
Abstract
This study aimed to investigate the chondrogenic transdifferentiation potential of human dermal fibroblasts (DFs) by stimulation with cartilage-derived morphogenetic protein 1 (CDMP1). Using CDMP1 (100 ng/mL) we induced human DFs at passage 5 in both monolayer and micromass culture. Chondrogenic-specific markers were detected via immunochemistry, reverse transcription–polymerase chain reaction, and Western blot analysis in the collected specimens. The expression profile of adhesion molecules including integrin α5, β1, and N-cadherin of DFs accompanying with chondrogenesis was further investigated. After 7 days of induction in monolayer culture, DFs acquired the polygonal chondrocyte-like shape with positive expression of chondrogenic-specific markers. Such a phenotypic transition of DFs was lost at 14 days. However, in micromass culture the chondrogenic transdifferentiation of DFs can be maintained even at 14 days. No chondrogenesis was detected in DFs without CDMP1 treatment under both culture conditions. By neutralization assay with blocking antibodies, it was further revealed that integrin α5 expression was in direct proportion to the degree of chondrogenic differentiation. Based on our findings, it can be ascertained that DFs are capable of transdifferentiating into a chondrogenic lineage by stimulation with CDMP1 in vitro. The integrin α5 mediating cell–cell and cell–matrix interactions is required for maintaining the chondrogenic phenotype of DFs.
Introduction
Transdifferentiation of DFs into other lineages under specific circumstances is particularly remarkable. Transdifferentiation, in definition and as reviewed by Tosh and Slack, 4 refers to the conversion of either stem cells from one germ layer to another or one fully differentiated cell type to another phenotype. Although transdifferentiation of mesenchymal stem cells (MSCs) into cells of a different embryonic lineage has been well documented, 5 investigation on the ability of DFs to transdifferentiate into other cell phenotypes is just on the horizon. It was reported that DFs were able to acquire osteocyte, hepatocyte, and macrophage phenotypes when exposed to specific differentiation cocktails, respectively.6–8 In our previous study, DFs were observed to undergo a phenotypic and functional switch to cornea stromal-like cells when implanted in a corneal stromal layer without induction. 9 These findings suggest that DFs may possess the capability of transdifferentiating into other cell types when offered with a suitable inducing agent or microenvironment.
The chondrogenic transdifferentiation of DFs, by which an alternative cell source with high proliferative potential and easy accessibility could be obtained for cartilage engineering, was proven in several reports.10,11 It was also found in our previous study that DFs underwent chondrogenic differentiation when cocultured with mature chondrocytes in micromass. 12 These results indicate that elements in the microenvironment (such as soluble factors and cell–matrix or cell–cell contact) may be responsible for chondrogenic differentiation of DFs. However, to induce chondrogenic transdifferentiation of DFs more efficiently for use as seed cells in cartilage engineering, a specific stimulator has to be identified.
As a member of bone morphogenetic protein (BMP) family, cartilage-derived morphogenetic protein 1 (CDMP1) was found to play an important role in appendicular skeleton formation during development, particularly at the early stage of limb chondrogenesis. 13 The expression of CDMP1 was mainly localized within the condensed mesenchyme of limb as well as in the forthcoming joint spaces during the developmental process. 14 In addition, the combination of CDMP1 and transforming growth factor-β1 could yield a synergistic induction effect on in vitro-cultured bone marrow MSCs. 15 We thus hypothesize that CDMP1 may function as a stimulator to induce the transdifferentiation of DFs into a chondrogenic phenotype. Hence, in this work, chondrogenic potential of human DFs was verified by subjecting DFs to CDMP1 stimulation in either monolayer or high-density micromass culture systems, and the relative mechanisms underlying such a transdifferentiation were also explored.
Materials and Methods
Isolation and culture of human DFs
Fresh human foreskin specimens were obtained from five donors (aged from 5 to 12 years) who received a routine circumcision procedure at the Shanghai Children's Hospital, China. The specimens were washed with sterile phosphate-buffered saline (PBS) and cut into small pieces (1–2 mm3), which were then digested with 0.1% dispase (Worthington, Lakewood, NJ) at 4°C overnight. The epidermal layers were removed, and the remaining dermal parts were further digested with 0.1% collagenase (Worthington) in Dulbecco's modified Eagle's medium (DMEM; Invitrogen, Grand Island, NY) for 3 h by gentle agitation at 37°C. The digested cells were then passed through a 100 μm cell strainer (BD Biosciences, San Jose, CA) and further centrifuged at 1500 rpm for 5 min. The cells were collected by resuspension in low-glucose DMEM supplemented with 10% fetal bovine serum (FBS; Hyclone, Logan, UT),
Chondrocytes isolated from discarded auricular cartilage of congenital microtia patients who received an ear reconstruction surgery at the Department of Plastic and Reconstructive Surgery of Shanghai 9th People's Hospital were cultured according to the methods described earlier 16 and similar analysis was carried out on these chondrocytes to serve as a positive control.
Flow cytometry analysis
The cells at passage 5 were collected and resuspended at a density of 1 × 106 cells in 100 μL flow cytometry buffer (PBS containing 2% FBS). The cell suspensions were then incubated for 30 min with fluorescein isothiocyanate (FITC)-conjugated anti-CD13 (Invitrogen), CD14 (Sigma-Aldrich), CD34 (BD Biosciences), CD44 (Serotec, Oxford, United Kingdom), CD90 (Serotec), CD133 (Invitrogen), CD166 (Serotec) or phycoerythrin-conjugated anti-CD29 (BD Biosciences), CD45 (Sigma-Aldrich), CD49d (Serotec), CD105 (Serotec), CD106 (Serotec), CD117 (Serotec) monoclonal antibodies, respectively. When cells were stained with nonlabeled mouse anti-Stro-1 antibody (R&D, Minneapolis, MN), FITC-conjugated goat anti-mouse immunoglobulin M (1:100; MilleGen Prologue Biotech, Labege, France) was used to amplify the signals. FITC- or phycoerythrin-conjugated isotype-matching immunoglobulins were used to determine nonspecific staining. The cells were analyzed on a fluorescence-activated cell sorting (FACS) Caliber (Becton-Dickinson, San Jose, CA), and the data were analyzed with CellQuest software (Becton-Dickinson).
