
Abstract
The liver and other organs are connected to each other through the bloodstream. Therefore, the connection between tissues is generally mediated by soluble molecules able to cross the endothelial wall of capillaries. We developed a multicompartmental device, multicompartmental bioreactor (MCB), designed to mimic the connection between different tissues in which crosstalk is mediated by soluble molecules transported through the blood. A comparative study of the crosstalk between hepatocytes (HepG2) and endothelial cells (human umbilical vein endothelial cells) in connected culture in the MCB and in a traditional static coculture system was performed by analyzing glucose consumption and secretion of albumin, urea, and nitric oxide. When hepatocytes and endothelial cells were cultured together, the production of albumin and urea increased, and the increase was higher in the MCB than in traditional static coculture. In spite of this enhanced metabolic activity, the crosstalk between hepatocytes and endothelial cell leads to decreased glucose consumption with respect to hepatocytes alone, both in static and in dynamic conditions. However, the dynamic connected culture has a higher rate of metabolite synthesis and secretion with respect to cocultures. This means a more efficient use of energetic substrates and enhanced hepatocyte function in the MCB.
Introduction
To address this gap, we developed a multicompartmental bioreactor (MCB) that represents a simplified downscaled in vitro human body. Our aim was to better understand the interaction between different tissues, with particular reference to the metabolic system.
The device was designed using biomedical engineering concepts based on biomimetics, as described elsewhere. 9 It is composed of cell culture chambers connected in series and in parallel to form a closed loop. The cell culture chambers are connected to each other through the flow of the culture medium that acts as the bloodstream, transporting molecules to and from the compartments. In the MCB, each chamber can be addressed and interrogated separately, and different cell types can be added stepwise to the system.
In a previous report 10 we described the design of the system through allometric scaling, focusing on glucose processing for its relevance to diabetes and metabolic disorders. We also described a connected culture of murine hepatocytes and human umbilical vein endothelial cells (HUVEC) to prove the concept of the system. In this work we compare a connected culture system with HepG2 and HUVEC to classic coculture. Our hypothesis is that in the dynamic connected culture the crosstalk between the two different cell types will be enhanced with respect to traditional static coculture. Thanks to the flow-mediated connection with endothelial cells, HepG2 will display higher levels of hepato-specific function. For this purpose, albumin secretion, urea synthesis, and glucose consumption were assessed in dynamic connected culture and in static coculture. Nitric oxide was also measured to evaluate if the presence of hepatocytes in connected culture influences endothelial response to flow.
Materials and Methods
Design and fabrication of the MCB device
The MCB system was developed taking into account the relationship between different parameters such as metabolism, time and volume distribution, and organ dimensions, to scale the physiological interactions present in human body. 11 Using allometric laws, times, lengths, and cell numbers were downscaled as reported in our previous work. 10
The bioreactor was designed using Comsol Multiphysics (Comsol AB, Stockholm, Sweden) to ensure laminar flow in central region of the cell culture compartment. At the flow rate used, 175 μL/min, the wall shear stress in the hepatic compartment was estimated to be in the range of 1–5 mPa. The MCB chamber was fabricated as described elsewhere.9,10 Briefly, polydimethylsiloxane (Sylgard 184®; Dow Corning, Midland, MI) was cast and cured in a mold (fabricated using stereolithography) by heating at 65°C and then carefully removed, washed, and sterilized with H2O2 gas plasma. After placing the cells in the compartments, the polydimethylsiloxane base was covered with a plexiglass plate, and the two parts were clamped together by a Teflon frame tightened with screws. The output tube was connected to a peristaltic pump (Instec P720, Boulder, CO), to a reservoir chamber, and finally to the input tube of the bioreactor. The total volume of the four-chamber system, including tubing and medium reservoir, was 30 mL. Figure 1 shows the bioreactor setup as well as a connection and flow scheme of the MCB system.
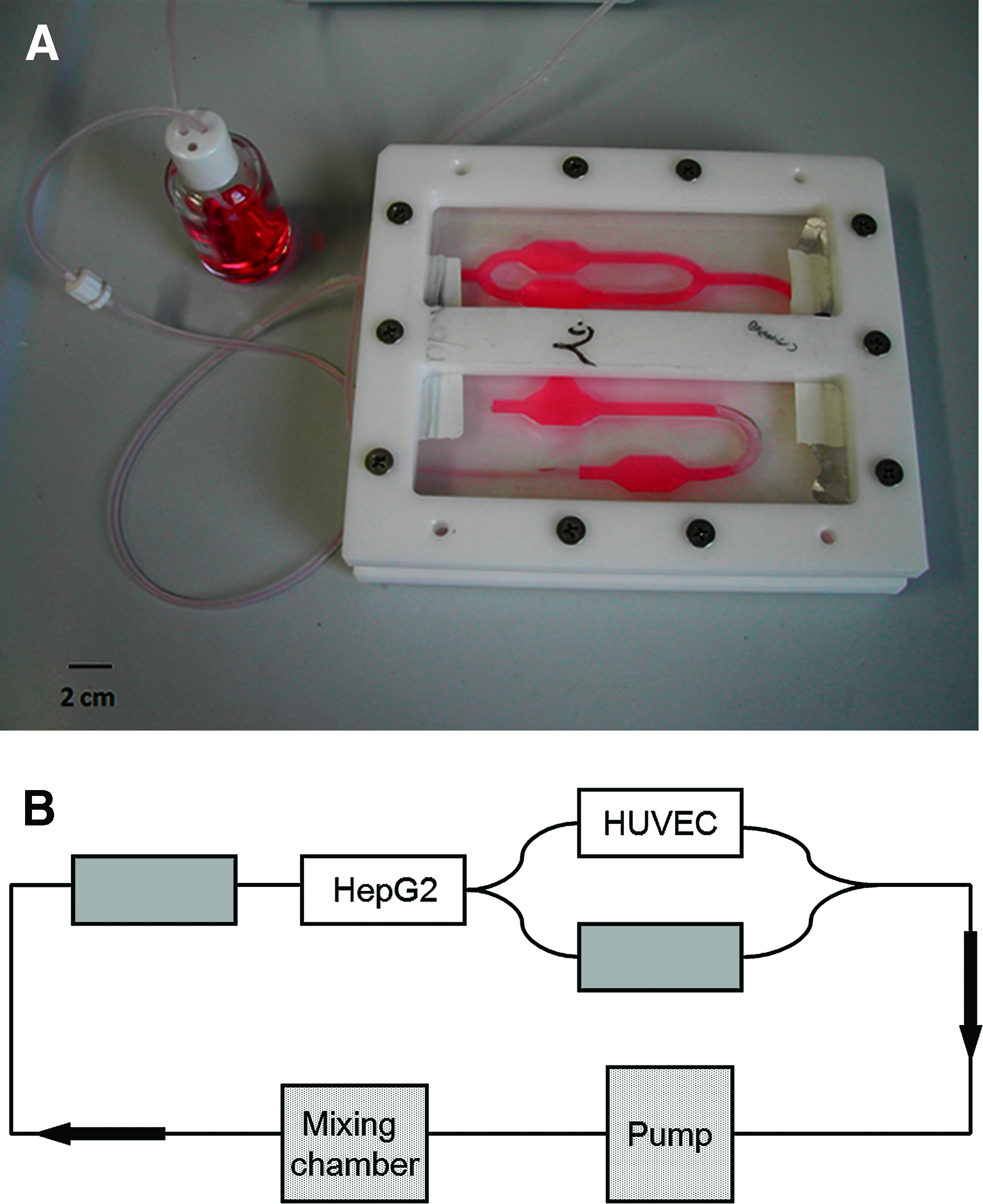
(
Cell culture supplies
Trypsin, ethylenediaminetetraacetic acid (EDTA), phosphate-buffered saline (PBS), cell culture medium M199, Eagle's minimal essential medium (MEM), fetal bovine serum (FBS), and bovine brain extract were purchased from LonzaBioscience (Basel, Switzerland). All other reagents were purchased from Sigma-Aldrich (St. Louis, MO).
HUVEC extraction
HUVEC were extracted using a trypsin-based treatment. 12 The umbilical vein was cannulated and washed with PBS. Then, it was closed at one end, filled with 0.05% trypsin with 0.02% EDTA in PBS, and left at 37°C for 15 min. The solution was recovered and centrifuged (900 rpm, 5 min), and the cell-containing pellet was resuspended in a fresh medium. The cells were counted using a Burker chamber, assessed for vitality through a trypan blue exclusion assay (routinely greater than 90%), and seeded for amplification in a 25 cm2 flask precoated with 0.5% gelatin. To maintain and grow the cells, M199 was supplemented with 10% FBS, 100 U/mL penicillin, 100 μg/mL streptomycin, 1% glutamine, and 1% bovine brain extract; this is referred to as complete M199.
