
Abstract
In this study we generated human skin equivalents (HSEs) under submerged conditions mimicking the aqueous in utero environment and investigated the morphology and differentiation process of the formed epidermis. Further, the skin barrier, which resides in the stratum corneum (SC), was characterized by its lipid content, hydration level, and natural moisturizing factor level. The submerged HSEs showed comparable tissue morphology and similar expression of several differentiation markers and SC lipid composition compared with HSEs grown at the air–liquid interface and native human skin. The SC of the submerged HSEs, however, contained more free water and less natural moisturizing factors compared with the air-exposed counterparts. These results show that the presented cell culture method can be utilized to generate HSEs under submerged conditions to study epidermal formation under aqueous conditions.
Introduction
It is ironic to notice that the barrier that protects the human body from desiccation develops in an aqueous environment, that is, the amniotic fluid. The formation of human skin starts early during embryogenesis and at first consists of only one cell layer of multipotent epithelial cells. This epithelium is covered by a second epidermal layer, called the periderm, which serves as a protective shield between the amniotic fluid and the epidermis while it keratinizes.1,2 The periderm is shed at approximately 24 weeks of estimated gestational age (EGA) when the epidermis is fully differentiated and comprises the first few SC layers. During the third trimester the SC is fully exposed to the surrounding amniotic fluid while it matures. At 34 weeks EGA barrier formation is complete. Full-term neonates (40 weeks EGA) are therefore born with a competent skin barrier that resembles that of adults.
Several attempts have been made to simulate the intrauterine epidermal development by culturing keratinocytes under submerged conditions in Petri dishes or culture flasks.3–7 In these culture systems the medium was supplied only from the apical side of the keratinocytes. The resulting cultures, referred to as human skin equivalents (HSEs), show drastic differences with human skin, such as a disorganized epithelium and an incomplete differentiation indicated by the lack of a SC, the absence of keratohyalin and membrane-coating granules, or the lack of expression of high-molecular-weight keratins. Only when keratinocytes are seeded on an appropriate substrate and subsequently cultured at the air–liquid interface do HSEs show high resemblance to human skin in many aspects such as morphology, expression of several differentiation markers, and formation of the SC.7–15 As the microenvironment of a HSE affects its differentiation process, in utero epidermal development may differ from epidermal development at the air–liquid interface. In this study we have developed a novel method to generate HSEs under aqueous conditions by submerging HSEs in culture media and ensuring that the apical and basolateral side of the developing epidermis are surrounded by an aqueous environment that mimics the in utero environment very closely. The morphology and differentiation process of the epidermis generated under these submerged conditions were examined by determining the expression of various differentiation markers. As the lipids of the SC are crucial for maintaining a proper barrier function, the lipid composition of the SC was examined as well. In addition, the natural moisturizing factor (NMF) and hydration level in SC were determined. Our study showed for the first time that under submerged conditions a well-organized epidermis, including a SC, can be formed.
Materials and Methods
Cell culture
Normal human keratinocytes (NHKs) and human dermal fibroblasts were obtained from adult donors undergoing mammary or abdomen surgery and were established as described previously. 16
Dermal equivalents
Dermal equivalents were generated as described earlier.16,17 In brief, 1 mL of a 1 mg/mL collagen type I solution was pipetted into filter inserts (Transwell cell culture inserts, membrane diameter 24 mm, pore size 3 μm; Corning Life Sciences, Amsterdam, The Netherlands) and was allowed to polymerize for 15 min at 37°C. Subsequently, 3 mL of a 2 mg/mL collagen type I solution, with a fibroblast suspension reaching a final density of 0.4 × 105 cells/mL collagen, was pipetted into the inserts. After polymerization, Dulbecco's modified Eagle's medium (Invitrogen, Breda, The Netherlands) supplemented with 5% fetal bovine serum (Hyclone, Logan, UT), 1% penicillin/streptomycin, and 0.45 μM vitamin C (Sigma, Zwijndrecht, The Netherlands) was added to each culture. The dermal compartments were kept under submerged conditions for 1 week. During this period the medium was refreshed once.
HSEs
Generated on dermal equivalents
HSEs grown on collagen matrices were generated as described previously.9,16 First or secondary NHKs were seeded onto fibroblast-populated collagen gels (0.5–1 × 106 cells/gel). To mimic the aqueous in utero environment, the cultures were kept under submerged conditions during the entire culture period. To investigate the epidermal development under submerged conditions over time, HSEs were grown for 8, 16, and 24 days after seeding of the NHKs at 37°C, 93% relative humidity (RH), and 8% CO2. As a control, HSEs were also generated under air-exposed conditions by seeding keratinocytes on the dermal compartment and lifting them to the air–liquid interface after 1 day. During the culturing period the submerged and air-exposed HSEs were nourished with different media, as described by Bouwstra et al. 16
Generated on inert filter
To generate HSEs under submerged conditions, NHKs (1.0 × 106 cells/filter) were seeded directly onto cell culture inserts (Transwell cell culture inserts, membrane diameter 24 mm, pore size 0.4 μm). After 4 days the culture medium was supplemented with 1 ng/mL epidermal growth factor (Sigma) during the remaining culture period, using the same incubator conditions as the HSEs generated on the collagen substrate. As a control, NHKs were also seeded onto cell culture inserts and were lifted to the air–liquid interface after 4 days. The filter HSEs were fed with the same media as the collagen HSEs.
Amniotic fluid-treated HSEs
Human amniotic fluid samples were obtained by amniocentesis, provided by the Leiden University Medical Centre, from women undergoing cesarean section at 37–39 weeks of pregnancy. Amniotic fluid samples were centrifuged for 6 min at 1000 rpm to obtain a cell-free solution and were stored at −20°C until use. To closely mimic the in utero environment, HSEs generated on a collagen substrate were kept submerged with medium or grown at the air–liquid interface for 8 days. Then a metal ring with a diameter of 10 mm was placed on top of the HSEs and 200 μL amniotic fluid of one donor was applied at the apical side of the HSE until day 24. Amniotic fluid was refreshed twice a week. The amniotic fluid-treated HSEs were nourished with the same media as the collagen HSEs.