Chondrogenic differentiation of DFs by CDMP1 induction in monolayer culture
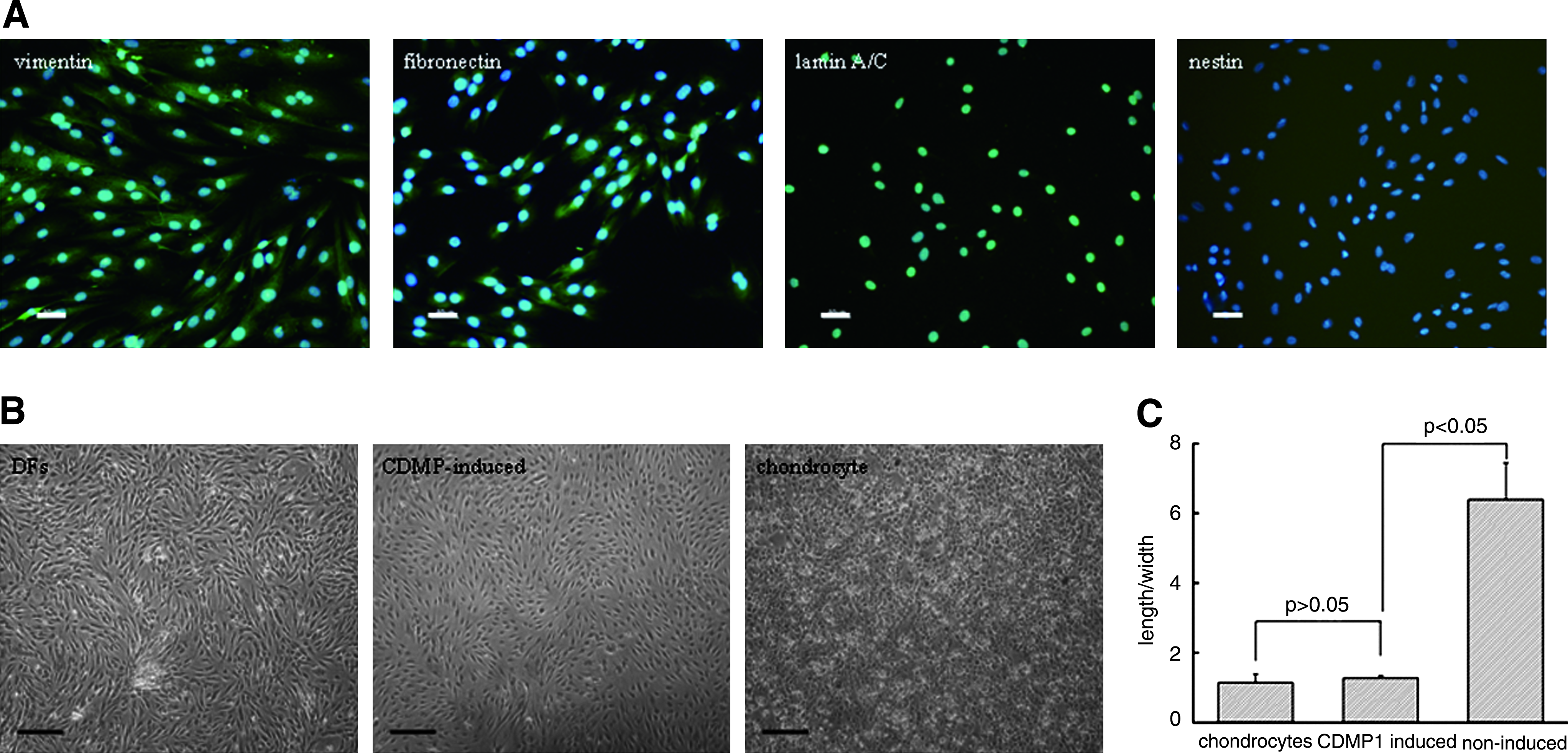
When cells at passage 5 reached confluence, the growth medium was first replaced with serum-free DMEM for 2 days and then replaced with the induction medium (DMEM containing 2% FBS and 100 ng/mL of CDMP1), which was changed every 3 days. After induction for 7 days, cell morphology was observed under phase-contrast microscope, and the maximum length and corresponding width (perpendicular to the cell axis) were measured according to the microscopic images using Image Plus software (Gaffney, SC). The average length/width ratio was then calculated in 20 randomly selected cells in each of triplicate dishes. To investigate whether the induction effect was dose dependent, CDMP1 at concentrations of 50 and 300 ng/mL was further used to compare with those of 100 ng/mL. DFs at passage 5 cultured in the growth medium were also analyzed to serve as a negative control.
Chondrogenic differentiation of DFs in micromass culture
To initiate chondrogenic differentiation of DFs in a three-dimensional environment, micromass culture was performed as described by Chang et al. 17 Briefly, DFs at passage 5 after reaching confluence were trypsinized, centrifuged, and collected by resuspension at a density of 2.0 × 107 cells/mL in the growth medium. The fresh cell suspension was then spotted in every 15 μL drop into dishes. The cells were incubated for 2 h at 37°C to allow attachment and then maintained in the induction medium.