Cell culture
The human hepato-blastoma cell line HepG2 was used as an alternative to primary human hepatocytes because, by virtue of its immortality, it is possible to make standard comparisons that remain valid over time and for all experiments. HepG2 cells maintain the main synthetic and endogenous functions of primary hepatocytes, as well as some exogenous metabolic functions of primary hepatocytes. 13 The cell culture medium, referred to as complete MEM, was MEM (glucose 1 g/L) supplemented with 5% FBS, 100 U/mL penicillin, 100 μg/mL streptomycin, 1% glutamine, 2% MEM nonessential amino acids, and 2% vitamins. Hepatocytes were coated as described below.
HepG2 were cultured in 75 cm2 flasks with complete MEM. For their use in static and dynamic experiments, they were detached with 0.05% trypsin with 0.02% EDTA, and seeded on glass slides of 12 mm diameter placed in 24-multiwell plate. HepG2 were seeded at a density of 5 × 104 per well for static monoculture tests, and at a density of 8 × 104 per well for static coculture and dynamic experiments.
When at confluence, HUVEC were detached with trypsin and seeded on glass slides pretreated with 0.5% gelatin at a density of 5 × 104 per well for static monoculture tests, at a density of 8 × 103 per well for static coculture experiments, and at a density of 2.4 × 105 per well for dynamic experiments. For the static coculture experiments, HUVEC and HepG2 were suspended together and seeded onto the slides. The HUVEC:HepG2 seeding ratio was 8 × 103:8 × 104. The 1:10 ratio used mimics the ratio between endothelial cells and hepatocytes in the human liver where the two types of cells have homotypic and heterotypic connections. 14 In the dynamic connected culture the ratio between HUVEC and HepG2 was 3:1 (2.4 × 105:8 × 104) to represent the anatomical ratio between liver compartment and endothelial compartment in the whole body.
HepG2 coating
HepG2 in all experiments were coated with an alginate gel. This gel was obtained by crosslinking alginate and CaCl2 at room temperature for 1 min. Different concentrations of alginate and CaCl2 were tested to obtain a thin coating that was resistant to the shear stress generated in the system by flow. Alginate was tested at concentrations of 1%, 1.5%, 2%, and 3%, while CaCl2 was used at concentrations of 0.1 and 0.05 M. Sodium alginate was prepared in a solution of serum-free medium and diluted 1:1 with PBS. CaCl2 solutions were prepared using cell culture medium directly. The use of the culture medium ensures that the pH remains constant during the crosslinking of alginate gel. The alginate gel was crosslinked directly on the surface of HepG2 seeded on glass slides placed in the bioreactor or in a 24-multiwell plate using 400 μL of alginate and 250 μL of CaCl2. After waiting 1 min for the gel to crosslink, the excess of CaCl2 was removed, the bioreactor closed, and dynamic experiments run for 24 h at a flow rate of 175 μL/min. After this period, the cells and the coating were examined under a microscope.
Culture medium tests
Three different culture media for HUVEC and HepG2 were tested in static and dynamic conditions to find a common culture medium to be used in the connected culture. The two cell types were tested with complete MEM, complete M199, and M199 supplemented with the same cocktail as complete MEM (5% FBS, 100 U/mL penicillin, 100 μg/mL streptomycin, 1% glutamine, nonessential amino acids, and vitamins), referred to as M199-2. Cells were seeded directly in the wells and on glass slides to be placed in the MCB device. Experiments were carried out for 24 h, at the end of which cells were tested for vitality and morphology.
Biochemical and morphological analysis
Vitality, viability, morphology, and metabolic activity were evaluated in the dynamic and static cell cultures. Metabolic activity was assessed analyzing glucose consumption and albumin and urea secretion. HUVEC were also assayed for nitric oxide production.
Vitality and cell counting
Vitality was assessed by trypan blue exclusion assay and cell counting with a Burker chamber. Cells were detached by trypsin/EDTA solution, and 50 μL of cell suspension was mixed with an equal volume of trypan blue solution and counted. Both vital and dead cells were counted.
Cell viability
Cell viability was assessed by the MTT assay. It uses a yellow compound that is oxidized by mitochondrial enzymes to a purple compound soluble in dimethyl sulfoxide. The cell suspension was centrifuged at 900 rpm for 5 min, and the pellet was resuspended in 200 μL of fresh medium and placed in 96-multiwell plate. Twenty microliters of MTT solution 5 mg/mL was added to the cell suspension. After 12 h, medium was removed, the pellet suspended in 200 μL dimethyl sulfoxide, and the absorbance measured at 540 nm.
Biochemical analyses
Metabolite concentrations were determined in the medium using commercial kits in accordance with the manufacturer's instructions and then converted to absolute quantities by multiplying by the total volume. Production and consumption rates are expressed as the quantity of metabolite produced or consumed over each 24-h period divided by the number of cells. In particular, the glucose concentration was assessed using the Glucose Test Kit (Megazyme International Ireland, Bray, Ireland), a colorimetric assay based on glucose-oxidase and peroxidase reactions. In all cases glucose consumption is the difference between fresh medium and sampled medium and is expressed as a daily consumption rate per cell. Human albumin concentration was evaluated by an ELISA immunochemical assay (Bethyl Laboratories, Montgomery, TX); urea and nitric oxide concentrations were measured using commercial colorimetric kits (Urea Kit; Sigma-Aldrich and Cayman Chemical [Ann Arbor, MI]).
Morphology
HUVEC were fixed with 4% formaldehyde and stained with crystal violet to highlight their typical elongated shape, while HepG2 were observed without any kind of stain. Cell morphology was analyzed using an optical microscope (AX70; Olympus Italia, Milan, Italy), with a 20 × objective.
Statistical analysis
Statistical analysis was performed using the Student's t-test when comparing between control and experimental groups in static and connected cultures and analysis of covariance (ANCOVA) for comparison of trends (Matlab Statistics Toolbox; The MathWorks, Natick, MA); a p-value of less than 0.05 was considered statistically significant.
Static baseline and coculture
The proliferation and metabolic activity of HepG2 and HUVEC were analyzed in traditional static multiwell cultures for 5 days to obtain a baseline to compare data obtained from dynamic connected cultures and to profile metabolic rates of cells over several days. Moreover, the proliferation and metabolic rate of a static coculture of HepG2 and HUVEC were also assessed. For the baseline, HepG2 and HUVEC were seeded in 24-multiwell at a density of 5 × 104 cells per well. Both complete MEM and M199 as well as M199-2 were used for the baseline cultures. For the cocultures, HepG2 and HUVEC were seeded at a density of 8 × 104 and 8 × 103, respectively, with complete MEM. Every 24 h, the medium was withdrawn for biochemical analysis, and cells were tested for proliferation and viability. All experiments were performed in triplicate.
Dynamic culture
Dynamic experiments were performed in the MCB with HepG2 or HUVEC alone and with both cell types together in the so-called the dynamic connected culture. Twenty-four hours after seeding, the 12-mm-diameter glass slide with HepG2 was inserted in the MCB liver chamber and covered with alginate gel as described previously. Then, the glass slide with HUVEC was inserted in the endothelial chamber. The bioreactor was assembled and connected to the mixing reservoir and the pump and filled with 30 mL of complete MEM. For these experiments, a flow rate of 175 μL/min was applied. Experiments were carried out for 24 h, during which 2 mL of medium was withdrawn from the outlet of the MCB for biochemical analysis at 2 h 40 min, 4 h 20 min, 6 h, and 24 h and, each time, replaced with 2 mL fresh medium. In dynamic experiments with only one type of cell, we used the same protocol and the same conditions, and either the liver or the endothelial chamber was empty of cells.
