
Abstract
The development of functional engineered cartilaginous tissues of sufficient size that can be used clinically to treat large defects remains a major and significant challenge. This study investigated if the introduction of microchannels into chondrocyte-seeded agarose hydrogels would result in the formation of a superior and more homogenous cartilaginous tissue as a result of enhanced nutrient transport. Microchanneled construct cylinders were fabricated via a molding process utilizing a pillared structure to create the required architecture. Constructs were subjected to either constant rotation in a rotational bioreactor system or free-swelling conditions. After 28 days of free-swelling culture the presence of microchannels did not enhance glycosaminoglycan accumulation within the core of the construct compared to solid constructs (0.317 ± 0.002% w/w vs. 0.401 ± 0.020% w/w). However, under dynamically rotating conditions, glycosaminoglycan accumulation in the cores (1.165 ± 0.132% w/w) of microchannel constructs were similar to that in the periphery (1.23 ± 0.074% w/w) of solid constructs, although still significantly lower than their corresponding periphery (1.64 ± 0.133% w/w) after 28 days. These results confirm that cellular nutrient consumption is primarily responsible for creating the spatial gradients in molecules regulating the biosynthetic activity of chondrocytes through the volume of hydrogels, and that changing the scaffold architecture alone may have little effect while the inherent diffusivity of the material remains high. Rather, a combination of forced convection and modified scaffold architecture is necessary to engineer large cartilaginous tissues in vitro.
Introduction
An alternative approach to enhancing the matrix composition and mechanical properties of tissue-engineered constructs is through the use of bioreactor systems, in which tissues are cultured under dynamic conditions. These applied dynamic culturing conditions (perfusion/fluid flow, hydrostatic pressure, or dynamic compressive loading) enhance developing neo-tissues through enhanced nutrient delivery and metabolic waste removal15–17 and/or by providing a mechanical stimulus to the cells.6,18–29 For example, Akmal et al. 17 showed that bovine encapsulated alginate hydrogels cultured in a rotating-wall vessel enhanced DNA, glycosaminoglycan (GAG), and collagen levels. However, histological analysis revealed enhanced Safranin-O staining in the peripheral regions of the constructs compared with the central region.
Hydrogels are a class of biomaterials that are commonly and widely used in cartilage tissue engineering and include, for example, alginate, agarose, poly(ethylene glycol), poly(vinyl alcohol), pluronics, chitosan, collagen, and fibrin.30,31 Hydrogels provide significant advantages over traditional porous-type sponges, including high water content and efficient transport of nutrients and waste removal, and possess the ability to effectively and homogeneously encapsulate cells as they are generally mixed with the gel before gelation. Hydrogels are three-dimensional networks swollen by a solvent (such as water). Agarose hydrogels, a polysaccharide extracted from marine red algae, have been shown to support the chondrogenic phenotype 32 and the synthesis of cartilage extracellular matrix. 33 Chondrocyte–agarose grafts have also been shown to be biocompatible 34 and to produce a morphologically stable hyaline cartilage in approximately half of all defects 18 months after implantation in one particular animal model study. 35 Despite these advantages, diffusion hindrance within agarose–chondrocyte constructs will increase progressively with culture time in parallel with extracellular matrix production, 36 potentially limiting their use as a scaffold material for engineering large cartilaginous tissue in vitro. In the present study, we hypothesized that the introduction of microchannels into agarose constructs, in conjunction with dynamic rotational culturing, would enhance nutrient delivery and fluid flow, thereby providing an environment in which significantly greater elaboration of matrix could be observed throughout the construct compared to free-swelling conditions. If successful, such a culture system could be used to engineer functional cartilaginous tissues of predefined clinically relevant dimensions.
Materials and Methods
Cell isolation and expansion
Adult bovine hock joints were obtained from a local abattoir within 4 h of sacrifice. Full-depth slices of cartilage were harvested under aseptic conditions from the metacarpophalangeal region of four hock joints, and rinsed thoroughly with phosphate buffered saline (PBS) containing penicillin/streptomycin (200 U/mL). Chondrocytes were isolated from cartilage slices via serial digestion with pronase (1 mg/mL) for 1 h with agitation and rinsing with PBS followed by incubation with Dulbecco's modified Eagle's medium (DMEM)/F-12 containing collagenase type II (0.5 mg/mL) (all from Sigma–Aldrich, Dublin, Ireland) for 16–18 h under constant rotation at 37°C. The cell suspension was passed through a 40-μm-pore cell sieve (Falcon, Sarstedt, Ireland), and the filtrate centrifuged and rinsed with PBS twice. Cells were seeded at a density of 50,000 cells/cm2 in 175-cm2 T flasks and cultured to passage one. Cell number and viability were determined using a hemocytometer and 0.4% trypan blue staining. Isolated chondrocytes from all joints (minimum of three animals) were pooled and maintained in DMEM/F-12 (Sigma–Aldrich) supplemented with 10% v/v fetal bovine serum and 100U/mL penicillin/streptomycin (GIBCO–Biosciences, Dublin, Ireland) during the expansion phase.
