this perfusion system. As previously seen in cultured arteries,15,17 cell death measured by TUNEL staining was minimal in all cases, while viability assessed by MTT assay displayed similar mitochondrial activity to fresh tissue. Select cultured arteries were also tested for vasoactive response to norepinephrine and sodium nitroprusside and demonstrated strong responses to both agonists similar to previous reported results.15,17 These data combined demonstrated maintenance of cell and tissue viability similar to previous reports.15,17
Protein isolation
Whole segments of arteries weighing 30–50 mg were flash frozen in liquid nitrogen immediately after harvest or ex vivo culture and stored at −80°C. Frozen tissue was hydrated with protein extraction buffer 31 and immediately homogenized using a rotor–stator homogenizer. The tissue homogenates were centrifuged, the supernatant was collected, and protein concentration was determined using the bicinchonic acid (BCA) assay according to manufacturer's directions (Thermo-Fisher, Rockford, IL). Protein was then aliquoted and frozen at −80°C to prevent excessive freeze-thaw cycles.
Protein analysis
To assess the activity and quantity of MMP-2 and MMP-9, gelatin zymography was performed as described previously elsewhere.20,22,32 Equal amounts of protein (10 μg) were run on each lane of 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gels containing 1% gelatin (Biorad, Hercules, CA), along with human MMP-2 and MMP-9 standards (Chemicon, Temecula, CA). Bands were imaged and quantified using the Biorad's Quantity One software, with all values normalized by the average value of the proMMP-2 band from fresh tissue. This allowed for direct comparison of expression levels between MMP-2 and MMP-9. Data were quantified for the proMMP-2, MMP-2, and proMMP-9 bands from the zymograms, and total MMP-2 refers to the sum of the expression levels for proMMP-2 and (active) MMP-2. To verify that the bands corresponded to MMP activity, identical control gels were developed in the presence of the MMP inhibitor Marimastat (Chemicon), which produced gels with no detectible degradation bands (data not shown).
To confirm that the degradative bands seen in the gelatin zymography corresponded with MMP-2 and MMP-9, Western blot analysis was performed. Briefly, equal amounts of protein (10 μg) were run on each lane of 10% SDS-PAGE gels (Biorad), along with MMP-2 and MMP-9 standards and a molecular weight ladder (Chemicon). The proteins were then transferred to nitrocellulose membranes, blocked, and stained with MMP-2 or MMP-9 antibodies according to manufacturer's instructions (Chemicon).
Immunohistochemical evaluation
Segments of arteries 2–3 mm in length were fixed in 10% formalin overnight, embedded in paraffin, cut into 5-μm-thick sections, and placed onto slides, as previously described.15,17 Sections from all conditions were stained with antibodies recognizing (1) pro and active MMP-2, (2) pro and active MMP-9, (3) p50, and (4) p65 (all from Chemicon), while serial sections were incubated either without primary antibodies or with mouse immunoglobulin (IgG) as controls. Antigen expression was visualized by using the Vectastain detection system and DAB kits according to manufacturer's instructions (Vector, Burlingame, CA).
Elastin quantification
Total elastin content was measured from 1 mm arterial segments, after treatment with cyanogen bromide, as previously described. 33 Briefly, samples were suspended in 70% formic acid, 50 mg/mL cyanogen bromide was added, and the mixture was incubated for 24 h at room temperature, washed thoroughly with boiling water, and then lyophilized, weighed, and normalized by the initial dry mass of the segment.
Evaluation of elastin degradation
Slides were deparaffinized, stained with eosin to enhance elastin's natural autofluorescence, and imaged at 100× using the blue filter (excitation 475 nm, emission 527 nm) on a Zeiss Axiophot 2 microscope (Carl Zeiss, Thornwood, NJ). Digital images were taken for the entire cross-sectional area, and the elastin chain length, the total length of each individual elastin chain, was quantified for three to four arteries in each group using ScionImage 5.0 (Scion, Frederick, MA). The average length was chosen as the unit measure because it is sensitive to small breaks in the lamina where the total length is not. Images were thresholded at a standard cutoff, converted to binary, and skeletonized to obtain a 1-pixel-wide representation of the cross sections of individual elastin laminae. From the resulting images, the average length of the skeletonized objects was then calculated, similar to the techniques used previously for quantifying cell networks. 34 A macro was written to automate this process and is available upon request. As a positive control, sections were treated with elastase (Sigma-Aldrich, St Louis, MO) for 1 h at room temperature, before eosin staining, and then analyzed as described.
Statistical evaluation
Data were imported into JMPin® software version 4.0 (SAS, Cary, NC) for determination of statistical significance. Groups were evaluated using ANOVA with Tukey–Kramer post hoc test with α set to 0.05.
Results
MMP-2 expression increased by hemodynamics or elongation
MMP-2 expression was determined using gelatin zymography then quantified using Biorad's Quantity One densitometry software (Fig. 1). On day 2, the expression of total (pro+active) and active MMP-2 increased significantly from fresh tissue, with total MMP-2 activity increasing roughly fourfold regardless of hemodynamic conditions. The MMP-2 expression levels continued to increase during culture, with all arteries having significantly greater total and active MMP-2 expression on day 9 than arteries sampled on day 2. However, by day 9, mechanical stimulation through increased axial strain, reduced hemodynamics, or the combination of these two stimuli all had the similar effect of significantly increasing active and total MMP-2 expression relative to arteries perfused under arterial hemodynamic conditions at in situ length (p < 0.05).