Histology and immunostaining
DFs induced in the monolayer culture were collected at different time points and fixed with 4% paraformaldehyde for 15 min. For the immunofluorescent staining, the fixed specimens were washed and incubated with primary monoclonal antibodies (anti-vimentin, lamin A/C, fibronectin, and nestin from Santa Cruz [Santa Cruz, CA]; anti-collagen type I [COL I], collagen type II [COL II], and aggrecan monoclonal antibodies from Sigma-Aldrich) diluted in 0.5% bovine serum albumin at 4°C overnight and further washed and incubated with secondary antibody (FITC-conjugated goat anti-mouse from Oncogene, Cambridge, MA) for 30 min at 37°C. Then the specimens were washed and mounted with a solution containing propidium iodide (Sigma-Aldrich) to detect nuclei. The specimens were examined with a Nikon Optiphot fluorescence microscope. Alternatively, paraffin sections of micromass culture after incubation with COL II primary antibody were incubated with horseradish peroxidase-conjugated anti-mouse immunoglobulin G (Dako, Glostrup, Denmark) for 30 min at 37°C and then color was developed with diaminobenzidine tetrahydrochloride. Further, the sections were stained with hematoxylin and eosin and alcian blue to reveal the histological structure and cartilage matrix (glycosaminoglycans [GAGs]) deposition, respectively.
Reverse transcription–polymerase chain reaction
Total RNA was isolated from cells in each group with Trizol reagent (Invitrogen) and reverse transcribed into cDNA with reverse transcription–polymerase chain reaction (RT-PCR) kit (TakaRa Biotechnology, Dalian, P.R. China). Amplification reactions specific for the following cDNAs were performed: COL I (F-tgttcagctttgtggacctc, R-cttggtctcgtcacagatca; 236 bp); COL II (F-cttgggcacctcgggctcctttag, R-tccccggcactcctggcactgat; 510 bp); aggrecan (F-tcctggaagctcttctcact, R-atgcccaagactaccagtgg; 510 bp); sex determining region Y-related high mobility group-box gene9 (SOX9) (F-gaacgcacatcaagacggag, R-tctcgttgatttcgctgctc; 631 bp); β-actin (F-atcatgtttgagaccttcaa, R-catctcttgctcgaagtcca; 318 bp) and β-actin for SOX9 (F-aggagcagactgtggatgg, R-gaacccggaatgggtgat; 580 bp). The reactions were performed in T3 thermocycler (Biometra, Goettingen, Germany). The amplified products were separated on 1.5% agarose gels and visualized by ethidium bromide (Sigma-Aldrich).
Quantitative real-time PCR
Total RNA was extracted as described earlier. The RNA concentration of the extract was determined from optical absorbance at 260 nm and the RNA was reverse transcribed into cDNA. PCRs were performed and monitored in T3 thermocycler (Biometra). For quantitative real-time PCR (qPCR), markers of chondrogenic (COL II, aggrecan, SOX9, and COL I) and osteogenic (collagen type X [COL X] [F-gcctgaagggagaaagag, R-atcctggaatgcctggtg; 431 bp], alkaline phosphatase [ALP] [F-acgtggctaagaatgtcatc, R-ctggtaggcgatgtcctta; 476 bp], osteocalcin [OCN] [F-caaaggtgcagcctttgtgtc, R-tcacagtccggattgagctca; 150 bp], and core-binding factor alpha 1 [cbfα-1] [F-gtcttacccctcctacctga, R-tgcctggctcttcttactga; 184 bp]) differentiation were analyzed using quantitative real-time amplification system (MxPro-Mx3000P; Stratagene, La Jolla, CA). The reaction mixture contained SYBR green (Molecular Probes, Foster City, CA), and β-actin housekeeping gene served as the control. Relative expression levels for each gene of interest were calculated by normalizing the quantified cDNA transcript levels (cycle threshold) to that of β-actin. Three independent experiments were performed.
Western blot detection
Total cell lysate was prepared from different cultures at various time points by extracting proteins with a lysis buffer (50 mM Tris-HCl, 150 mM NaCl, 0.2 g/L NaN3, 1 g/L sodium dodecyl sulfate, 0.1 g/L aprotinin, 10 g/L NP-40, 5 g/L sodium deoxycholate, 0.1 g/L phenylmethanesulfonyl fluoride (PMSF), pH 8.0). Proteins of 20 μg (determined by Pierce BCA protein assay kit, Rockford, IL) were size-fractionated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane (Millipore, Billerica, MA). After being blocked with PBS containing 2% nonfat dry milk and 3% bovine serum albumin for 2 h at room temperature, the membranes were incubated with 1:1000 diluted primary monoclonal antibodies at 4°C overnight and then with 1:2000 diluted IRDye800-conjugated goat anti-mouse secondary antibody (Rockland, Gilbertsville, PA) for 1 h at room temperature. Signals were detected using the Odyssey Infrared Imaging System (LI-COR Biosciences, Lincoln, NE). For chondrogenesis detection, antibodies used were anti-COL I, COL II, aggrecan, SOX9, and β-actin (Sigma-Aldrich). For detection of cell adhesion molecules, antibodies of mouse anti-human N-cadherin (Sigma-Aldrich), integrin α5, and integrin β1 (both from Chemicon, Billerica, MA) were used.
Neutralization assay with cell adhesion-specific antibodies
To further identify the role of integrin α5 during chondrogenesis, monoclonal anti-human integrin α5 (10 ng/mL; Chemicon) was added in culture medium to neutralize the respective cell adhesion molecule. After 7 days, CDMP1 was added in the medium for 14 days in both monolayer culture and micromass culture. Then specimens were collected for Western blot analysis.
Statistical analysis
Data were presented as mean ± standard deviation from three or more experiments. The statistical significance was assessed by paired Student's t-test with SPSS 11.0 software (Chicago, IL), and a p-value of less than 0.05 was considered statistically significant.
