
Abstract
This study investigated the effect of acemannan (Aloe vera gel polysaccharide) on dentin formation. Primary human dental pulp cells were treated with acemannan. New DNA synthesis, bone morphogenetic protein-2, alkaline phosphatase activity, dentin sialoprotein expression, and mineralization were determined by [3H]-thymidine incorporation, enzyme-linked immunosorbent assay, biochemical assay, western blotting, and Alizarin Red staining, respectively. Then the upper first molars of 24 male Sprague Dawley rats were intentionally exposed and capped with either acemannan or calcium hydroxide. At day 28, the teeth were histopathologically examined and evaluated for the degree of inflammation, dentin bridge formation, and pulp tissue organization. The results revealed that acemannan significantly increased pulp cell proliferation, bone morphogenetic protein-2, alkaline phosphatase activity, dentin sialoprotein expression, and mineralization, compared with the untreated group. The acemannan-treated group also exhibited a complete homogeneous calcified dentin bridge and good pulp tissue organization, whereas neither was detected in the calcium hydroxide-treated and sham groups. In the acemannan-treated group, either mild or no inflammation was found, whereas the other groups had various degrees of inflammation. The data suggest that acemannan promotes dentin formation by stimulating primary human dental pulp cell proliferation, differentiation, extracellular matrix formation, and mineralization. Acemannan also has pulpal biocompatibility and promotes soft tissue organization.
Introduction
Generally, dental pulp healing consists of inflammatory, formative, and remodeling phases. In the dentin formation phase, dental pulp cells undergo proliferation and differentiation into odontoblast-like cells. They secrete dentin matrix proteins and induce dentin mineralization. 9
Aloe vera, also known as Aloe barbadensis Miller, has long been used as an herbal medicine for soft tissue treatment such as burns and wounds. 10 Acemannan, a major polysaccharide of A. vera gel, is recognized for its cytocompatibility and use as a wound-healing inducer. Recently, our team reported the effect of acemannan on oral wound healing in animals by stimulating gingival fibroblast proliferation and collagen and vascular endothelium growth factor expressions. 11 However, the effect of acemannan on hard tissue regeneration, such as bone and dentin, has not yet been investigated. This study reveals the effect of acemannan on dentin formation both in vitro and in vivo.
Materials and Methods
Preparation, purification, and characterization of acemannan
A. barbadensis Miller was obtained from a local herbal supplier in Thailand, and the specimen (AC-05-2007) was deposited at the Oral Biology Research Center, Faculty of Dentistry, Chulalongkorn University. It was identified as A. vera by Assoc. Prof. Dr. Chaiyo Chaichantipyuth, Department of Pharmacognosy, Faculty of Pharmaceutical Sciences, Chulalongkorn University.
Acemannan was extracted from fresh A. vera pulp gel by homogenization, centrifugation, and alcohol precipitation, as previously described.11–13 The molecular weight was analyzed using high-performance liquid chromatography with a reflective index detector (Shimadzu, Kyoto, Japan). Separation was performed with a Shodex Sugar KS-804 column and compared with Shodex standard P-82 (Showa Denko K.K., Yokohama, Japan). Monosaccharide compositions were analyzed using gas chromatography–mass spectroscopy and 13C nuclear magnetic resonance spectroscopy, as previously described.13,14 The data obtained were consistent with previous results showing that the polysaccharide extracted from fresh A. vera gel was acemannan.13,14 The yield of acemannan extraction was about 0.2%.
For the cell culture, acemannan was solubilized in distilled water and sterilized by autoclave. For the animal study, solubilized acemannan was frozen and then lyophilized for 24 h to create an acemannan sponge.
Cell culture
This study was approved by the Ethics Committee of the Faculty of Dentistry, Chulalongkorn University (no. 16/2007).
Primary human dental pulp cells (PDPCs) were isolated from explanted healthy pulps of impacted third molars. PDPCs were pooled and cultured in a growth medium: Dulbecco's modified medium supplemented with 10% fetal bovine serum, 1%
DNA synthesis assay
DNA synthesis was investigated by [3H]-thymidine incorporation assay, as previously described. 11 Briefly, PDPCs (6 × 104 cells/well) were seeded in 24-well plates containing growth medium and cultured until 70–80% confluence. The cells were treated with different concentrations of acemannan for 24 h. During the last 4 h of incubation, cells were labeled with 0.25 μCi/well of [3H]-thymidine (Amersham Biosciences, Little Chalfont, United Kingdom). The cells were washed three times with phosphate-buffered saline (PBS), fixed with 10% trichloroacetic acid for 20 min, and solubilized in 0.5 M NaOH overnight. After neutralizing with 0.5 M HCl, the lysate was thoroughly mixed with scintillation fluid (OptiPhase HiSafe; Fisher Scientific, Milton Keynes, United Kingdom). [3H]-thymidine incorporation was determined using a liquid scintillation counter (Wallac, Turku, Finland).
Determination of bone morphogenetic protein-2 expression
PDPCs (8 × 104 cells/well) were seeded in 24-well plates. The cells were treated with acemannan in mineralizing medium. Test media were replaced with fresh media every 3 days. The conditioned media were collected for measurement of the amount of bone morphogenetic protein-2 (BMP-2) with an ELISA kit (R&D Systems, Minneapolis, MN). The detection limit for BMP-2 was 11 pg/mL.
Alkaline phosphatase activity assay
Alkaline phosphatase (ALPase) activity was determined at days 3 and 9 of incubation. The cell layer was washed twice with PBS and incubated with glycine buffer (100 mM glycine, 2 mM MgCl2, pH 10.5) containing 0.35 mg/mL p-nitrophenylphosphate (Sigma-Aldrich) at 30°C for 50 min. 16 After terminating the reaction with 1 M NaOH, p-nitrophenol (p-NP) production was measured at 405 nm using a spectrophotometer. ALPase activity was reported in terms of p-NP production and normalized with total cellular protein (nmol p-NP/min/mg protein).
Western blot analysis
The whole cell lysate was extracted by RIPA lysis buffer (Thermo Scientific–Pierce Biotechnology, Rockford, IL). The protein concentration was determined using Lowry method (Bio-Rad Protein Assay; Bio-Rad Laboratories, Hercules, CA). Protein (50 μg) of each sample was resolved by 10% sodium dodecyl sulphate–polyacrylamide gel electrophoresis, transferred to a polyvinylidene difluoride membrane (Immun-Blot; Bio-Rad Laboratories), and then immunoblotted with affinity-purified goat polyclonal anti-human dentin sialoprotein (DSP) or osteopontin antibodies or β-actin (Santa Cruz Biotechnology, Santa Cruz, CA). The β-actin was used as an internal control. Detection was performed using SuperSignal® West Pico (Thermo Scientific–Pierce Biotechnology), according to the manufacturer's instructions.