Morphology and immunohistochemistry
Harvested HSEs were fixed in 4% (w/v) paraformaldehyde (Lommerse Pharma, Oss, The Netherlands) overnight and sequentially dehydrated in 70%, 80%, 90%, 96%, and 100% ethanol, xylene, and liquid paraffin for 1 h each. The dehydrated samples were subsequently embedded in paraffin. 5 μm sections were cut, deparaffinized and rehydrated in preparation for morphological analysis and immunohistochemical staining of keratin 10 and 16, filaggrin, involucrin, and loricrin. The primary and secondary antibodies used in this study are listed in Table 1. Morphological analysis was performed by light microscopic examination of hematoxylin and eosin-stained sections. Immunohistochemical analysis for keratin 10, keratin 16, and filaggrin was performed as described previously. 16 Staining of involucrin and loricrin was performed as described with the following modifications: treatment with the citrate buffer was omitted, an additional wash with 0.5% Triton X-100 (Sigma) was performed prior to the incubation with human serum, and incubation with the first antibody was performed for 1 h at room temperature.
Lipid extraction and analysis
Human epidermis and the epidermis of HSEs were consecutively extracted according to a modified Bligh and Dyer procedure, 18 with the addition of 0.25 M KCl to extract polar lipids. The extracted lipids were redissolved in a suitable volume of chloroform:methanol (2:1) and stored at −20°C until use. The lipid extracts of three HSEs generated under the same culture conditions were pooled to obtain a sufficient amount of lipids for analysis. Amniotic fluid of three donors was extracted accordingly. The extracted lipids were analyzed by means of one-dimensional high-performance thin layer chromatography as described previously, 19 with the solvent systems provided in Table 2A, B. Cochromatography of a standard lipid mixture was performed to identify the various lipid classes.
A, acetone; C, chloroform; DCM, dichloromethane; DE, diethyl ether; E, ethanol; EA, ethyl acetate; H, hexane; HA, hexyl acetate; M, methanol.
2P, 2-propanol; EMK, ethyl methyl ketone; W, water.
Cryo-scanning electron microscopy
The harvested HSEs were processed as described elsewhere. 16 In brief, the samples were cryo-fixed at −180°C and planed perpendicular to the skin surface. The surface of the samples was visualized at −190°C with a field scanning electron microscope (Jeol 6400F; Jeol, Tokyo, Japan). As a control, dermatomed human skin (300 μm) was incubated for 24 h at 37°C and 93% RH. Cryo-scanning electron microscopy (cryo-SEM) images of at least four different HSEs were taken per culture condition.
NMF content determination
The pyrrolidone carboxylic acid (PCA) content, one of the main components of NMF, in HSEs was determined. The HSEs were clamped between two plates with an opening of 1 cm in diameter in the upper plate and tape-stripped with Scotch Magic Tape 810 tape-strips (3M, Zoeterwoude, The Netherlands). The PCA and the protein content per strip were determined as described previously. 16
Results
Formation of a fully differentiated epidermis, including a SC, under submerged conditions
To determine whether all the epidermal layers are generated under conditions mimicking the in utero environment, HSEs were cultured under submerged conditions with amniotic fluid or medium. As a control, HSEs were also generated under air-exposed conditions. Vertical sections stained with hematoxylin and eosin were used to evaluate the general tissue architecture. At day 24, the amniotic fluid-treated and submerged HSEs showed the presence of all epidermal strata, including a SC. These HSEs showed a similar appearance as the air-exposed HSEs, although some small differences were noticed (Fig. 1).
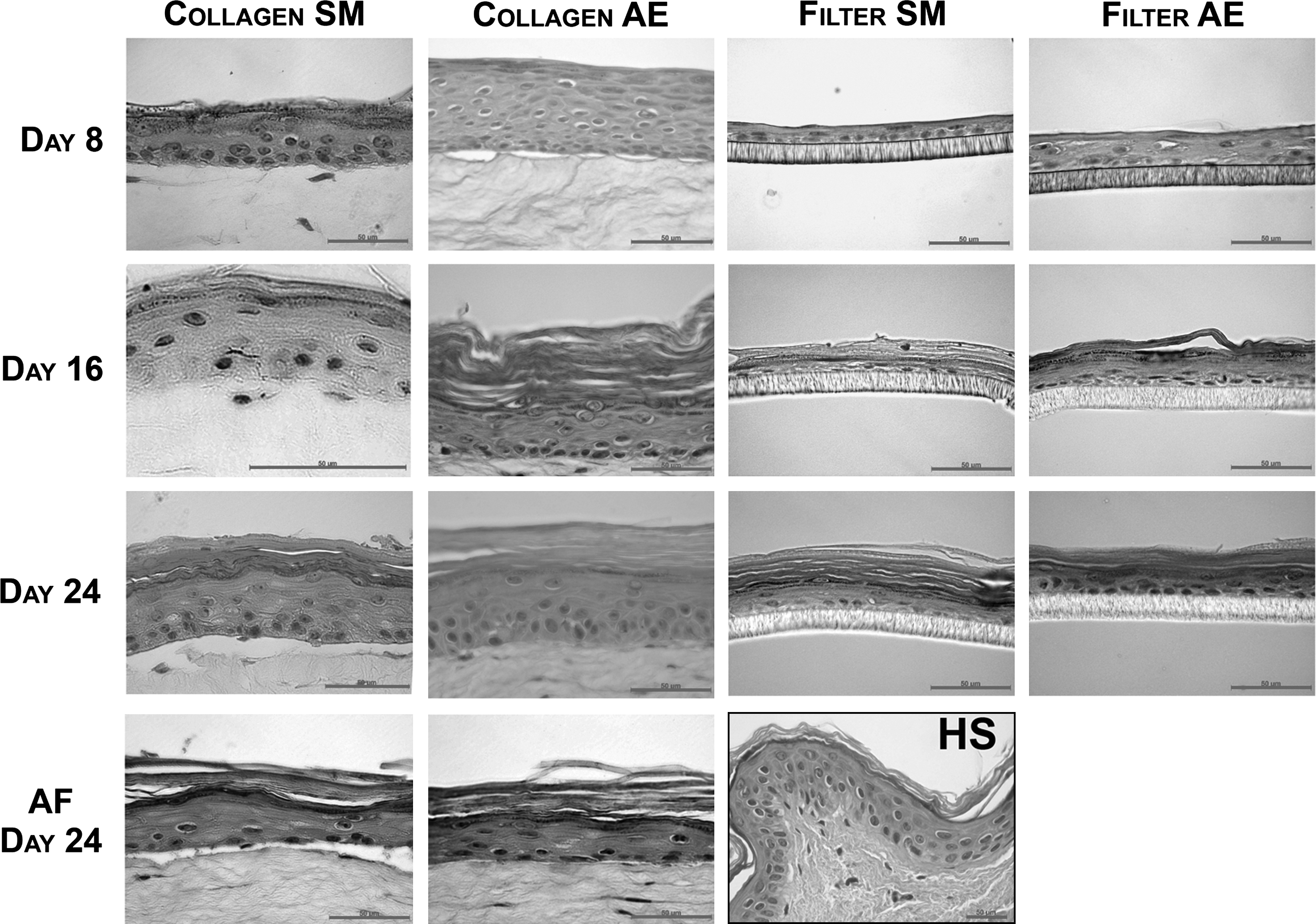
Morphology of submerged and air-exposed HSEs. Hematoxylin and eosin staining of HSEs generated under submerged conditions with amniotic fluid or medium, and air-exposed conditions, and harvested after 8, 16, and 24 days after seeding of normal human keratinocytes on a fibroblast-populated collagen matrix (Collagen) or inert filter (Filter). The morphology of at least three HSEs per culture condition was examined. Scale bars represent 50 μm. SM, submerged; AE, air-exposed; AF, amniotic fluid; HS, human skin.
