
Abstract
Fibrin is a hydrogel carrier widely used in cartilage tissue engineering. It is rapidly degraded by plasmin, which is produced by the cells. ɛ-Aminocaproic acid (EACA) can be used to inhibit this enzyme and thus save the fibrin carrier. In this study we investigated the effect of EACA on the transforming growth factor β-1–induced chondrogenic differentiation of human bone marrow–derived mesenchymal stem cells (hMSCs). To assess this, we used the standard pellet culture system, and EACA treatment was compared to an untreated chondrogenic control. To investigate differentiation, real-time RT-PCR was used on chondrocytic marker genes: aggrecan, collagen types II and X, and the SRY-related HMG-box gene 9 (SOX9). Also, specific glycosaminoglycan production was measured. Safranin-O/fast green staining was used to localize proteoglycans and collagens within the pellet. All results concur that EACA did not affect the chondrogenic differentiation process at 5 μM concentration, which is adequate to inhibit fibrin degradation. Therefore, it is a useful plasmin inhibitor for cartilage tissue engineering with hMSCs.
Introduction
Because of good manageability and its role in the natural healing process, the fibrinogen–fibrin system is a logical choice for scaffolding and delivery agent for cells with regenerative capacity. The components of the tissue engineering design can be conveniently mixed with fibrinogen and will only solidify when exposed to thrombin.
Fibrin, however, degrades very rapidly in the presence of plasmin, which is produced by many cell types. 1 While the removal of the clot is desirable during tissue healing, it is a drawback in the tissue engineering process. Thus, protease inhibitors must be applied in studies longer than a few hours.
A commonly used inhibitor is aprotinin. It is a polypeptide, routinely isolated from bovine lung. This competitive inhibitor blocks the active site of serine proteases such as trypsin, chymotrypsin, kallikrein, and plasmin.
Another well-known inhibitor is ɛ-aminocaproic acid (EACA, 6-aminohexanoic acid), a lysine analog. It promotes the dissociation of plasmin, thereby inhibiting the activation of plasminogen and subsequent fibrinolysis. 2 Apart from plasmin it is known to affect carboxypeptidase B and a component of the complement system.3,4 The isolation of aprotinin is expensive, and it is difficult to reach high purity. On the contrary, the synthesis of EACA is much more cost-effective. According to the information from the manufacturer, it also affects fewer enzymes than aprotinin; thereby it is a more specific inhibitor of plasmin and fibrinolysis. Based on these features EACA appears to be suitable for use in fibrin-based constructs for cartilage tissue engineering, and it has been used for this purpose before. 5
However, a recent study by Huang et al. reported that aprotinin and EACA influenced the transforming growth factor beta-1 (TGF-β1)–induced chondrogenic differentiation of rabbit bone marrow mesenchymal stem cells. 6 Should this also occur in the human counterparts of these cells, it would render EACA unfeasible for the engineering of cartilaginous constructs.
The goal of this investigation was to find out whether EACA interferes with the chondrogenic differentiation process of human bone marrow–derived mesenchymal stem cells (hMSCs). To examine this, we employed the standard chondrogenic pellet culture model devised by Johnstone et al. 7 and Yoo et al. 8
Materials and Methods
Isolation and expansion of hMSCs
Bone marrow was obtained with ethical approval and the written consent of the patients. The bone marrow was diluted 1:5 with Dulbecco's modified Eagle's medium (DMEM) containing 5% fetal calf serum. The mixture was layered on a Ficoll cushion and centrifuged at 800 g for 20 min. Mononuclear cells were collected from the interface, and washed with DMEM. The isolated cells were plated in polystyrene cell culture flasks and supplemented with α-modified essential medium, 10% fetal calf serum, and 5 ng/mL recombinant human basic fibroblast growth factor, and then cultured at 37°C, 5% CO2, and 95% humidity. The medium was changed after 3–5 days, and twice a week afterward. The third-passage cells were used in three experiments for pellet culture.
