
Abstract
The aim of this study was to investigate the roles of Smad2/3 and Smad1/5/8 phosphorylation in transforming growth factor-beta-induced chondrogenic differentiation of bone-marrow-derived mesenchymal stem cells (BMSCs) to assess whether specific targeting of different Smad signaling pathways offers possibilities to prevent terminal differentiation and mineralization of chondrogenically differentiated BMSCs. Terminally differentiated chondrocytes produced in vitro by chondrogenic differentiation of BMSCs or studied ex vivo during murine embryonic limb formation stained positive for both Smad2/3P and Smad1/5/8P. Hyaline-like cartilage produced in vitro by articular chondrocytes or studied in ex vivo articular cartilage samples that lacked expression for matrix metalloproteinase 13 and collagen X only expressed Smad2/3P. When either Smad2/3 or Smad1/5/8 phosphorylation was blocked in BMSC culture by addition of SB-505124 or dorsomorphin throughout culture, no collagen II expression was observed, indicating that both pathways are involved in early chondrogenesis. Distinct functions for these pathways were demonstrated when Smad signaling was blocked after the onset of chondrogenesis. Blocking Smad2/3P after the onset of chondrogenesis resulted in a halt in collagen II production. On the other hand, blocking Smad1/5/8P during this time period resulted in decreased expression of matrix metalloproteinase 13, collagen X, and alkaline phosphatase while allowing collagen II production. Moreover, blocking Smad1/5/8P prevented mineralization. This indicates that while Smad2/3P is important for continuation of collagen II deposition, Smad1/5/8 phosphorylation is associated with terminal differentiation and mineralization.
Introduction
After traumatic injury or in diseases such as osteoarthritis or rheumatoid arthritis, cartilage can be damaged and the limited reparative capacity of cartilage is a major problem. Therefore, bone-marrow-derived mesenchymal stem cells (BMSCs) are widely studied for their potential use in cartilage tissue engineering to repair cartilage defects. Chondrogenic differentiation of BMSCs follows a process similar to embryonic limb development. The cells condensate, 3 start expressing SOX9 and collagen type II, but eventually the cells become terminally differentiated expressing matrix metalloproteinase 13 (MMP13), collagen X, and alkaline phosphatase (ALPL).4–6 This terminal differentiation of BMSCs may be advantageous for tissue engineering of bone through the endochondral route. 5 However, for cartilage reconstructions it is highly unwanted as terminally differentiated cartilage produced by BMSC mineralizes when implanted in vivo. 4 So far, inhibition of terminal differentiation of BMSCs was only demonstrated through the addition of PTHrP.7,8 Elucidation of the mechanisms regulating chondrocyte differentiation and especially terminal differentiation are crucial to have a better control over the chondrogenic differentiation of BMSCs.
For in vitro chondrogenic differentiation of BMSCs, transforming growth factor-beta (TGF-β) is commonly used.9,10 TGF-β signaling requires binding to complexes of type II and type I serine/threonine kinase receptors, followed by receptor-Smad phosphorylation at their C-terminus. The canonical TGF-β pathway is the Smad2/3 pathway. However, by use of an alternate type I receptor (ALK1 instead of ALK5) Smad1/5/8 is phosphorylated instead of Smad2/3.11–13 The Smad1/5/8 route is commonly known as the route activated by bone morphogenetic proteins (BMPs), also a member of the TGF-β superfamily and a very potent inducer of bone formation. 14
The Smad2/3 route has been found to block terminal differentiation,15,16 whereas the Smad1/5/8 route is known to stimulate terminal differentiation in murine embryonic endochondral ossification. 17 We have recently found that the Smad2/3 route is protective for postnatal articular cartilage and that loss thereof is potentially a hallmark of cartilage degradation.18,19 In contrast, in osteoarthritic (OA) chondrocytes ALK1 expression was related to MMP13 expression, a marker of terminally differentiated chondrocytes. 19
We postulate that Smad2/3 signaling is important during all stages of chondrogenic differentiation and that Smad1/5/8 signaling is a crucial factor for terminal differentiation. We suggest that blocking Smad1/5/8 signaling is a potential tool to prevent terminal differentiation of BMSCs, thereby generating tissue-engineered cartilage that does not mineralize and more closely resembles the desired stable hyaline cartilage.