HSEs were also harvested after 8 and 16 days to investigate the epidermal morphogenesis over time. HSEs generated on a collagen substrate under submerged conditions with medium contained all epidermal cell layers including the stratum basale, stratum spinosum, stratum granulosum, and even a few SC layers at day 8, similar to the air-exposed HSEs (Fig. 1). The number of viable cell layers, however, was less when compared with the air-exposed counterparts. At day 16, these submerged HSEs had a fully differentiated epidermis, including a clearly visible SC. No difference was detected in the number of viable cell layers between days 8 and 16. Between days 16 and 24, the SC of the submerged HSEs increased in thickness, but the number of viable cell layers did not change. HSEs generated under air-exposed conditions showed a decrease in the number of viable cell layers from days 8 to 16, whereas the number of SC layers increased. Although the submerged and air-exposed HSEs developed differently over time, at day 24, the submerged and air-exposed HSEs showed many similarities: all viable epidermal cell layers and the SC were generated, which resembled the morphology of human skin.
The submerged HSEs generated on a filter consisted of only three to four cell layers at day 8, similar to the air-exposed HSEs generated on filter (Fig. 1). At this time point a stratum granulosum and SC were not yet uniformly present. However, at day 16, all the strata in the submerged and air-exposed HSEs were formed, including the stratum granulosum and the SC. Nevertheless, the stratum spinosum consisted of only one to two cell layers and the stratum granulosum was thicker than observed in human skin. At day 24, the submerged and air-exposed HSEs generated on filters showed a similar morphology. Under both culture conditions the number of stratum spinosum cell layers did not normalize at the end of the culture period.
Submerged and air-exposed HSEs show similar expression of protein differentiation markers
To determine whether the differentiation process of the HSEs that were kept submerged with amniotic fluid or medium was similar to that of the air-exposed HSEs and human skin, the expression of several specific differentiation markers was investigated. Only HSEs treated with amniotic fluid from days 8 to 24 were stained to examine the effect of amniotic fluid on the differentiation process. Further, HSEs generated on a filter and harvested after 8 days were not stained, as these cultures did not develop a fully differentiated epidermis yet.
HSEs generated on a collagen substrate that was kept submerged with amniotic fluid or medium showed similar expression of the protein differentiation markers keratin 10, keratin 16, filaggrin, loricrin, and involucrin as the air-exposed HSEs, irrespective of the culture period. Keratin 10, a differentiation-specific marker, was located in all the viable cell layers, except the basal layer, whereas filaggrin and loricrin expression was confined to the granular layer. The expression of these proteins is similar to the expression detected in human skin. In contrast, the expression of involucrin and keratin 16 differed from the expression observed in human skin. The submerged as well as the air-exposed HSEs showed suprabasal expression of keratin 16 and involucrin at days 16 and 24. In human skin, involucrin is only expressed in the stratum granulosum and keratin 16 is only expressed under pathological conditions.
The submerged HSEs generated on filters also showed similar expression of keratin 10, filaggrin, and loricrin as the air-exposed HSEs and human skin from days 16 to 24 (Fig. 2). Again the expression pattern of involucrin and keratin 16 in HSEs generated on filters differed from the expression detected in human skin. From days 16 to 24, involucrin could be detected throughout the entire viable epidermis of both the submerged and air-exposed HSEs. At day 16, keratin 16 expression was also detected in the entire viable epidermis of the submerged HSEs, but only in the suprabasal layers of the air-exposed HSEs. However, at day 24, the air-exposed HSEs also showed weak keratin 16 expression in the basal layer.