Mineralization staining
Mineralization of cultured PDPCs was determined by Alizarin Red (AR) staining. 17 After day 15, the cell layer was washed with PBS, fixed with 70% ethanol, stained with 2% AR (pH 4; Wako Pure Chemical Industries, Osaka, Japan), and then photographed. After that, the cells were destained with 100 mM cetylpyridinium chloride for 1 h. 18 The absorbance of the released stain was measured at 570 nm. The concentration of AR staining in the samples was determined by comparison with the AR standard.
Animals
Forty-eight healthy, noncarious upper first molars of 24 male Sprague Dawley rats (8 weeks old, average weight 350 g) were used in the study. The animals were obtained from the National Laboratory Animal Centre, Nakhon Pathom, Thailand. They were given access to water and diet ad libitum and maintained at 25°C ± 1°C under a 12 h light/12 h dark cycle. Throughout the experiment, the animals remained healthy and gained more weight (up to 50%).
Response of pulp tissue in rat molars
This study was approved by the Animal Ethics Committee, Faculty of Pharmaceutical Sciences, Chulalongkorn University (no. 302/2007).
The procedures were performed according to Decup et al., 19 with minor modifications. Briefly, the animals were anesthetized with an intraperitoneal injection of 80 mg/kg Zoletil® (chloral hydrate tiletamine and chloral hydrate zolazepam; Virbac Laboratories, Carros, France). Cavities were prepared on the cervical 1/3 of the mesial surface of right and left upper first molars. Under loupes at 2.5 × magnification (SurgiTel®; General Scientific Corporation, Ann Arbor, MI), half-moon class V-like cavities were drilled using a sterile round tungsten carbide bur, 0.6 mm in diameter (ISO 500104; Hager & Meisinger GmbH, Neuss, Germany). The cavities created were around 3/4 the size of the bur diameter, and the procedure was performed under copious sterile water. Then, pulp exposures were accomplished by applying gentle pressure with the tip of a sterile steel probe.
The teeth were divided equally into three groups: Group I was treated with acemannan (300 μg/tooth); Group II was treated with Ca(OH)2 (Dycal®; Dentsply Caulk, Milford, DE); Group III was a sham group. Then all cavities were filled with glass-ionomer cement (Fuji Lining LC; GC, Tokyo, Japan).
At days 7 and 28 of treatment, four animals from each group were sacrificed. The upper jaws were removed, fixed in 10% neutral formalin buffer, and demineralized with 4% nitric acid in 10% neutral formalin buffer. Dehydration of tissue was carried out by ethanol–acetone dehydration, followed by paraffin embedding. Serial sections of 7 μm thickness were cut. Pulp healing was assessed using hematoxylin and eosin and Masson's trichrome staining. Histopathological evaluation was done according to the grading criteria of Tarim et al. 20 and Kitasako et al., 21 which, with minor modifications, consist of inflammatory cell response, dentin bridge formation, and soft tissue organization (Table 1).
Statistical analysis
Statistical analysis was performed using the SPSS program for Windows, version 15.0 (SPSS, Chicago, IL). For the in vitro study, the results of three or four independent experiments were expressed as mean ± standard error and analyzed by one-way analysis of variance and Scheffé multiple comparisons. For the in vivo study, the scores of histopathological features were analyzed by Kruskal–Wallis and Bonferroni multiple comparisons. Values of p < 0.05 were considered to be statistically significant.
Results
Acemannan induced PDPC proliferation
After 24 h of incubation, acemannan at concentrations of 1, 2, and 4 mg/mL significantly stimulated new DNA synthesis in PDPCs by about 2.0, 2.1, and 1.7 times, respectively, compared with the untreated group (Fig. 1). A dose-dependent effect of acemannan was observed. Acemannan at a concentration of 2 mg/mL exhibited the maximum effect on de novo DNA synthesis.
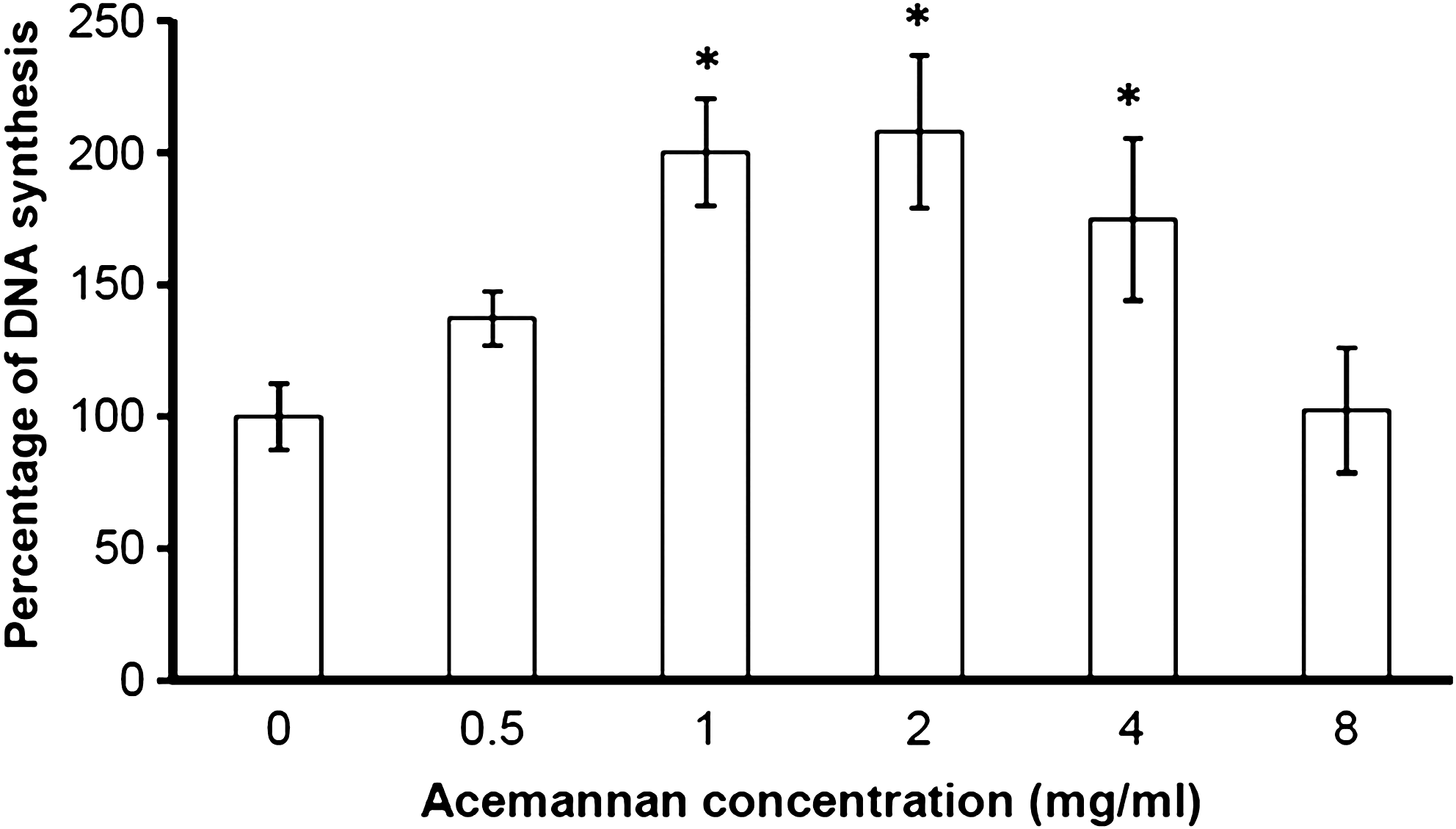
Acemannan induced PDPC proliferation. PDPC proliferation was examined by incorporation of [3H]-thymidine after 24 h of incubation with various concentrations of acemannan. Acemannan significantly stimulated PDPC proliferation at concentrations of 1, 2, and 4 mg/mL. *Compared with the untreated group; p < 0.05, n = 3. PDPC, primary human dental pulp cell.