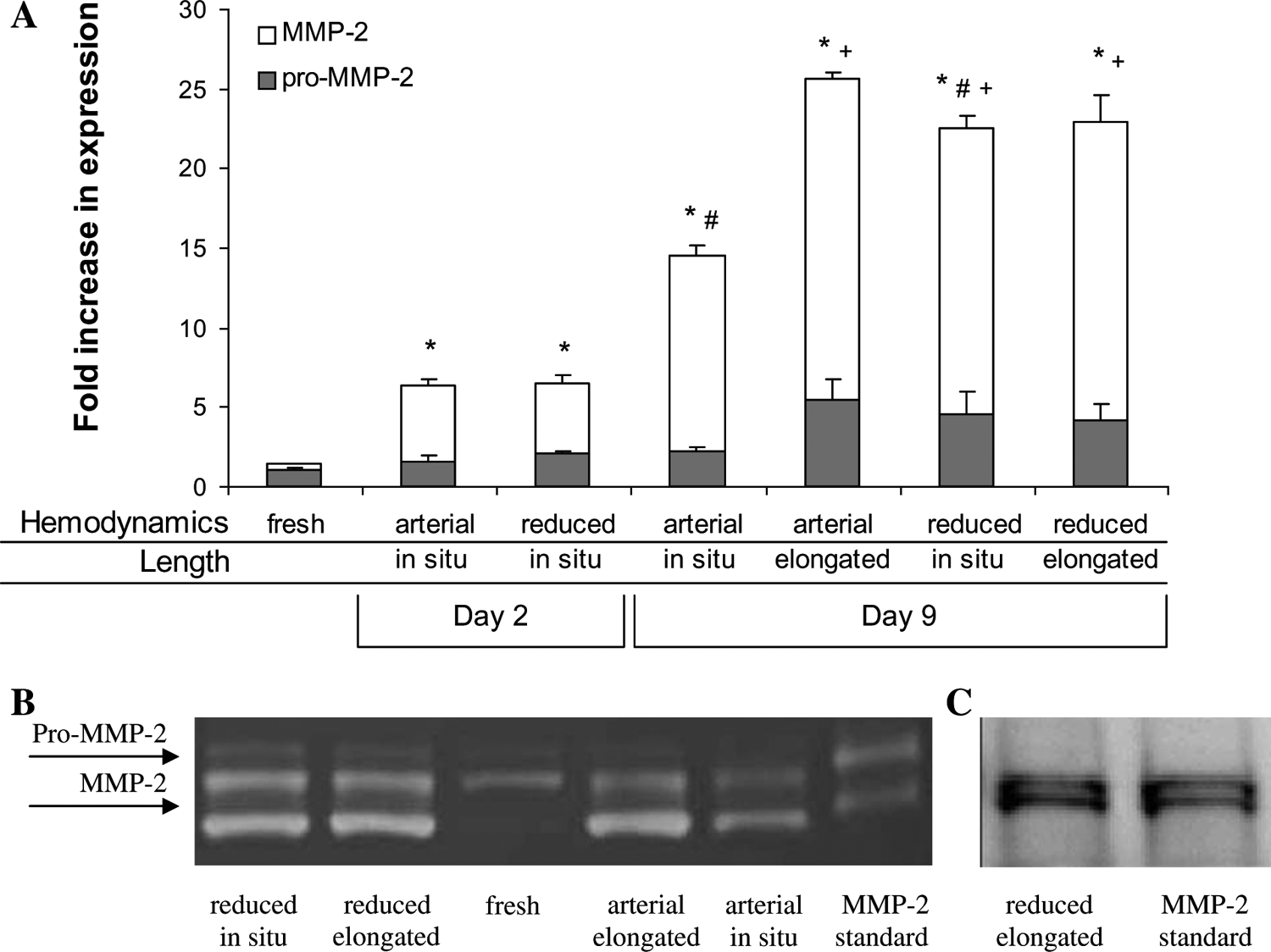
MMP-2 expression. (
MMP-9 increased by hemodynamics and elongation; combination caused additive increase
MMP-9 was undetectable in fresh arteries even when the total protein loaded was doubled to 20 μg. Similarly, while active MMP-9 was detected in some samples, expression levels were below reliable detection limits, and therefore all MMP-9 expression reported refers to proMMP-9. By day 2, arteries perfused at in situ length and with either arterial or reduced hemodynamics demonstrated MMP-9 activity, with reduced hemodynamics causing significantly greater expression than arterial hemodynamics at day 2 (Fig. 2, p < 0.05). This significant difference was even more substantial on day 9, where reduced hemodynamics caused roughly a threefold increase in MMP-9 expression compared to the expression on day 2 (p < 0.05), while arterial hemodynamics caused no further increase. Similarly, arteries elongated under arterial hemodynamics had significantly greater MMP-9 expression on day 9 by roughly threefold than arteries perfused at in situ length (p < 0.05). Similar to the MMP-2 expression, stimulating arteries with either increased axial strain or reduced hemodynamic conditions lead to a similar significant increase in MMP-9 expression, which was not significantly different from each other. However, the MMP-9 expression induced by the combination of reduced hemodynamics and increased axial strain was significantly greater than either stimulus alone and roughly equal to the combination of the two individual stimuli (p < 0.05). These data suggest that axial strain and hemodynamics have an additive effect on MMP-9 expression, but not on MMP-2 expression.