Results
HepG2 coating
Hepatocytes in the liver are not subject to direct blood flow, but O2, CO2, and metabolites are exchanged with the blood by passage through the capillary sinusoid wall. Shear stress generated by flow of the medium directly on the cell surface can damage hepatocytes. Therefore, the HepG2 cells were protected by a thin permeable alginate coating that allows metabolite and gas exchange between cells and the culture medium. This is one of the most commonly used materials to encapsulate cells.15–17 Different concentrations of sodium alginate and CaCl2 were tested to obtain a gel able to protect HepG2 from the direct effect of shear. Coated cells were cultured in multiwells or in the bioreactor for 24 h, and cellular viability and morphology were assessed. Crosslinking sodium alginate with 0.05 M CaCl2 led to the formation of gels unable to resist to the shear stress induced by flow. However, alginate concentrations between 1.5% and 2%, crosslinked with 0.1 M CaCl2 solution, formed a gel that was thin and porous enough to allow metabolite exchange and to preserve HepG2 viability and morphology, but compact enough to protect cells (Fig. 2). Moreover, using PBS as solvent for alginate and CaCl2 led to a decrease in pH that caused cell damage. The use of the medium itself as a diluent for both the CaCl2 solution and PBS-alginate solution was the best way to buffer any pH changes due to addition of CaCl2. The lowest concentration of alginate that resisted flow and maintained HepG2 integrity was 2% with 0.1 M CaCl2, and this composition was used for all further experiments.
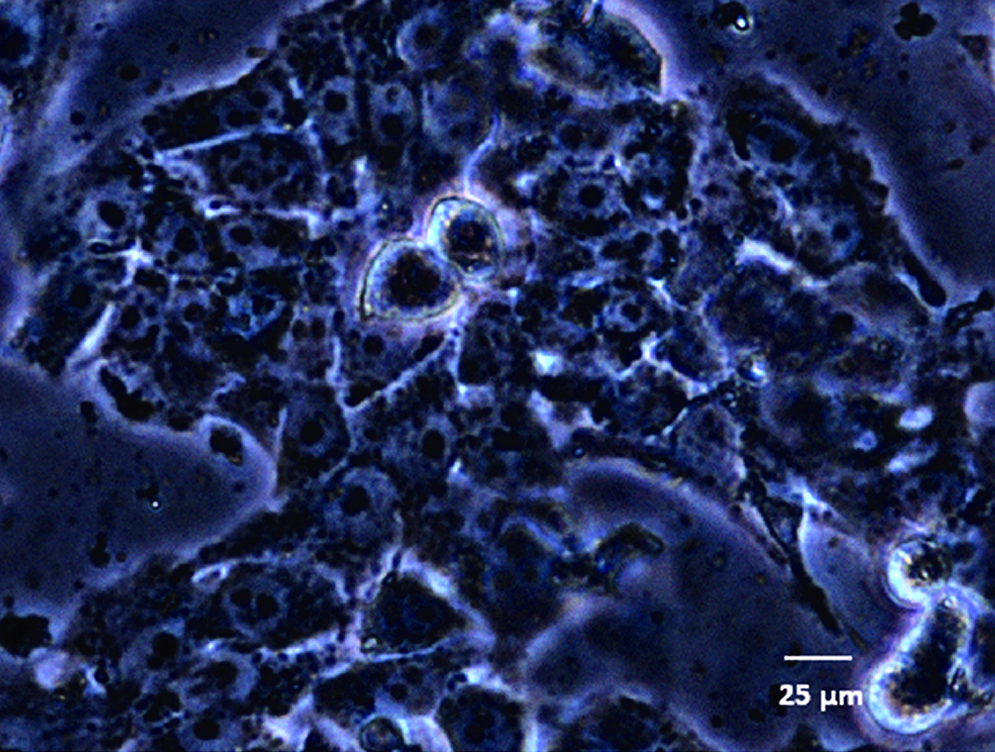
HepG2 after 24 h of dynamic culture. Cells were coated with 2% alginate and 0.1 M CaCl2. Color images available online at www.liebertonline.com/ten.
Culture medium tests
Three different cell culture media were tested on HUVEC and HepG2 separately: M199 and MEM completed with their specific supplements for HUVEC and HepG2, respectively (as described above), and the M199 medium supplemented as MEM for HepG2 (M199-2). In complete M199 medium, HUVEC adopted their typical elongated shape, while this feature is lost in M199-2 (data not shown). In complete MEM, their shape and vitality are more similar to HUVEC in complete M199. On the other hand, in both complete M199 and M199-2, HepG2 viability decreased with respect to that in complete MEM, and cells lost their phenotypic shape. In these cells, membrane integrity was compromised and the presence of little vesicles suggested the beginning of necrotic processes (Fig. 3). Therefore, complete MEM was chosen as the common medium for dynamic connected cultures and static cocultures since it preserved both hepatic and endothelial shape and viability.
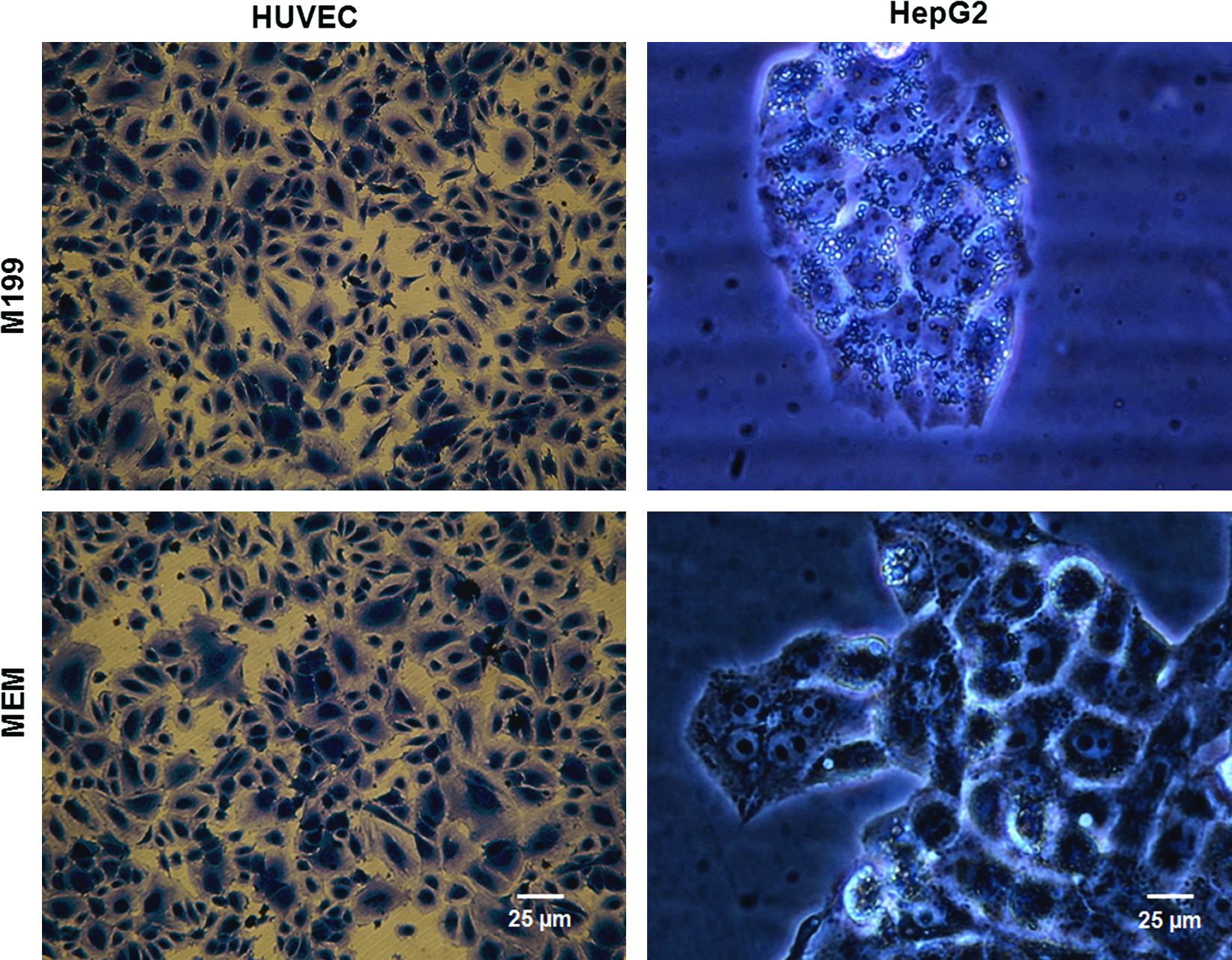
Human umbilical vein endothelial cells (HUVEC) and HepG2 cells after 24 h culture in complete M199 and MEM medium. HUVEC were stained with crystal violet dye. Color images available online at www.liebertonline.com/ten.