Acemannan increased DSP and BMP-2 expressions
At day 3 of incubation, acemannan at concentrations of 1, 2, 4, and 8 mg/mL significantly upregulated DSP expression by about 1.1, 1.6, 1.9, and 1.7 times, respectively, compared with the untreated group (Fig. 2). Acemannan induced DSP expression in a dose-dependent manner. Treatment with acemannan also resulted in a slight increase of osteopontin expression (Fig. 2).
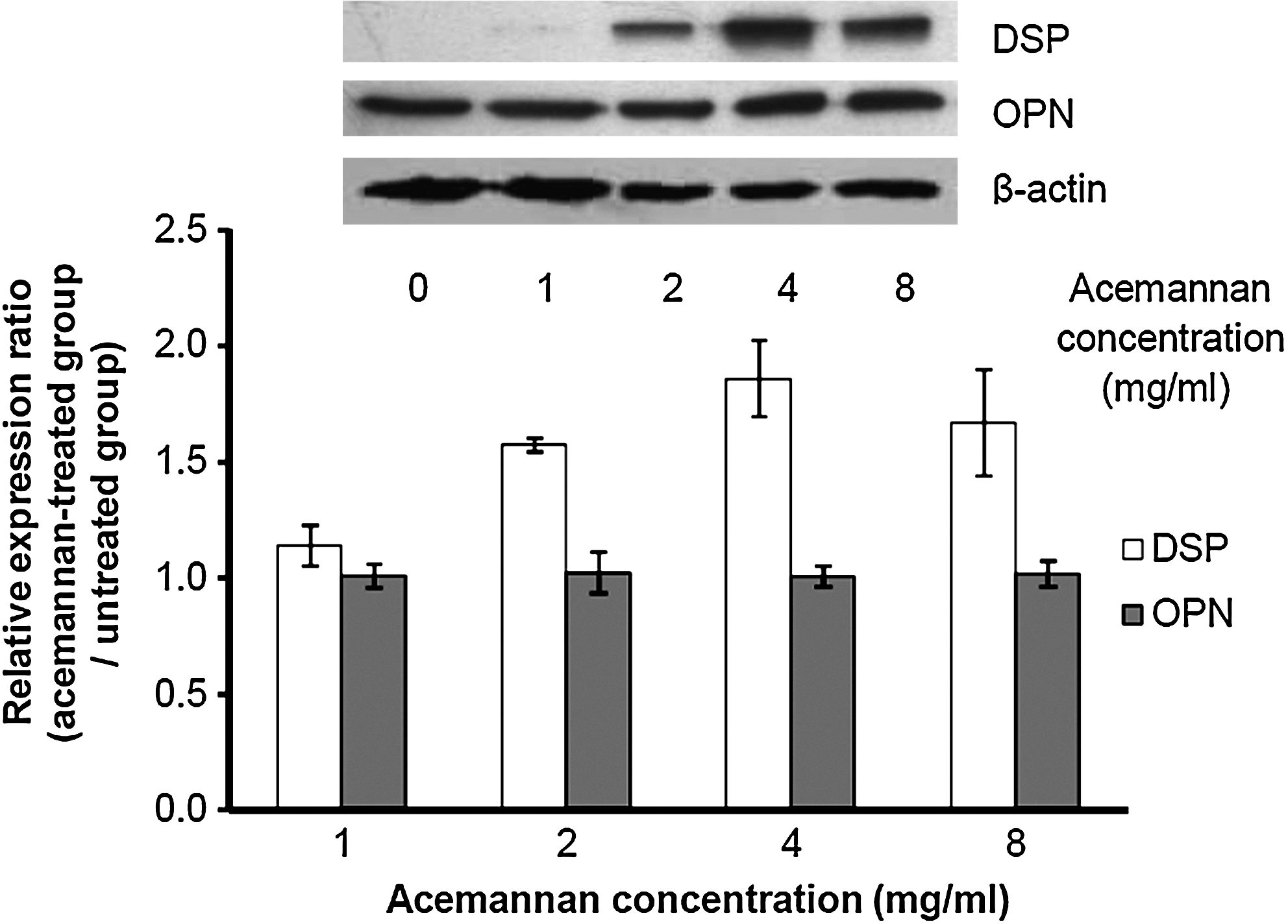
Western blot analysis of DSP and OPN expression. The whole cell lysate from acemannan-treated cells was separated by 10% sodium dodecyl sulphate–polyacrylamide gel electrophoresis, transferred to a polyvinylidene difluoride membrane, and immunoblotted with goat polyclonal anti-human DSP or OPN antibody. β-Actin was used as an internal control for each group (n = 3). DSP, dentin sialoprotein; OPN, osteopontin.
At day 3 of incubation, only acemannan at a concentration of 4 mg/mL significantly enhanced BMP-2 expression by around 1.2 times when compared with the untreated group. At days 9 and 12 of incubation, acemannan at concentrations of 2, 4, and 8 mg/mL significantly stimulated BMP-2 expression by 30–50% when compared with the untreated group. Only acemannan at a concentration of 4 mg/mL significantly upregulated BMP-2 expression in PDPCs from day 3 to 12 of the study (Fig. 3).