Pellet culture of hMSCs
Third-passage hMSCs were trypsinized, and 2 × 105 cell aliquots were spun down at 500 g for 5 min in 15 mL polypropylene conical tubes. The pellets were cultured in three groups:
Noninduced control: DMEM with 4.5 g/L glucose, ITS + 1 (10 μg/mL insulin from bovine pancreas, 5.5 μg/mL human transferrin [substantially iron-free], 5 ng/mL sodium selenite, 0.5 mg/mL bovine serum albumin, and 4.7 μg/mL linoleic acid; Sigma-Aldrich Chemie GmbH, Buchs, Switzerland), 1% nonessential amino acids (Sigma), 50 μg/mL ascorbic acid 2-phosphate, and 10−7 M dexamethasone. Chondrogenic control: Same as noninduced control plus 10 ng/mL recombinant human TGF-β1. EACA: Same as chondrogenic control, with the addition of 5 μM EACA.
Each pellet was cultured in 0.5 mL medium at 37°C, with 5% CO2 and 95% humidity. The culture medium was changed three times a week, and pellets were harvested at days 14 and 21. Conditioned medium was collected for biochemical analysis.
DNA
Cell number was evaluated by measuring the DNA content at different time points. Pellets were digested with 0.5 mg/mL recombinant Proteinase-K (Roche Diagnostics GmbH, Mannheim, Germany) at 56°C for approximately 16 h. DNA concentrations were determined with the Hoechst method as described by Labarca and Paigen. 9 Fluorescence intensity was measured with an HTS 7000 Perkin-Elmer Bio Assay Reader (Norwalk, CT).
Glycosaminoglycan content of the medium and pellets
The amount of glycosaminoglycan (GAG) was determined by the dimethylmethylene blue dye method, using bovine chondroitin sulfate as the standard. 10 Proteinase-K digests were used to measure the GAG content of the pellets. The total GAG content of the culture media, collected at each media change, was also measured to assess the release of matrix molecules from the sample into the media. These total values were normalized with the DNA content to characterize cell activity. Then, the activity values were compared to the chondrogenic control pellet of the respective experiment. This step was necessary because of the inter-individual variability.
RNA isolation and real-time RT-PCR
Pellets were lysed with TRI Reagent® (Molecular Research Center, Cincinnati, OH), and the RNA isolation was carried out according to the protocol from the manufacturer. RNA was reverse transcribed with TaqMan reverse transcription kit (Applied Biosystems, Foster City, CA) using random hexamers to initiate transcription. For real-time PCR we used TaqMan PCR analysis kits, on a GeneAmp 7500 Real-Time PCR System (Applied Biosystems). SRY-related HMG-box gene 9 (SOX9) gene expression assay was ordered from Applied Biosystems (product no. Hs00165814_m1). Aggrecan (ACAN), collagen type II α1 chain (COL2A1), and collagen type X α1 chain (COL10A1) primers and probes are listed in Table 1.
Approximation of pellet size
Before histological analysis, the pellets were placed on a standard microscope slide, and transmitted light images were taken with a Zeiss Axiovert25 microscope (Carl Zeiss MicroImaging GmbH, Jena, Germany) using the same settings for all samples. On these images, the dense pellets appeared black or nearly black in front of a clearly distinguishable bright background. Then, the images were gated so that a black-and-white image was acquired, on which the largest contiguous black region was the projection of the pellet. The area of this region (pixels) was quantified with the ImageJ's Nucleus Counter plugin (Bethesda, MD). 11
Safranin-O staining
Histology samples were fixed in 100% methanol, and then rehydrated in PBS, containing 5% sucrose before cryosectioning at 6 μm. The sections were sequentially stained with Meyer's hematoxylin, 0.02% fast green, and 0.1% safranin-O to visualize the proteoglycan and collagen accumulation. The slides were mounted with a xylene-based mounting medium (Eukitt; O. Kindler GmbH, Freiburg, Germany). Light micrographs were taken with a Zeiss Axioplan2 microscope at 5 × magnification, with consistent settings to gain comparable results.