Static baseline and coculture
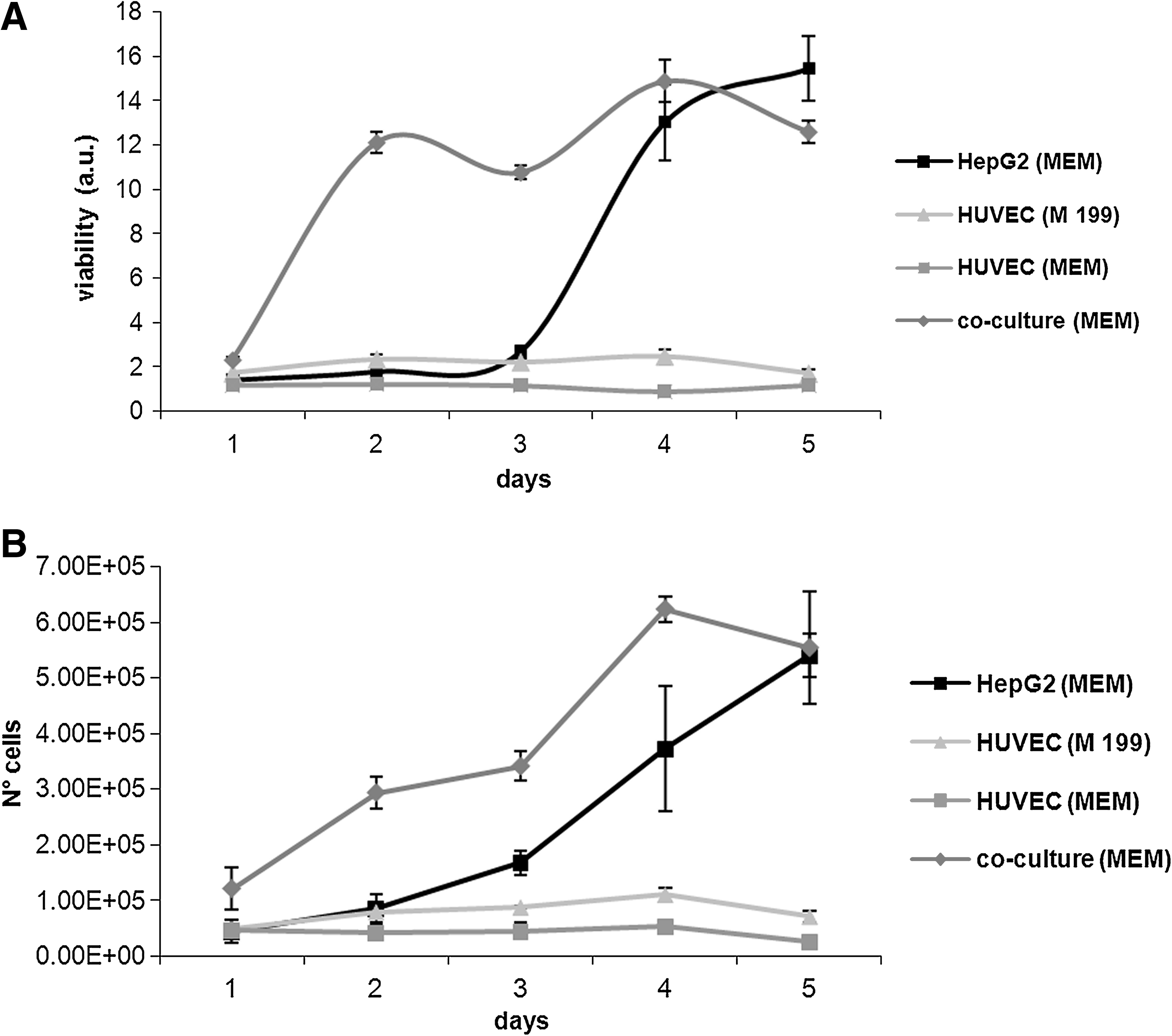
HepG2 and HUVEC were cultured in multiwells individually and in coculture for 5 days. The aim was to define a baseline for proliferation and metabolic markers such as glucose, albumin, and urea, to compare classic static culture and dynamic connected cultures in the MCB. As expected, the HepG2 proliferation rate was higher than HUVEC (Fig. 4A, B): the former is an immortalized cell line, while the latter is a primary culture. Notably, both the viability and proliferation rate increased at a much higher initial rate in the cocultures, and it is likely that the proliferation is principally due to HepG2. HUVEC were tested using both complete MEM and complete M199 to assess if the use of a nonstandard medium could compromise cell proliferation and viability over 5 days.
(
There was only a slight difference between the two media as far as proliferation was concerned, but glucose consumption rate was increased in complete MEM compared with complete M199 (5.76 ± 0.86 ng/cell/day and 3.03 ±0.86 ng/cell/day for the first 3 days, p = 0.003, ANCOVA), as shown in Figure 5. In the coculture, the glucose consumption rate was significantly lower than that of HUVEC alone in the same medium (1.86 ± 0.86 ng/cell/day compared with 5.76 ±0.86 ng/cell/day, p = 0.0001), and more similar to that of the HepG2 monoculture. These data together with the ones from the proliferation analysis suggest a more efficient use of this energetic substrate in coculture compared to the single monocultures. In coculture, in fact, while cell proliferation increased at a high initial rate, glucose consumption was constant.
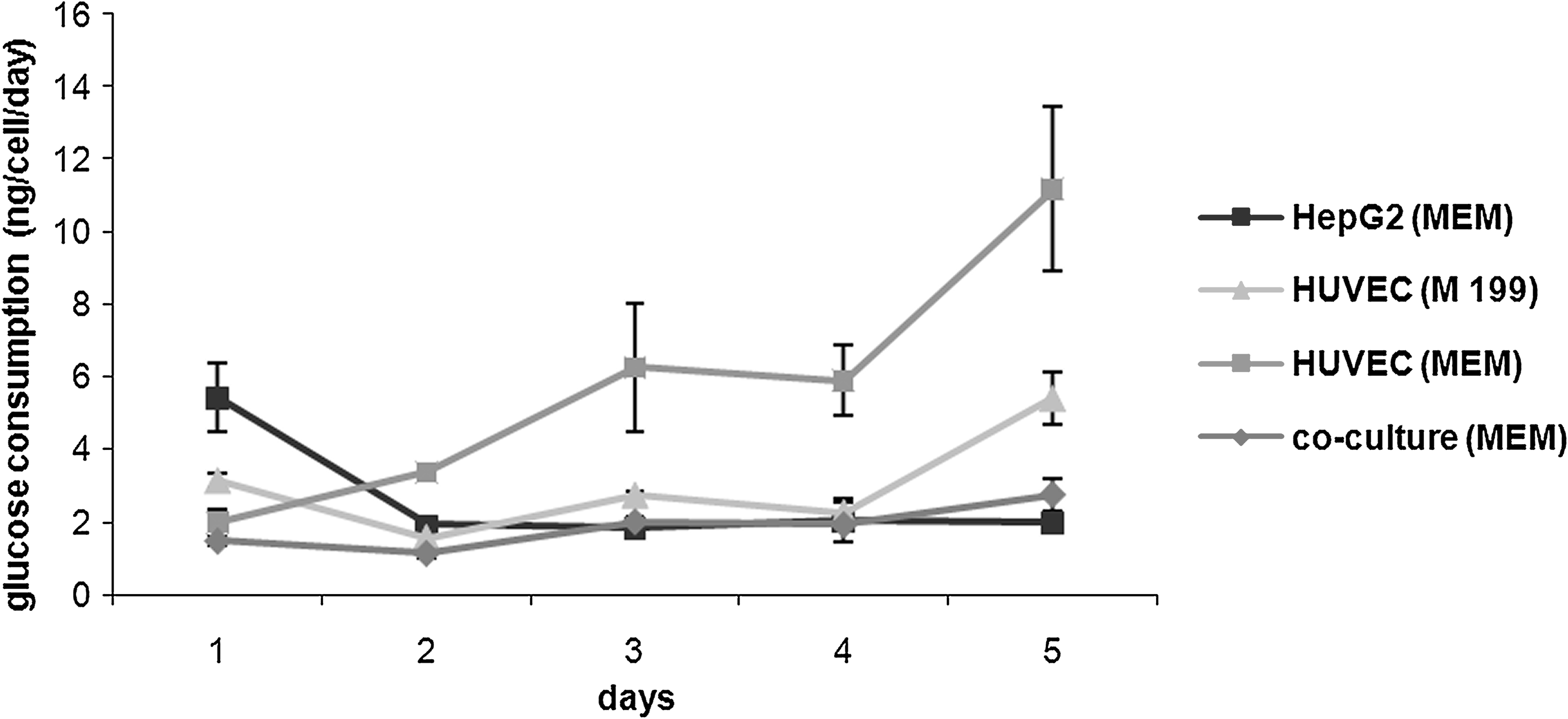
Glucose consumption rate per cell in baseline and coculture studies, n = 3.
