
Abstract
The parathyroid-hormone-related protein (PTHrP) is an important modulator of bone formation and bone remodeling. High and/or prolonged glucocorticoid (GC) treatments inhibit PTHrP expression in osteoblastic cells and bone formation and repair. We assessed the ability of the N- and C-terminal PTHrP fragments to restore the GC-altered bone regeneration after bone marrow ablation in mice. Animals were administered 3-methylprednisolone or vehicle and PTHrP (1–36) or PTHrP (107–139) every other day, beginning 4 days before marrow ablation in the tibia, and euthanized 12 days later. GC-treated mice showed in the ablated tibia a decrease in bone formation and in osteoblast and sclerostin-positive osteocyte numbers, reduced expression of osteoblastic factors, decreased osteogenesis of bone-marrow-derived cells, an increase in the numbers of multinucleated osteoclasts and adipocytes, and decreased cortical vascularization, as well as altered bone structure (measured by microcomputerized tomography) in the intact femur. These effects were reversed at least in part by either PTHrP peptide. The present novel findings support the use of both PTHrP peptides tested as putative bone regenerative therapies in GC-related bone diseases.
Introduction
Chronic glucocorticoid (GC) exposure is a major cause of bone mass loss through mechanisms incompletely understood. High GC dosing induces an early increase in bone resorption followed by a decrease in bone formation and bone remodeling.15,16 GCs can act on osteoclasts, both indirectly via increasing receptor activator of NF-κB ligand (RANKL) and decreasing osteoprotegerin (OPG) in osteoblasts, and directly by diminishing their bone-degrading activity.17–19 However, the bone sparing action of GCs seems to be mainly accounted for by a deficit in the number of functional osteoblasts and thus in bone formation.19–22 GCs at supraphysiological levels (>100 nM) inhibit PTHrP expression while not affecting or even increasing PTH1R expression and/or signaling in osteoblastic cell cultures.16,23–26 However, the putative relationship between an altered PTHrP/PTH1R system and GC-related osteopenia has not yet been explored. In any event, both anabolic and antiresorptive therapies are efficient in improving bone mass in both experimental animals and randomized controlled trials in humans with GC-related bone disease.16,27,28
Due to the deleterious effects of GCs in bone metabolism, an altered bone repair occurs in experimental models undergoing GC treatment.29,30 Thus, in a bone marrow ablation model in rats, high-dose GC for 3 days inhibited the newly formed bone and the subsequent bone resorption occurring within 2 weeks in the ablated marrow cavity. 29 Of interest in the context of the present study, a recent report has shown that daily administration of PTH (1–34) (40 μg/kg/day) for 3 weeks promotes bone regeneration after marrow ablation in the rat femur. 31 Considering the aforementioned actions of different domains of PTHrP in bone metabolism, we hypothesized that both its PTH-like and its C-terminal domains would promote bone healing after marrow ablation under GC treatment. Hence, we evaluated the efficacy of PTHrP (1–36) and PTHrP (107–139) in this setting in mice as an attempt to provide alternative bone regeneration therapies in GC-related bone diseases.
Materials and Methods
Animal model
Male C57BL/6J mice (16 weeks old) from Harlan Laboratories (Harlan Interfauna Ibérica, Barcelona, Spain.) were stabilized in the Animal Research Facility at Fundación Jiménez Díaz for 2 weeks. Animals were given free access to water and fed a standard diet (8.8 g/kg calcium and 5.9 g/kg phosphate; Panlab, Reus, Spain) in a room maintained at 22°C on 12-h light/12-h dark cycles. Mice were randomly divided into four groups of 10 mice. Three of these groups were administered 3-methylprednisolone (3-Mpred; 10 mg/kg, every other day, subcutaneously), a synthetic GC largely used in humans that lacks mineralocorticoid activity, 32 and the other group was administered phosphate-buffered saline (PBS) as vehicle (control) for 16 days. Similar doses have previously been used in other studies evaluating the skeletal actions of GCs.20,22,33,34 GC-treated mice were also given PTHrP (1–36) or PTHrP (107–139) (100 μg/kg/every other day, subcutaneously), or vehicle (50 mM KCl, pH 4.5; control) from day 1 after the start of 3-Mpred administration. On day 5, all mice in the study were anesthetized with ketamine/xylazine (2:1, v/v) before undergoing mechanical marrow ablation in both tibiae, as recently described. 4 A time period of 12 days for bone regeneration after marrow ablation was chosen based on previous observations in rats, 29 and our initial data in mice (not shown). Thus, on day 17 animals were euthanized (24 h after the last treatment with each PTHrP peptide or its vehicle). Both ablated tibiae from each animal were removed and cleaned of soft tissues. One of them was assigned for histological evaluation; ex vivo marrow cultures were obtained from the other tibia as described below, and the remaining hard bone tissue was kept in liquid nitrogen before total RNA extraction. Intact femurs were also harvested, cleaned, and kept at −20°C until microcomputerized tomography (μCT) analysis. Body weight of mice was recorded daily throughout the study. Animal protocols were approved by the Institutional Animal Care and Use Committee at Fundación Jiménez Díaz.