Nonchanneled and channeled cell-encapsulated hydrogel constructs
Bovine chondrocytes (passage one) were suspended in DMEM and mixed with 4% agarose (Type VII, Sigma–Aldrich) at a ratio of 1:1 at ∼40°C, to yield a final gel concentration of 2% and a density of 15 million cells/mL. The agarose–cell suspension was cast in a stainless steel mold to produce regular nonchanneled construct cylinders (Ø 6 × 4 mm).
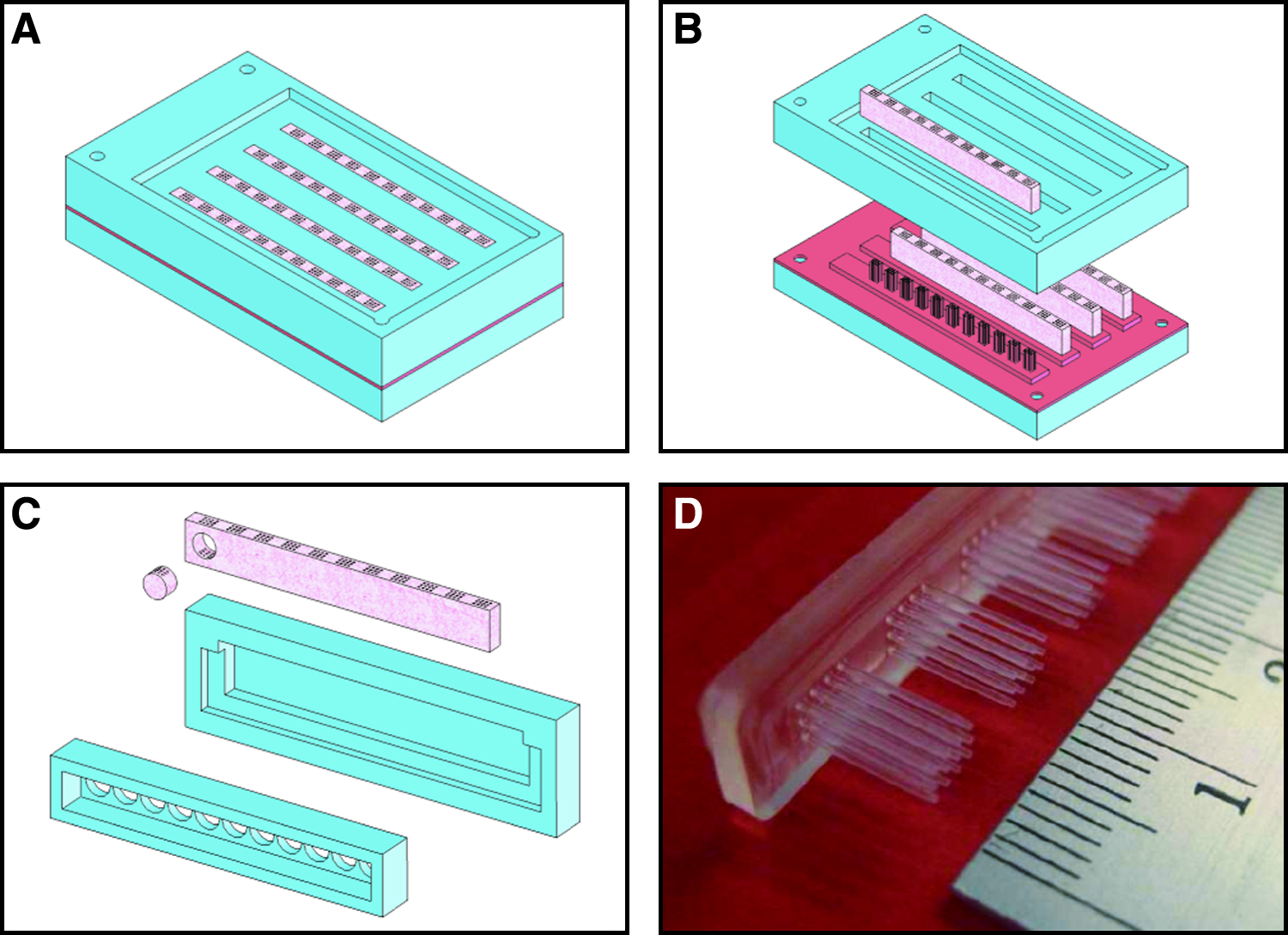
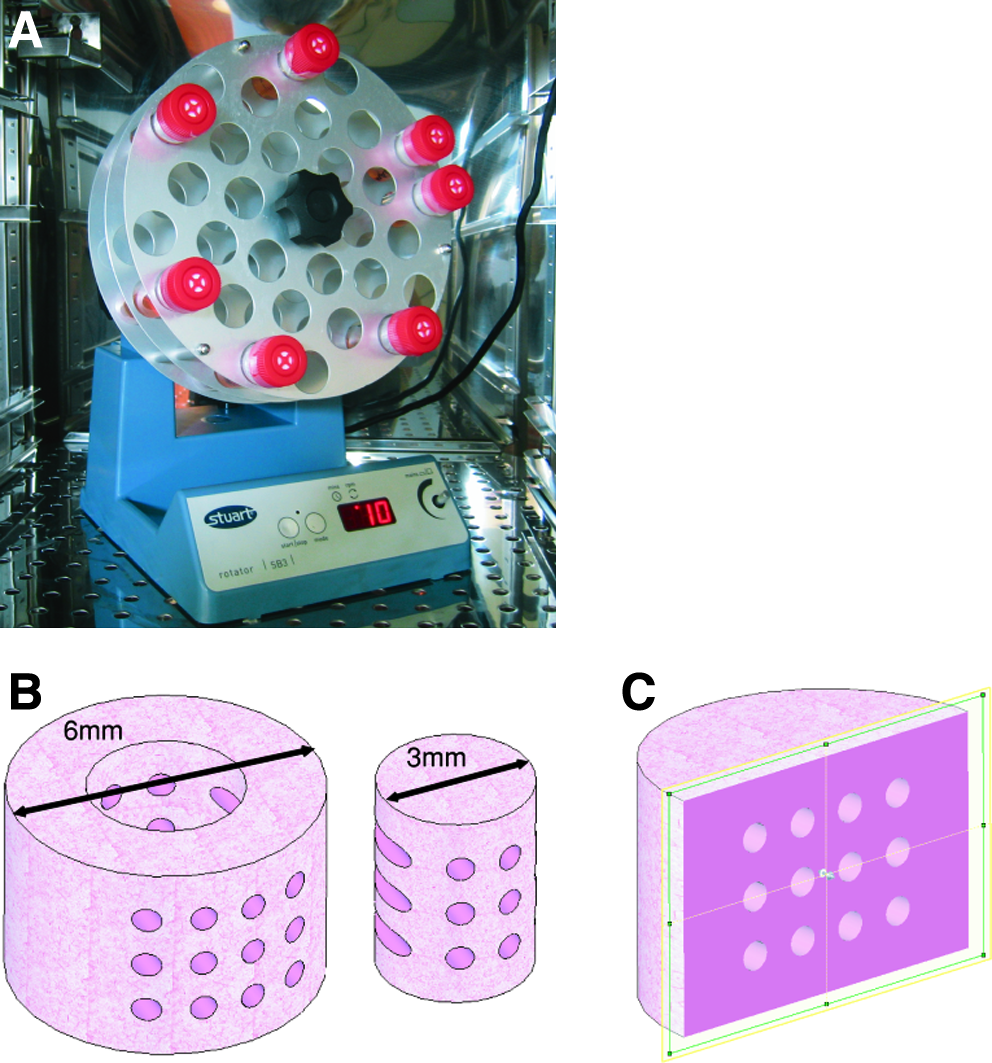
Micro-channeled construct cylinders were fabricated via a molding process (Fig. 1) utilizing a pillared array polydimethylsiloxane structure (fabricated through indirect casting of a computer numeric control (CNC) machined mold) to create the architecture (Fig. 1D). Extraction of the mold from the cast agarose–cell hydrogel produced a unidirectional channeled array in the transverse direction with diameters of 500 μm and a center–center spacing of 1 mm. All chondrocyte-seeded agarose constructs were allowed to equilibrate for 3 days before experiments were initiated (referred to as day 0). Constructs were either subjected to constant rotation (10 rpm) in a rotational culturing system or left in free-swelling conditions. For the dynamic rotational culturing regime, constructs were placed into the bottom of 30 mL polypropylene tubes, of length 107 mm and diameter 25 mm (Sarstedt LTD, Wexford, Ireland) with 15 mL of supplemented medium. The 30-mL tubes were capped with tissue flask filter caps and inserted into a rotator wheel (Stuart® Rotator SB3; Lennox Laboratory Supplies, Dublin, Ireland) inclined at an angle of approximately 15° (Fig. 2A). Each tube contained two hydrogel constructs, and rotation was performed clockwise around the central axis of the rotator wheel with a rotation radius of 10 cm. The rotation speed of the system was set to 10 rpm and maintained in a humidified atmosphere at 37°C, 5% CO2. In all groups, supplemented medium consisting of DMEM/F-12 (Sigma–Aldrich), 10% v/v fetal bovine serum, 100 U/mL penicillin/streptomycin (GIBCO–Biosciences), 1 μg/mL insulin, and 50 μg/mL ascorbic (Sigma–Aldrich) was replaced at a rate of 50% three times per week. Nonchanneled agarose constructs were also cultured in parallel under both culturing regimes for comparison purposes. Both construct types were assessed at 0, 14, and 28 days.