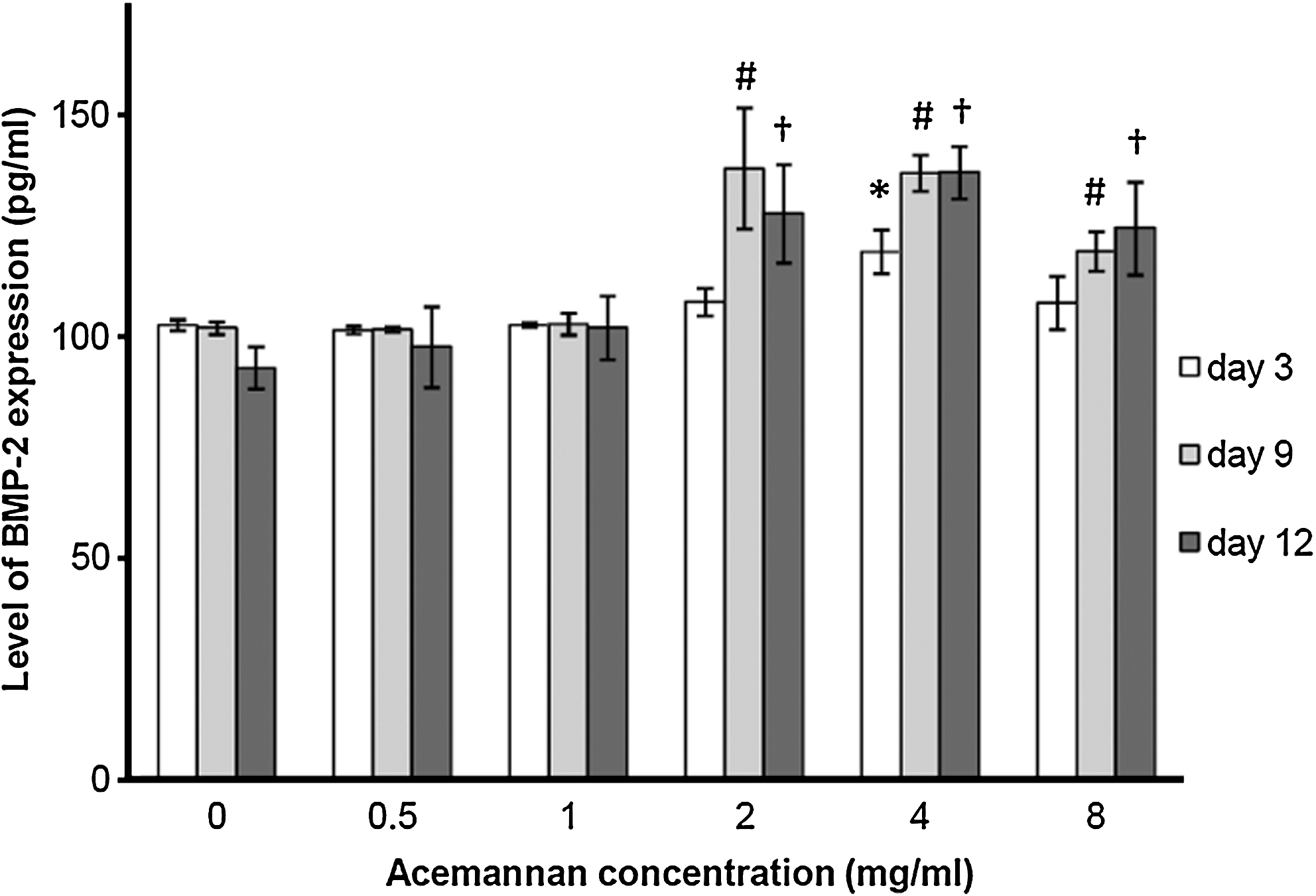
Acemannan induced BMP-2 expression in PDPCs. BMP-2 production was determined using the conditioned media at days 3, 9, and 12 of treatment by enzyme-linked immunosorbent assay method. Acemannan significantly enhanced BMP-2 expression at day 3 (*), day 9 (#), and day 12 (†); p < 0.05, n = 4. BMP-2, bone morphogenetic protein-2.
Acemannan stimulated ALPase activity and mineral deposition
When compared with the untreated group, acemannan at a concentration of 4 mg/mL significantly enhanced the ALPase activity of PDPCs by approximately 1.4 and 1.8 times at days 3 and 9 of the study, respectively. Acemannan at a concentration of 8 mg/mL also significantly augmented ALPase activity by approximately 1.4 times at day 9 of the study (Fig. 4).