Statistics
The normality of the sample distributions was determined with the Shapiro–Wilk normality test. In case of normal distribution, ANOVA followed by pairwise t-tests were used to judge significance between the treatment groups. In case of nonnormal distribution, Kruskal–Wallis rank sum test followed by pairwise Wilcoxon rank sum tests were calculated. When using pairwise tests, the p-values were adjusted by Bonferroni's method. Statistical analysis was performed with the help of the R package for statistical computing. 12
Results
TGF-β1 efficiently induced chondrogenic differentiation in the pellet cultures. The pellet size proved to be a good marker of enhanced anabolic activity (Fig. 1). The TGF-β1–treated pellets were significantly larger than the noninduced controls. EACA did not influence the pellet size.
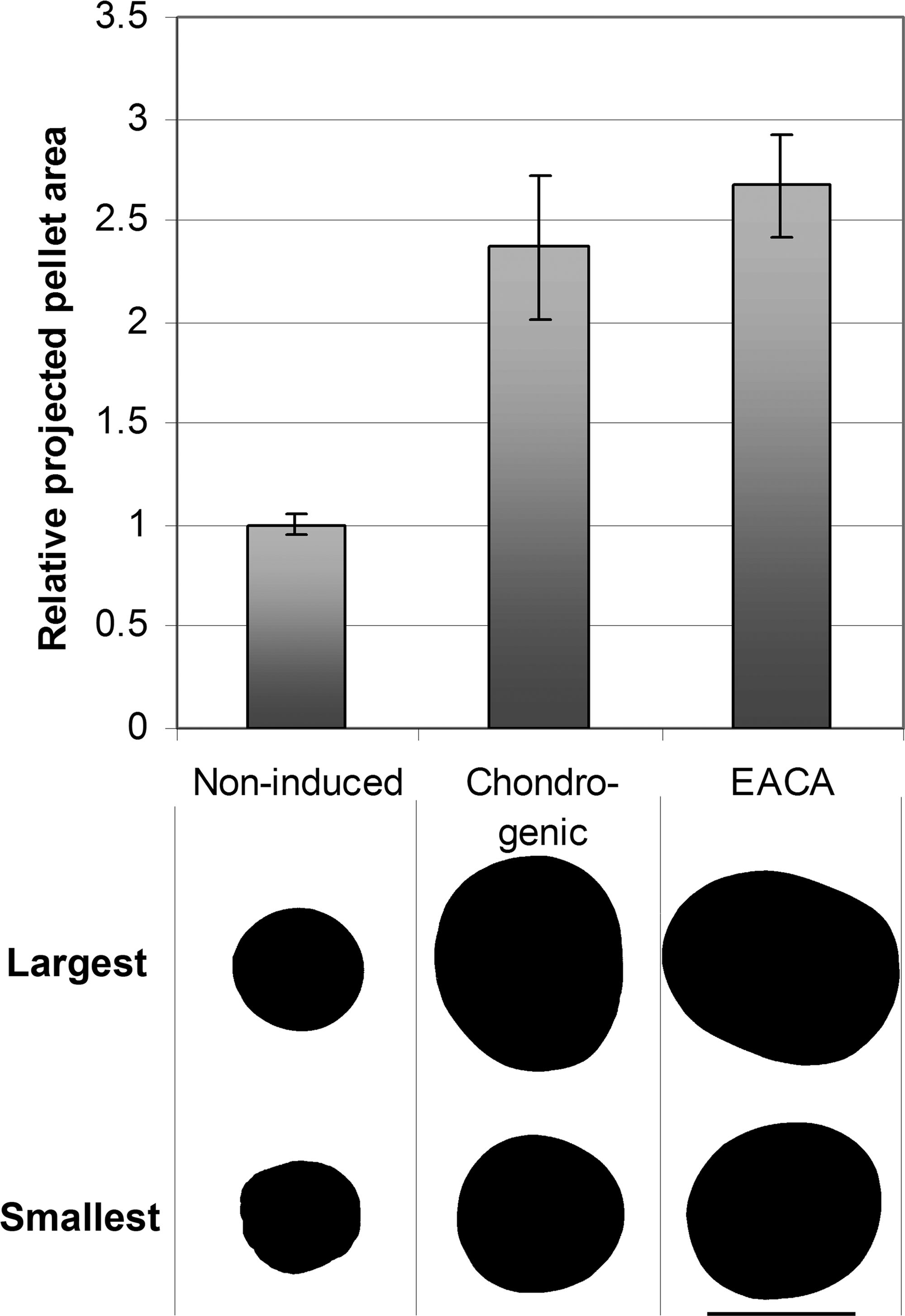
Pellet size variation. Transmitted light images of all pellets were acquired at the same magnification at day 14. The areas of the pellets' projections were calculated with the ImageJ image analysis program, and are plotted relative to noninduced control. Each group contained eight pellets from the same donor. The error bars represent ± SD. The projections of the largest and the smallest pellets are shown under each group. The scale bar in the bottom right corner is 1 mm. Noninduced, pellet without TGF-β; chondrogenic, TGF-β1 induced; EACA, chondrogenic + EACA.