(
(
Mechanical testing
Constructs were mechanically tested in unconfined compression using a standard materials testing machine with a 5-N load cell (Zwick Z005, Roell, Germany). Agarose constructs were kept hydrated through immersion in a saline (0.9%) bath maintained at room temperature. A preload of 0.01 N was applied to ensure that the surfaces of the gel constructs were in direct contact with the impermeable loading platens. Stress relaxation tests were performed, consisting of a ramp and hold cycle with a ramp displacement of 0.001 mm/s until 10% strain was obtained and maintained until equilibrium was reached (∼30 min). The compressive equilibrium modulus was calculated by application of the standard uniaxial stress–strain relationship whereby the stress determined at full relaxation is divided by the applied strain. Dynamic tests were performed immediately after the stress relaxation cycle. The strain was maintained at 10% for the start of the dynamic test. A cyclic strain of 1% was applied for 10 cycles at 0.1 and 1 Hz. Dynamic moduli at each frequency were calculated through the ratio of the determined stress amplitude and the applied strain amplitude. After mechanical testing, constructs were weighed wet and the total mass recorded. Constructs were cored using a 3-mm biopsy punch and separated from the annulus (Fig. 2B); the wet mass of both the core and annulus was recorded and frozen for subsequent biochemical analyses. At each time point, one sample was fixed for histological analysis.
Biochemical analysis
The biochemical content of cultured agarose constructs was assessed at each time point. Annuli and inner cores were digested separately with papain (125 μg/mL) in 0.1 M sodium acetate, 5 mM cysteine HCl, and 0.05 M ethylenediaminetetraacetic acid, pH 6.0 (all from Sigma–Aldrich), at 60°C under constant rotation for 18 h. The proteoglycan content was estimated by quantifying the amount of sulfated GAG in constructs using the dimethylmethylene blue dye-binding assay (Blyscan; Biocolor, Co Antrim, Northern Ireland), with a chondroitin sulfate standard normalized to the tissue wet weight.
Histology
Samples for histology were fixed in formalin and dehydrated in a graded series of ethanol, embedded in paraffin wax, sectioned to 10 μm, and affixed to microscope slides. Sections were stained with Safranin-O to view proteoglycan distribution. Stained specimens were imaged with a color CCD camera and an inverted microscope. Dimensional characterization of microchannel diameters was performed using Scion™ image analysis software (Scion, Frederick, MD).
Statistical analysis
The entire experimental regime was performed once with a minimum of three replicates for both construct type and culture regime. All statistical analyses were performed using GraphPad Prism (Version 4.3) software. Numerical and graphical results are reported in the form of mean ± standard error from the mean. Groups were analyzed with one- or two-way analysis of variance with Bonferroni posttests, with culture time and culturing regime (free-swelling or dynamic rotation) as the independent variables.
Results
Acellular (2% agarose) mechanical tests were first performed to assess the influence of incorporating microchannels on the overall functional properties. It was observed that the presence of microchannels significantly reduced the equilibrium (8.50 ± 0.23 vs. 13.83 ± 0.53), 0.1 Hz (28.37 ± 0.36 vs. 43.76 ± 0.21), and 1 Hz (30.49 ± 0.31 vs. 44.42 ± 0.14) dynamic moduli, when compared to solid constructs.
Since the introduction of microchannels was observed to reduce the global functional mechanical properties, all mechanical properties presented from the cell culture experiments were normalized to those obtained at day 0 to gain an appreciation of the relative increases (see Fig. 3). At day 14, no statistical difference was observed between free-swelling and rotational culture regimes within individual groups (i.e., solid and microchannel architectures) for equilibrium modulus, 0.1 Hz, and 1 Hz dynamic moduli. However, solid free-swelling constructs had a greater equilibrium modulus compared to microchannel free-swelling constructs at day 14 (p < 0.01). By day 28, microchannel rotational constructs were significantly stiffer compared to microchannel constructs cultured under free-swelling conditions (p < 0.01). In addition, for 0.1 Hz dynamic modulus, rotational microchannel constructs exhibited greater increases compared to all other groups (p < 0.05). No difference was found in mechanical properties between solid constructs cultured under free-swelling or rotational conditions by day 28.