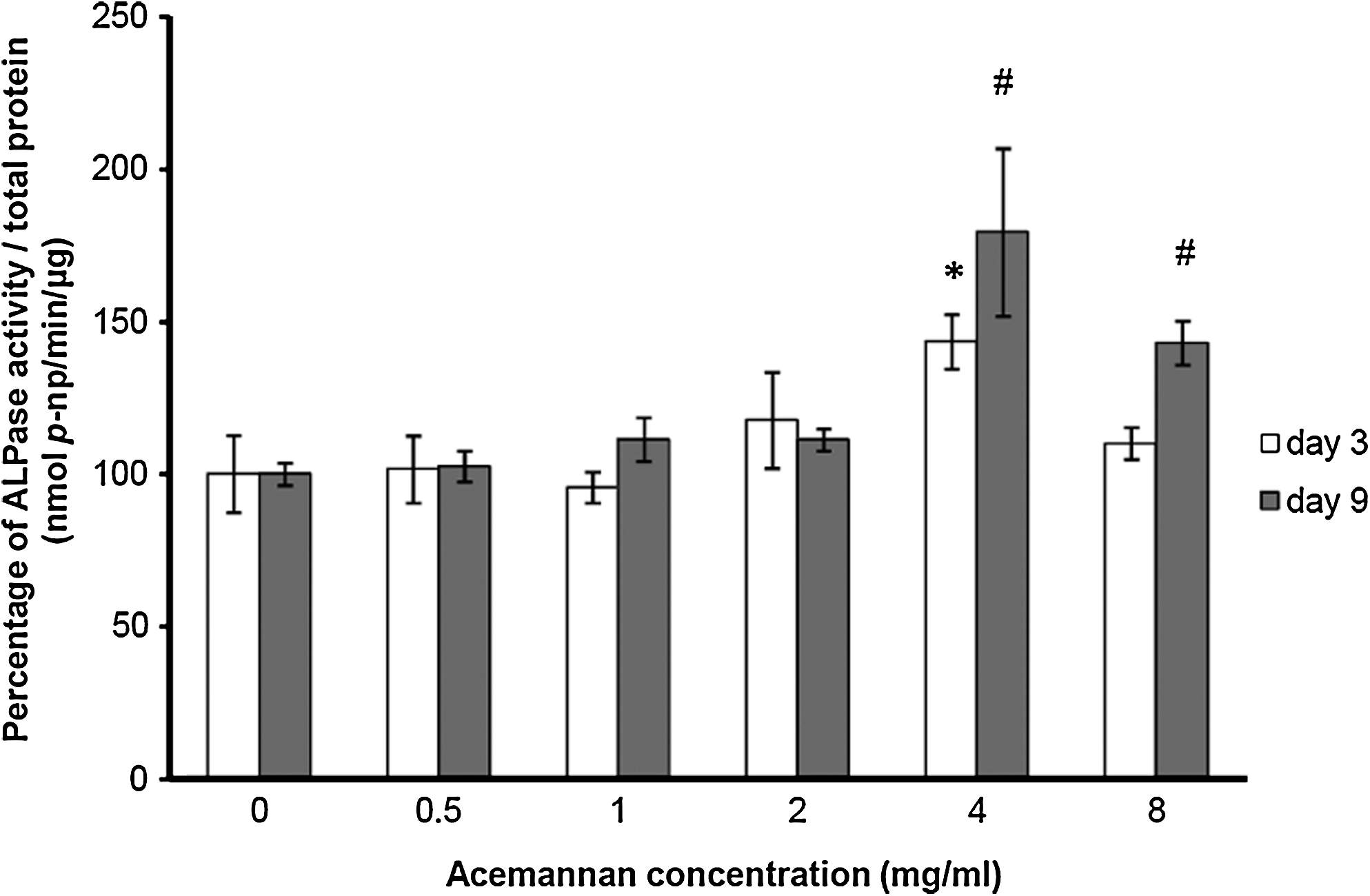
Acemannan stimulated ALPase activity of PDPCs. After 3 and 9 days of incubation with various concentrations of acemannan, the ALPase activity with total protein correction was examined by biochemical assay. Acemannan significantly enhanced ALPase activity of PDPCs at day 3 (4 mg/mL) and day 9 (4 and 8 mg/mL). The control ALPase activity was 0.05 ± 0.006 and 0.04 ± 0.010 nmol p-np/min/μg protein at days 3 and 9, respectively. *Compared with the untreated group at day 3; #compared with the untreated group at day 9; p < 0.05, n = 4. ALPase, alkaline phosphatase; p-np, p-nitrophenol.
After 15 days of incubation, acemannan effectively induced mineral deposition by PDPCs. Large and intense areas of AR staining were detected in the cells treated with acemannan (Fig. 5a–h). The amount of mineralization significantly increased in all acemannan-treated groups, by about five- to sixfold when compared with the untreated group (Fig. 5i).
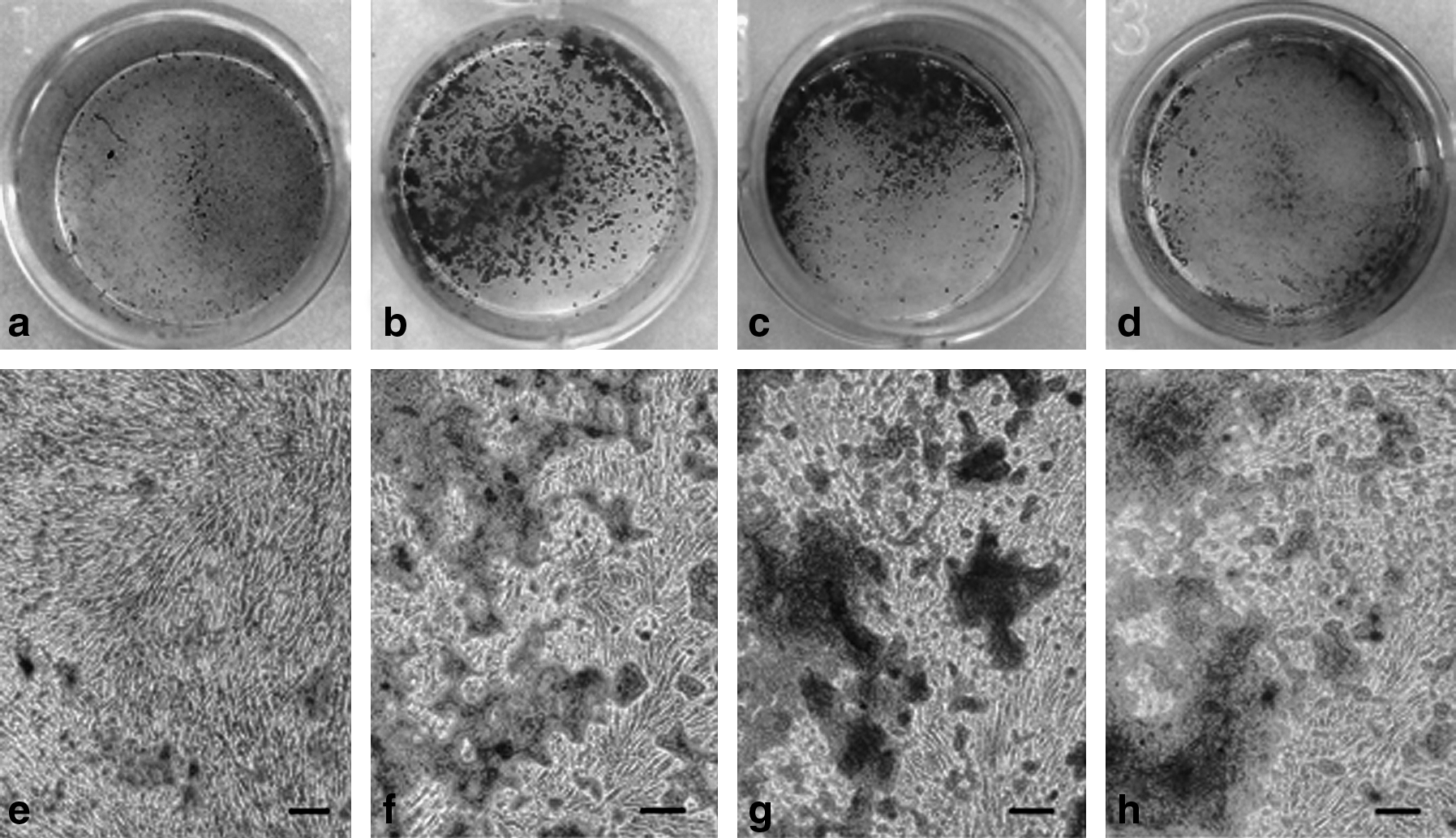
Acemannan induced PDPC mineralization. After 15 days of incubation with various concentrations of acemannan, mineral deposition was stained with AR. PDPCs treated with acemannan at concentrations of 2, 4, and 8 mg/mL (
Acemannan stimulated dental pulp healing by inducing tissue organization and dentin bridge formation and by reducing inflammation
During the experiment, at day 7 of treatment, a rat in the sham group died because of anesthetic overdose. Also one tooth sample in the acemannan-treated group at day 7 of treatment and one in the sham group at day 28 of treatment were found to have been damaged during the histologic sectioning process. Therefore, on day 7, the numbers of tooth samples in the acemannan-treated, Ca(OH)2-treated, and sham groups were seven, eight, and six, respectively, and on day 28, sample numbers were eight, eight, and seven, respectively.