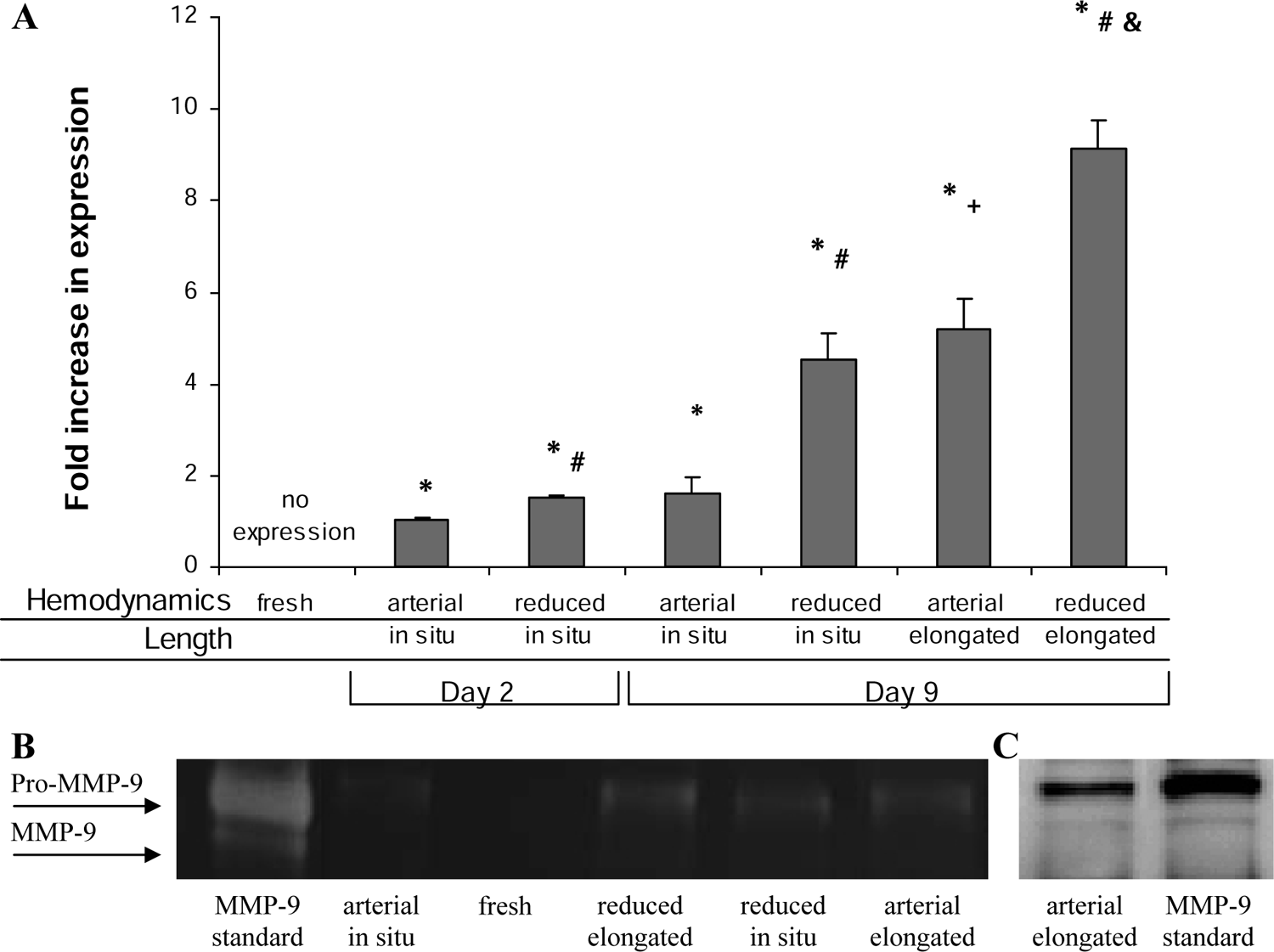
MMP-9 expression. (
MMP-2 and MMP-9 histological expression correlates roughly with protein expression
Histological sections from fresh arteries and those cultured for 9 days under each condition were stained using an antibody recognizing both pro and active MMP-2 (Fig. 3) to determine the spatial expression of MMP-2 throughout the artery wall. Fresh tissue and arteries perfused under arterial hemodynamics at in situ length showed similar levels of expression (Fig. 3E, A). Similar expression of MMP-2 was found in arteries elongated under arterial hemodynamics or subjected to reduced hemodynamics (Fig. 3B, C). The strongest expression was seen in arteries elongated under reduced hemodynamics (Fig. 3D), with especially strong staining observed along the internal elastic lamina (IEL)/endothelial region.