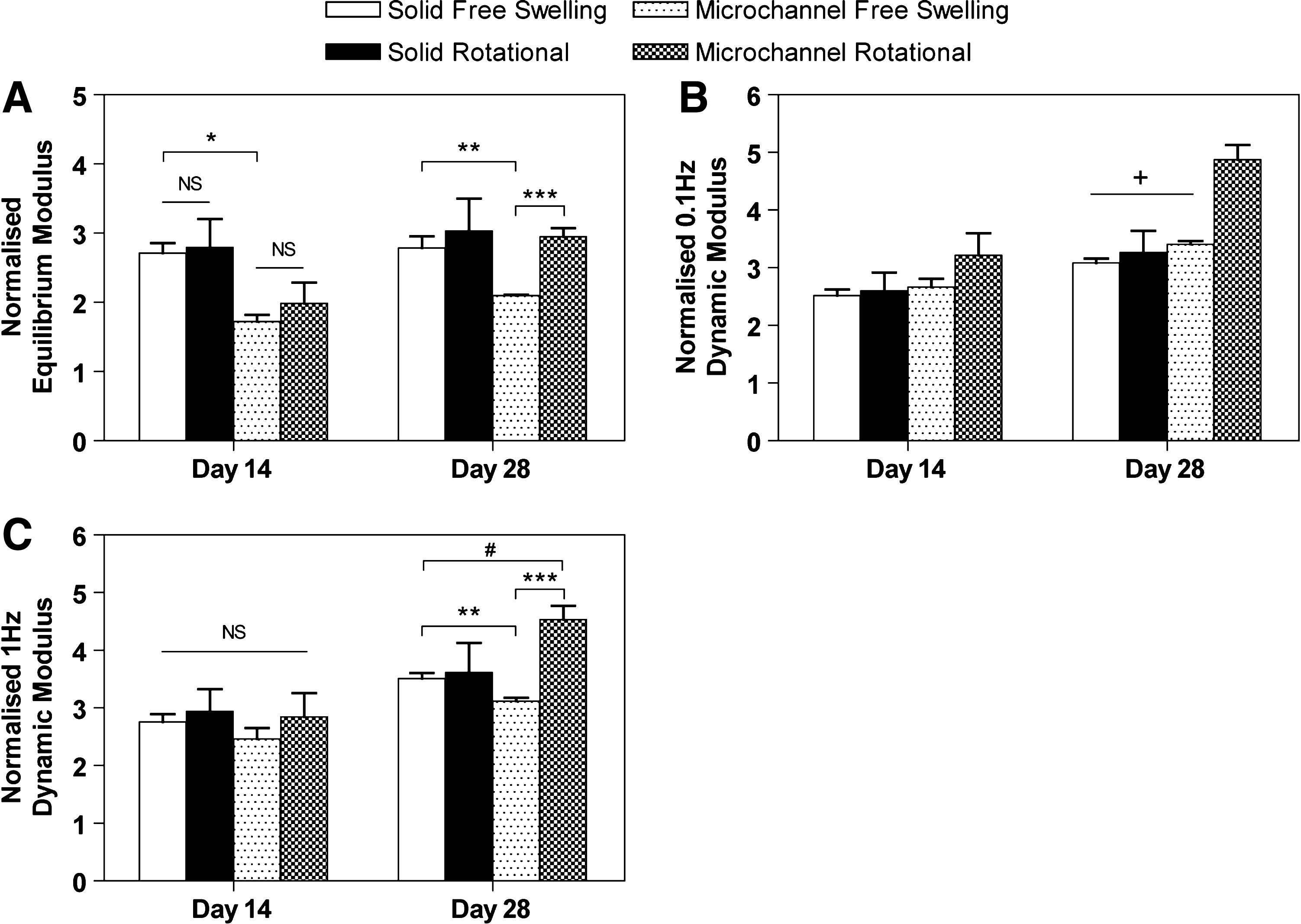
(
Under free-swelling conditions the microchannel constructs did not demonstrate enhanced GAG accumulation within the core region compared to solid constructs (Fig. 4A). The annular regions of both construct types exhibited significantly greater GAG accumulation compared to the core, with no significant differences based on construct type subjected to a free-swelling culture regime. Overall, the core region GAG content (0.317 ± 0.002% w/w for microchannel, 0.401 ± 0.020% w/w for solid) was approximately half that of the respective annular regions (0.921 ± 0.061% w/w for microchannel, 1.067 ± 0.088% w/w for solid) after 28 days of free-swelling culture. Dynamically rotated constructs demonstrated superior GAG accumulation compared to free-swelling culture for both construct types. The core regional GAG content (1.165 ±0.132% w/w for microchannel and 0.653 ± 0.066% w/w for solid, day 28) was significantly greater than free-swelling controls (p < 0.05), although GAG content in the annulus also increased (1.635 ± 0.133% w/w for microchannel and 1.229 ± 0.074% w/w for solid, day 28). In general, the greatest increases in GAG production due to rotational culture were observed in the microchannel constructs. Histological results confirmed that dynamic rotational culturing enhanced GAG synthesis and accumulation within the central regions of constructs compared to free-swelling cultures after 14 and 28 days (Fig. 5). Predominant staining was observed around the periphery of the microchannels. No significant qualitative difference was observed in the periphery between microchannel and solid constructs for both solid free-swelling and rotational cultures. In addition, intense staining was not observed in the core regions of solid constructs, unlike that observed around the periphery of the unidirectional channels of microchannel constructs. Figure 5L represents a microchannel close to the edge of a dynamically rotated construct. Here it can be observed that there is more a diffuse pattern of GAG staining, presumably due to its closer proximity to the construct edge.