Therefore, we set out to investigate the activation of both the Smad2/3 and the Smad1/5/8 pathways in in vitro chondrogenically differentiated BMSCs, which are known to generate terminally differentiated cartilage. We investigated these pathways in in vitro cultured articular chondrocytes, which are known to produce stable hyaline-like cartilage 4 and compared Smad signaling in in vitro cultured cartilage with in vivo terminally differentiated cartilage in murine embryonic limbs, and in vivo human healthy articular cartilage. Then, we assessed the outcome of blocking either Smad2/3 or Smad1/5/8 phosphorylation during different stages of in vitro chondrogenic differentiation of BMSCs. Finally, we tested whether blocking Smad1/5/8 phosphorylation not only prevented hypertrophic differentiation but also resulted in a more stable cartilage construct that does not mineralize.
Materials and Methods
Harvesting cartilage samples
To study Smad signaling in healthy hyaline and terminally differentiated cartilage in vivo, cartilage was processed directly for immunohistochemistry. Human healthy articular cartilage was harvested from left-over material of an amputated knee of a 19-year-old patient. Forelimbs of E16 murine embryo's were used to study terminal differentiated cartilage in the central region of the metacarpal bone. C57/BL6 female mice were intercrossed with CBA males (Charles River Laboratories). Gestational age was estimated by the vaginal plug method, with the day of plug occurrence designated as day 1 (E1). After gravid female mice had been killed by isoflurane inhalation, embryos were excised after cervical dilatation on E16 and immediately placed into Dulbecco's phosphate-buffered saline (PBS) (Gibco).
Cell isolation and expansion
BMSCs were isolated from a bone marrow biopsy from the femoral shaft of five patients (age 20–44) undergoing total hip replacement, and articular cartilage was obtained from two donors (age 3–8) undergoing triple arthrodesis to treat clubfoot deformity, after informed consent had been obtained in accordance with the local ethics committee of Rotterdam (MEC-2007-032 and MEC 2004-142).
For isolation of BMSCs, the heparinized aspirate from the femoral shaft biopsy was seeded at a density of 2–5 × 105 cells/cm2 in Dulbecco's modified Eagle's medium (DMEM)-low glucose (LG) (Gibco), supplemented with 10% fetal calf serum (FCS; Gibco), 1 ng/mL fibroblast growth factor 2 (AbD Serotec), 25 μg/mL ascorbic acid-2-phosphate (Becton Dickinson), 1.5 μg/mL fungizone (Gibco), and 50 μg/mL gentamicin (Gibco). After 24 h, nonadherent cells were washed off and adherent cells were further expanded. At subconfluency, BMSCs were trypsinized, seeded at a density of 2300 cells/cm2, and further expanded. BMSCs from passages 2–4 were used.
For isolation of chondrocytes, cartilage slices were rinsed with saline and subsequently digested through incubation for 2 h at 37°C with 2 mg/mL Pronase E (Sigma-Aldrich), followed by overnight incubation with 1.5 mg/mL Collagenase B (Boehringer) in DMEM containing Glutamax (Gibco), and 10% FCS (Gibco). Next, cell suspensions were filtered through a 100 μm filter, centrifuged, and washed with saline. For expansion, chondrocytes were seeded at a density of 7500 cells/cm2 in DMEM containing Glutamax, with 10% FCS and 1.5 μg/mL fungizone (Gibco) and 50 μg/mL gentamicin (Gibco). When chondrocytes reached subconfluency they were trypsinized and further expanded. Chondrocytes from passage 4 were used for the experiments.
In vitro chondrogenic differentiation
Cells were cultured in pellets, formed by centrifuging aliquots of 2 × 105 cells in 0.5 mL medium at 200 g for 8 min in a polypropylene tube. Chondrogenic differentiation was induced by DMEM containing Glutamax, ITS+1 (B&D Bioscience), 40 μg/mL L-proline (Sigma-Aldrich), 1 mM sodium-pyruvate (Gibco), 5 μg/mL fungizone, and 50 μg/mL gentamicin, 25 μg/mL ascorbic acid-2-phosphate, 10 ng/mL TGF-β2 (R&D Systems), and 10−7 M dexamethasone (Sigma-Aldrich). The medium was changed twice a week. Pellets were cultured for 35 days in total and harvested in triplicates for immunohistochemistry and gene expression.