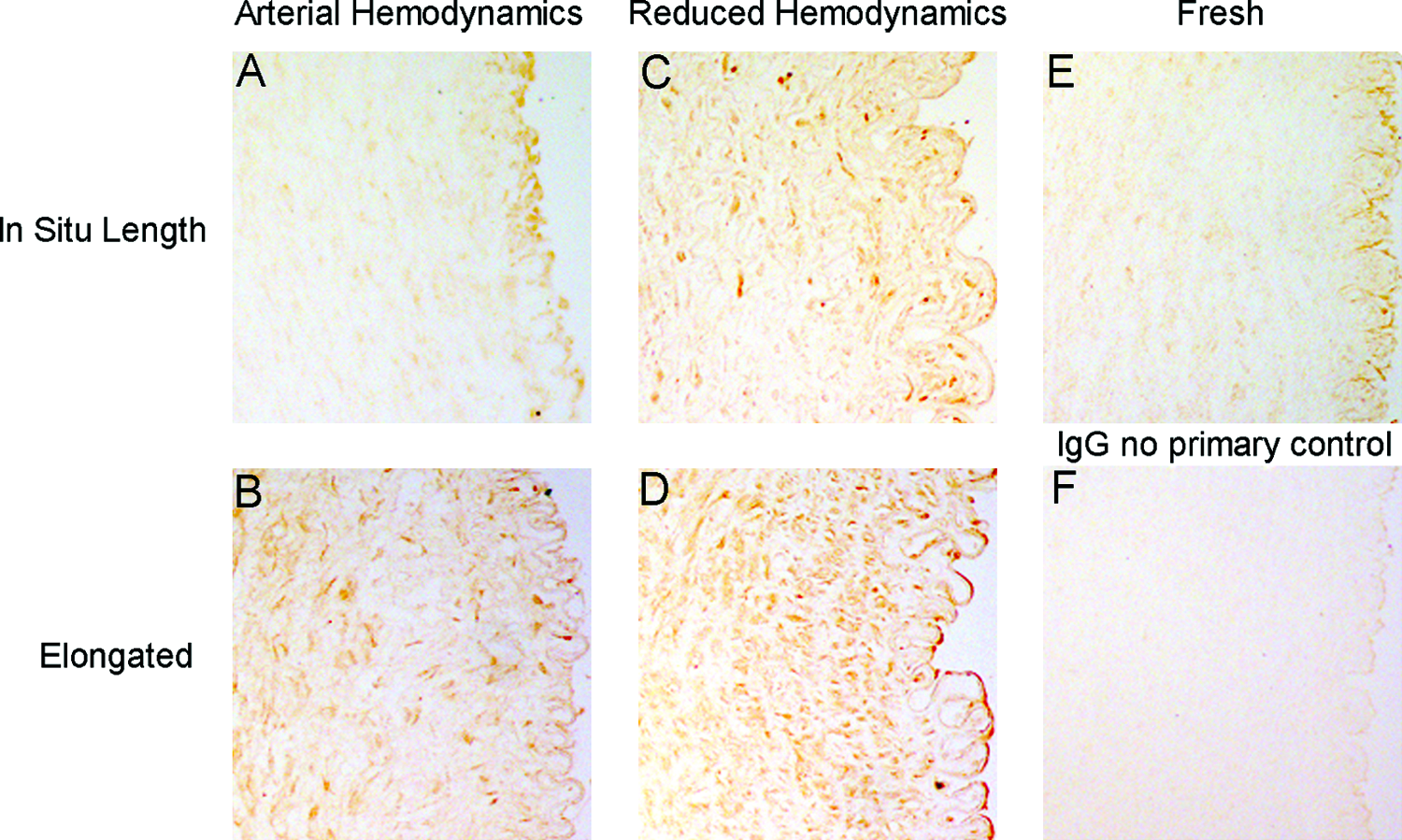
MMP-2 immunohistochemical staining. Representative cross-sectional images of MMP-2 in (
Immunostaining for pro and active MMP-9 demonstrated expression that was dependent on the mechanical environment (Fig. 4). Fresh tissue and arteries perfused under arterial hemodynamics at in situ length displayed basal MMP-9 staining with expression being relatively uniform throughout the cross section (Fig. 4E, A). In arteries perfused under arterial hemodynamics, elongation caused increased MMP-9 staining throughout the artery wall (Fig. 4B). Reduced hemodynamics also increased overall MMP-9 expression throughout the artery wall; however, the presentation was punctate and located near the nuclei rather than the diffuse expression seen in arterial hemodynamic samples. The combination of reduced hemodynamics and elongation displayed strong diffuse MMP-9 expression throughout the artery wall and strong localized punctate expression, exhibiting expression similar to that of both reduced hemodynamic and arterial elongated arteries.