Result
Characterization of human DFs
The immunofluorescent staining results revealed that DFs were positive for vimentin, fibronectin, and lamin A/C expression, but not for nestin (Fig. 1A). Thus, a mesenchymal origin of these cells could be confirmed, while the possibility of contamination by other dermis-derived multipotent stem cells could be ruled out as reported by Toma et al. 18 To further characterize DFs, the expression profile of surface antigens was examined by FACS. As shown in Table 1, cells displayed a phenotypic single population characterized by the expression of CD13 (71.5% ± 0.7%), CD29 (78.7% ± 0.6%), CD44 (64.0% ± 1.2%), CD49d (66.7% ± 0.8%), CD166 (79.7% ± 1.1%), and a trivial level of CD105 (6.3% ± 0.5%). Additionally, homogenous expression of CD90, which was identified as a fibroblast-specific antigen epitope, 19 was detected to reach a high level (72.6% ± 0.8%). In contrast, either hematopoietic stem cell subpopulations (CD14, CD34, CD45, and CD133) or MSC-related markers (CD117 and Stro-1) could not be observed.
(
Chondrogenic differentiation of DFs by CDMP1 induction in monolayer culture
To evaluate the chondrogenic differentiation potential of DFs, cells at passage 5 were cultured in monolayer in the presence of CDMP1 (100 ng/mL) for 7 days. Interestingly, the morphology of these DFs changed gradually from spindle-like appearance to a chondrocyte-like polygonal shape, which was similar to that of human normal chondrocytes (Fig. 1B). However, DFs cultured in the growth medium without CDMP1 remained in spindle shape. By Image Plus software analysis, it was found that the length/width ratio decreased significantly from 6.39 ± 1.25 of DFs before induction to 1.27 ± 0.07 of DFs at 7 days postinduction (p < 0.05), whereas no significant difference was found between the induced DFs and the normal chondrocytes (1.14 ± 0.25, p > 0.05; Fig. 1C).
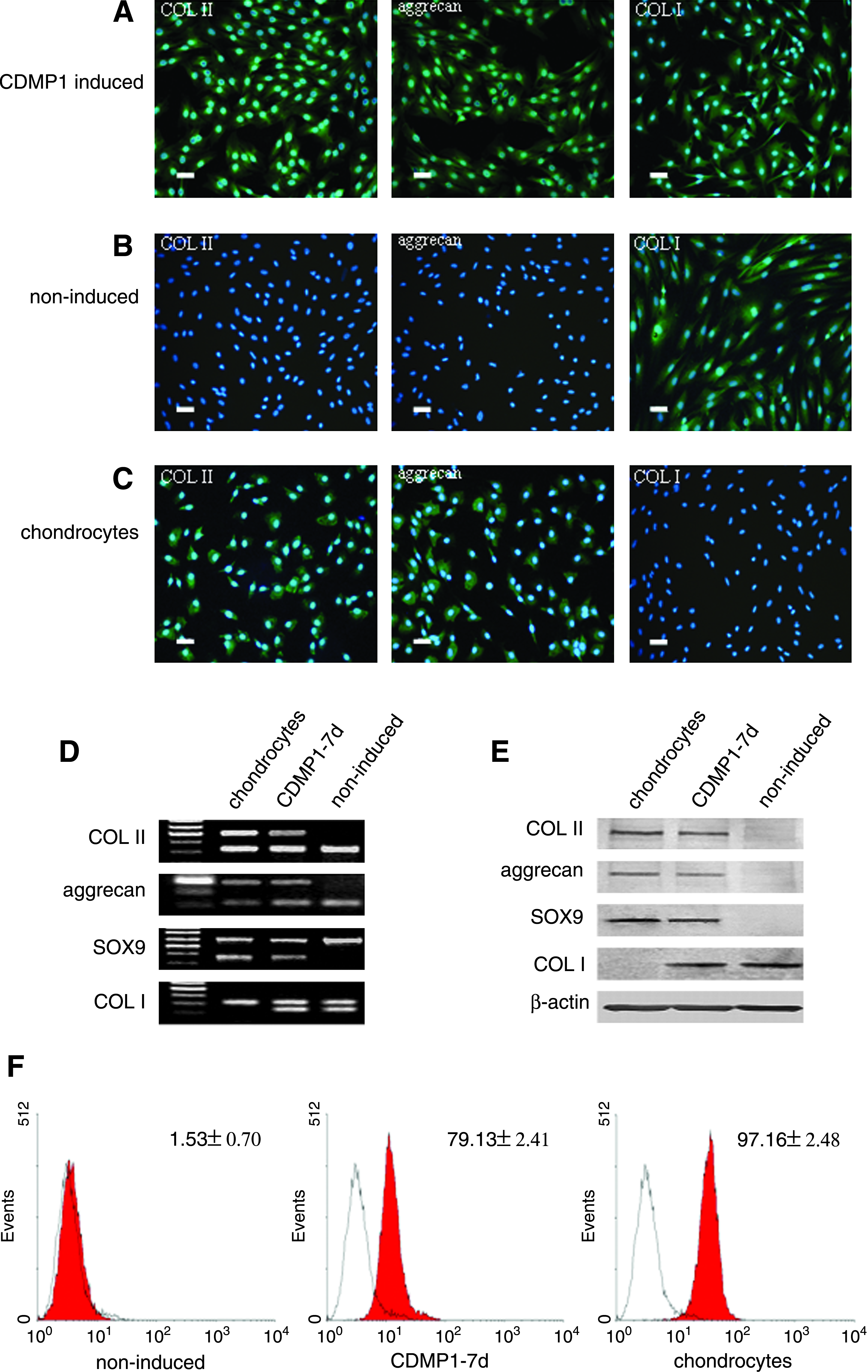
The normal DFs do not express COL II and aggrecan as revealed by immunofluorescent staining (Fig. 2B). But expression of COL II and aggrecan was visualized in the induced DFs (Fig. 2A). The expression of COL I was observed in both normal and induced DFs. In the normal chondrocytes, the expression of COL II and aggrecan was detected as expected, whereas no COL I could be observed in these cells (Fig. 2C).