Histological analysis and immunohistochemistry
One ablated tibia per mouse was fixed in 4% p-formaldehyde at 4°C; subsequently, it was decalcified in Mielodec (Bio-Optica, Milan, Italy), dehydrated, and embedded in paraffin. All histological and immunohistological stainings were carried out on serial saggital 4-μm sections from each mouse in at least five mice per experimental group. Newly formed bone was stained by Masson's and quantified in cross-sectional × 400-fields within the regenerating bone area (1 mm2) at the metaphysis, starting immediately below the growth plate, using Leica QwinPro® V 3.5.0 software (Leica, Heerbrugg, Switzerland). Osteoblasts, identified by their cuboidal shape and localization on newly formed bone and endosteal surfaces,20,34 were also counted in this area, using the same magnification (× 400). Blood vessels in cortical bone were counted in 10 consecutive × 200-fields between the growth plate and the mid-diaphysis. They were identified as elongated or round lectin-positive structures with luminal blood cells. A horseradish peroxidase (HRP)–conjugated lectin from Bandeiraea simplicifolia (L5391; Sigma-Aldrich, St. Louis, MO), showing affinity for endothelial cells, was used. 35 Adipocytes were identified as unstained round spaces void of blood cells, and confirmed by positivity for S-100 (by using immunostaining as described below),4,36 in four to six random × 200-fields between mid- and distal diaphysis.
For immunohistochemistry, antigen was retrieved by incubation with 0.1% trypsin (Lonza, Verviers, Belgium) in PBS for 30 min at 37°C (PTHrP and PTH1R, tartrate-resistant acid phosphatase [TRAP]), with 10 mM sodium citrate buffer (pH 6.0), for 4 min, at 95°C (calcitonin receptor [CTR]) or with target retrieval solution (pH 6.5) (Dako, Glostrup, Denmark), for 20 min, at 95°C. Tissue samples were first incubated in 10% goat serum (PTHrP and PTH1R), rabbit serum (sclerostin), horse serum (CTR), or 0.25% bovine serum albumin and 2.5% goat serum (TRAP) in PBS for 1 h at room temperature. The following primary antibodies were used (dilution factor): rabbit polyclonal anti-PTHrP antiserum C13 (300); 16 rabbit polyclonal antibodies against human PTH1R (H-125; Santa Cruz Biotechnology, Santa Cruz, CA) (250), S100 (RZ0311; Dako) (500), and TRAP (H00000054-D01P; Abnova, Jhongli City, Taiwan) (300); goat polyclonal anti-CTR (sc-8860; Santa Cruz Biotechnology) (500) and anti-sclerostin (AF1589, R&D, Minneapolis, MN) (100) antibodies. Primary antibodies were incubated overnight in a humidified chamber, except for sclerostin and S100, which were incubated for 2 h and 20 min, respectively, at room temperature. Endogen peroxidase was blocked with 3% H2O2 in PBS for 15 min. Sections were subsequently incubated with biotinylated donkey anti-goat IgG (CTR) or goat anti-rabbit IgG (PTH1R), followed by avidin–biotin–HRP complex (Dako), HRP-rabbit anti-goat IgG (sclerostin), HRP-goat anti-rabbit IgG (PTHrP), or Envission-Flex® (Dako) (S-100, TRAP). In all cases, 3,3′-diaminobenzidine was used as chromogen. Sections were counterstained with hematoxylin (Sigma-Aldrich). Some tissue samples were incubated without the primary antibody as negative controls.
PTHrP and PTH1R staining were determined in four to six random ×400-fields per sample in the trabecular area at the proximal metaphysis. Intensity of osteoblastic staining was evaluated according to a semiquantitative score, graded from 0 to 3. Multinuclear osteoclasts (with three or more nuclei) and mononuclear osteoclast precursors were identified as showing CTR or TRAP positivity37–40 and counted in 10–20 consecutive × 400-fields in the same area. Sclerostin-positive and total osteocytes, 41 as well as empty and osteocyte-containing lacunae were counted in four to six random × 400-fields per sample in a cortical bone segment between the growth plate and the mid-diaphysis. All stainings were evaluated by two to three independent observers in a blinded fashion, and the corresponding mean score value was obtained for each mouse.
Ex vivo bone marrow stromal cell cultures
Bone marrow stromal cells (BMSCs) from the ablated tibia were cultured as recently described. 4 Cell suspensions were seeded at 106 (for mineralization studies) or 2.5 × 106 cells/well onto six-well plates. After 3 days in culture and until the end of the experimental period, cells were grown in osteogenic medium containing 50 μg/mL ascorbic acid and 10 mM β-glycerophosphate. Half of the volume of the medium was exchanged every other day. BMSCs were grown in osteogenic medium for 14 days, and the colonies formed were fixed with Merckofix® (Merck, Darmstadt, Germany). Alkaline-phosphatase (ALP)-positive colonies were stained with a histochemical kit (Sigma-Aldrich), following the manufacturer's instructions; total cell colonies per well were estimated in these cultures by crystal violet stain, eluted with 0.2% Triton X-100, and measuring absorbance at 540 nm. 4 The number and area of mineralized nodules at day 21 were determined by staining with 40 mM alizarin red (pH 4.2). ALP-stained and alizarin-red-stained areas were scanned and quantified using Photoshop cs 8.0.1. These analyses were carried out in BMSC cultures from 8 to 10 mice per experimental group.