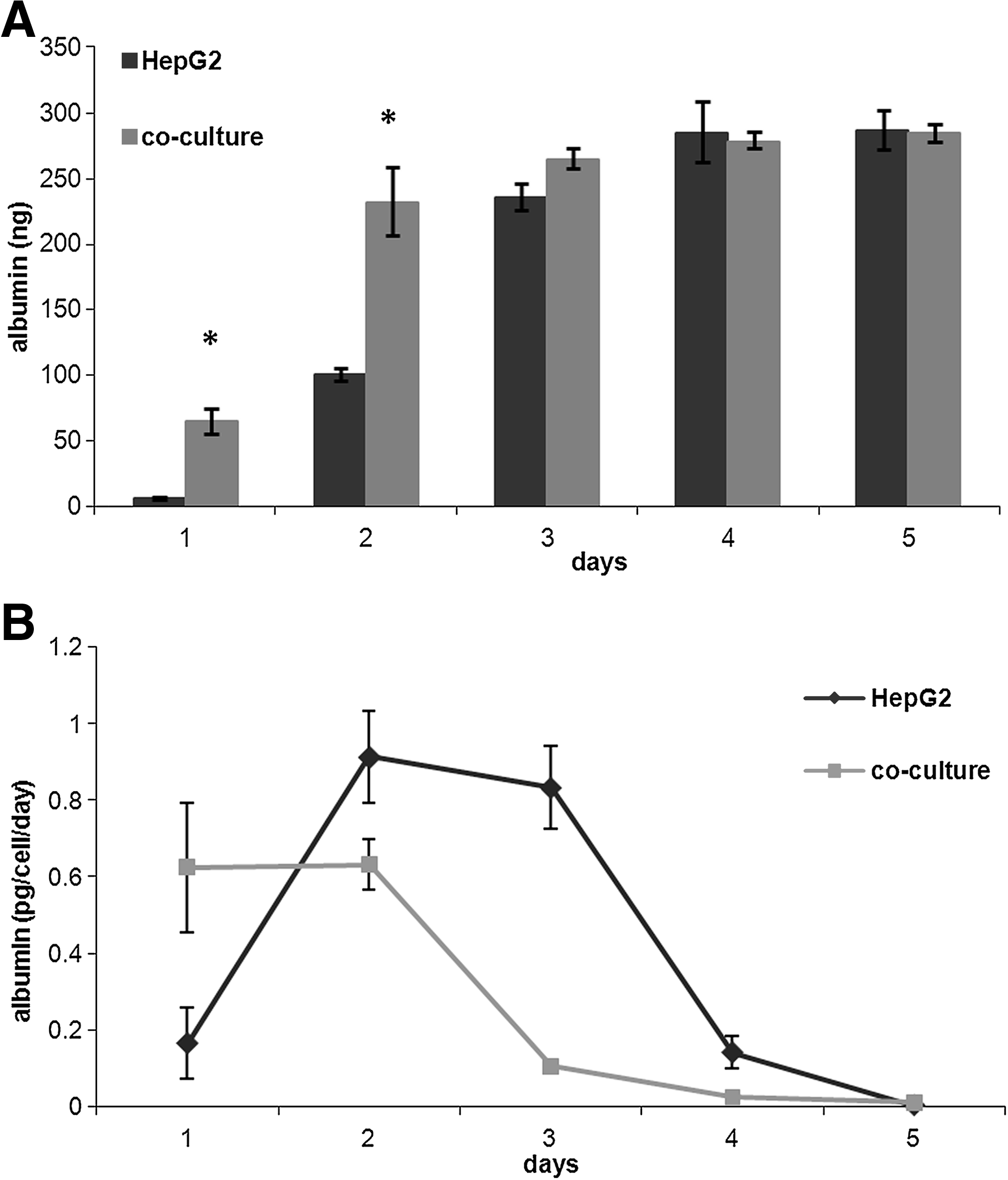
As far as albumin is concerned, both HepG2 monocultures and cocultures show an initial increase in production and then almost zero production from days 4 to 5 (Fig. 6A). Figure 6B shows the albumin production rate per cell, calculated by dividing the quantity of albumin detected by the total number of cells in culture per day. Together, the two figures show that although production rates are different, the total quantity of albumin produced over 5 days was the same for the two cultures.
(
Urea production increased gradually during the 5 days of culture (Fig. 7A), but not at the same rate as cell proliferation, so the urea production rate per cell decreased after the second day (Fig. 7B). Moreover, on the first 2 days the urea production rate was higher in the HepG2 monoculture than in coculture (monocultures 0.28 ± 0.10 ng/cell/day, cocultures 0.15 ± 0.10 ng/cell/day, p = 0.006 ANCOVA). As for albumin, there is a high initial rate of release due to the increase in cell number, after which metabolic and synthetic activity falls probably because saturation concentrations are reached or due to substrate limitations.
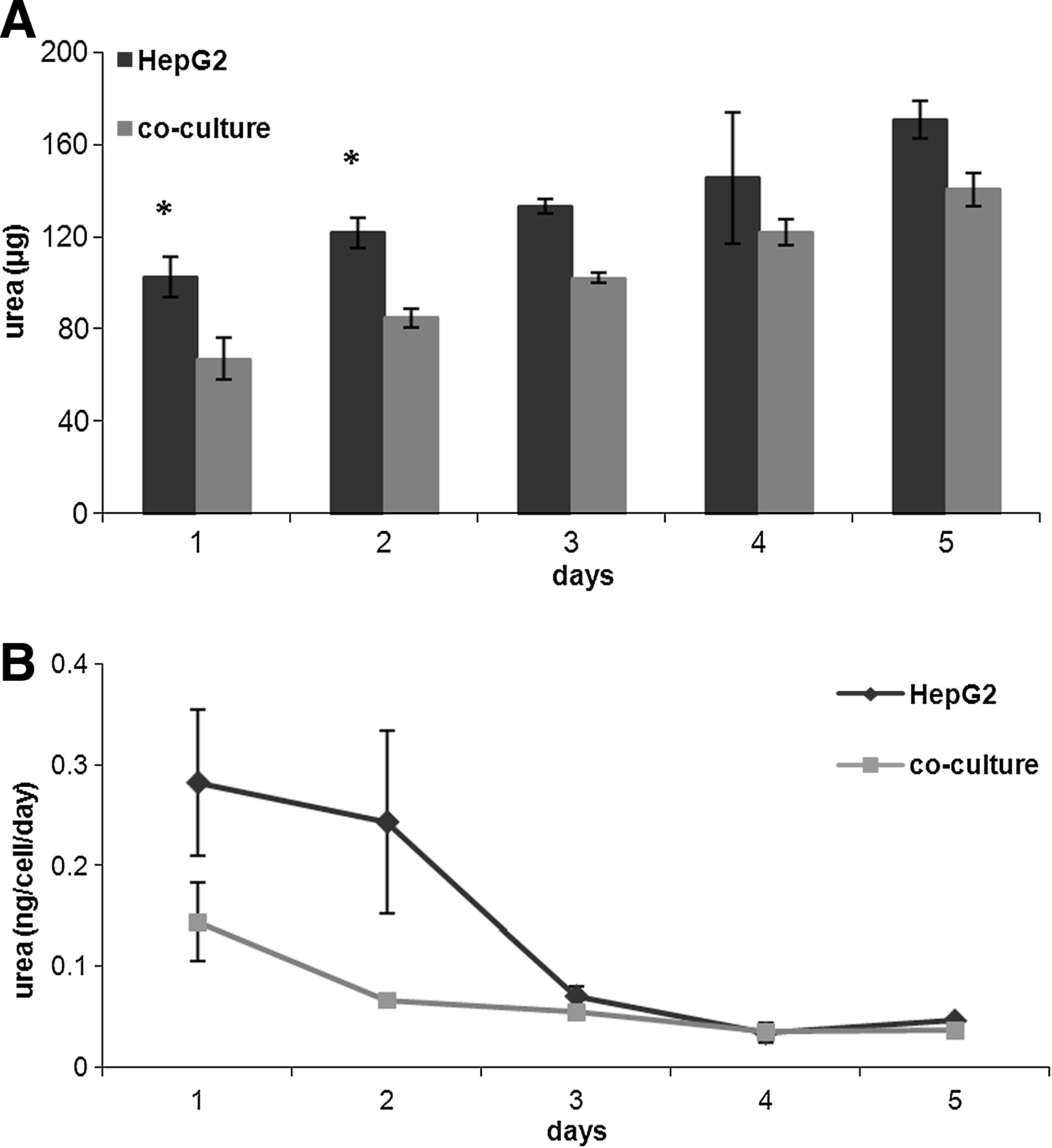
(
Dynamic culture
Having established a common medium and a suitable coating for cells, HepG2 and HUVEC were tested individually and connected together in the MCB. The baseline tests indicated that cell metabolism and proliferation are highest in the first 2 days of culture; therefore, these studies were conducted over a period of 24 h, 1 day after seeding. The medium was withdrawn at the outlet of the bioreactor four times in the 24 h as described previously and assayed for glucose, albumin, urea, and nitric oxide. Comparing the glucose consumption per cell at the end of the 24 h of dynamic culture to the same data obtained from the static baseline and coculture studies, we found that dynamic cultures consistently consume significantly higher quantities of glucose per cell (about sixfold more) than their static counterparts (Fig. 8). On the other hand, comparing the three types of dynamic culture (HUVEC monoculture, HepG2 monoculture, and HUVEC-HepG2 connected culture), glucose consumption per cell in the connected culture is less than that in the two separate dynamic cultures, and this result is similar to that observed for the static cocultures. Albumin production per HepG2 cell in the dynamic connected culture and monoculture are reported in Figure 9 and compared with the corresponding data at 24 h in static cocultures and hepatocyte monocultures. In dynamic culture the presence of endothelial cells increased hepatocyte protein synthesis, and albumin release per cell at the end of the 24 h was threefold higher than the corresponding dynamic monoculture. This marked difference was also observed between static cocultures and monocultures, but notably, albumin release per cell in dynamic connected culture is double that in static coculture (Fig. 9). Urea secretion per cell, another marker of hepatic function, was twofold higher in dynamic connected culture than in dynamic monoculture. Moreover, urea production per cell in the dynamic connected culture was about 10 times higher than in static coculture (Fig. 10). Overall, the continuous supply of nutrients as well as convection-mediated crosstalk between hepatocytes and HUVEC lead to increased albumin and urea secretion with respect to the HepG2 dynamic monocultures, static monocultures, and static cocultures.