The dental pulp response scores of all groups are shown in Table 2. At day 7, the scores for inflammatory response, dentin bridge formation, and soft tissue organization were not statistically different among the three groups. The acemannan-treated group revealed the presence of dense collagen fibers around the exposed sites (see Fig. 8a). Remarkably, dental pulp cells of the acemannan-treated group displayed more hyperplasia and hypertrophy than those of the other groups (Fig. 6).
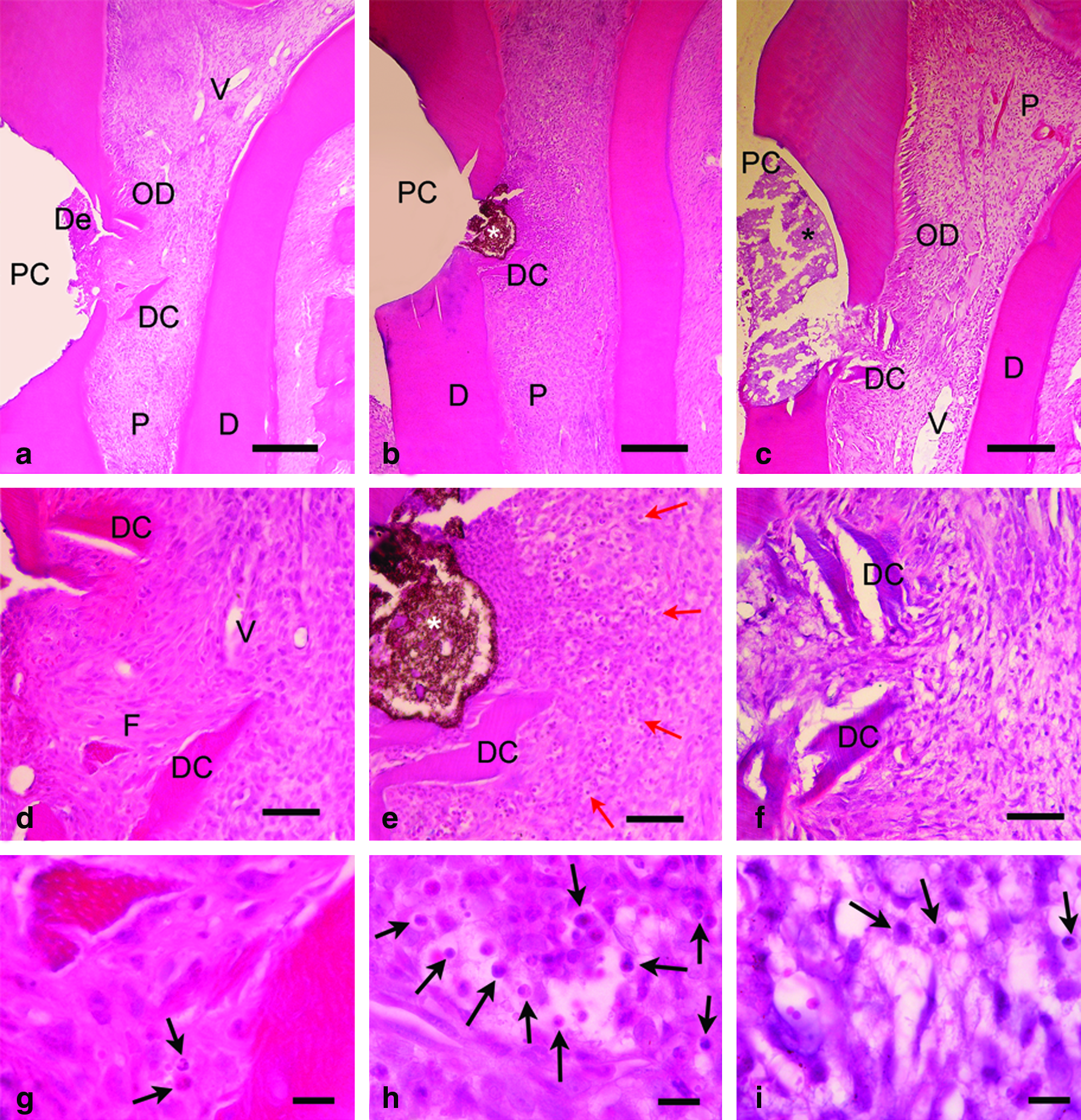
Histopathology of dental pulp at day 7 of the acemannan-treated group (
At day 28, the acemannan-treated group revealed significant inflammatory reduction, complete dentin bridge formation, and good tissue organization when compared with both Ca(OH)2-treated and sham groups (Figs. 7 and 8d–f). Pulps treated with acemannan had dense, regularly arranged collagen fibers within the dentin bridges, whereas the pulps of the Ca(OH)2-treated and sham groups revealed less-dense collagen fibers that contained some defects (Fig. 8d–f). All of the teeth in the acemannan-treated group had little or no inflammatory cell infiltrate (Fig. 7a, d), most of which successfully synthesized a homogeneous hard dentin bridge that completely covered exposure sites (seven of eight teeth or 87.5%; Fig. 8d). The pulp tissue was well organized, with new odontoblast-like cells (seven of eight teeth; Fig. 7a, g). In the Ca(OH)2-treated group, moderate inflammation (five of eight teeth) and poorly organized soft pulp tissue (seven of eight teeth) were observed (Fig. 7b, e). A dentin bridge appeared, but in incomplete heterogeneous patterns covering the exposure site (partial: four of eight teeth; almost complete: three of eight teeth; Fig. 8e).