Expression and localization of protein differentiation markers. Immunohistochemical staining of keratin 10, keratin 16, filaggrin, loricrin and involucrin of HSEs generated under air-exposed and submerged conditions either on fibroblast-populated collagen matrices or on filters, and collagen HSEs treated with amniotic fluid. Gray-colored squares indicate that the differentiation marker is expressed in the corresponding layer of the viable epidermis. The expression of the majority of proteins was similar irrespective of the substrate used, with the exception of K16 and involucrin. Weak and strong expression, indicated by the half-tone and black squares respectively, of these differentiation markers were additionally detected in the basal cell layer in HSEs generated on an inert filter. White squares indicate that no expression of the differentiation marker was detected in the specified epidermal layer. HSEs generated on filter and harvested after 8 days are not included in this figure. Examples of SM collagen HSE sections are shown. AF, amniotic fluid; AE, air-exposed; SM, submerged; VE, viable epidermis; NHS, normal human skin; K10, keratin 10; K16, keratin 16; Fil, filaggrin, Lor, loricrin; Inv, involucrin; SG, stratum granulosum; SS, stratum spinosum; SB, stratum basal. Color images available online at www.liebertonline.com/ten.
Submerged and air-exposed HSEs have similar SC lipid profiles
The lipid profiles of the HSEs generated under submerged conditions with amniotic fluid or culture medium and under air-exposed conditions were investigated to determine whether qualitative changes occurred when culturing under different conditions. Lipid analyses showed that the SC lipid profiles of HSEs grown submerged in culture medium and air-exposed conditions, both in the case of HSEs generated on a collagen substrate or filter, are very similar at day 24 (Fig. 3A). The lipid profiles of the submerged and air-exposed HSEs reveal the presence of all barrier lipids—cholesterol, free fatty acids, and ceramides—which are also present in human SC. Moreover, all ceramide subclasses are present in the submerged and air-exposed HSEs, but the free fatty acid levels in the HSEs are lower than in the native tissue. Further, the lipid profile of the submerged HSEs shows a lower level of free fatty acids and esterified ω-hydroxyacid sphingosin (EOS) ceramide than the air-exposed counterparts. Additionally, when cultured on a collagen substrate, the presence of an additional ceramide with a retention factor (Rf value) between that of ceramide EOS and the non-hydroxy fatty acid sphingosine A (NS-A) ceramide can be detected in the submerged HSEs. The detailed structure of the unidentified lipid remains to be established. Lipid analyses of amniotic fluid-treated HSEs (Fig. 3B) showed the presence of all barrier lipids, which appeared to be present in slightly higher amounts than in HSEs that were not treated with amniotic fluid (data not shown). However, further analysis showed that all barrier lipids were also present in amniotic fluid (Fig. 3B).
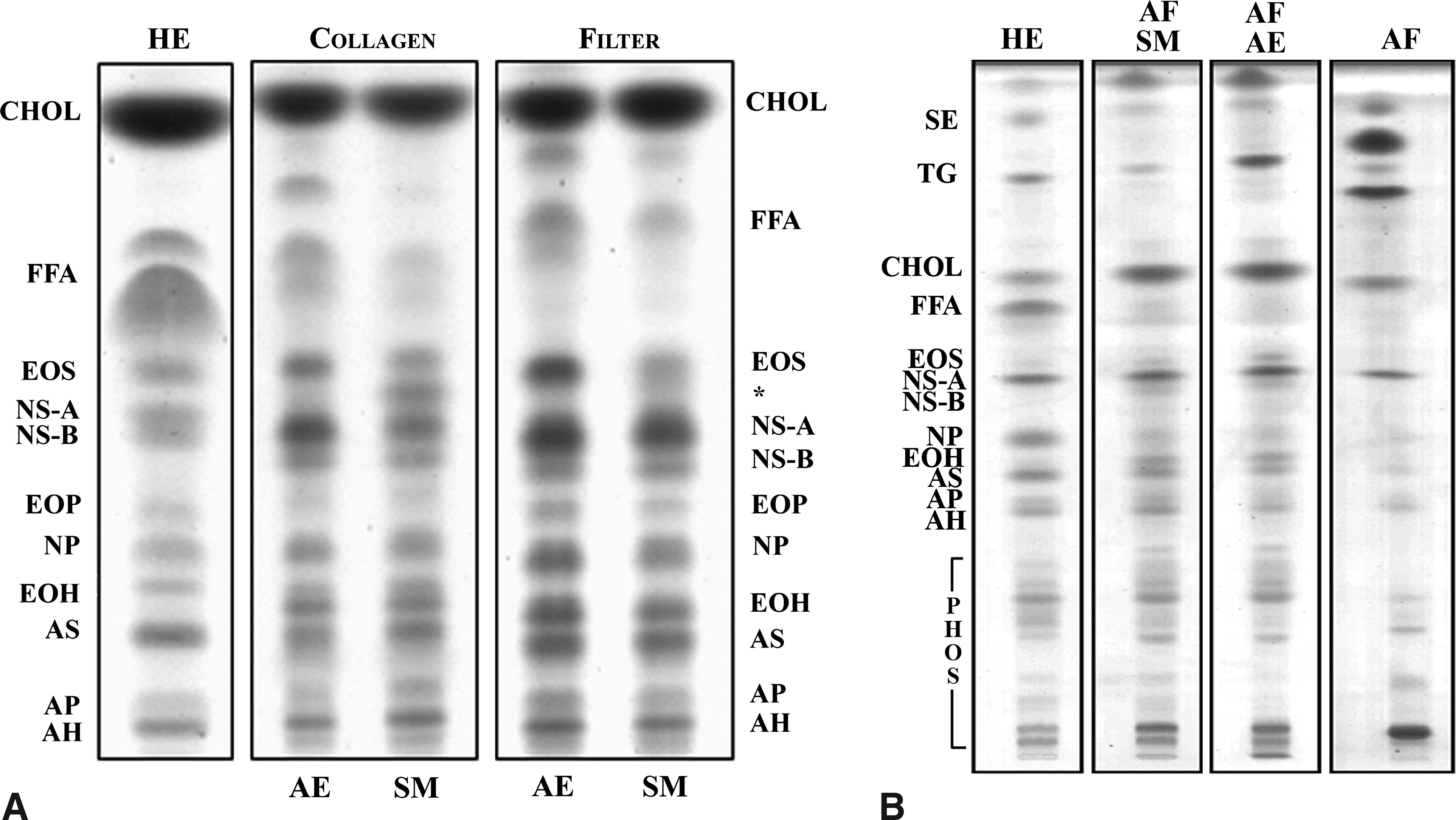
Lipid profiles of HSEs and human epidermis. Lipid profiles of human epidermis, submerged HSEs, generated on fibroblast-populated collagen matrices (Collagen) or an inert filter (Filter), and air-exposed HSEs harvested after 24 days of culturing are shown in (
Submerged and air-exposed HSEs acquire more barrier lipids as the culture period is prolonged
The lipid profiles of HSEs submerged with medium and air-exposed HSEs were quite similar at day 24. To investigate the generation of the epidermal barrier with time under submerged and air-exposed conditions, the changes in barrier lipid content were also monitored. For this purpose the HSEs were harvested after 8, 16, and 24 days of culturing. HSEs generated on a filter and harvested after 8 days were not examined, because these HSEs had not formed a SC yet.