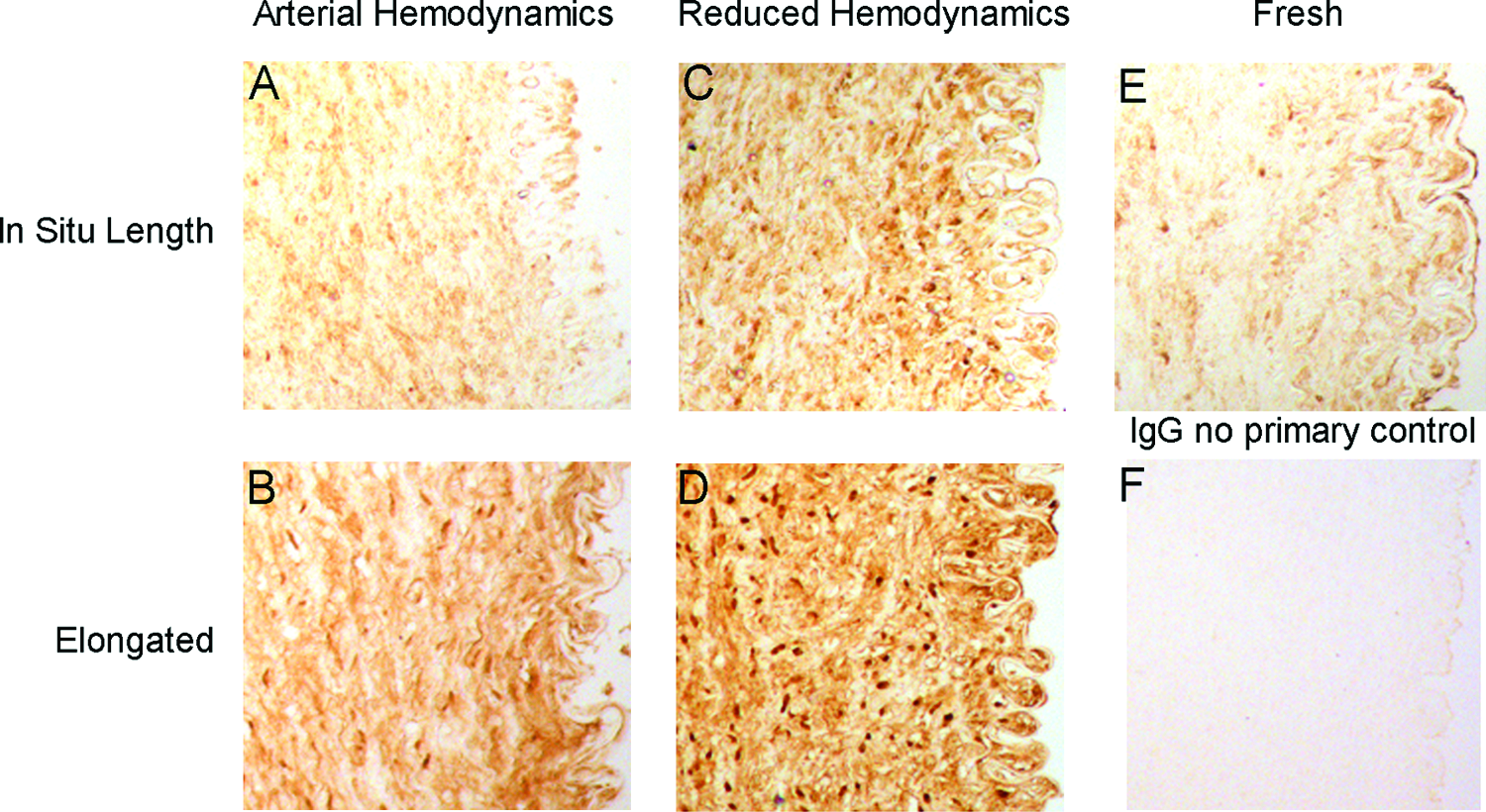
MMP-9 immunohistochemical staining. Representative cross-sectional images of MMP-2 in (
Average elastin content similar regardless of hemodynamic conditions or elongation
Elastin is abundant in the arterial wall, is a major target for MMP-2, MMP-9, as well as other MMPs, and almost certainly will not achieve appreciable replacement in only 9 days, making maintenance and assessment of native elastin content and orientation paramount to the creation of engineered arteries. Elastin chains were visualized in histological sections (Fig. 5) to determine if and where degradation might have occurred, and the total quantity was determined through a biochemical assay (Fig. 5G). Biochemical quantification of total elastin revealed no significant differences in the relative elastin mass under any conditions tested (Fig. 5G). However, quantification of average chain length determined that fresh arteries and arteries perfused under arterial hemodynamics at in situ length had a significantly longer average chain length than did arteries perfused at in situ length under reduced hemodynamics or elongated under arterial or reduced hemodynamics (Fig. 5H, p < 0.05). This was visualized as small gaps in otherwise continuous concentric rings of elastin throughout the media layer. Taken together, the absence of appreciable gain or loss in total elastin content, combined with a significant decrease in elastin chain length in arteries with significantly greater MMP expression, suggests that MMP-mediated elastin remodeling is occurring, while widespread, nonspecific elastin degradation is not.