To study differentiation and Smad expression during the course of chondrogenic differentiation of BMSC, pellets of donor 2 and 5 were harvested days 7, 14, 21, 28, and 35. In addition, pellets of BMSC donors 3 and 4 were harvested on days 14, 21, and 35 and articular chondrocytes were harvested on days 7, 14, 21, 28, and 35.
Modulation of Smad-signaling
To study the effect of inhibition of Smad2/3 phosphorylation on chondrogenic differentiation of BMSCs, 10 μM SB-505124 (Sigma-Aldrich) was added to the medium during chondrogenic differentiation of BMSC from three different donors (donors 1–3). SB-505124 blocks the kinase domains of ALK4, -5, and -7, thereby preventing Smad2/3 phosphorylation. To study the effect of inhibition of Smad1/5/8 phosphorylation, 10 μM dorsomorphin (Biomol International) was added to the medium of BMSC from three donors (donors 2–4). Dorsomorphin blocks kinase domains of ALK1, -2, -3, and -6, thereby preventing Smad1/5/8 phosphorylation. Before using these compounds in the chondrogenic differentiation of BMSC, their specific inhibitory effect on phosphorylation of either Smad2/3 or Smad1,5,8 was confirmed by Western Blotting cell cultures in monolayer (data not shown).
As we have previously demonstrated the importance to modulate chondrogenesis of BMSC stage specifically, 20 these compounds were added to the medium not only throughout the culture period (days 0–35) but also from days 14 to 35 and days 21 to 35. After 2 or 3 weeks of chondrogenic differentiation of BMSC, some collagen II production is usually seen, but this is donor dependent. Therefore, we used two different time points for the addition of dorsomorphin or SB505124 to study their effect on cartilage production and terminal differentiation without influencing the initial onset of chondrogenic differentiation. The effect of blocking Smad signaling during onset of differentiation only was not investigated as the aim of our study was to investigate the involvement of different Smad pathways in terminal differentiation.
In the donor with the best chondrogenic capacity, blocking of Smad1/5/8 phosphorylation on days 14–35 resulted in inhibition of markers of terminal differentiation (donor 4). To study whether stage-specific blocking of Smad1/5/8 after the onset of chondrogenic differentiation results in more stable cartilage construct that does not mineralize, 10 μM dorsomorphin was added to the culture medium from day 14 on as in the previous experiment using cells from the donor with the best chondrogenic capacity (donor 4). In this experiment culture continued after 35 days with the addition of β-glycerophosphate to the medium (containing TGF-β ± dorsomorphin) to allow for mineralization. After a total culture period of 49 days pellets were harvested for histology.
(Immuno)histochemistry
Samples were fixed in 4% formalin in PBS and embedded in paraffin. Pellets were first set in 2% agar before embedding in paraffin. To enable reliable direct comparison of intensity of the staining sections of treated pellets and controls were put on the same slide.
Antigen retrieval for collagen II was performed through incubation with 0.1% pronase (Sigma-Aldrich) in PBS for 30 min, whereas antigen retrieval for collagen X required 0.1% pepsin (Sigma-Aldrich) in 0.5 M acetic acid (pH 2.0) for 2 h. Both stainings continued with incubation with 1% hyaluronidase (Sigma-Aldrich) in PBS. Sections were incubated overnight with primary antibodies for collagen II (II/II6B3, Developmental Studies Hybridoma Bank) or collagen X (2031501005; Quartett) after blocking of nonspecific binding sites with 10% goat serum in PBS. An ALPL-conjugated secondary antibody was used for these immunohistochemical stainings, followed by incubation with Neu Fuchsin substrate (Chroma) to demonstrate ALPL activity with a red stain. To allow the use of these monoclonal mouse antibodies on murine specimens, an earlier described method 21 was used to link the antibody with antimouse antiserum before immunohistochemistry.
MMP13 staining was preceded by incubation of the sections in chondroitinase ABC (Sigma-Aldrich) 250 units/mL for 1 h at 37°C. For Smad2P, Smad1/5/8P, and MMP13, endogenous peroxide was blocked with 1% hydrogen peroxide in methanol for 30 min, followed by antigen retrieval through incubation in citrate buffer (0.1 M sodium citrate +0.1 M citric acid) for 2 h at room temperature. Sections were incubated overnight with primary antibodies against Smad2P, Smad1/5/8P (both Cell Signaling Technology), or MMP13 (Santa Cruz Biotechnology) after blocking of nonspecific binding sites with 5% goat serum. A biotin-labeled secondary antibody was used (DAKO) for these immunohistochemical stainings, followed by a biotin–streptavidin detection system according to manufacturer's protocol (Vector Laboratories). DAB reagent was used to demonstrate bound complexes with a brown stain.