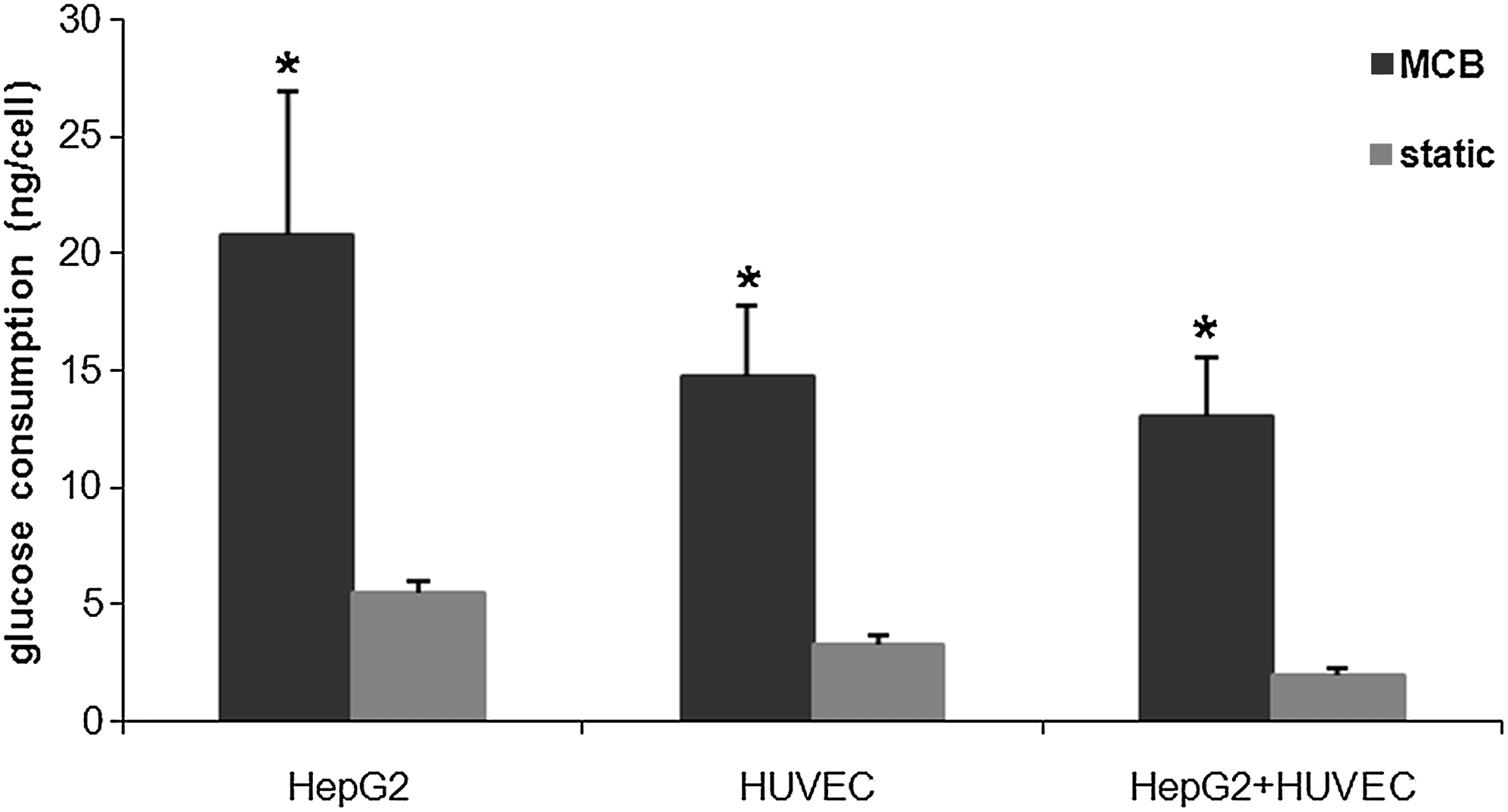
Glucose consumption per cell after 24 h in static mono- and cocultures and in dynamic mono- and connected cultures. The static data are the results obtained from baseline and coculture studies, in which the first point of the curve, on day 1 is considered. n = 3, *p < 0.05 with respect to the corresponding static values.
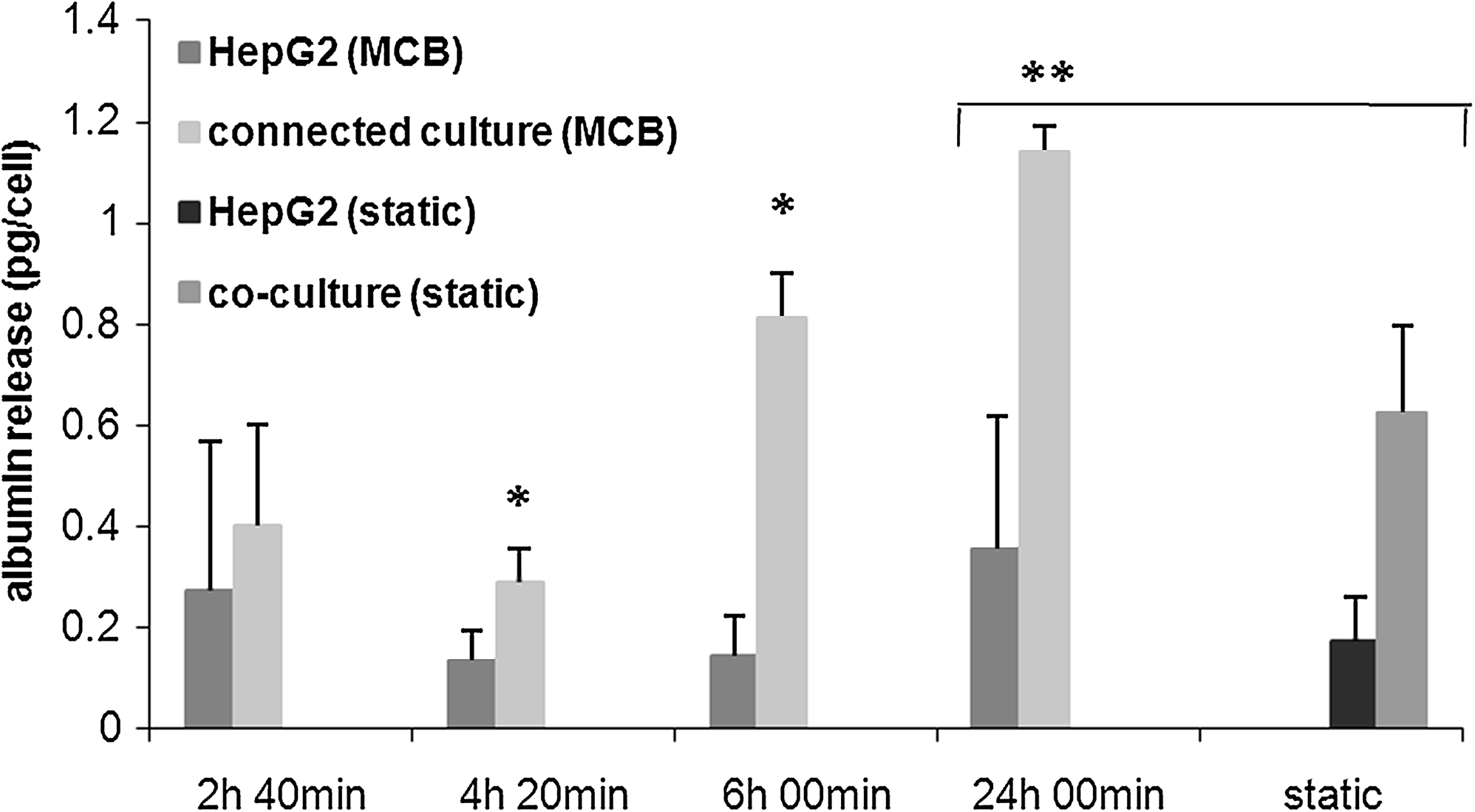
Albumin release per cell in hepatocyte mono- and connected culture in the MCB, and comparison between dynamic and static conditions, n = 3, *p < 0.05 with respect to the corresponding dynamic monoculture values at the same time. **p < 0.005.