(
In addition, the expression of specific markers for chondrogenesis was also identified in DFs treated with CDMP1 for 7 days, including COL II, aggrecan, and SOX9, as revealed both by RT-PCR and Western blot analysis (Fig. 2D, E). Similar to the results of immunofluorescent staining, none of those markers were observed in the noninduced DFs. On the other hand, both the induced and noninduced DFs were accompanied with COL I expression. The results of RT-PCR and Western blot were in well agreement.
Further, by FACS analysis, it was found that the percentage of SOX9-positive cells for DFs was extremely enhanced from 1.53% ± 0.70% to 79.13% ± 2.41% after induction with CDMP1 for 7 days, whereas the SOX9 expression for normal chondrocytes was 97.16% ± 2.48% (Fig. 2F).
Dose dependence of CDMP1 stimulatory effect on transdifferentiation of DFs
The expression of COL II, aggrecan, and SOX9 in DFs induced by different doses of CDMP1 was compared using qPCR measurement (Fig. 3). No expression of COL II, aggrecan, and SOX9 except COL I could be detected in cells induced with CDMP1 at a concentration of 50 ng/mL. However, the amount of expressed COL II, aggrecan, and SOX9 was impressively enhanced when CDMP1 was increased to 100 ng/mL, although the corresponding expression of COL I was decreased. With further increase in CDMP1 concentration to 300 ng/mL, the expression of COL II, aggrecan, and SOX9 was increased only to approximately 1.1-fold (COL II: 1.104-fold; aggrecan: 1.099-fold; SOX9: 1.078-fold), respectively, of those of cells treated with 100 ng/mL. Thus, a CDMP1 concentration of 100 ng/mL was fixed to induce chondrogenic differentiation of DFs in the following study.
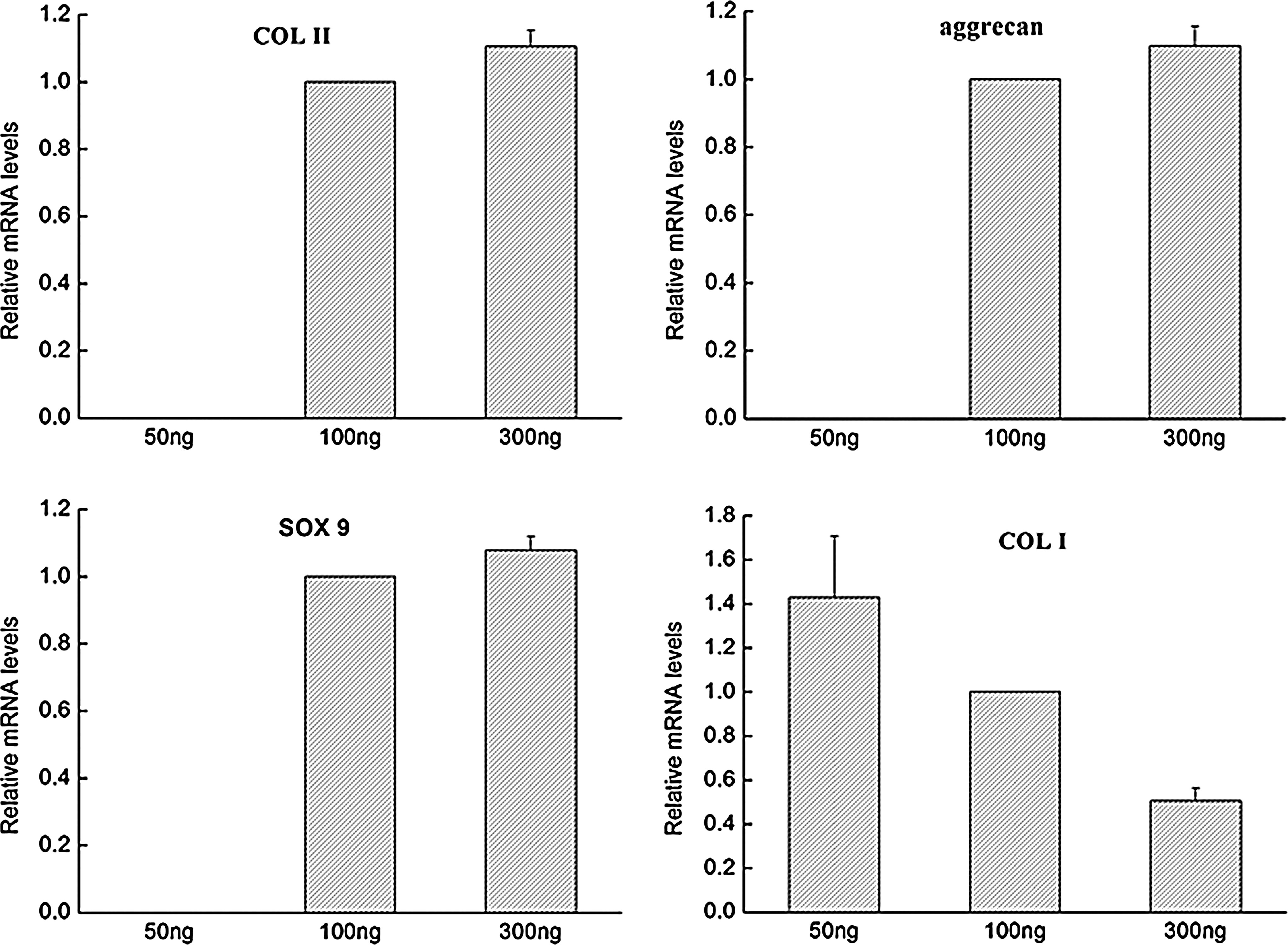
Dose dependence of CDMP1 on chondrogenic differentiation of DFs determined by real-time PCR analysis. DFs were treated with CDMP1 for 7 days at concentrations of 50, 100, and 300 ng/mL.