Sections were critically studied for presence or absence of a positive staining. In case of collagen II and collagen X immunohistochemistry, a red staining was considered positive, whereas in the case of Smad2P, Smad1/5/8P, and MMP13 a brown staining was positive. As the antibody for Smad2P detects Smad3 phosphorylated at equivalent sites according to the manufacturer, we considered positivity for this antibody as positivity for Smad2/3P.
To evaluate mineralization a von Kossa staining and a Alizarin Red staining were performed. For the von Kossa staining, slides were immersed in 5% silver nitrate solution (Sigma-Aldrich) for 10 min, rinsed in ultra pure water, and exposed to light for 10 min. Excess silver nitrate was removed with 5% sodium-thiosulfate (Sigma-Aldrich) and cells were rinsed in distilled water. For the Alizarin Red staining, slides were immersed in a saturated Alizarin Red S solution (Sigma-Aldrich) (5 g/L) for 1 min, and rinsed in 70% ethanol.
All (immuno)histochemical stainings were quantified using Bioquant Osteo 7.2 (Bioquant Image Analysis Corporation). Quantification was performed on a section through the middle of the pellet. First color-definitions were set per staining to define a positive staining. In the case of staining for collagen II, collagen X, MMP13, von Kossa, and Alizarin Red, the positive staining was then automatically selected and the size of the positive staining area was defined. Next, the total area of the pellet was manually selected and the size defined. The positive staining fraction over the total pellet area was calculated and used for further analysis.
Positive staining for Smad2/3P or Smad1/5/8P was observed in the cell, indicating active signaling via this pathway. Quantification using Bioquant Osteo was performed on a 200 × magnified image in a collagen II rich area of the pellet. Positive cells were automatically selected by the software and the percentage of positive cells was calculated. For the aim of this article we focused on whether active signaling was present or not, and the intensity of the staining was not further quantified.
We have attempted to perform Western blots for Smad2/3P and Smad1/5/8P on pellets, but unfortunately these were not successful due to lengthy protein extraction procedure and because the extraction contained large amounts of matrix constituent. Further, baseline Smad levels were not studied, as the aim of this study was to investigate the role of altered active Smad signaling, as indicated by phosphorylated Smads. We did not study whether altered Smad phosphorylation was caused by alteration in basal Smad levels since, although interesting, this will not alter the conclusions of this study.
Gene expression analysis
For total RNA isolation, pellets were manually homogenized in RNA-Bee™ (TEL-TEST). RNA was extracted with chloroform and purified from the supernatant using the RNAeasy Micro Kit (Qiagen), according to the manufacturer's guidelines with on-column DNA-digestion.
RNA concentration and purity were assessed on a spectrophotometer (NanoDrop® ND-1000 UV-Vis Spectrophotometer; Isogen Life Science B.V.). cDNA was made using RevertAid™ First Strand cDNA Synthesis Kit (MBI Fermentas) and polymerase chain reactions were performed using TaqMan® Universal PCR MasterMix (Applied Biosystems), as described earlier, 22 but on a ABI PRISM® 7000 (Applied Biosystems). Primers used for real-time reverse transcriptase (RT)-polymerase chain reactions for SOX9, CBFA1, collagen II, collagen X, and ALPL were reported previously by Farrell et al., 5 and for MMP13 by de Mos et al. 23
Amplification efficiencies of all assays were between 90% and 98% (data not shown). After testing GAPDH, 18S, HPRT, ACTB, UBC, and B2M as housekeeping genes on selected samples, GAPDH was chosen to normalize the data. Relative expression was calculated according to the 2-ΔCT formula. 24
Since in previous experiments we have seen that undifferentiated BMSCs express the genes of interest at a very low or undetectable level (unpublished data) these conditions were not included.
Statistical analysis
Statistical analysis was conducted using SPSS 15.0 and the GraphPad Prism 5 software for Windows. Values are represented as mean ± standard error of the mean. Mixed model analysis and independent sample t-test were performed and the level of significance was set at a p-value of <0.05.