Real-time polymerase chain reaction
Total RNA was extracted from the bone samples with Trizol (Invitrogen, Groningen, The Netherlands). Retrotranscription was done with the high-capacity cDNA reverse transcription kit (Applied Biosystems, Foster City, CA), and real-time polymerase chain reaction (PCR) was performed in an ABI PRISM 7500 system (Applied Biosystems). Unlabeled mouse-specific primers for Runx2, ALP, osterix (OSX), OC, OPG, RANKL, adipocyte fatty acid-binding protein (Ap2), PTHrP, PTH1R, as well as VEGF and its receptors, VEGFR1 and VEGFR2, and TaqMan MGB probes were obtained by Assay-by-Design™ (Applied Biosystems). Results were expressed in mRNA copy numbers, calculated for each sample using the cycle threshold (Ct) value, and normalized against 18S rRNA, as reported.4,12
μCT analysis
Mouse distal femurs from eight mice per experimental group were scanned with a high-resolution microtomographic system (SkyScan 1172; Skyscan N.V., Aartselaar, Belgium) at Trabeculae®, as recently described. 4 SkyScan™ CT-analyzer software, version 1.7.0.5, was used for determination of the three-dimensional microarchitectural properties of trabecular and cortical bone regions of interest, corresponding to a 1.5-mm length starting at 0.6 mm from the growth plate (excluding primary spongiosa), or a 0.6-mm length region of the diaphysis at 2.5 mm from the growth plate, respectively. The following parameters were calculated: cortical thickness (Ct.Th), cortical bone area (B.Ar), and cortical porosity (Po), as well as trabecular bone volume fraction (BV/TV), trabecular thickness (Tb.Th), trabecular number (Tb.N), trabecular bone pattern factor (Tb.Pf ) (an index of trabecular connectivity), and degree of anisotropy (DA) (a measure of the preferential spatial alignment of trabeculae).
Statistical analysis
Results are expressed as mean ± standard error of the mean. Statistical analysis was performed by Kruskal–Wallis nonparametric analysis of variance followed by Dunn's post hoc test or Mann–Whitney test with statistical software (GraphPad InStat™ V2.04a+). p < 0.05 was considered significant.
Results
The mean (±standard error of the mean) body weight of all mice at the beginning of the study was 27 ± 2 g. A slight increase in body weight occurred in control mice throughout the period of study, whereas a significant decrease in body weight occurred after marrow ablation and continued up to the end of the study in mice administered 3-Mpred (Fig. 1).
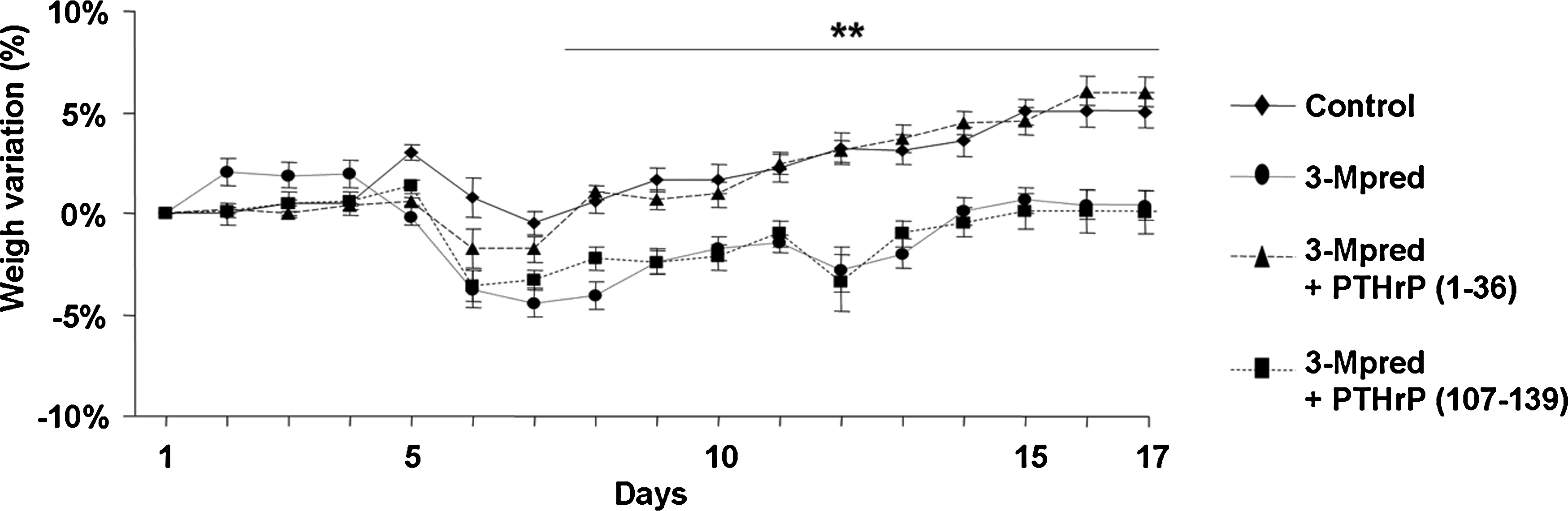
Mouse weight gain through different treatments during the period of study. Values are mean ± SEM (n = 10). **p < 0.01 versus corresponding control value. SEM, standard error of the mean.