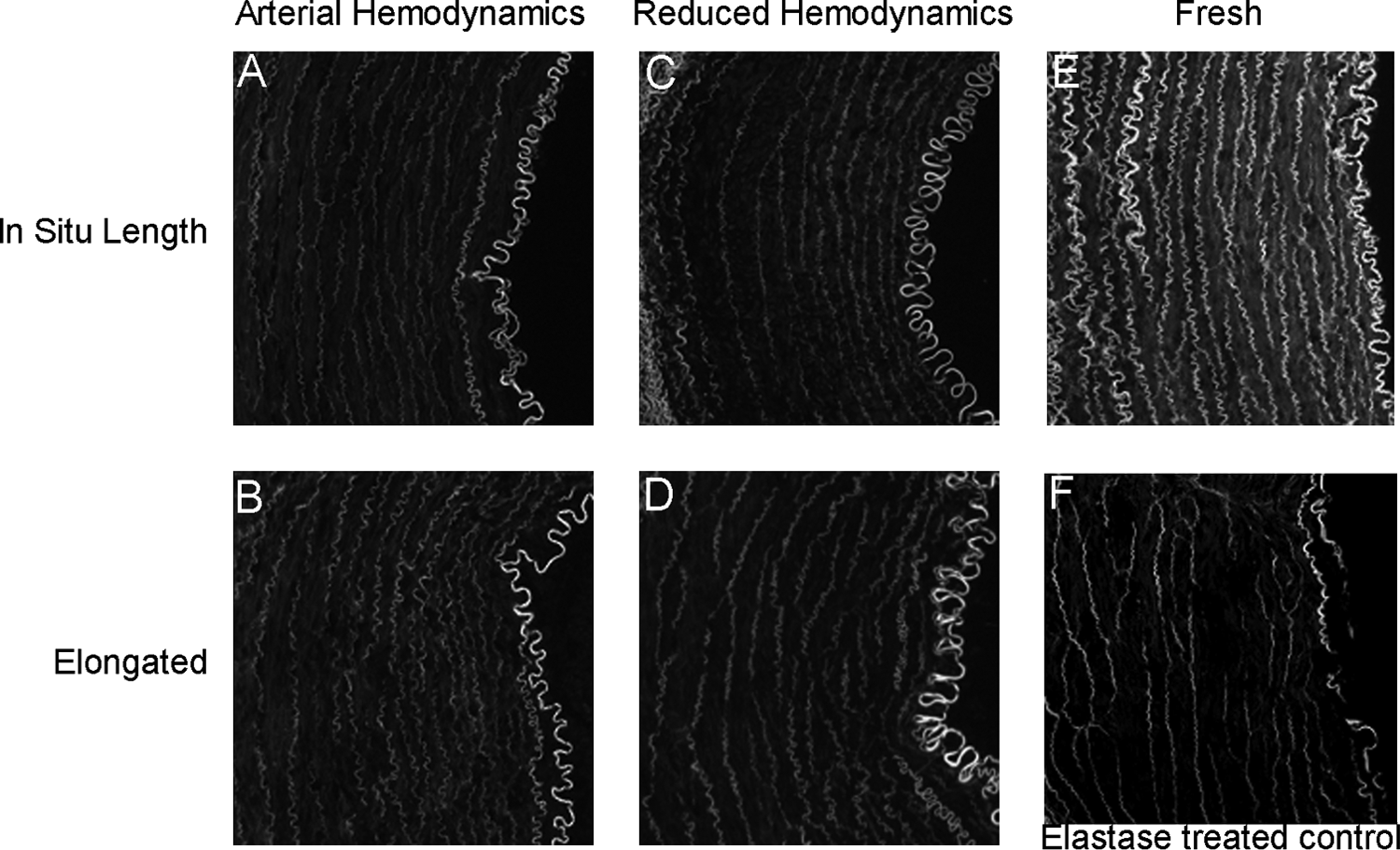
Elastin analysis. Representative cross-sectional images of elastin in arteries perfused under arterial hemodynamics at in situ length (
Reduced hemodynamics causes nuclear localization of p50, while axial strain induces nuclear localization of p65
To investigate possible mechanisms for mechanically induced stimulation of MMP-9, sections of arteries were stained with antibodies to the p50 and p65 subunits of NF-kB (Fig. 6). Fresh tissue and arteries perfused under arterial hemodynamics at in situ length showed little to no expression of either p50 or p65 (Fig. 6A–D). Arteries elongated under arterial hemodynamics showed little p50 expression (Fig. 6E); however, there was substantial diffuse and nuclear localized p65 expression (Fig. 6F). Arteries perfused at in situ length under reduced hemodynamics showed strong expression of p50 throughout the artery wall, especially around the nuclei (Fig. 6G), contrasted with little p65 expression (Fig. 6H). Combining reduced hemodynamics and elongation resulted in increased staining for both p50 (Fig. 6I) and p65 (Fig. 6J) throughout the artery wall and localized in nuclear regions. This demonstrated that the simultaneous reduction of hemodynamics and elongation lead to increases in both diffuse and nuclear localized expression of p50 and p65 subunits. In contrast, increased p50 expression alone was observed with reduced hemodynamics only, and increased p65 expression alone was observed with elongation only.