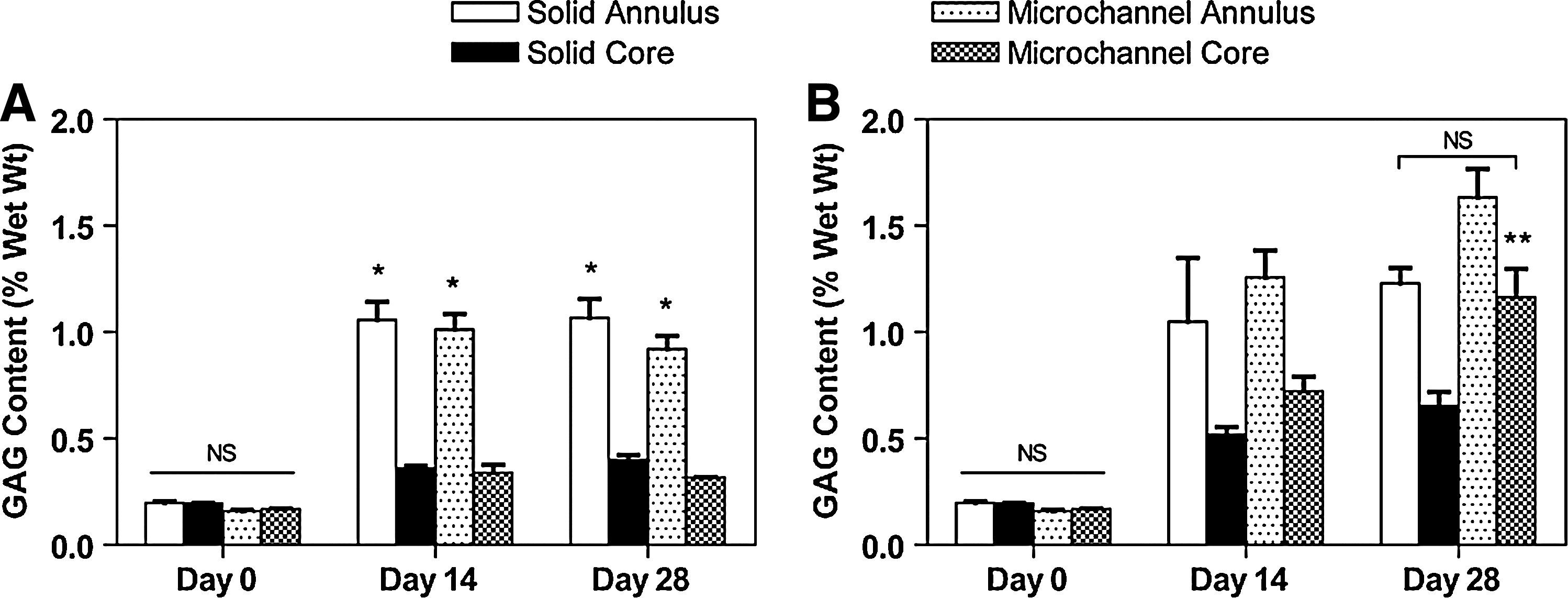
Glycosaminoglycan (GAG) content. (
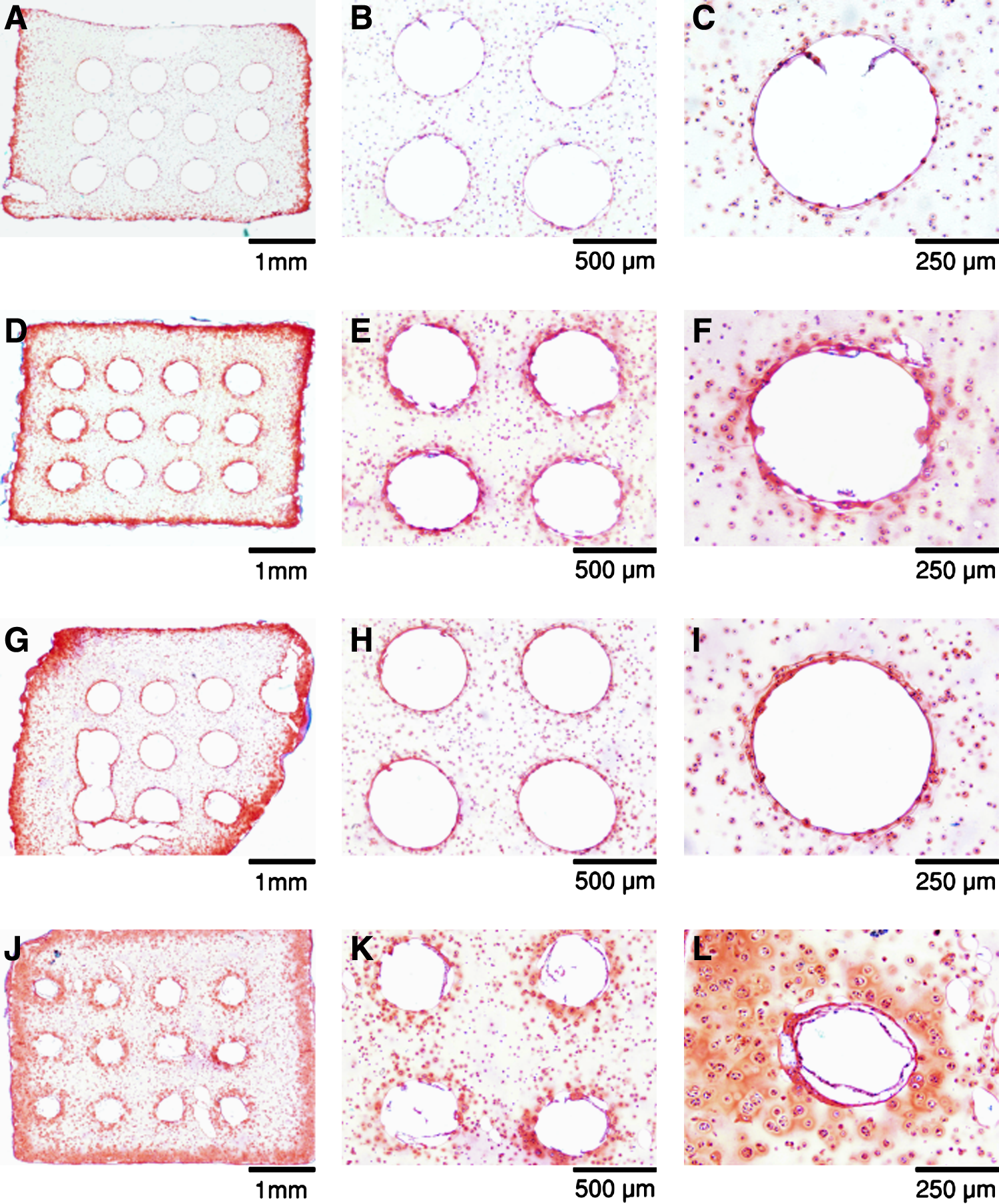
Safranin O histological sections of glycosaminoglycan distribution for free-swelling and dynamically rotated microchannel constructs after 14 and 28 days. (
A significant GAG layer was observed on the internal walls of the microchannels (Fig. 5). In addition, for dynamically rotated microchannel constructs, this increase in GAG accumulation appeared to reduce the overall dimensions of the microchannels with increasing culture periods. From image analysis, at day 0 the diameter of microchannels was calculated to be 490.33 ± 4.34 μm, decreasing to 459.05 ± 14.65 μm by day 14 and 380.20 ± 5.37 μm by day 28.
Discussion
The present study investigates the synergistic effects of dynamic rotational culturing and hydrogel architecture on the in vitro development of engineered cartilage tissue. The motivation behind this study was based upon the consistent literature reports of peripheral tissue formation,4–6 and the assumption that this was due to insufficient nutrient delivery and metabolic waste removal. 7 In an attempt to address this issue, we incorporated unidirectional microchannels (in the transverse plane) into a commonly employed chondrocyte-encapsulated agarose hydrogel system. To this end, we developed a novel micro-molding technique that permitted the creation of a microchannel array (500 μm diameter, center–center spacing of 1 mm). Although this work utilized agarose hydrogel, the technique could also be adapted for use with other hydrogels. It was initially hypothesized that these free channels would permit greater nutrient delivery and metabolic waste removal from the central regions of constructs. However, under free-swelling conditions it was observed that the incorporation of microchannels did not significantly enhance GAG matrix synthesis. This result suggests that changes to the physical architecture of a scaffold are not sufficient to overcome nutrient delivery limitations in free-swelling