Abundant newly formed bone tissue and osteoblasts were observed at the tibial metaphysis of control mice 12 days after marrow ablation, consistent with previous observations in another mice strain, 42 but they significantly decreased in GC-treated mice (Fig. 2A, B). On the other hand, a sharp increase in the number of adipocytes in the mid-distal diaphysis occurred in the regenerating tibia from the latter mice (Fig. 2C). These alterations induced by 3-Mpred in the ablated mouse tibia were also associated with a reduced number of sclerostin-positive osteocytes related to abundant empty lacunae (Fig. 3A), and an augmented number of multinucleated osteoclasts at the expense of less mononuclear precursors (as showing CTR and TRAP positivity) (Fig. 3B).

Effect of PTHrP (1–36) and PTHrP (107–139) administration for 16 days reversed several changes induced by 3-Mpred in the regenerating mouse tibia at day 12 after marrow ablation: the decrease in newly formed bone (

Immunostaining for sclerostin, CTR, and TRAP in the ablated tibiae at day 12 in mice with or without 3-Mpred treatment. Some GC-treated mice were also administered either PTHrP (1–36) or PTHrP (107–139). Immunohistochemistry was carried out in paraffin-embedded tissue sections using specific antibodies as described in Materials and Methods. (
We next evaluated the expression of several osteoblastic genes in the regenerating tibia after marrow ablation in these mice. The mRNA levels of Runx2, a key transcription factor to commit BMSCs toward osteogenesis, 43 and those of OC, a late osteoblastic marker, were decreased in the ablated mouse tibia of GC-treated mice. In addition, expression of other osteoblastic genes controlled by Runx2,43,44 namely, OSX, ALP, OPG, and RANKL, were significantly decreased by 3-Mpred treatment in the healing mouse tibia (Fig. 4A). On the other hand, a significant increase in the gene expression of Ap2, an adipocyte marker, 45 occurred in GC-treated mice (Fig. 4A), consistent with the augmented number of adipocytes as described above. A