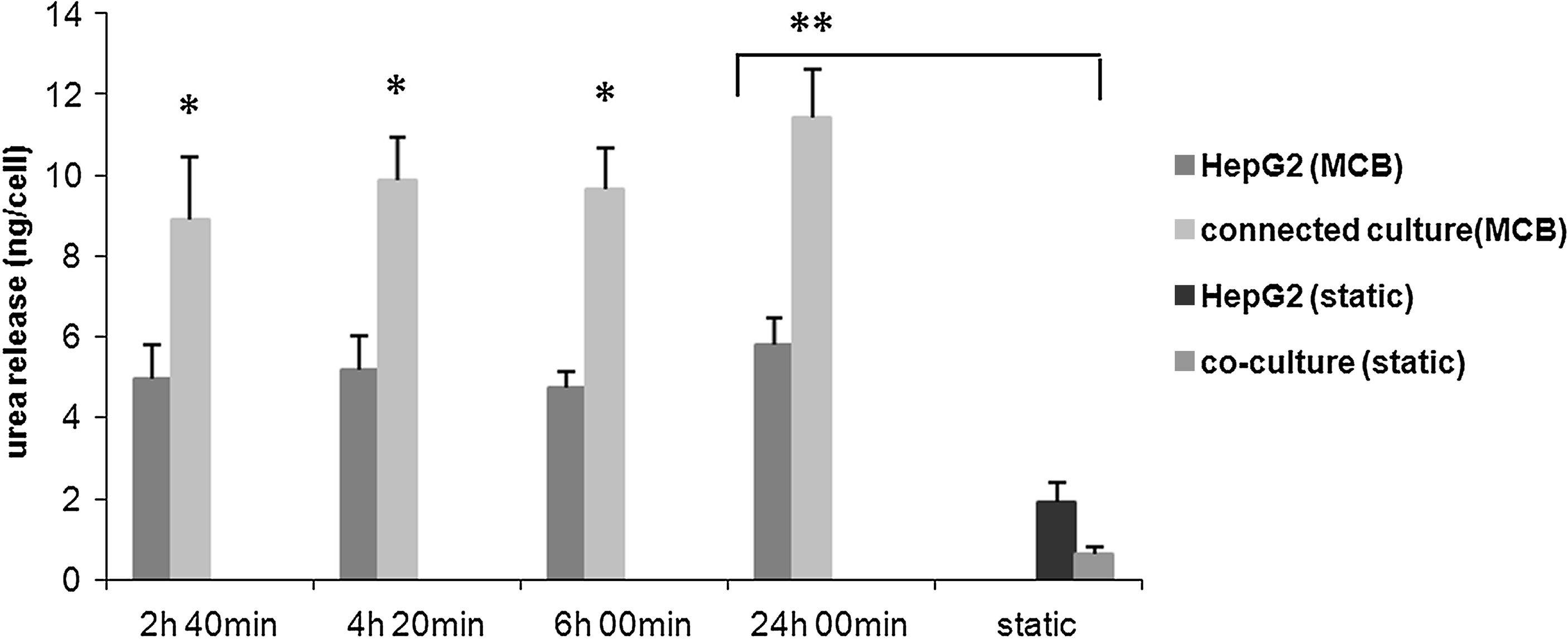
Urea production per cell in HepG2 dynamic mono- and connected culture in the MCB, and comparison between dynamic and static conditions, n = 3, *p < 0.05 with respect to the corresponding dynamic monoculture values at the same time. **p < 0.005.
Finally, we measured nitric oxide concentration to determine if crosstalk influences endothelial response to the flow. The data were compared with those obtained in static HUVEC monoculture and HUVEC and HepG2 coculture. 9 As shown in Figure 11, the quantity of nitric oxide released in both static conditions is much lower than in presence of flow; however, we did not find any significant difference in nitric oxide concentration per cell between HUVEC in mono- and coculture. Therefore, in these experimental conditions, the nitric oxide release is entirely mediated by flow, and the presence of hepatocytes does not appear to have any effect on this vasodilatory response of HUVEC.
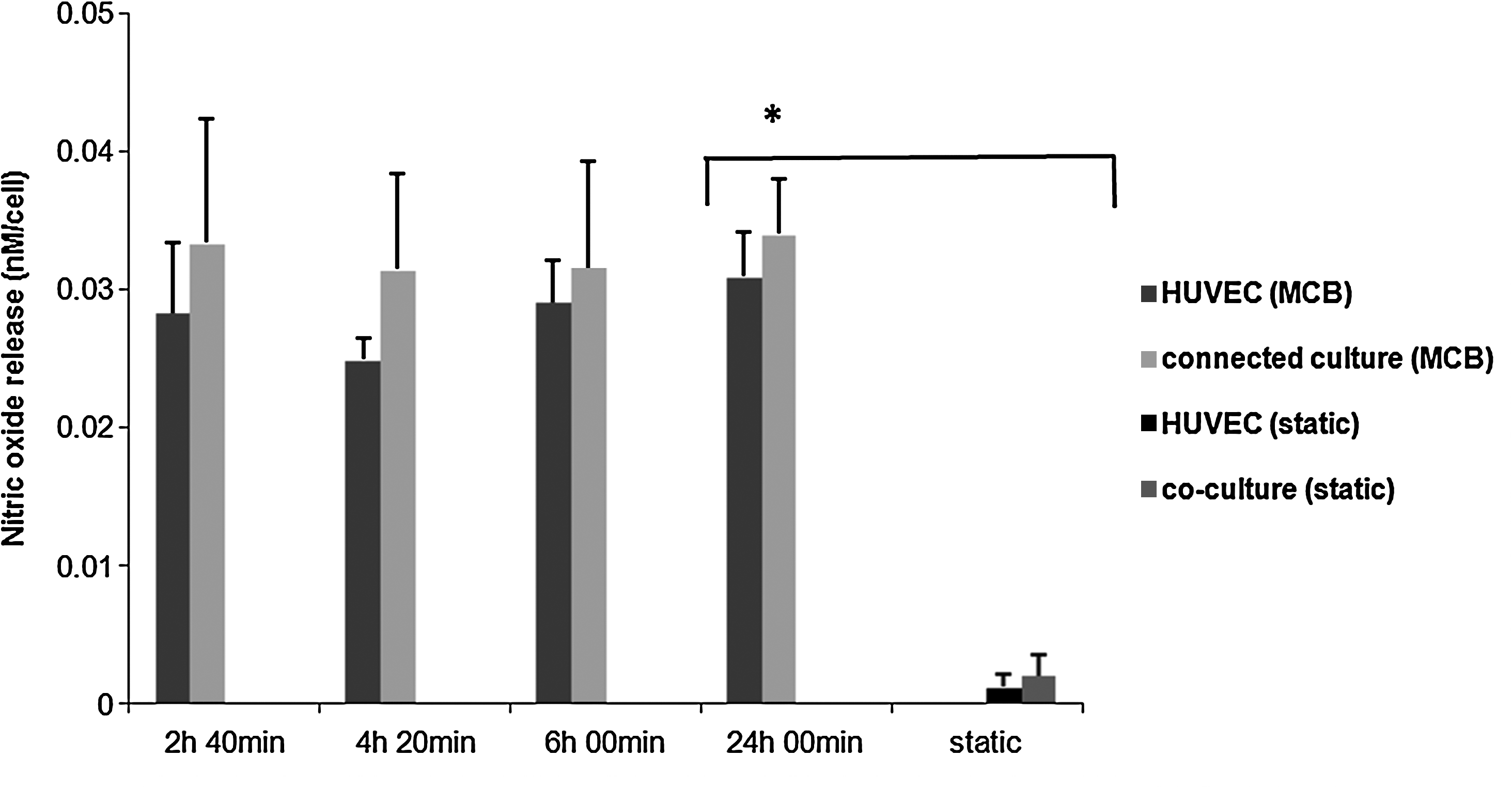
Nitric oxide release per cell in HUVEC mono- and connected culture in the MCB and comparison between dynamic and static condition, n = 3, *p < 0.005 with respect to the corresponding static monoculture values at the same time.