Gene expression analysis demonstrated that the expression levels of chondrogenic genes ACAN, COL2A1, and SOX9 were increased by TGF-β1 treatment, as was the hypertrophic marker COL10A1. None of these genes showed any response to additional treatment with EACA (Fig. 2).
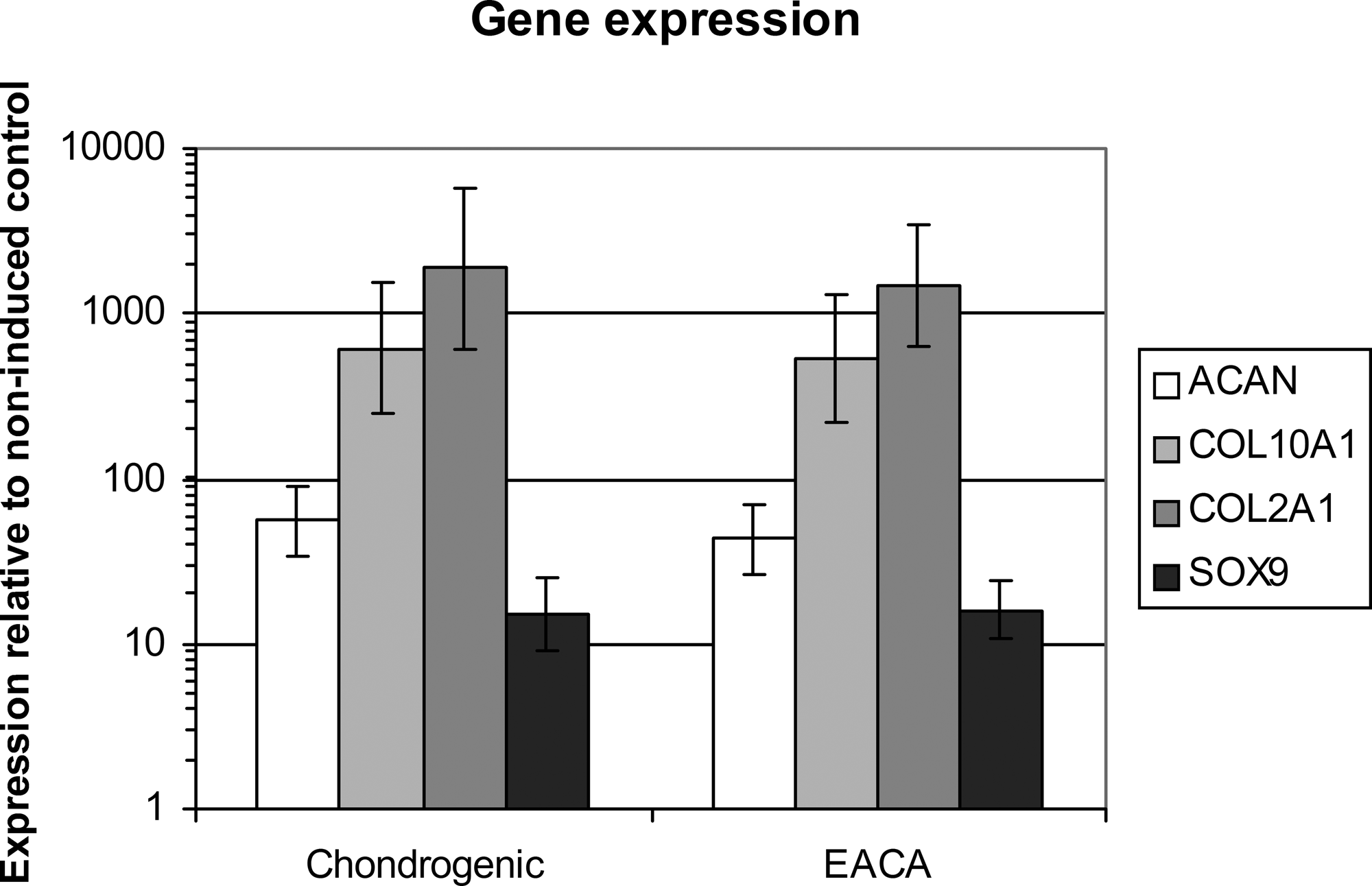
Gene expression patterns. The relative gene expression of chondrogenic and EACA pellets is shown compared to noninduced control (fold increase on a logarithmic scale, day 14). The different shadings represent different genes; the error bars are located at ± SD; n = 6. Chondrogenic, TGF-β1 induced; EACA, chondrogenic + EACA.
There was no difference in GAG release into the medium between any of the treatment groups. The TGF-β1–treated pellets retained 2–3 times more GAGs than the noninduced group, but addition of EACA, again, did not cause a significant difference (Figs. 3 and 4).
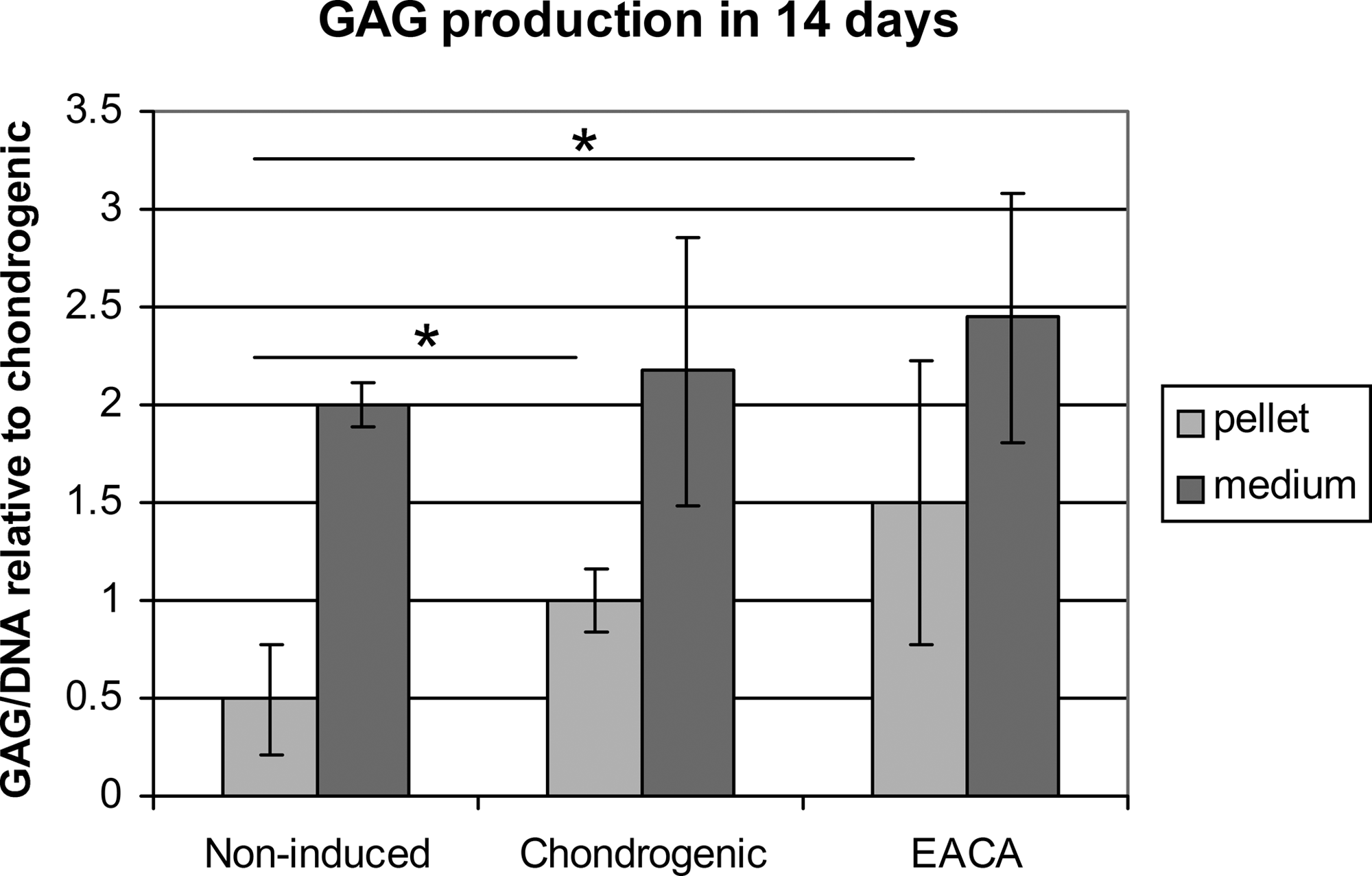
GAG production. GAG amounts were measured in the medium (dark gray) and in the pellets (light gray). They were both normalized to pellet DNA content. GAG/DNA is shown relative to chondrogenic control. The error bars represent ± SD. Nmedium = 15 and Npellet = 7; *p ≤ 0.05. Noninduced, pellet without TGF-β; chondrogenic, TGF-β1 induced; EACA, chondrogenic + EACA.
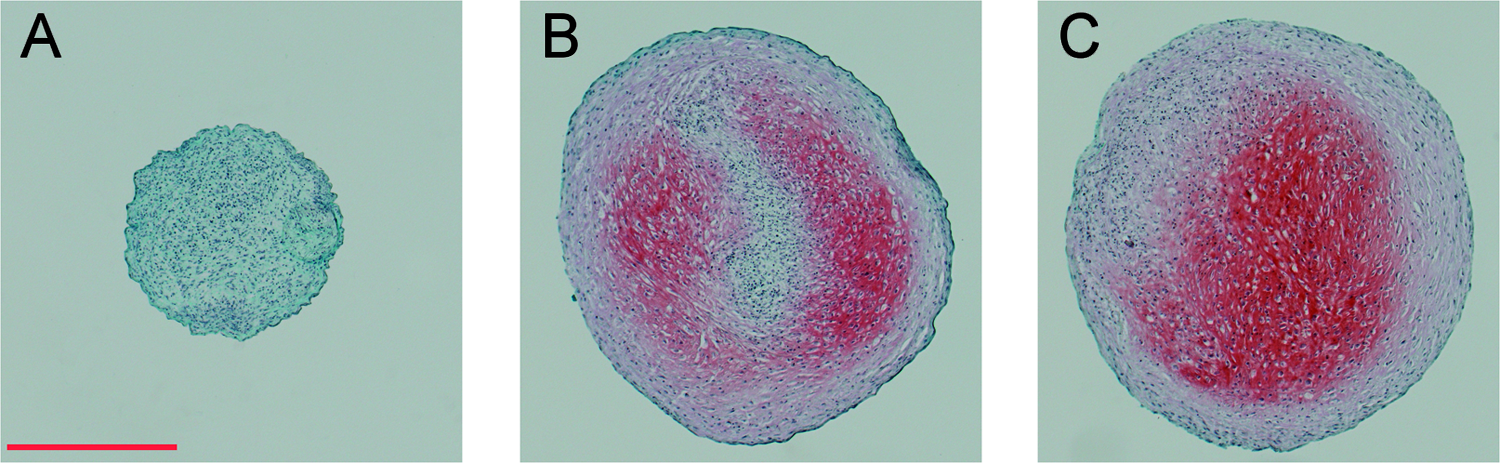