culture, particularly for hydrogels such as agarose where the inherent diffusivity of key nutrients is already relatively high.37,38 The diffusion coefficient for key nutrients within 2% agarose is of a similar order of magnitude as that in the medium, and hence a microchannel construct in static conditions will not improve the nutrient supply. A different result may have been obtained had we utilized a material with a lower diffusivity. It would appear that cellular consumption is primarily responsible for creating gradients of key biomolecules such as oxygen through the hydrogel in this study, and in the case of channeled constructs, along the length of the microchannels. This leads to lower core GAG production in both microchannel and solid constructs. This result will depend on the choice of scaffold material, which can influence cellular metabolism leading to changes in the consumption rates of key molecules such as oxygen. 38 This may explain why increasing the thickness of chondrocyte-seeded poly(ethylene oxide) hydrogels has been shown not to affect the biochemical composition of the construct, 39 albeit with lower levels of GAG production as observed in this and other studies (e.g., Ref. 40 ).
As chondrocytes synthesize specific cartilage matrix molecules (e.g., proteoglycan and collagen), the effective diffusivity of the constructs to molecules of different sizes will decrease,36,37 leading to gradients because of both cell consumption and lower construct diffusivity. It has been shown that growth of chondrocytes on polyglycolic acid scaffolds plateaus after 3–4 weeks, due in part to decreases in the diffusivity of the construct as new tissue is generated. 41 Therefore, had the culture period been extended beyond 28 days, or had more rapid matrix synthesis occurred, enhanced GAG synthesis may have been observed in microchannel constructs in free-swelling culture by increasing the effective diffusivity of the scaffold; however, concentration gradients because of cellular consumption would still exist.