decrease in PTHrP, but not in the PTH1R, at gene (assessed by real-time PCR) and protein (evaluated by immunostaining) levels, also occurred in this setting (Fig. 4A, B). Since alterations in angiogenesis have been reported to contribute to the deleterious effects of GCs in bone,46,47 we assessed the putative alterations in bone vascularization by 3-Mpred, related to the expression of the VEGF system in our experimental setting. A downregulation of the latter system (Fig. 4A), associated with a lower number of blood vessels (Fig. 2D), was observed in the ablated tibia from GC-treated mice with respect to controls.
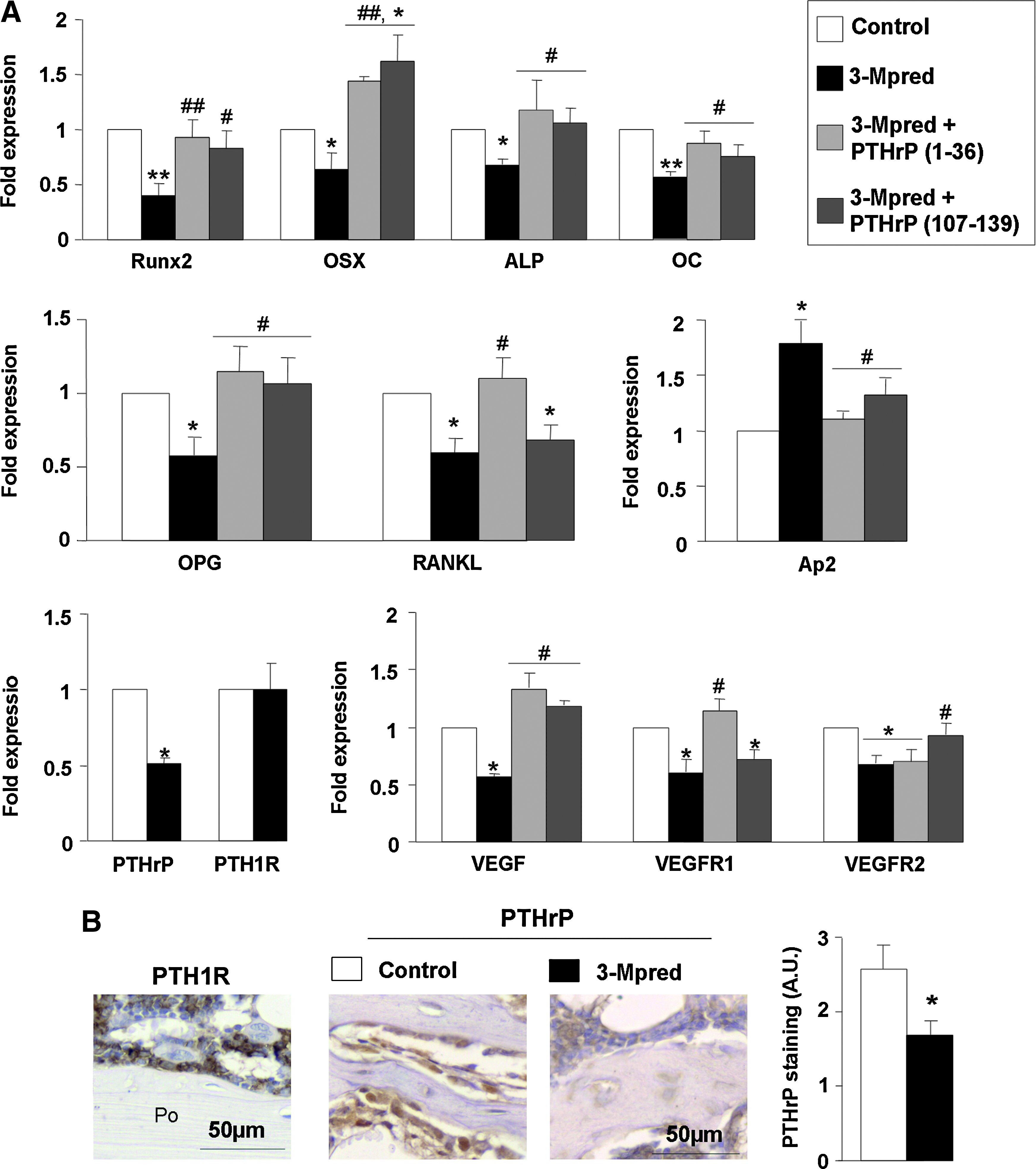
Effect of PTHrP (1–36) and PTHrP (107–139) administration for 16 days on GC-induced alterations in the expression of various bone-related factors in the ablated mouse tibia at day 12. (
These negative effects of 3-Mpred on bone regeneration were also analyzed in ex vivo BMSC cultures from the ablated mouse tibia. A decrease in ALP-positive area, without significant changes in the formation of total colonies—as assessed by crystal violet staining—was observed in these cultures from the GC-treated mice as compared to those from controls (Fig. 5A). The former cultures also showed a significant decrease in both the number and area of mineralized nodules (Fig. 5B).
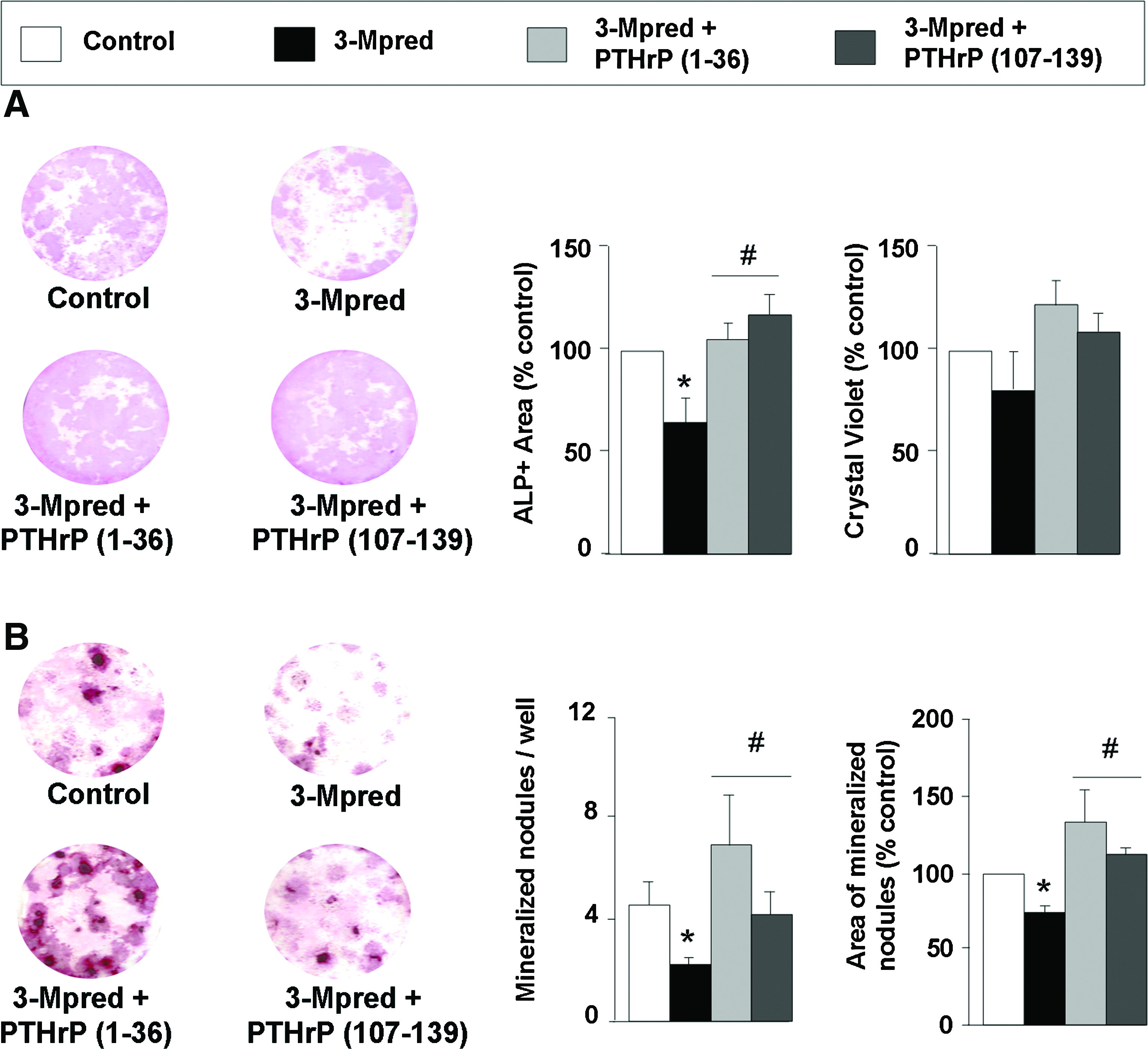
Effect of in vivo administration of PTHrP (1–36) and PTHrP (107–139) on GC-related changes in total and ALP-positive colonies and matrix mineralization in ex vivo BMSC cultures from the ablated mouse tibia. BMSCs were isolated from mice after 3-Mpred treatment, with or without administration of each PTHrP peptide for 16 days, and control mice as described in the text. Total colonies (assessed by eluting crystal violet stain with 0.2% Triton X-100, and measuring absorbance at 540 nm) and ALP-positive colonies (by histochemistry) were assessed in these cultures at day 14 (