Safranin-O/fast green staining images. Pellets were fixed in methanol after 14 days of culture, and stained with safranin-O, fast green, and Mayer's hematoxylin. (
Discussion
In MSC-based cartilage tissue engineering studies, chondrogenic differentiation of the cells is necessary. Although, the gold-standard chondrogenic system, the pellet culture, is feasible to study cell differentiation, it has certain drawbacks when it comes to tissue engineering. First, the size of the construct is restricted to ∼200,000 cells, due to the limited nutrient supply in the middle of the pellet. The other constraint is that the resulting chondrocytes rapidly progress toward the hypertrophic phenotype, which ultimately leads to apoptosis and calcification.7,8 This is to be avoided in cartilage tissue engineering.
This led to a number of ways to increase the size of the construct, most of which use cells embedded in hydrogels. Alginate, agarose, and fibrin are widely used. The degradation of preformed fibrin can be prevented by micromolar concentrations of EACA. 2 In this study, 5 μM EACA was applied; this concentration is sufficient to inhibit fibrin degradation during 21 days of hMSC culture based on our preliminary experiments.
This compound was found to cause a slight decrease in GAG production and ACAN expression compared to the aprotinin-treated control group of a study performed in rabbit MSCs pellet culture. It is desired to investigate this question in hMSCs, because in the clinically inclined field of tissue engineering, in vitro results from animal models should be verified in human cell culture.
Further, in this type of study it is important to compare the EACA-treated cultures to a nontreated control and not to another plasmin inhibitor, which might also affect differentiation. For this reason, our experiments were carried out in a fibrin-free pellet culture, because it is not possible to maintain fibrin matrix without inhibitors.
Our results show that the standard pellet culture system worked well in our hands, and all the chondrogenic genes investigated (including the hypertrophic marker COL10A1) were upregulated by TGF-β1 treatment. This was supported by increased GAG content. The addition of an adequate amount of EACA to inhibit fibrinolysis did not influence the gene expression pattern or the GAG production of TGF-β1–induced chondrogenic pellets. Further, the macroscopic appearance and size of the pellets, as well as staining by safranin-O, remained unchanged by EACA.
These results may seem to be in contrast with those of Huang et al. However, the two studies are not directly comparable due to the difference in the investigation strategies. We decided to compare EACA to an untreated control, but this prevented us from using fibrin in the cell culture. They chose to apply fibrin gel, but as a result they were unable to use a nontreated control group. Hence, the two investigations more complement rather than contradict each other.
Further, in the rabbit study the EACA concentration was 2 mg/mL (15 mM) and above, while we found that 5 μM EACA inhibits fibrin degradation by human MSCs adequately. In our view, it is beneficial to use the lowest satisfactory concentration of any compound in the culture medium. In addition, even though EACA was shown to be statistically slightly inferior to aprotinin in the work of Huang et al. it is so to such a small extent that it does not necessarily imply biological significance.
In our study the EACA pellets performed at the same level as the nontreated controls; their synthetic activity equaled that of the cultures stimulated by TGF-β1. Because the report by Huang et al. may discourage researchers from using EACA in combination with human MSCs, we find it important to confirm the status of this simple and cost-effective plasmin inhibitor.
All our observations unanimously show that EACA does not interfere with the chondrogenic differentiation of human MSCs, and thus it is a feasible inhibitor of fibrin degradation in cartilage tissue engineering constructs using human cells.
Footnotes
Disclosure Statement
No competing financial interests exist.