Maintenance of the chondrogenic phenotype of induced DFs in micromass culture
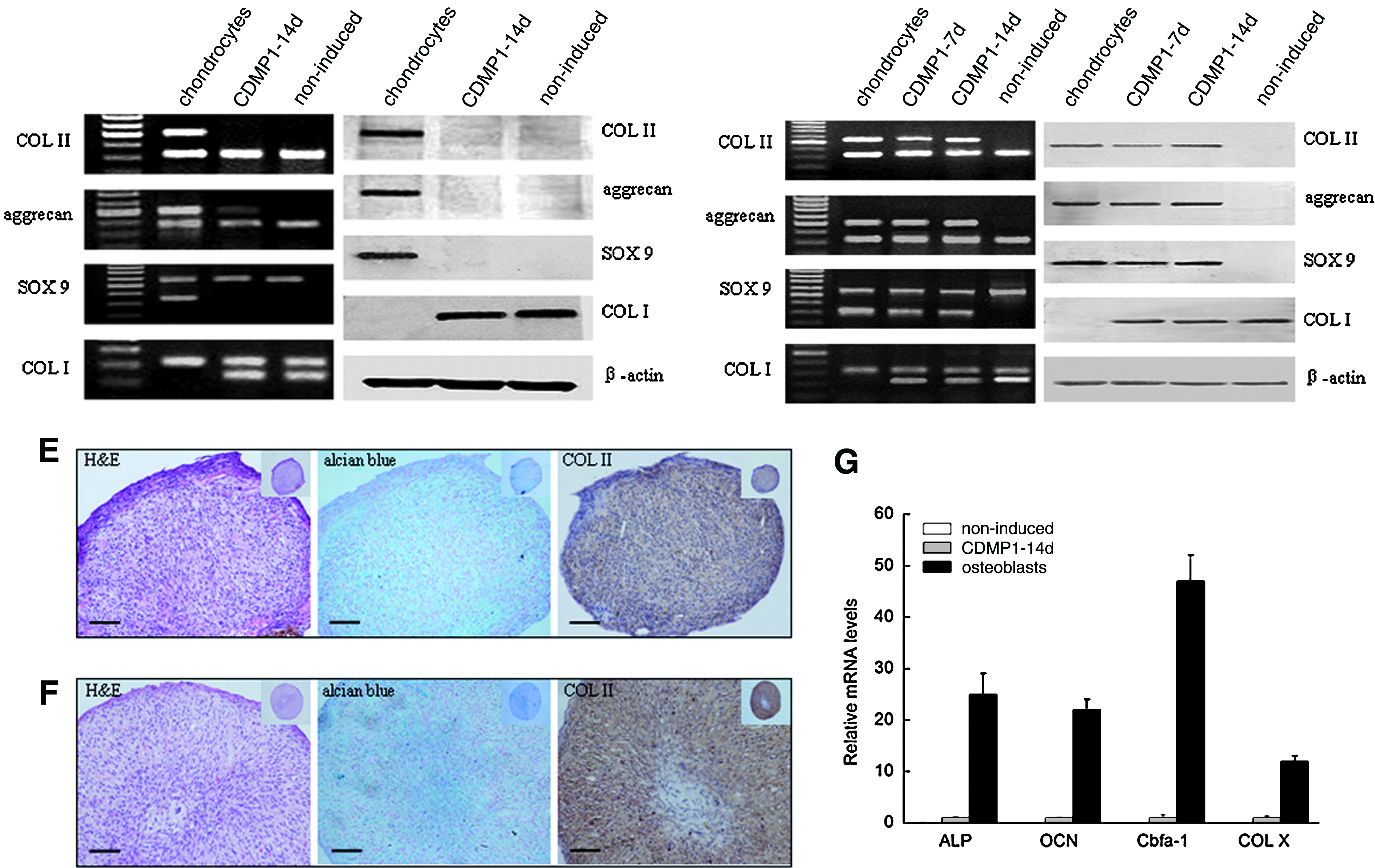
To further gain insights on the chondrogenic differentiation process of DFs, we extended the duration of CDMP1 stimulation (100 ng/mL) on DFs in monolayer culture to 14 days. Unfortunately, expression of COL II and SOX9 could no longer be detected by RT-PCR analysis, and only a trivial expression of aggrecan remained (Fig. 4A). Similarly, no expression of COL II, aggrecan, and SOX9 could be observed by Western blot analysis (Fig. 4B). Thus, the loss of chondrogenic phenotype of these DFs could be ascertained, indicating that monolayer culture is not a favorable microenvironment for CDMP1-treated DFs to maintain their chondrogenic phenotype. It was demonstrated by many investigations on chondrogenesis of mesenchymal progenitor cells that a high-density culture manner such as micromass or pellet is essential for the chondrogenic differentiation of these cells.17,20,21 Thus, to maintain the chondrogenic phenotype of CDMP1-treated DFs, DFs of passage 5 were then subjected to micromass culture and it was later found that cell aggregates could be generated with their sizes maintained stable in the culture medium. As DFs without CDMP1 stimulation failed to generate cell nodules, only histological results from CDMP1-treated group were presented, and the noninduced cells in micromass culture were also collected to serve as the negative control for RT-PCR and Western blot. Expression of COL II, aggrecan, and SOX9, which were lost by the induced DFs in monolayer culture at 14 days, was highly maintained by DFs in micromass culture at 14 days as shown by both RT-PCR (Fig. 4C) and Western blot (Fig. 4D) examination.
(
The morphology of the induced DFs in high-density culture was further revealed by hematoxylin and eosin and immunochemistry staining. After 7 days in micromass culture with CDMP1 induction, DFs deposited GAG and COL II as revealed by alcian blue and immunochemistry staining, respectively (Fig. 4E). When the induction time was extended to 14 days, the cell aggregates became quite mature characterized by the formation of lacunae structures with chondrocyte-like cells located inside. The expression of COL II was observed in most of the cells within the cell aggregates (Fig. 4F).
To exclude the possibility of osteogenesis, we therefore examined the expression of osteogenic-related genes including ALP, OCN, and cbfα-1 as well as hypertrophic-related COL X in induced DFs under micromass culture for 14 days. Compared with osteoblasts, no obvious expression of these genes could be detected by qPCR in either induced or noninduced DFs (Fig. 4G).