Treatment with PTHrP (1–36) for 16 days was found to reverse the body weight loss and histological alterations induced by 3-Mpred in the ablated mouse tibia, related to changes in bone formation (Figs. 1–3) and mRNA levels (except for the decreased VEGFR2 expression) (Fig. 4A). PTHrP (107–139) failed to affect the diminished body weight but reversed the histological changes in the GC-treated mice (Figs. 2 and 3). In these mice, this peptide also normalized the gene expression of the majority of the factors tested, but not that of RANKL and VEGFR1 in the regenerating tibia (Fig. 4A). The osteogenic action of these PTHrP peptides in vivo was confirmed using ex vivo BMSCs from the mouse regenerating tibia. Both PTHrP (1–36) and PTHrP (107–139) were found to reverse the GC-induced decrease in ALP-positive area and matrix mineralization in these cultures (Fig. 5A, B).
To obtain further insights on the osteogenic effects of the PTHrP peptides tested, bone structure was also analyzed by μCT in the intact distal femur from GC-treated mice at the end of the study by μCT. In these mice, a significant decrease in cortical B.Ar and Ct.Th, and an increased Po occurred at the femoral diaphysis (Table 1). In addition, %BV/TV showed a tendency to decrease (p = 0.08; n = 8), related to a reduced Tb.N in GC-treated mice compared to control animals. Moreover, DA and Tb.Pf, representing a measurement of preferential alignment of trabecular structures and trabecular connectivity, respectively, were also significantly altered by 3-Mpred treatment (Table 1). Administration of PTHrP (1–36) reversed these structural changes in GC-treated mice, and it even increased Tb.Th to a higher-than-control value (Table 1). PTHrP (107–139) also restored the cortical alterations in these mice, in contrast to the assessed trabecular parameters that were unaffected, except for DA, by this peptide (Table 1). These alterations in the distal femur of GC-treated mice and the osteogenic effects of each PTHrP peptide are depicted by corresponding μCT images (Fig. 6).
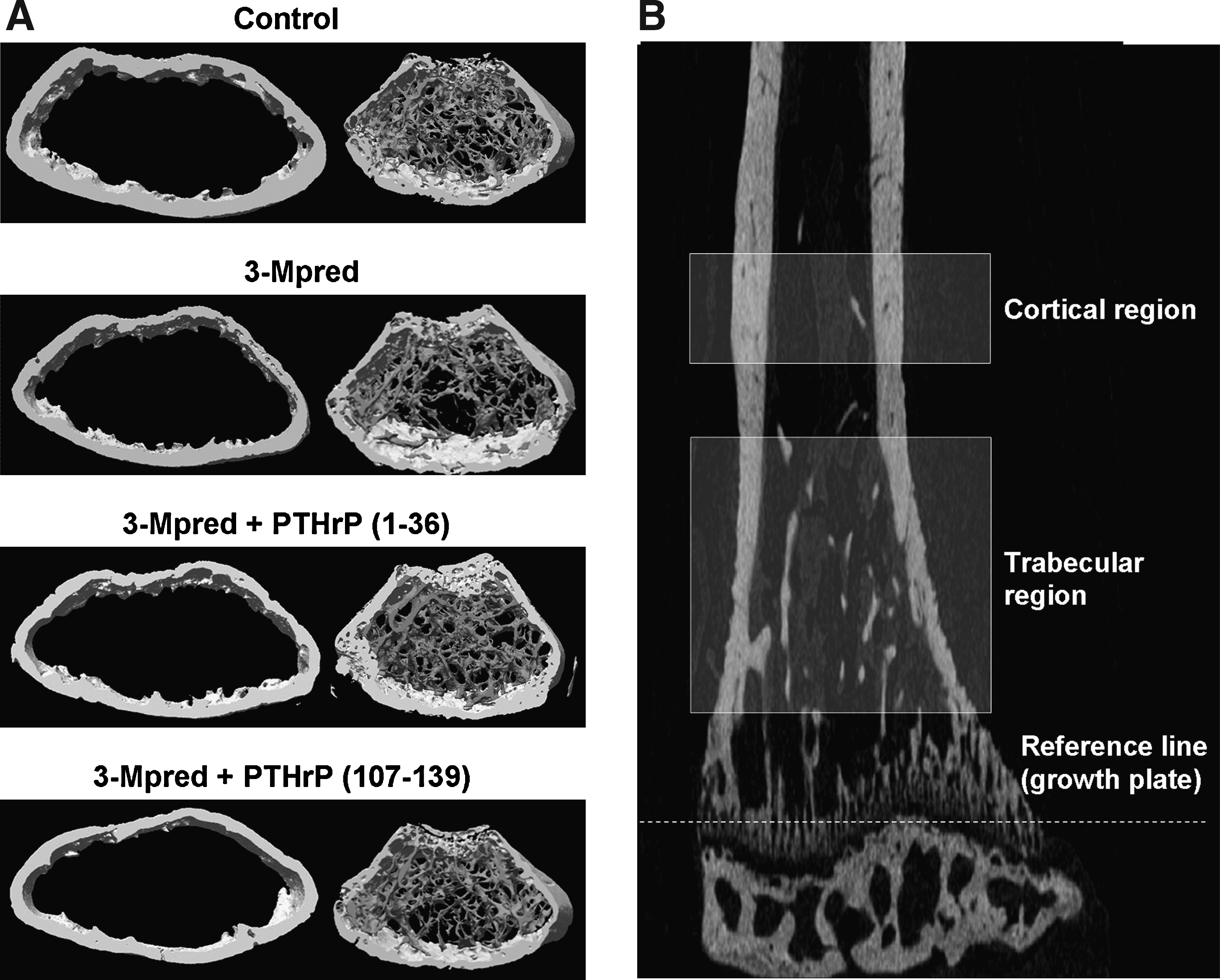
(
Values are mean ± SEM of four to eight animals per group.