The HSEs generated on collagen substrates and filters, under submerged and air-exposed conditions, showed an increase in all barrier lipid classes over time (Fig. 4). The HSEs generated on a collagen substrate even clearly showed the presence of ceramide NS-A and NS-B at day 8. However, the HSEs show a difference in one ceramide subclass over time. At day 8, both the submerged and air-exposed collagen HSEs showed the presence of the unidentified lipid, with an Rf value between that of ceramide EOS and NS-A. In the submerged HSEs the presence of this lipid diminishes at day 16, but increases again at day 24. Under air-exposed conditions, the collagen HSEs showed the presence of this lipid only at day 8.
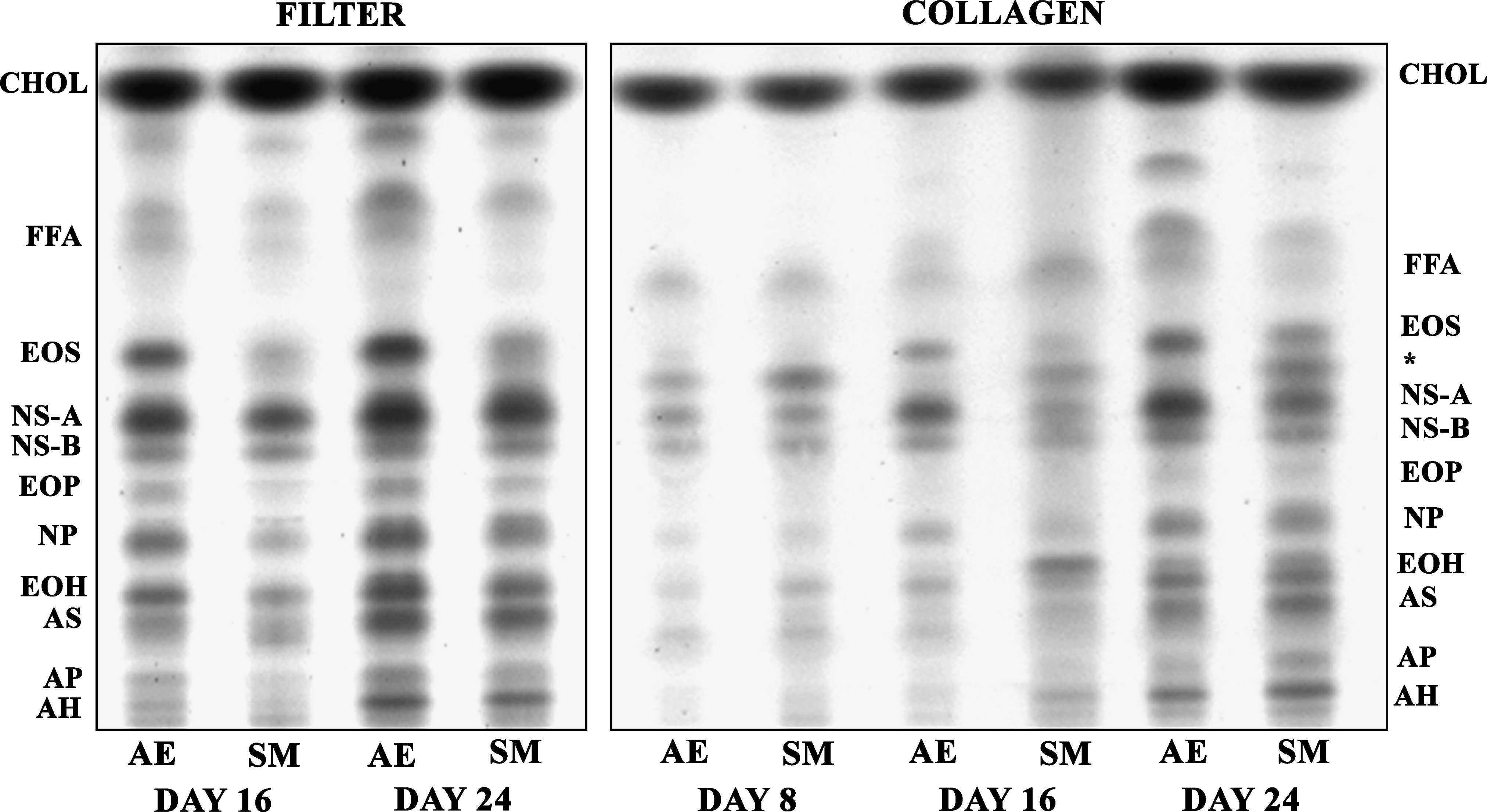
Changes in lipid profile over time. Lipid profiles of air-exposed (AE) and submerged (SM) cultures generated on fibroblast-populated collagen matrices or inert filters harvested after 8, 16, and 24 days of culturing. Lipid extracts of three HSEs grown under the same conditions were pooled. *Unidentified ceramide. CHOL, cholesterol; FFA, free fatty acids. Ceramides abbreviations: E, ester-linked fatty acid; O, ω-hydroxy fatty acid; S, sphingosine; N, non-hydroxy fatty acid; P, phytosphingosine; H, hydroxysphingosine; A, α-hydroxy fatty acid.