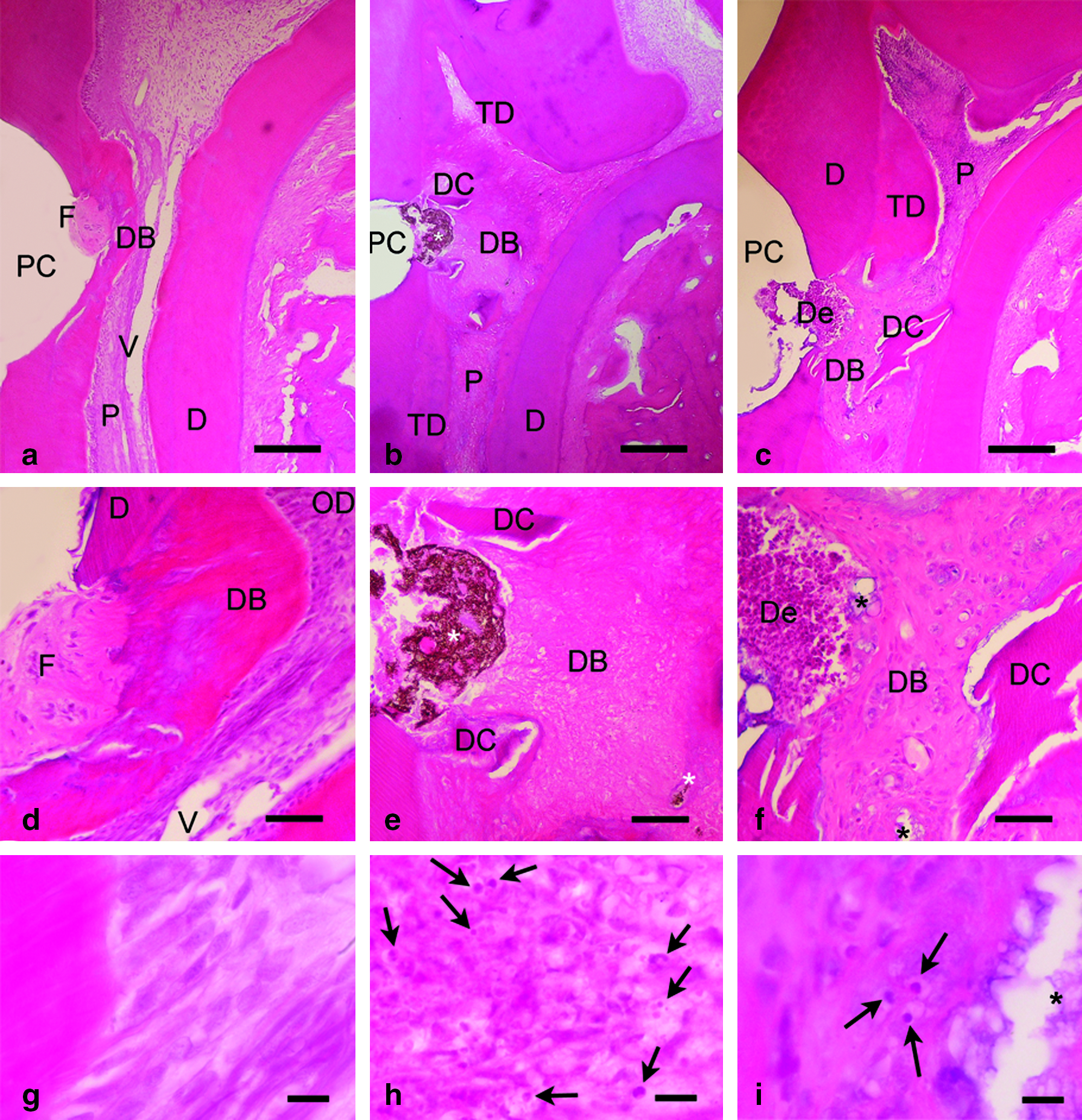
Histopathology of dental pulp at day 28 of the acemannan-treated group (
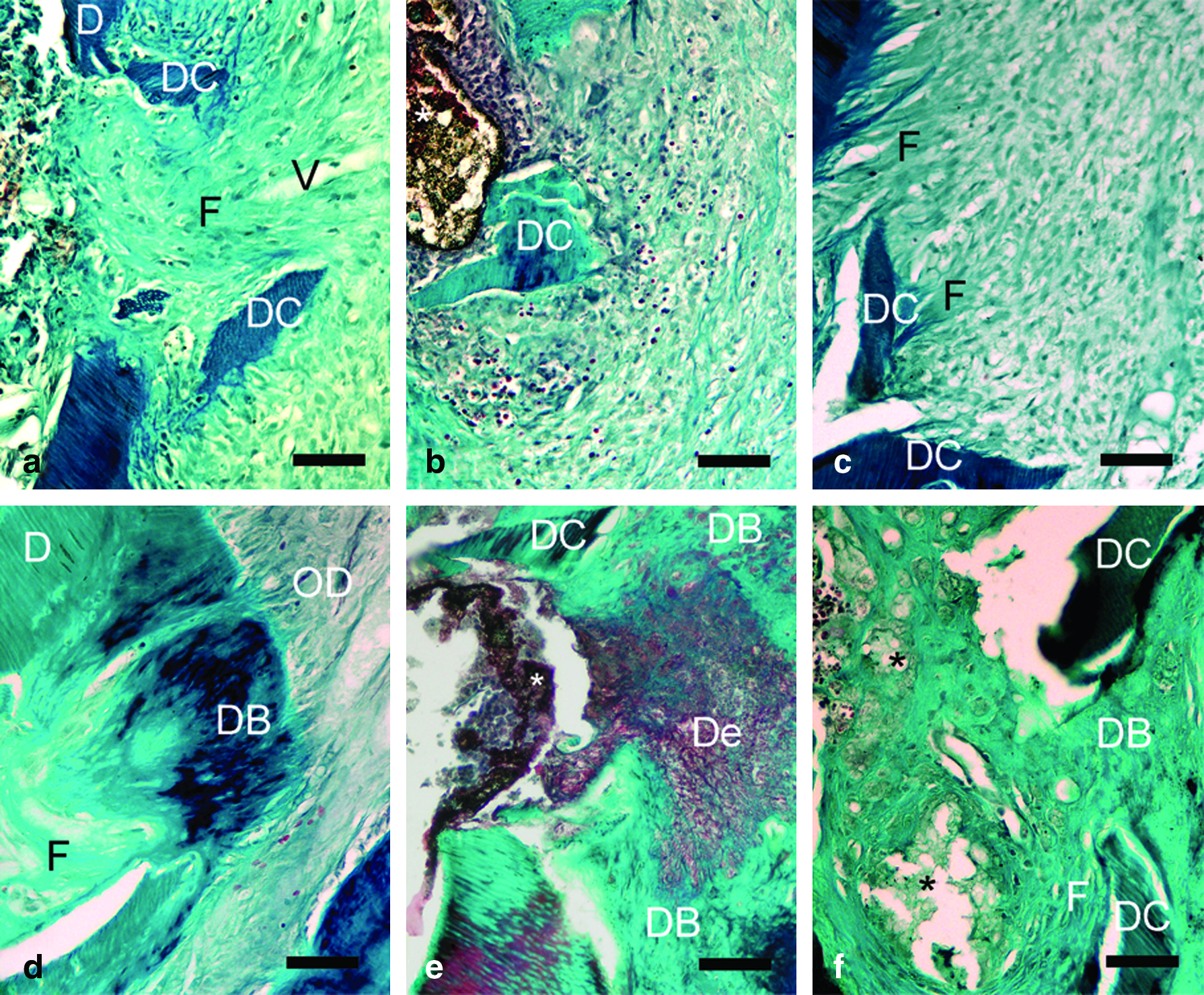
Histopathology of dental pulp at day 7 (
Discussion
Acemannan, β-(1,4)-acetylated polymannose, is the major polysaccharide component of A. vera gel. 22 The safety and healing efficacy of acemannan has been studied in humans and animals.23–26 Recent reports state that acemannan stimulates wound healing in the mouse footpad 27 and rat hard palate. 11 Dental pulp cells, or mesenchymal fibroblast-like cells, are considered to be odontoblast progenitor cells. Dental pulp cells proliferate, differentiate into odontoblast-like cells, and produce new dentin. 1 Therefore, in this study, PDPCs were selected as odontoblast progenitor cells to investigate the potential effect of acemannan on dentin formation, including proliferation, differentiation, and mineral deposition.