p < 0.05 versus control; bp < 0.05; cp < 0.01 versus Mpred.
Ct.Th, cortical thickness; B.Ar, bone area; Po, porosity; BV/TV, trabecular bone volume/tissue volume; Tb.Th, trabecular thickness; Tb.N, trabecular number; Tb.Pf, trabecular bone pattern factor; DA, degree of anisotropy; 3-Mpred, 3-methylprednisolone; PTHrP, parathyroid hormone-related protein.
Discussion
GC administration in mice reproduces most of the alterations in intact bone as observed in humans.19,20,33,34 In fact, a decrease in cortical and trabecular bone parameters was observed here by μCT analysis in the intact femur of 3-Mpred-treated animals, in agreement with previous findings using longer dosing of dexamethasone or prednisolone in mice.27,33 Our present data show that bone regeneration after marrow ablation in mice was impaired by 3-Mpred, related to a diminished osteogenic differentiation capacity of BMSCs. A decreased OC mRNA expression was found to occur in the regenerating tibia from GC-treated mice. In this regard, low levels of circulating OC have been reported in mice under CG treatment.19,20,27,33 Gene expression of earlier osteoblast differentiation markers, namely, Runx2, OSX, and ALP, 44 were also downregulated by 3-Mpred in this setting. In agreement with the current notion that factors controlling osteoblastogenesis also affect adipogenesis in an opposite manner, 44 Ap2 gene was upregulated in the regenerating tibia from GC-treated mice.
A reduced PTHrP and VEGF system were observed in the regenerating tibia from GC-treated mice. A similar downregulation has recently been found in mice with diabetes-related osteopenia. 4 Moreover, a parallel decrease in PTHrP and VEGF production has also been demonstrated in differentiating human osteoblastic cells from aging subjects. 3 In addition, PTHrP can stimulate the VEGF system in these and other cell types.12,13,48,49 These aggregated findings suggest that the GC-induced decrease in PTHrP might be in part responsible for the concomitant reduction in the VEGF system. Of note, VEGF has recently been shown to have a key role in the coordinated regulation of angiogenesis and bone formation during distraction osteogenesis in mice. 50 In fact, the decreased number of vessels in the metaphysis of the regenerating tibia from GC-treated mice as observed herein is also consistent with this notion.
Some in vitro studies have shown inhibition of OPG and induction of RANKL by GCs in human osteoblastic cells, but other investigators failed to find such effects in murine osteoblastic cell preparations.18,40,51 In the present in vivo study, a similar decrease in both OPG and RANKL mRNA, resulting in an unchanged OPG to RANKL mRNA ratio, occurred in the regenerating tibia from GC-treated mice. Thus, considering the well-characterized role of this ratio in osteoclastogenesis, it is not surprising that the total number of CTR- and TRAP-positive cells (including committed osteoclast precursors) was unmodified in the ablated tibia from these mice. Interestingly, a significant increase in the relative abundance of multinucleated cells showing positivity for these markers (mature osteoclasts) was found in the GC-treated mice, consistent with previous findings in nonregenerating mouse bone.17,19,34 Considering the deficient bone regeneration in the ablated mouse tibia here, and the current notion that osteoclast activity appears to be important for maintaining bone formation, 52 a deficient osteoclast activity might well occur in GC-treated mice. In this regard, recent reports have shown that the most important effect of GCs on osteoclasts might be inhibiting their activity.19,29,40 Since no histomorphometry analysis was performed to evaluate bone resorption, however, we cannot arrive at any conclusion on whether this is also the case in our model.
Our present finding that a decreased number of sclerostin-positive osteocytes occur in the regenerating tibia from the GC-treated mice might be explained by an increased osteocyte apoptosis related to GC-induced bone loss, as previously suggested.20,21,53 As evaluation of osteocyte apoptosis presents some technical difficulties (reviewed in Jilka et al. 54 ), we assessed instead the relative abundance of empty lacunae in the mouse cortical bone, 55 and found it significantly increased in the GC-treated mice.