Analysis of the phospholipids in the submerged and air-exposed collagen HSEs at days 8, 16, and 24 showed that the phospholipids form the major lipid fraction in the early phase of epidermal development. However, their presence decreases over time (data not shown), especially from days 16 to 24. The ceramide precursors, (acyl)glucosphingolipids (AGC and GSL, respectively), are both present at day 8. The overall GSL content does not change with increasing culturing time, whereas the AGC content shows a decrease after day 16. The level of both lipid classes is higher in submerged cultures than in the air-exposed cultures.
Generating HSEs under submerged conditions increases the hydration level of SC
To investigate whether the culture conditions affect the water level and water distribution in SC, cryo-SEM images were taken of HSEs generated under submerged conditions with medium and of air-exposed HSEs harvested after 16 and 24 days. These time points were chosen because morphological examination showed a substantial SC layer. As the composition of amniotic fluid varies, the amniotic fluid-treated HSEs were not analyzed. Human skin equilibrated in an incubator at 37°C and 93% RH for 24 h served as a control, as the HSEs were also generated under these conditions.
Cryo-SEM images can be interpreted by the contrast in an image, which is created at the planed surface by the sublimation of free water. Regions with contrast correspond to areas where free water was located prior to its evaporation. Areas without free water appear as low-contrast regions in the image.
The HSEs generated on a collagen substrate and grown under submerged conditions revealed a high hydration level of the SC (Fig. 5). At day 16 the upper layers of the SC were hydrated, while the inner most layers showed low levels of water. The water present in the hydrated regions is mainly present within the corneocytes at this time point. At day 24, the submerged HSEs showed a high hydration level in the central region of the SC. The water in the hydrated central region of the SC was mainly present inside the corneocytes, but was occasionally also observed in the intercellular space. The SC of collagen HSEs grown under air-exposed conditions increases in thickness between days 16 and 24, but continues to have low water levels throughout the entire SC, as indicated by the low contrast in the images. Similar results were obtained from submerged and air-exposed HSEs generated on a filter (data not shown).
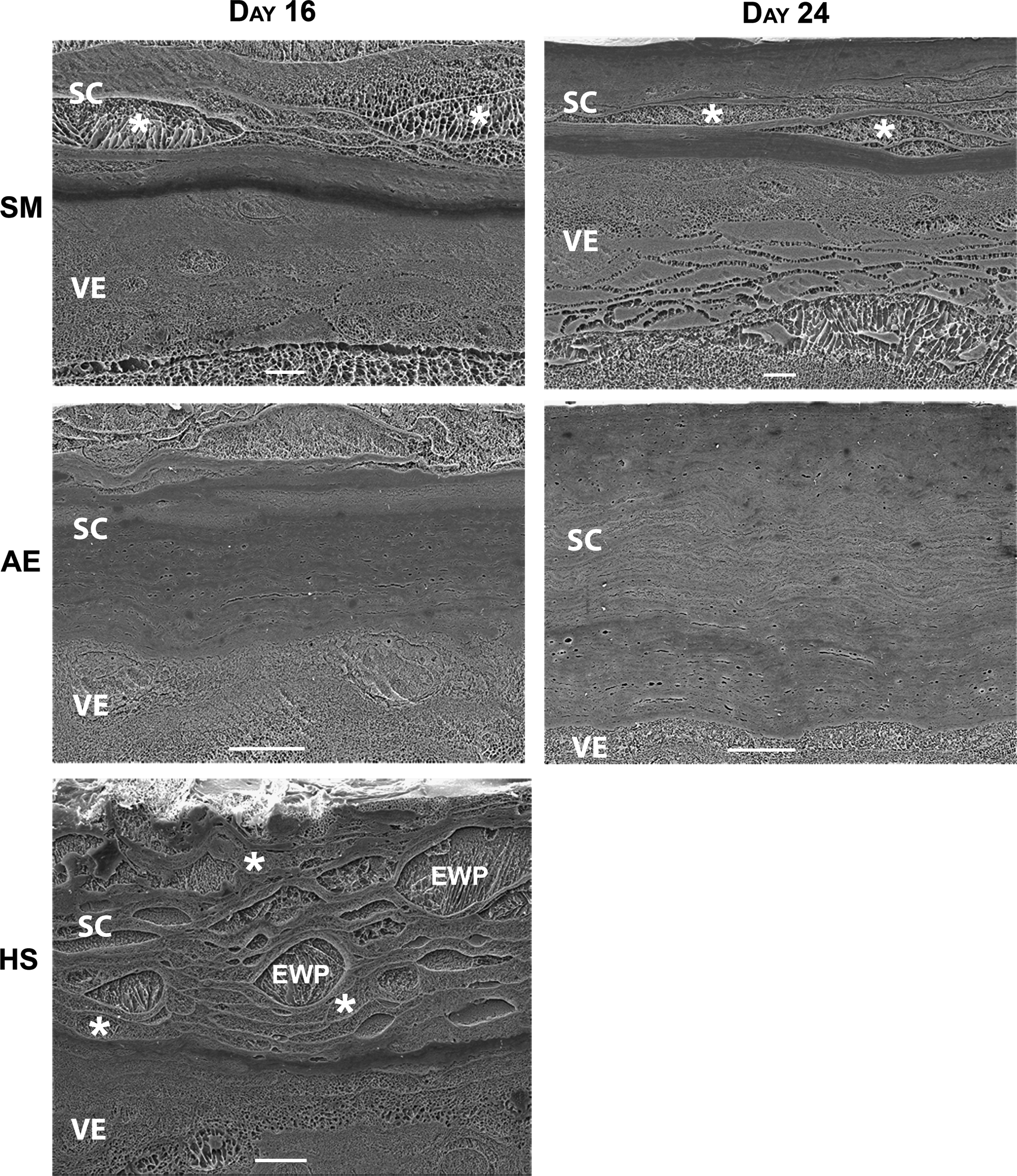
Hydration level of SC. Cryo-scanning electron microscopy images of submerged and air-exposed collagen HSEs harvested after 16 or 24 days and human skin. Human skin was equilibrated in an incubator for 24 h as a control. Scale bars represent 10 μm. AE, air-exposed; HS, human skin; SC, stratum corneum; SM, submerged; EWP, extracellular water pool; VE, viable epidermis; *Water-containing corneocyte.