[3H]-thymidine incorporation assay was used to determine cell proliferation. This assay is an accurate and simplified method of measuring the incorporation of radioactive precursors of DNA. 28 Our data show that acemannan significantly stimulated PDPC proliferation. This finding is consistent with our recent report that acemannan induced gingival fibroblast proliferation. 11
In this study, acemannan significantly induced ALPase activity, production of BMP-2 and DSP, and mineral deposition. This finding is consistent with our previous report that acemannan upregulated BMP-2 expression in mRNA levels in both pulpal fibroblasts and periodontal fibroblasts after 24 and 48 h of incubation. 13
ALPase activity and DSP are considered to be odontoblast differentiation markers. 29 ALPase activity is present during the early differentiation phase and plays an important role in mineral deposition.30,31 An immunohistochemical study found that odontoblasts and dental pulp cells express ALPase activity during dentin formation in developing teeth. 32 DSP is a dentin extracellular matrix protein that functions as an initiator of mineralization. 33 DSP production is widely recognized as one of the most specific markers of the odontoblast phenotype.29,34,35 DSP is also expressed in bone, but at a level around 1/400th of that in dentin. 36
BMP-2 plays an important role in tooth and bone formation. This protein has been successful in inducing bone and dentin formation in both in vitro and in vivo studies.6,37 BMP-2 promotes odontoblast differentiation and mineral deposition.38,39 Therefore, secreted BMP-2 may have an autocrine effect that induces PDPC differentiation and mineral deposition. Mineral deposition has been considered to be a late stage of odontoblast differentiation. 29
Taken together, our laboratory data suggest that acemannan not only induces dental pulp cell proliferation but also stimulates dental pulp cell differentiation into mature odontoblast-like cells, which synthesize dentin. To confirm the potential activity of acemannan on dentin formation, an in vivo test was done in a rat model. This animal model has been accepted for investigating the biocompatibility of dental materials and the dentin formation of bioactive molecules.20,40,41 Glass-ionomer cement was used to seal the cavities against bacterial invasion. 19 To compare the effect of acemannan on dentin formation, Ca(OH)2 was used in the control group. To date, Ca(OH)2 is regarded as the gold standard treatment for direct pulp capping.
After 28 days of treatment, the acemannan-treated group demonstrated a success rate for homogeneous complete mineralized dentin bridge formation of around 87.5%. There was no complete dentin bridge formation in either the Ca(OH)2-treated group or sham group. This is consistent with our in vitro data indicating that acemannan could stimulate cell proliferation, differentiation, and mineral deposition. None of the acemannan-treated samples showed any inflammation around the exposure site or under the dentin bridge. Remnants of acemannan sponge were not detected in any of the treated samples. These findings suggest that acemannan has good biocompatibility with pulp tissue, which corresponds with a previous study in which acemannan was placed into extraction sockets of patients. 26
In this study, the results obtained from the Ca(OH)2-treated group were poor. All samples revealed incomplete hard dentin bridge formation, with moderate to severe inflammation and dystrophic calcification of the pulp tissue. This is consistent with the study of Asgary et al., 42 in which inflammation and incomplete dentin bridge formation were found in all cases of dogs with Ca(OH)2 pulp capping.
At day 7 of treatment, there was no statistical difference in pulp responses among the three groups. A possible explanation is that the first week of reparative dentin formation is the inflammatory phase. 19 Like the inflammatory phase of wound healing, only macrophages and white blood cells play major roles in the clearing of necrotic tissue and cell debris. 43 The formative phase of dentin formation in rats is thought to begin late in the first week, lasting until the fourth week. 19 This corresponds with our in vitro and in vivo data showing that acemannan-treated dental pulp cells have higher cellular activity and larger cell volume than other groups.
It should be noted that this animal study was performed in healthy pulp tissue conditions with iatrogenic exposure. In clinical practice, pulp exposure usually occurs as a result of caries and is accompanied by a severe inflammation. In the case of direct pulp capping, it is generally accepted that iatrogenic pulp exposures through intact dentin have better prognosis than infected carious pulp exposures. Finally, to verify its reproducibility, further studies of acemannan on dentin formation in infected carious teeth should be performed.
In conclusion, acemannan accelerated new dentin formation via pulp cell proliferation, differentiation into odontoblast-like cells, upregulation of BMP-2 and DSP expression, and mineral deposition.
Footnotes
Acknowledgments
The authors thank Professor Dr. Visaka Limwongse and Associate Professor Dr. Dolly Methathrathip for their valuable suggestions, and also Dr. Ngampis Six and Dr. Suwimon Jettanacheawchankit for their outstanding advice. This work was supported by the Thai Government Research Fund 2008–2009 and by the 90th Anniversary of Chulalongkorn University Fund (Ratchadaphiseksomphot Endowment Fund) 2007.
Disclosure Statement
No competing financial interests exist.