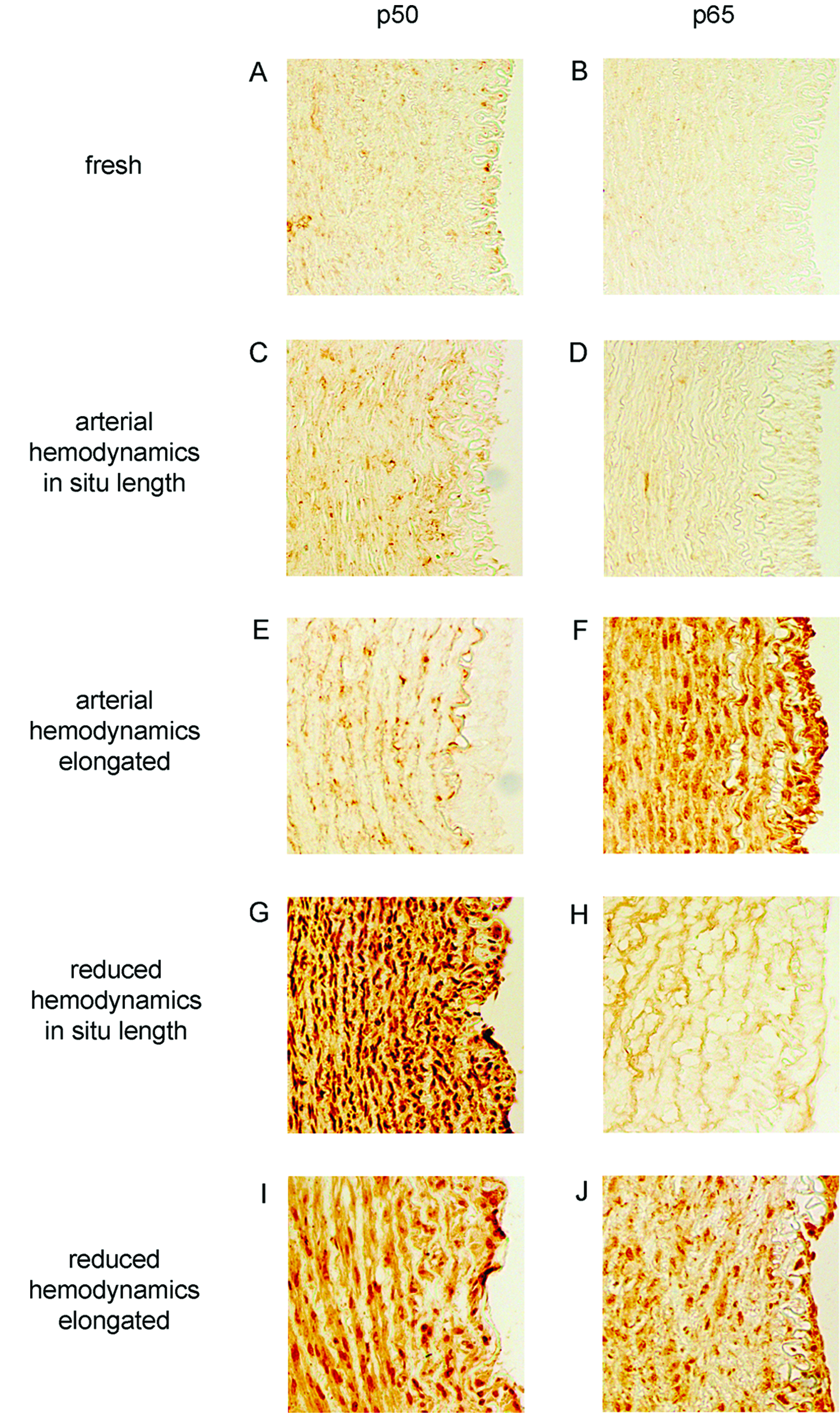
Expression and nuclear localization of p50 and p65. Expression is largely not present in fresh arteries (
Discussion
The significant findings from this study were (1) increased axial strain and reduced hemodynamics were each individually sufficient to cause a significant increase in MMP-2 and MMP-9 protein expression in engineered arteries; (2) combining reduced hemodynamics with increased axial strain caused no further increase in MMP-2 expression, but caused a significant additive increase in MMP-9 expression, correlating with previously reported synergistic remodeling; (3) expression and nuclear localization of p50 and p65 subunits responded differently to reduced hemodynamics and increased axial strain, which may suggest a potential mechanism for the additive MMP-9 expression. These data provide evidence of molecular mechanisms regulating mechanically inducedarterial remodeling, which could be exploited to further direct the remodeling in engineered arteries.
MMP expression in ex vivo and in vivo elongation
Recent studies have investigated the effect of increased axial strain on arteries both in vivo and ex vivo;15–18,35 however, few have investigated the role of MMPs in the remodeling process. In one such study, a one-time length increase of roughly 20% in rabbit carotid arteries caused increased ECM deposition and cell proliferation in vivo. 23 MMP-2 expression was elevated early and remained increased for 7 days; however, the axial strain normalized entirely in that amount of time through retraction and remodeling. Despite substantial MMP-2 expression, this study showed only transient MMP-9 expression immediately after the surgery that was no longer present after 1 day. In our system, the goal was to create engineered arteries using increased axial strain such that the arteries were unable to reduce their axial strain through retraction. Further, arteries were subjected to greater than twice the increase in axial strain (50% vs. ∼20%) in comparison to the previous study by Jackson et al. 31 The use of gradual step-wise elongation versus one-time application of mechanical elongation may explain why MMP-9 expression increased over time in our ex vivo results but not in the in vivo study of Jackson et al. 31
MMPs increased by ex vivo perfusion alone are significantly less than mechanically induced remodeling
While MMP-2 and MMP-9 expression was significantly increased by either reduced hemodynamics or increased axial strain, arteries cultured at in situ length under arterial hemodynamics displayed significantly higher MMP expression than fresh tissue, suggesting that some component(s) of the ex vivo perfusion system or the vessel harvest process was inducing MMP activity. Clearly, the harvest process is a traumatic event because it removes all axial tension for 1–2 h, instantaneously retracts the length ∼33%, and much of the adventitia and vaso vasorum are removed. Others have shown that a one-time stretch of human saphenous veins before culture can cause a significant increase in MMP-2 and MMP-9 activity after 7 days of ex vivo perfusion, demonstrating how even a short mechanical stimulus can cause prolonged MMP expression long after the stimulus is removed.31,36 Similar to the findings presented in this report, Meng et al. also showed that levels of proMMP-9, proMMP-2, and MMP-2 all were elevated by day 7 in both control and stretched vein segments, yet stretched veins still showed a significantly greater increase in MMP expression. 31 In addition, Meng et al. also reported the inability to detect active MMP-9 in zymograms, hypothesizing that this was possibly due to quick release from or degradation within the tissue. These data suggest not only that one isolated mechanical stimulus can cause sustained MMP expression up to a week later, but also that factors associated with combination of harvest and ex vivo culture might increase MMP expression in perfused vessels.