Corneocytes in SC of human skin equilibrated in the incubator for 24 h appeared swollen and showed a high contrast intracellularly, indicating that these cells contained a considerable amount of free water. The corneocytes in the inner part of the SC contained almost no free water, as these cells showed almost no contrast.
Generating HSEs under submerged conditions leads to decreased NMF levels
As the NMF level is reported to be one of the factors that influences the hydration level of the SC, 20 its content in the SC of the submerged and air-exposed HSEs generated on a collagen substrate was also determined. The NMF level is expressed as the PCA/protein ratio, in which PCA is one of the main components of the NMFs. Figure 6 shows that HSEs grown under submerged conditions have a lower total PCA/protein ratio compared with the HSEs grown under air-exposed conditions. This indicates that the submerged HSEs contained less NMFs compared with the air-exposed HSEs. However, both air-exposed and submerged HSEs contained less NMF than human skin.
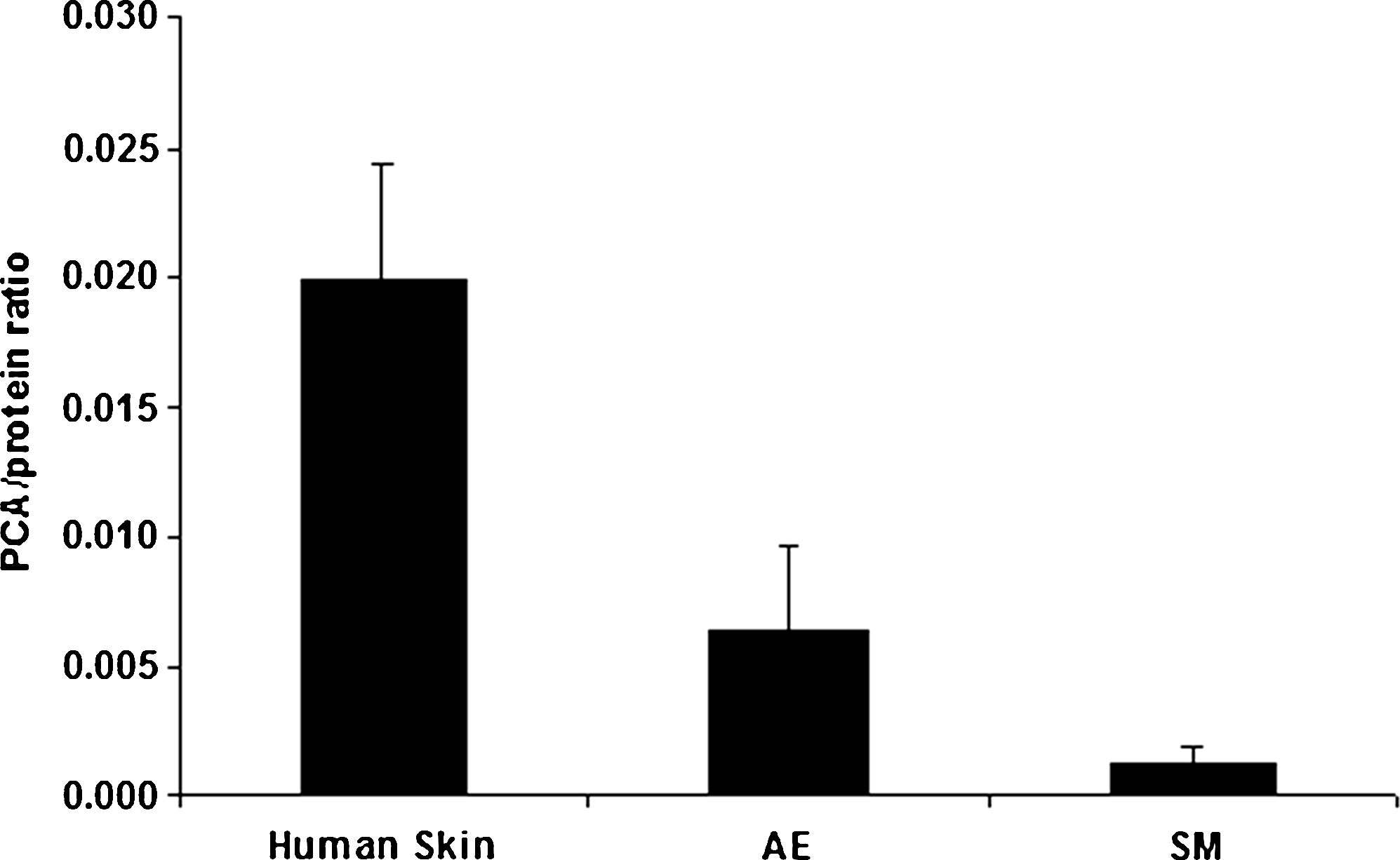
Total SC natural moisturizing factor content. Total SC natural moisturizing factor content, represented by PCA/protein ratios, of human skin, and air-exposed (AE) (n = 18) and submerged (SM) (n = 9) HSEs. All HSEs were generated on fibroblast-populated collagen matrices and harvested after 24 days of culturing. The PCA/protein ratio for human skin was obtained from Bouwstra et al. 16 The error bars represent standard deviations. PCA, pyrrolidone carboxylic acid.