The role of cell adhesion molecules in chondrogenesis of DFs
To evaluate whether the chondrogenesis of DFs is related to the expression of cell adhesion molecules, Western blot was carried out to detect N-cadherin, integrin α5, and β1 in the induced DFs. In monolayer culture (Fig. 5A), the expression of N-cadherin, which mediates cell–cell interactions, was extremely low in chondrocytes, compared with that in DFs. With CDMP1 induction, its expression by DFs was reduced with the proceeding of chondrogenesis. As COL II expression by the induced DFs was high when assayed at 7 days in monolayer culture and then declined to an undetectable level at 14 days, the temporal expression profile of N-cadherin in monolayer culture was in an opposite way as that of COL II. However, the expression profile of integrin α5 was similar to that of COL II. There was no obvious change in the expression of integrin β1 during the whole period. In micromass culture (Fig. 5B), a gradual decrease in N-cadherin and increase in integrin α5 was observed with the process of chondrogenesis, while an almost constant expression of integrin β1 was similarly observed as that in monolayer culture.
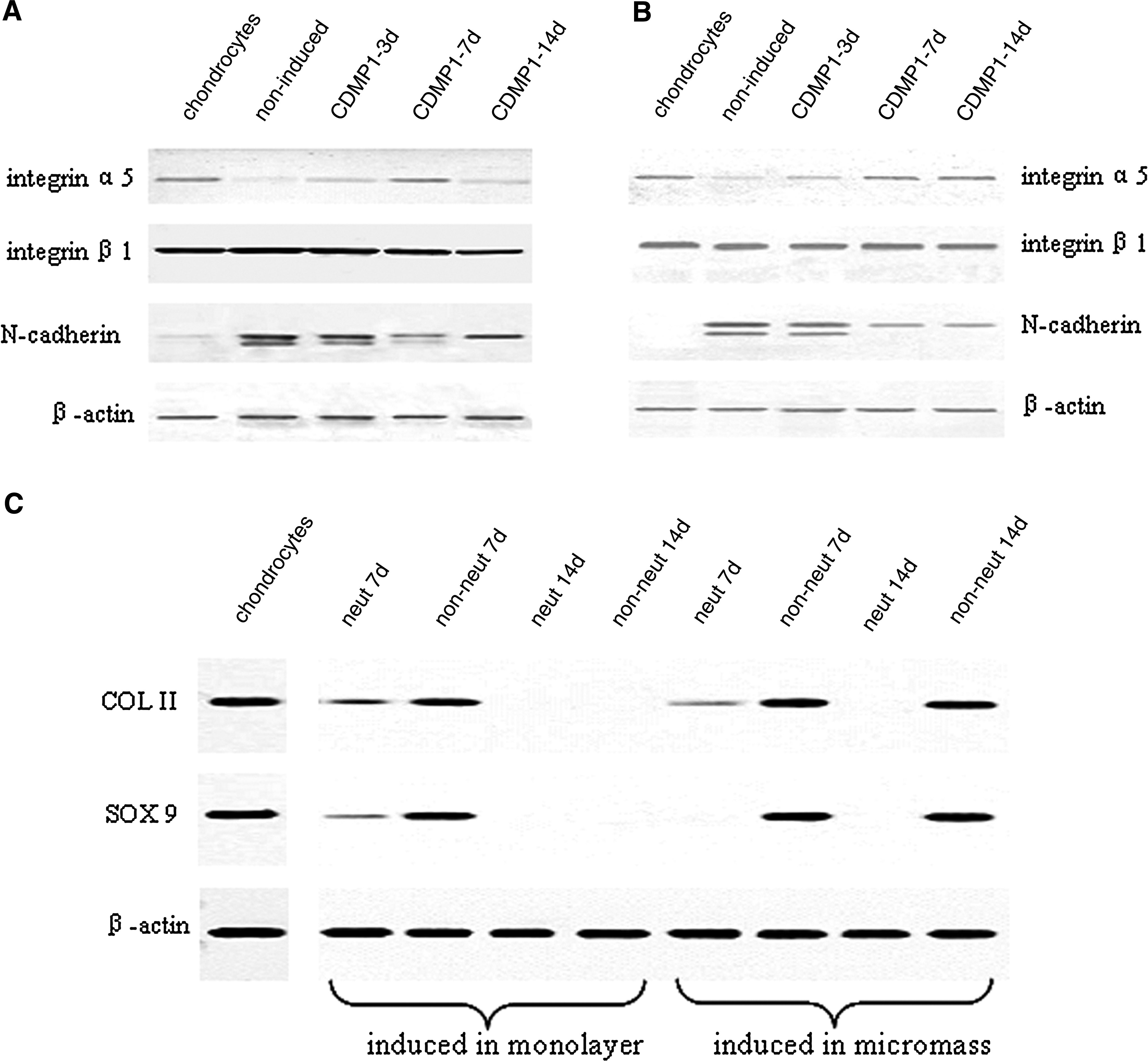
(
To further reveal the role of cell adhesion molecules, Western blot was carried out to detect COL II and SOX9 after integrin α5 was neutralized as the expression of this molecule was in a parallel way as the proceeding of chondrogenesis (Fig. 5C). In monolayer culture, the expression of COL II and SOX9 was extremely inhibited during chondrogenesis in the neutralized group, compared with that in CDMP1-induced nonneutralized group. In micromass culture, similar phenomenon was observed as that in monolayer culture.