Results
Both Smad2/3 and Smad1/5/8 signaling are active in terminally differentiated cartilage, whereas only Smad2/3 is active in hyaline cartilage
Pellets of BMSCs of all five donors contained abundant collagen II and collagen X after 35 days of culture (Fig. 1A). Sox-9, CBFA1, collagen II, collagen X, and ALPL mRNA increased in time with limited variation between donors (Fig. 2) and were highly expressed in all pellets at day 35 confirming the chondrogenic and subsequent terminal differentiation of these in vitro differentiated BMSCs. Moreover, MMP13 was highly expressed on both protein and mRNA level (Figs. 1B and 2). Staining for both Smad2/3P and Smad1/5/8P demonstrated positive cells dispersed throughout the pellets at all time-points (data not shown). A strong positive staining for both Smad2/3P and Smad1/5/8P was seen in all terminally differentiated cells embedded in collagen II and collagen X matrix (Fig. 1C, D). In vivo terminally differentiated cartilage in the central region of embryonic (E16) metacarpal bones, rich in collagen X and MMP13, stained positive for both Smad2/3P and Smad1/5/8P (Fig. 3A–C). Therefore, both Smad signaling pathways are active in terminally differentiated cartilage, both in in vitro differentiated BMSC as well as in vivo during endochondral ossification.
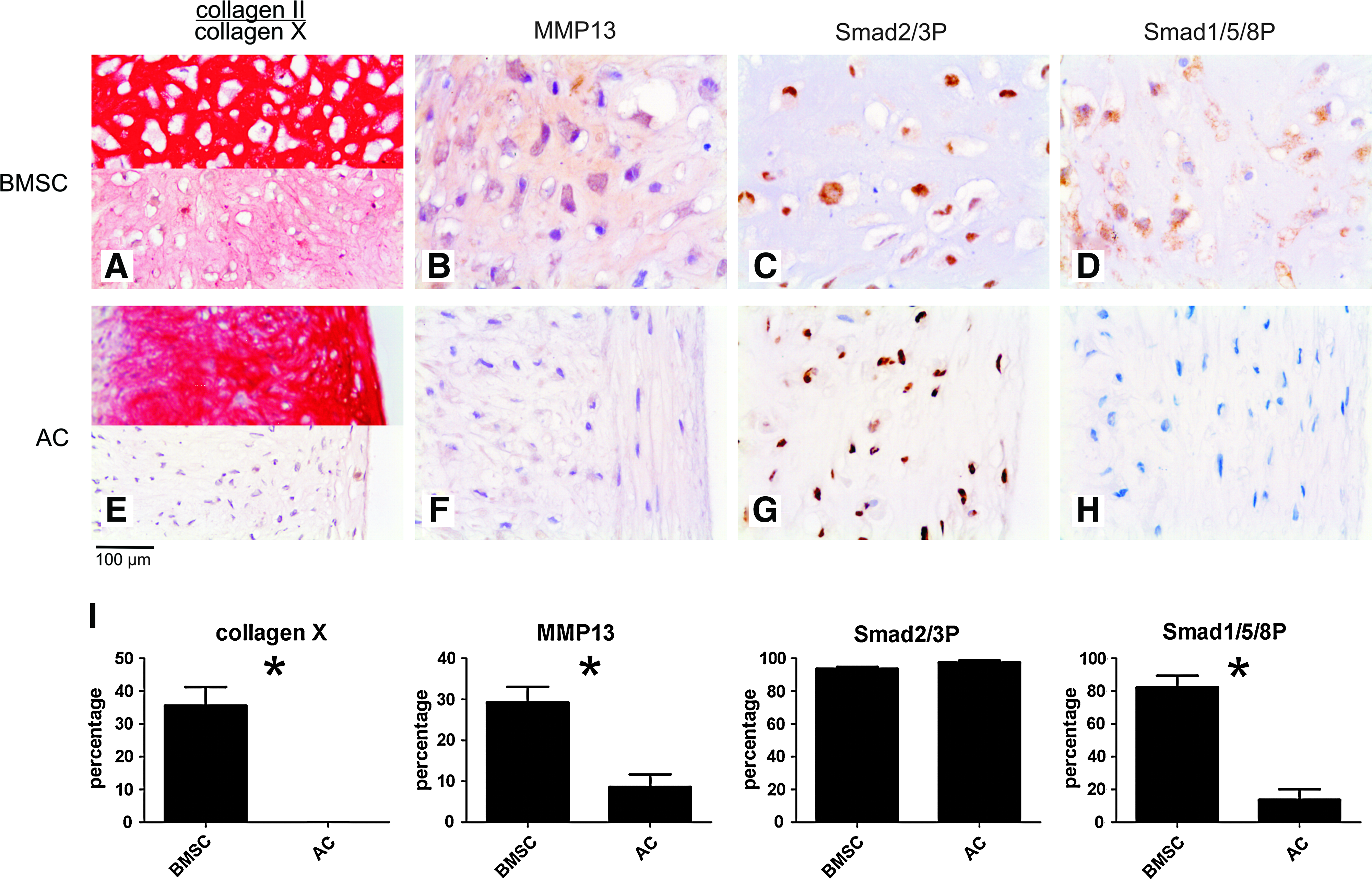
Immunohistochemistry of tissue-engineered cartilage from bone-marrow-derived mesenchymal stem cell (BMSC) and articular chondrocytes. (
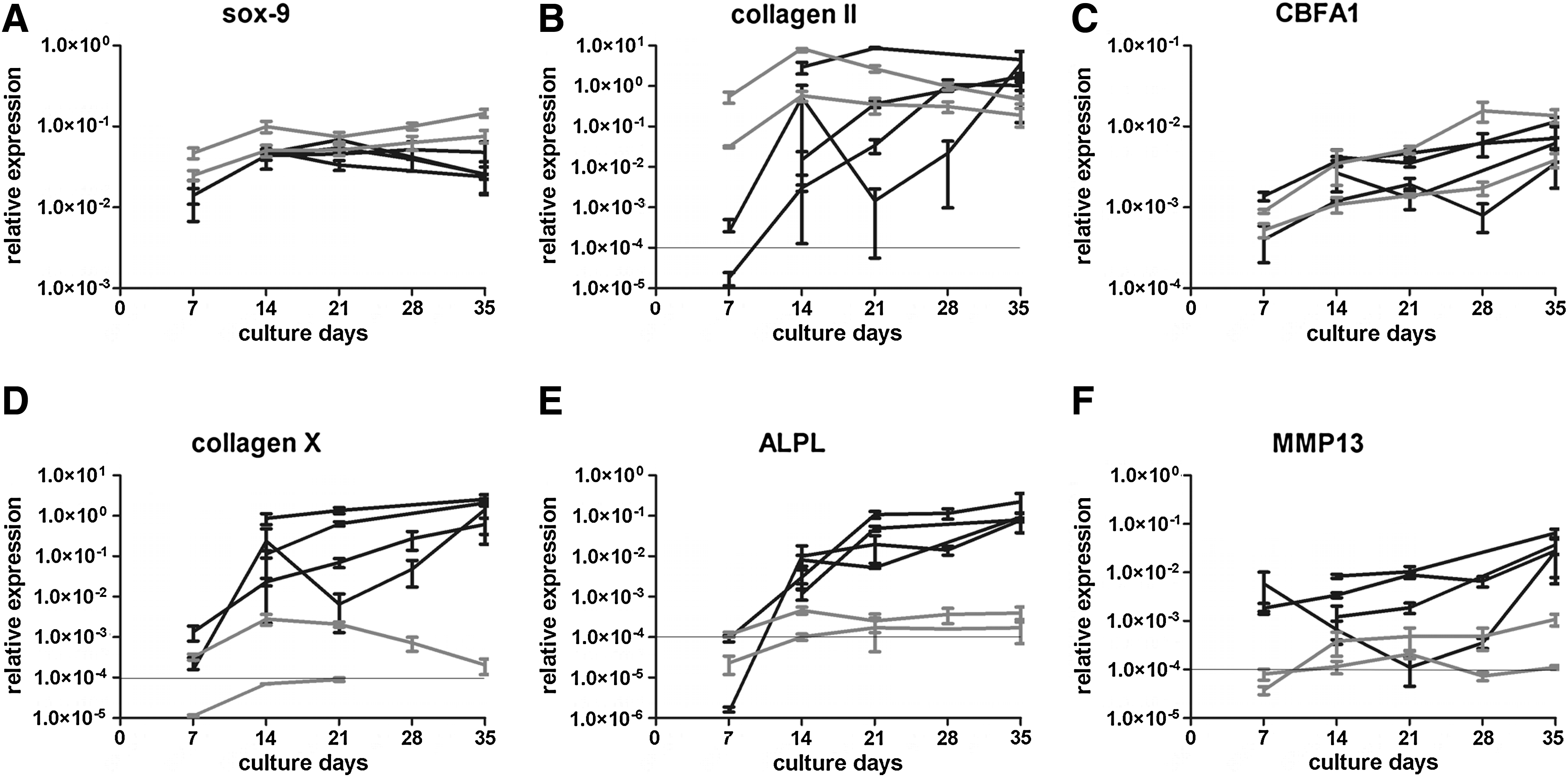
Gene expression during time course of chondrogenic differentiation of BMSC and articular chondrocytes. Gene expression for sox-9 (
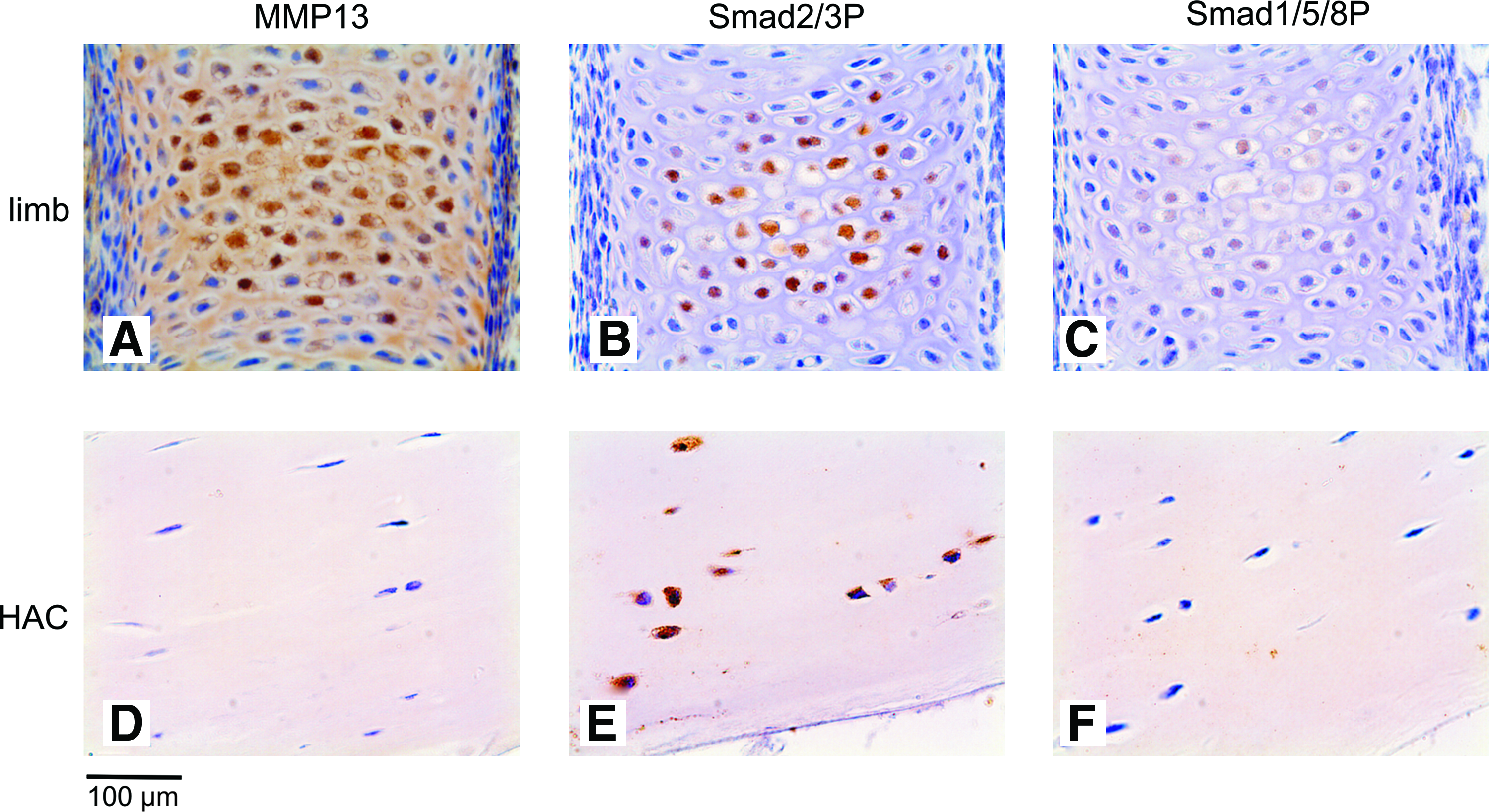
Immunohistochemistry of healthy terminally differentiated and hyaline cartilage in vivo. A brown staining demonstrates positivity for the antibody. (
Pellet cultures of expanded articular chondrocytes showed abundant collagen type II at day 35 (Fig. 1E). Chondrogenic differentiation of these cells was confirmed by a high expression of mRNA for sox-9 and collagen II. Articular chondrocytes reached a high expression of these chondrogenic markers earlier in culture than BMSCs, but the level of expression was comparable (Fig. 2). Collagen X, MMP13, and ALPL expression was only minimal on protein and mRNA level (Figs. 1E, 1F, and 2), indicating that tissue-engineered cartilage from articular chondrocytes resembles hyaline cartilage. Chondrocytes in these pellets stained strongly positive for Smad2/3P, whereas no Smad1/5/8P immunostaining was observed (Fig. 1G, H). This was similar to the healthy human hyaline articular cartilage samples (Fig. 3E, F).