Discussion
It is now recognized that the crosstalk between different tissues is important in modulating and enhancing cell function, and the use of conditioned media, or the addition of growth factors generated by stromal or other cells is increasingly common, as are cocultures. Several studies have already shown that the culture of hepatocytes in collagen sandwich configurations or with nonparenchymal cells enhances liver cell function, particularly albumin and urea secretion.3–5 Some recent reports also use coculture models of cells that are not found together in vivo to provoke pathological situations, such as the adipose induction of insulin resistance in hepatocytes. 18 Traditional in vitro models, which use hepatocyte monocultures or cocultures, can be used to study the effects of materials or cells able to mimic the extracellular matrix on cell proliferation, differentiation, and metabolism.19,20 Further, the influence of homotypic and heterotypic interactions on hepatocyte function can also be investigated. Although these models are useful to study interactions between cells housed in the same tissues, such as hepatocytes and fibroblasts, they are not suitable for the study of crosstalk between cells belonging to different organs. To address this gap, an MCB was developed using biomedical engineering concepts based on biomimetics. Metabolic homeostasis in the body depends on the crosstalk and reciprocal interactions between many different tissues; the MCB was designed to reproduce salient features of the metabolic system with particular regard to the metabolism of glucose, fat, and proteins. The connected culture system can house different cell types in separate chambers, and allows cell–cell crosstalk to be studied by addressing each cell type separately. Besides this biological stimulus, cells are submitted to biomechanical and biochemical stimuli due to the flow of the medium, which increases nutrient supply and removal of catabolites. The main parameters such as time, length, and cell numbers were downscaled using allometric laws, as reported in our previous work. 10 In allometry, mass and cell numbers are scaled linearly, and since cell numbers are simple to control and measure, this was our starting point. The second step was to choose the volume of our system and thus scale fluid and metabolite passage times with respect to the human body using the M1/4 power law for time. 11 Cell cultures are typically carried out in multiwell plates, where volumes may range from 0.3 to 10 mL. So as to conserve cell culture protocols as much as possible, each compartment of the MCB was designed to hold a 12-mm-diameter glass or plastic coverslip and contain 2–3 mL of medium; the total volume of the system was 30 mL. The metabolite of interest was glucose, and there are several references that describe physiological glucose transport and clearance times between different organs. 21 As the liver is the principal organ of metabolic regulation, hepatocytes are the most important cells of our in vitro metabolic system. The liver was thus the focal point for the scaling and design of the allometrically scaled bioreactor. In the model used here, only the hepatic and endothelial compartments were considered, and the endothelial compartment represents the endothelium of the entire body. Given an MCB volume of 30 mL, the downscaled liver to systemic endothelium transport time is about 25 min. The volume to be moved between the liver and endothelial compartment is about 4.3 mL, which corresponds to a flow rate of 175 μL/min. Using this allometric scaling method, it is simple to establish working cell numbers, volumes, and flow rates in a connected cell culture system.
For a given set of cell types, the overall system response is a function of three variables: cell numbers and cell ratios, passage times or flow rates, and total volume. In the bioreactor, cell ratios and medium passage times are scaled using allometry. In this way, cell ratios are conserved to obtain physiologically meaningful proportions between cell types. Having established the ratios, we then use cell numbers and densities that are compatible with standard 12- or 24-multiwell plates to facilitate medium handling and assays, and to allow direct comparisons between the different experimental groups. The flow rate and volume will determine the residence and passage times of molecules, as well as the shear stress, and the level of crosstalk is likely to depend strongly on these variables. For a given volume, the allometric scaling model allows us to establish a flow rate that is high enough to provide a dynamic stimulus in the form of flow and concentration gradients 10 but low enough to ensure that crosstalk signals can be transduced by the cells.
In this work we analyzed the effect of medium flow and the crosstalk between hepatocytes and endothelial cells separately and together, to compare monocultures, cocultures, and connected cultures. Intercellular communication between these two cell types is driven by the flow of interstitial fluid through blood vessel walls and across intercellular spaces, and there is an extensive degree of crosstalk between them, as suggested by the endothelial cell–induced regulation of LDL uptake in hepatocytes 22 and the maintenance of endothelial growth and differentiation in the presence of hepatocytes. 23 In addition, endothelial cells are also known to regulate hepatic function through the release of nitric oxide, a powerful antioxidant,24,25 whose production in physiological conditions is largely flow induced. 26 Comparing the data from the baseline monocultures in static conditions to those obtained from the HepG2 dynamic cultures, we observed an increase in glucose consumption and in albumin and urea secretion as a result of an increased availability of substrates and more efficient catabolite removal (Table 1). This suggests that the flow itself is a stimulus for hepatic cells and leads to an increase in metabolic rate in terms of glucose metabolism, protein synthesis, and nitrogen turnover, even in the absence of endothelial cells. Successively, the effect of culturing hepatocytes and endothelial cells (HUVECs) in the presence of flow, which we term “connected cultures,” was investigated. The presence of HUVEC increased hepatic protein synthesis over 24 h with respect to the static cocultures and the dynamic monocultures. Similarly, urea secretion was much higher in the dynamic connected culture. Therefore, both flow and the presence of HUVEC act synergically to upregulate these two endogenous hepatic functions. To investigate the reciprocal interaction between the two cells, endothelial nitric oxide production was measured in monocultures and connected cultures. In these experiments we confirmed that nitric oxide release is indeed flow-induced, and that the presence of hepatocytes in connected cultures, or cocultures does not mediate the production of this antioxidant, vaso-active factor. Finally, glucose consumption was higher in dynamic conditions than in static culture for all three types of culture; however, the connected culture showed lower glucose consumption with respect to the dynamic HepG2 monoculture (Table 1). Similar results were obtained in, 10 and indicate that crosstalk between endothelial cells and hepatocytes may play a role in glycemic homeostasis.
Results are expressed as means ± standard deviation, n = 3.
MCB, multicompartmental bioreactor.
The results demonstrate that the flow of the medium, which simulates the bloodstream in the MCB model, increases protein synthetic function and nitrogen metabolism, as well as nitric oxide production. Further, crosstalk between HepG2 and HUVEC in connected cultures leads to a further increase in metabolic activity of hepatocytes, and to a more efficient use of the energetic substrate glucose.
Conclusions
The MCB was designed to explore crosstalk between tissues and cells in spatially separate cultures. In the MCB system, different types of cells can be cultured individually but connected to one another through the flow of the medium. This enables each culture to be addressed and interrogated individually. While cocultures are useful for the study and optimization of paracrinal cell function, they are not a suitable model for investigating interactions between cell types belonging to different tissues. In this sense, the MCB represents a more physiological model. In this work we used HepG2 and endothelial cells in static monocultures in 24-multiwell plate as our baseline and compared static cocultures, dynamic monocultures, and connected cultures. Connected cultures enhance albumin synthesis and urea production in hepatocytes compared with static mono- and cocultures and dynamic monocultures. Moreover, in both static and dynamic conditions, the presence of HUVEC and HepG2 decreases the unit consumption of glucose with respect to monocultures. Therefore, as demonstrated here, the connected culture in the MCB system combines the dynamic stimulus of flow with cell crosstalk through soluble ligands so that the unit production of albumin and urea is more than double that of the static cocultures.
Footnotes
Disclosure Statement
No competing financial interests exist.