In another study from the same group, perfusing carotid artery segments from 100 kg pigs ex vivo under normal arterial hemodynamics for 48 h elevated MMP-9 expression and significantly increased MMP-2 expression. 22 This provides further evidence that factors associated with ex vivo perfusion (e.g., initial injury, differences in mechanical environment, and serum/growth factor stimulation) can cause increased basal levels of MMP expression. In our system, as shown in the other studies, all experimental alterations of hemodynamics or axial strain caused a further and significant increase in MMP-2 and MMP-9 expression as compared to arteries perfused under arterial hemodynamics at in situ length that cannot be explained by ex vivo perfusion alone. Further, the fact that arteries perfused at in situ length under arterial hemodynamics showed no significant decrease in the average elastin chain length, while all mechanically stimulated arteries displayed a significant decrease, provides further support that substantial remodeling was not occurring in the ex vivo control.
Expression patterns of MMP-9 correlate with mechanical stimuli
Histological evaluation of MMP-9 expression demonstrated that while overall levels of protein expression caused by reduced hemodynamics or increased axial strain may be similar in magnitude, the location and clustering of the protein were quite different. The fact that expression was diffuse and equally distributed throughout the arterial media in the elongation cases suggests that increased axial strain is being sensed roughly equally by all smooth muscle cells, causing them to equally increase MMP expression to reduce the increased axial stress through remodeling. In contrast, when only hemodynamics was altered, MMP-9 expression was more strongly expressed in the endothelial/IEL region, suggesting that the MMP expression was being influenced more strongly by endothelial signaling. The spatial distribution of MMP expression is consistent with in vivo observations that endothelial denudation blocks MMP-mediated inward remodeling caused by decreased flow 29 but not remodeling due to decreased 24 or increased 23 axial loading. The combination of mechanical stimuli appears to express MMP equally throughout the artery wall while also demonstrating strong nuclear staining, seemingly combining the presentation profiles of the individual stimuli.
NF-kB subunit localization specific to mechanical stimuli correlates with MMP-9
Reduced hemodynamics has been shown to decrease NO release, which in turn leads to contraction of the lumen, initiating the inward remodeling response mediated by MMP expression.20,21 NO inhibition has been shown to increase MMP-9 production, 26 through decreased binding of NO to IkB, 37 an NF-kB nuclear chaperone, suggesting that NF-kB nuclear localization induces MMP-9 activity. Similarly, in vascular smooth muscle cells, it was reported that inhibition of NF-kB through overexpression of IkB nearly eliminated MMP expression, suggesting that NF-kB is required for MMP induction. 38 It was also shown that NO inhibits nuclear localization of the p50 subunit of NF-kB in endothelial cells in vitro. 37 Interestingly, constant stretch in endothelial cells in vitro induced nuclear localization of NF-kB subunit p65. 39 This, combined with the knowledge that the MMP-9 gene contains individual promoter regions for each the p50 and p65 subunits, made p50 and p65 attractive potential mediators to investigate for dual pathway–induced MMP-9 production. 40 The fact that p50 and p65 were differentially expressed in response to altered hemodynamics and axial strain or stretch, yet both subunits were expressed when the stimuli were combined, suggests that NF-kB is at least partially responsible for the demonstrated additive MMP-9 expression. This potential relationship merits further investigation to elucidate the mechanisms involved.
Overall, the differential regulation of MMP-9 in response to different combinations of mechanical stimuli provides new evidence into how arterial remodeling may occur in response to both physiological and pathological stimuli. These mechanisms are important to our understanding of remodeling in native and engineered arteries, and provide evidence as to why arteries perfused under reduced conditions were more responsive to increased axial strain. The presented work also demonstrates the ability of ex vivo perfusion to investigate the remodeling responses to both individual and combined mechanical stimuli in a controllable manner to better evaluate and direct the growth remodeling of native and engineered tissues. With better knowledge of the molecular response to mechanical stimuli, these mechanisms can be more effectively exploited to create the optimal remodeling environment for creating engineered arteries and other tissues.
Footnotes
Acknowledgments
We would like to thank the Whitaker Foundation (Biomedical Engineering Research Grant [K.J.G.], Predoctoral Fellowship [J.W.N.]) and the National Institutes of Health (R01 HL064388-01A1) and the American Heart Association (0655323B, 0555538U) for funding this work.
Disclosure Statement
No competing financial interests exist.