Our aggregated data clearly indicate that GC treatment hampers osteoblastic differentiation during bone healing in the ablated mouse tibia. A recent study in rats failed to detect any significant effect of 3-Mpred on ALP and type 1 collagen as assessed by immunohistochemistry and in situ hybridization, respectively, 1 week after bone marrow ablation. 29 At this time period, the newly formed bone volume was lower in the GC-treated group than in controls, but it was less rapidly remodeled within the following week, related to a diminished bone resorption, in the former. 29 In this study, a much higher dose of 3-Mpred than that used herein was administered for only 3 days; thus, differences in GC treatment patterns and/or species (rat and mouse) might account for the observed differences between this study and the present work.
Daily administration of PTHrP (1–36) for 16 days was shown to reverse the deleterious effects triggered by 3-Mpred in the femoral metaphysis, consistent with the anabolic action of this PTHrP fragment in various in vivo scenarios.5–7 A similar treatment with PTHrP (107–139) in these mice was found to compensate these effects on cortical bone and also on trabecular alignment (but not on the other trabecular parameters tested). In this regard, a single previous study has compared the putative osteogenic effects of daily administration of the closely related peptide osteostatin and PTHrP (1–34), each at 30 nmol/kg, for 13 days to OVX rats. Both PTHrP peptides restored femoral bone mass and calcium content, but osteostatin did so apparently due to effects on cortical bone. 5 These osteogenic effects of each PTHrP peptide as observed herein do not seem to depend on changes in mouse weight, which were slight (about 5%) and had a different outcome at the end of the treatment with each PTHrP peptide.
In this study, treatment with either PTHrP peptide was able to reverse the following GC-related changes in the ablated mouse tibia: the decrease of osteoblast number and function, the decrease of sclerostin-positive osteocytes, and increase of empty lacunae, as well as the altered angiogenesis and adipogenesis, which were induced by 3-Mpred in the ablated mouse tibia. Moreover, both PTHrP (1–36) and PTHrP (107–139) were found to normalize the GC-related alterations in the formation of both ALP-positive and mineralized colonies in ex vivo BMSC cultures from the regenerating mouse tibia. Interestingly, treatment with each PTHrP peptide was shown to prevent the formation of mature osteoclasts, an event apparently more related to normalization of OPG than to changes in RANKL expression, in the regenerating tibia from GC-treated mice. These findings are consistent with the known inhibitory effect of PTHrP (107–139) on osteoclastogenesis and osteoclast activity,10,56,57 and also with the fact that the anabolic effect of the N-terminal PTHrP fragment in previous studies apparently occurs independently of an increased bone resorption.7,31 In addition, PTHrP (1–36) increased VEGFR1 but not VEGFR2 gene expression, whereas the opposite occurred for the action of PTHrP (107–139) in the present in vivo setting. This further supports the idea that each PTHrP domain modulates VEGFR1 and VEGFR2 in a specific manner, as recently suggested.12,13,49
In summary, the present results demonstrate that an impaired bone healing after marrow ablation occurs in mice undergoing 3-Mpred administration, and both the N- and C-terminal domains of PTHrP exert similar osteogenic effects to enhance this process. Although each PTHrP peptide on osteopenic bone might differ in a nonregenerative scenario, consistent with previous findings. 5 These findings expand the role of these PTHrP peptides as putative regenerative therapeutic agents to promote bone tissue regeneration in the context of GC-related bone diseases.
Footnotes
Acknowledgments
We thank A.F. Stewart and A. García-Ocaña (Division of Endocrinology and Metabolism, University of Pittsburgh School of Medicine, Pittsburgh, PA) for providing human PTHrP (1–36), and F. Roncal (Proteomics Unit, Centro Nacional de Biotecnología, Madrid) for human PTHrP (107–139) synthesis. We are also indebted to J. Arrieta (Departamento de Estomatología, Fundación Jiménez Díaz) for generously supplying the dental drills; T. Carrizosa (Departamento de Anatomía Patológica, Fundación Jiménez Díaz) for performing S-100 immunostaining; I. Mahillo (Departamento de Epidemiología, Fundación Jiménez Díaz) for assistance in statistical analysis; M. Corcuera and F. Gómez (Departamento de Anatomía Patológica, Hospital Carlos III, Madrid) for assistance in image analysis; and M. Davis for proofreading the article. This study was supported in part by grants from Spanish Instituto de Salud Carlos III (PI050117, PI080922, and RETICEF RD06/0013/1002), the Spanish Ministerio de Educación y Ciencia (SAF2005-05254), and Fundación de Investigación Médica Mutua Madrileña. L.F. de C. and D.L. are predoctoral fellows of Fundación Conchita Rábago. S.P.-N. was the recipient of a research contract from Instituto de Salud Carlos III (RETICEF RD06/0013/1002).
Disclosure Statement
The authors have nothing to disclose.
Portions of this study were presented at the 35th European Symposia on Calcified Tissues, May 24–28, 2008 (this short oral paper was awarded with an European Calcified Tissue Society Travel Award), and the 30th Annual Meeting of the American Society for Bone and Mineral Research, September 12–16, 2008, in Barcelona (Spain) and Montréal (Québec, Canada), respectively.