Dynamic rotational culturing proved to enhance GAG synthesis in both solid and microchannel constructs. Higher levels and more homogeneous distribution of GAG were observed throughout the depth and width of microchannel constructs compared to solid constructs when dynamically cultured. The accumulated GAG content (% w/w) within the core of microchannel constructs was equivalent to that in the annular regions of the solid constructs at day 28. This is most likely due to enhanced delivery or supply of key nutrients such as oxygen and glucose and more effective metabolic waste removal of by-products such as lactate from the interior regions of the microchannel constructs via forced convection. In addition, the rotational culturing regime provides for a well- mixed medium environment, thereby altering the boundary conditions and limiting large gradients from developing due to cellular consumption. Measurement and modeling of the transport of these key molecules will need to be undertaken in the future to verify this. The higher GAG values in the cores of microchannel constructs compared to solid constructs suggests that the forced convection alone may not guarantee homogenous construct formation, and that optimization of the architecture in addition to the flow environment must also be made when engineering large cartilaginous constructs. In addition, as revealed through histological analysis, superior GAG penetration depths (∼350 μm) exist at the peripheral fluid–structure interface for dynamically rotated constructs compared to free-swelling constructs (Fig. 5J). On inspection of centrally located microchannels from the dynamic rotation culture regime (Fig. 5K), it is clear that the GAG penetration depths (∼100 μm from the microchannel–gel interface) are not as pronounced. This further supports the conclusion that optimization of the rotational culturing regime or scaffold architecture is required to recapitulate the superior exchange rate environment that clearly exists at the boundary interface of the constructs. Application of a direct perfusion system may ensure a more uniform flux through the microchannel array, with comparable exchange rates inside and outside the hydrogel. Such an enhancement of the dynamic culture regime would most likely result in higher GAG levels within the agarose gel interspaces, thereby creating a more functional cartilaginous tissue.
In addition to enhancing nutrient transfer, rotational culture also induces a fluid shear stress along the surface of the scaffold. In the case of a microchannel construct, the total surface area subjected to shear stress is obviously increased, although the magnitude will vary with position on the surface. Whether this stimulus, in combination with enhanced mass transfer, contributes to higher levels of GAG accumulation is debatable. Although low levels of shear stress may be beneficial, 42 the observation that rotating wall vessel bioreactors, which provide efficient mixing but minimal shear stress, enhance chondrogenesis as compared to mixed cultures with high shear, 43 has led to the suggestion that the shear stress not only has less of an effect than transport, but may also actually be detrimental.37,42 This view is supported by our observation that cells near the surface of hydrogels subjected to rotational culture took on a more fibroblastic morphology. Definitive conclusions, however, cannot be reached in this study as the flow conditions and associated fluid shear stresses generated in the bioreactor system cannot yet be accurately quantified. Theoretical models of tissue differentiation would also suggest that high magnitudes of fluid flow can lead to fibrous tissue formation.44,45 Optimizing both the nutrient and biophysical environment is a key challenge in tissue engineering.
The presence of microchannels within an agarose hydrogel reduces the apparent modulus values obtained from mechanical testing. This complicates our understanding of developing structure–function relationships within such constructs. To clarify matters, the mechanical properties of acellular hydrogels, both solid and channeled, fabricated using either 2% or 4% agarose were compared (unpublished results). The Young's modulus of acellular solid agarose hydrogels was found to increase ∼3.5-fold by increasing the agarose concentration from 2% to 4%, while the dynamic modulus increased ∼2.7-fold. For the microchannel constructs, the Young's modulus increased ∼4.4-fold, while the dynamic modulus increased ∼3.2-fold. The greater increase observed in the microchanneled hydrogels implies that the channels have less of an effect on the apparent mechanical properties of the structure as the inherent properties of the material (or tissue) increase. Given that greater increases were observed in the dynamic properties of the microchannel constructs compared to solid constructs after 4 weeks in rotational culture suggests that the inherent properties of the cartilaginous tissues forming inside the channeled construct are greater than that in the solid construct (Fig. 3). The higher GAG content within the microchanneled construct is presumably contributing to this finding (Fig. 4).
The microchannels were observed to close in or narrow in diameter with increasing time in culture due presumably to matrix synthesis and tissue swelling associated with the fixed charge density of the accumulated GAG. For longer culture periods, it is expected that these channels will completely fill in, at which point they will obviously have no further value in enhancing nutrient transfer within the engineered tissue. This is of course a desired attribute as, ideally, the channels will be entirely or very nearly closed before implantation. This eventuality could therefore be viewed as a simple measure for when in vitro culture should cease, as further culture could lead to cell death caused by nutrient transfer limitations. Future studies will therefore focus on altering both the size and number of channels, in combination with changes to the culture time and environment, to engineer tissues with controlled properties. In addition, changes in the diffusivity and permeability of the tissue at different points in the culture period will be assessed. In conclusion, this paper demonstrates a methodology to engineer large cartilaginous tissues using feasible numbers of cells (15 million cells/mL) derived from skeletally mature tissue that have first undergone monolayer expansion to better mimic the expected clinical scenario. The use of a hydrogel overcomes the problem of obtaining an efficient and homogenous cell-seeded construct before culture. This approach could also be used with anatomical molds to engineer tissues of more complex geometries.46,47 It is also possible that this culturing system could be extended to engineer different tissue types of clinically relevant dimensions other than cartilage tissue.
Footnotes
Acknowledgments
Funding was provided by Science Foundation Ireland (07-RFP-ENMF142) and Enterprise Ireland (PC/2006/384).
Disclosure Statement
No competing financial interests exist.