Discussion
CDMP1, also known as BMP14, is the human homolog of mouse growth and differentiation factor 5, which belongs to the BMP superfamily. 22 During embryonic early limb development, expression of CDMP1 is found to be restricted to the primordial cartilage of appendicular skeleton. 13 Besides its involvement in osteochondrogenesis development, CDMPs were also detected in early postnatal, adult healthy, and diseased osteoarthritis articular cartilage, 14 indicating a promising role of CDMPs in regulating and maintaining chondrogenic phenotype. Therefore, inspired by these studies, we administered recombinant CDMP1 to human DFs and the transdifferentiation potential of DFs toward chondrogenic lineage was tested. It was shown that under stimulation with CDMP1, terminal differentiated human DFs were capable of differentiating into chondrogenic phenotype in vitro.
To eliminate the potential contamination of multipotent precursors, a serial passage of fibroblasts was carried out with the phenotypic characterization being preformed at the initial stage of this study. Results of FACS analysis correlated well with findings of other studies, which confirmed a high purity of DFs with positive expression of relative CD markers instead of hematopoietic as well as MSCs' relevant ones.8,23,24 In combination with the positive immunofluorescent staining of lamin A/C, a marker of terminally differentiated cells, 25 a fibroblast phenotype in the cell population could be identified and detectable contamination of dermis-derived MSCs could be ruled out.
Morphologically, we found that in the presence of CDMP1, the spindle-shaped fibroblasts gradually switched to the cuboidal chondrocyte-like shape, which was quantitatively confirmed by software evaluation. Moreover, clear expression of COL II and aggrecan, the two main chondrogenic marker genes, by DFs treated with CDMP1 was demonstrated by RT-PCR analysis, immunofluorescent staining, and Western blot detection. Further, we found that expression of SOX9 was initiated in DFs with addition of CDMP1. FACS analysis revealed that nearly 80% of induced DFs were SOX9 positive. Taken together, these results suggest that DFs cultured in monolayer with CDMP1 for 7 days acquired a chondrogenic phenotype.
During chondrogenic differentiation of embryonic stem cells 26 and adult MSCs 27 in vitro, cell–cell contact was observed to play a critical role. In this study, we found that chondrogenically differentiated DFs in monolayer culture failed to express COL II, aggrecan, and SOX9 when treatment with CDMP1 was prolonged to 14 days, indicating that the microenvironmental lack of cell–cell contact might be responsible for the induced DFs to withdraw their chondrogenic phenotype. Thus, micromass culture was adopted to culture these cells with CDMP1. As a result, such a high-density culture system that offered more cell–cell contact resembling embryogenesis leads to the maintenance of chondrogenic phenotype of DFs treated with CDMP1 for 14 days. Cartilaginous-like structure with the formation of typical lacunae and deposition of COL II and GAG was observed in such a culture system. More importantly, expression of SOX9 was observed to be at a relatively high level even at day 14. Hence, these results indicate that transdifferentiation of DFs into chondrogenic lineage takes place in a similar way as those adopted by embryonic stem cells (ES cells) and adult MSCs in their chondrogenic differentiation.
In the process of endochondral ossification, mature chondrocytes undergo terminal hypertrophic differentiation characterized by COL X expression, 28 followed by initiation of osteogenic-related genes such as ALP, OCN, and cbfα-1.29,30 To exclude an accompanying endochondral ossification process, we invested whether hypertrophic-related COL X, osteogenic-relevant ALP, OCN, and cbfα-1 were expressed by DFs treated with CDMP1 for 14 days in micromass culture. Expression of COL X cannot be detected, suggesting that the chondrogenic transdifferentiation was not accomplished with hypertrophic maturation. Only similar baseline levels in expression of cbfα-1, ALP, and OCN were detected in CDMP1-treated cells to those in noninduced ones without any observable increase. Thus, such chondrogenic transdifferentiated DFs could be safely used for cartilage engineering without initiating ossification in the duration of 14 days.
During chondrogenesis, mesenchyme condensation is a well-programmed stepwise process involving extensive cell–cell interactions in the early stage and cell–matrix interaction in the late stage in which adhesion molecules including N-cadherin, integrin α5, and β1 are believed to play crucial roles in maintaining chondrogenic phenotype. 31 N-cadherin was observed to be expressed in the early period of chondrogenic-induced MSCs and its expression decreased as chondrogenesis proceeded. 32 In this study, expression of N-cadherin was observed at 3 days and diminished at 7 days in CDMP1-treated DFs in either monolayer or micromass cultures, indicating that the involvement of N-cadherin in cell–cell interaction at the early stage of chondrogenic transdifferentiation of DFs occurred in a similar manner as that in cartilage morphogenesis as well as in chondrogenesis of MSCs. We then also investigated the expression of integrin α5 and β1 in CDMP1-treated DFs, as these two were the most prominently expressed integrins in mature chondrocytes and responsible for a signal switch from cell–cell to cell–matrix interactions in late stage of chondrogenesis. 33 We found that both induced and noninduced DFs expressed integrin β1 subset at a high and constant level in the duration of experiment. However, expression of integrin α5 subunit was found to be in a direct proportion to chondrogenic differentiation. When this molecule was neutralized, the consequent expression of COL II and SOX9 was significantly blocked in CDMP1-induced DFs cultured in either monolayer or micromass. Thus, the above results suggest that chondrogenic transdifferentiation of DFs induced by CDMP1 was closely mediated by integrin α5 expression.
In summary, this study demonstrated that terminal differentiated human DFs are capable of transdifferentiating into chondrogenic lineage by stimulation with CDMP1 in vitro. The microenvironment facilitating cell–cell and cell–matrix interactions is further required for maintaining chondrogenic phenotype of CDMP1-treated DFs. And such a phenotype maintenance is evidently mediated by integrin α5 expression. Modulation of the chondrogenic differentiation of DFs may further pave the way for application of DFs as an alternative cell source for cartilage tissue engineering and gain more insight into the regulation of cartilage tissue repair.
Footnotes
Acknowledgments
This study was supported by the Natural Science Foundation of China (grant no. 30970743), the Science and Technology Commission of Shanghai Municipality Foundation (grant no. 09xd1402800), and the National “973” Project Foundation (grant no. 2005CB522700).
Disclosure Statement
No competing financial interests exist.