Discussion
Several studies have been performed in which human keratinocytes have been cultured in Petri dishes or culture flasks under submerged conditions.3–7 However, the degree of stratification or the organization and lipid content of the SC of these submerged cultures generally showed key differences between cultures grown at the air–liquid interface or human skin. In this study we have developed a culturing method to mimic more closely the aqueous conditions found in utero than in previous studies to determine whether a fully differentiated epidermis can also be formed under submerged conditions in vitro. The most important result was that the submerged HSEs, generated on a collagen substrate or filter, were able to form a fully differentiated epidermis that contained all the strata present in human epidermis, including the SC. The submerged HSEs generated on a collagen substrate even showed the presence of all viable epidermal layers as well as a few SC layers after 8 days of culturing. Further, not only all main lipid classes were present in HSEs generated under submerged conditions, but even all the ceramide subclasses were synthesized under these conditions. To mimic the in utero environment even closer, additional studies were performed with HSEs generated on a collagen substrate that were kept submerged with amniotic fluid at the apical side. These amniotic fluid-treated HSEs showed a fully differentiated epidermis with a similar morphology, a similar expression of the differentiation markers, and same lipid profile as the HSEs that were kept submerged with culture medium. However, as the biological amniotic fluid varies in composition, it was decided to carry out most of our studies under well-defined medium conditions.
It is shown that the expression of several receptors is related to the differentiation state of keratinocytes.21,22 Basal keratinocytes in submerged cultures grown in Petri dishes or culture flasks receive nutrients only from the apical side. Therefore, the epidermal layers that develop above the basal layer in these cultures may decrease the availability of essential nutrients from the culture medium toward the basal keratinocytes, which may hamper the epidermal development. In our study, keratinocytes receive stimulating factors, which are present in the medium or secreted by fibroblasts, at the basal side in addition to the apical side. This tissue culture method mimics the in utero situation closer than the described method in Petri dishes or culture flasks. Feeding of the keratinocytes from the basal side might therefore be a crucial prerequisite to develop a fully stratified epidermis under submerged conditions. This hypothesis is in agreement with previous studies that reported an incomplete differentiation of submerged keratinocyte cultures that received nutrients only from the apical side.3–7
This study also shows that HSEs generated on an inert filter are also able to form a differentiated epidermis under submerged conditions in the absence of fibroblasts. This finding further supports our hypothesis that feeding of keratinocytes from the basal and apical side is important to generate a differentiated epidermis. However, it should be noted that the presence of fibroblasts improves the epidermal morphology. Several literature findings indicate that fibroblasts have a stimulatory effect on keratinocyte proliferation and appearance of the epidermis.9,10,23,24
Although the submerged and air-exposed HSEs show similarities in their morphology, some differences were observed concerning the expression of several differentiation markers and SC lipid composition. In the early stage of epidermal development the submerged HSEs generated on a collagen substrate had fewer viable epidermal layers compared with the air-exposed HSEs. This indicates that exposure of HSEs to air results in enhanced proliferation and differentiation of the keratinocytes in the first few days compared with the submerged conditions.
At present the function of each ceramide class remains to be elucidated. However, it is suggested that ceramide EOS is important for the barrier properties of the skin, as this ceramide promotes the formation of the characteristic 13-nm-long periodicity phase.25,26 Further, free fatty acids are also thought to play a crucial role in the tight packing of the SC lipids. 27 The lower level of ceramide EOS and free fatty acids in the submerged HSEs may therefore lead to decreased barrier properties compared with the air-exposed HSEs. The higher ceramide precursor levels in the submerged HSEs may be a result of the aqueous environment surrounding the developing epidermis.
The submerged HSEs had a much higher SC water content and a paradoxally lower NMF level compared with the air-exposed HSEs. Both the submerged and air-exposed HSEs were grown at a RH of 93%. This environment has a higher humidity level than that generally found in vivo. As the humidity of the surrounding environment directs the conversion of filaggrin to NMFs, it is plausible that the current culturing conditions lead to decreased NMF levels in the HSEs compared with human SC. 20 However, it is likely that the SC water content of the submerged HSEs nevertheless increased because of the direct contact with culture medium. This increased water level could consequently lead to a further decrease in the conversion of filaggrin to NMFs.
Conclusion
This study shows for the first time that HSEs grown under submerged conditions are able to form a fully differentiated epidermis. The submerged HSEs show similar expression of many differentiation markers and comparable lipid profiles as the air-exposed HSEs and human epidermis. During the last weeks of pregnancy, the developed epidermis is fully exposed to the surrounding amniotic fluid. The described culture method mainly mimics this stage of epidermal development and can therefore be used to study the effect of an aqueous environment on the development of the epidermis during that period. In addition, the presented culturing method may also provide information on the changes that occur in the epidermis during the transition from an aqueous to a terrestrial environment.
Footnotes
Acknowledgments
The authors thank Hendrik W. Groenink and Nazmoen Mahmood for their technical assistance.
Disclosure Statement
No competing financial interests exist.