Thus, signaling through Smad2/3 seems related to hyaline-like chondrogenic differentiation and signaling through Smad1/5/8 is related to terminal differentiation and specifically the upregulation of MMP13 expression.
Inhibition of Smad2/3 phosphorylation blocked chondrogenic differentiation of BMSCs
To investigate the importance of Smad2/3 signaling for chondrogenic differentiation, SB-505124 was added to BMSCs from three donors (donors 1–3) in chondrogenic pellet cultures. Exposure to SB-505124 from the onset of the culture completely blocked chondrogenic differentiation and collagen II production (data not shown).
Although no clear donor-to-donor differences were observed in gene expression of collagen II in the time-course experiment, timing of collagen II deposition in the matrix as seen on immunohistochemistry differed from donor to donor. Only in donor 1 collagen II was already present at day 14. Inhibition of Smad2/3 phosphorylation with SB-505124 prevented further collagen II production. While collagen II was present throughout the pellets in the controls, very limited collagen II production was seen in SB-505124-treated pellets after 35 days of culture (Fig. 4A, B). Donors 2 and 3 did not have a collagen II containing matrix at day 14, and addition of SB-505124 from day 14 on resulted in pellets without collagen II at day 35 (data not shown). When SB-505124 was added to these donors from day 21 on, when the first spots of collagen II depositions were seen in the matrix, further collagen II production was completely blocked as was the case in the first donor (Fig. 4C). Collagen X production was not seen in pellets treated with SB-505124, either from day 14 or from day 21 on, whilst no inhibitory effect was seen on MMP13 expression.

Immunohistochemistry and gene expression of MSC pellets at day 35 of chondrogenic differentiation, with or without blocking Smad2/3 phosphorylation days 14–35. (
These protein data were supported by gene expression analysis. In all three donors, the addition of SB-505124 form day 14 on significantly inhibited gene expression of collagen II, whereas expression of Sox-9 was not significantly influenced (Fig. 4G). Collagen X expression was inhibited on mRNA level corresponding to protein level in treated pellets of all three donors, and so was gene expression of ALPL. This is most likely due to the inhibited chondrogenic differentiation, hence delaying further terminal differentiation. Staining for MMP13 was positive in both treated and control pellets (Fig. 4E, F), and its gene expression was not significantly inhibited by addition of SB-505124 (Fig. 4G). In Figure 4 gene expression is displayed for pellets treated with SB-505124 from day 14 as addition from day 21 on resulted in similar trends.
Summarizing, we conclude that inhibition of Smad2/3 phosphorylation by the addition of SB-505124 prevented further cartilage differentiation and cartilage matrix production after the onset of chondrogenic differentiation, whereas it had no inhibiting effect on MMP13 expression.
Inhibition of Smad1/5/8 phosphorylation inhibited terminal differentiation and mineralization in differentiated chondrocytes but completely blocked early chondrogenic differentiation
To test the hypothesis that Smad1/5/8 signaling is related to terminal differentiation, dorsomorphin was added to the chondrogenic differentiation medium of BMSCs of three donors (donors 2–4) to prevent phosphorylation of Smad1/5/8. Addition of dorsomorphin throughout the culture period completely prevented chondrogenic differentiation of BMSCs. Thus, Smad1/5/8P is important during the onset of chondrogenesis.
Addition of dorsomorphin from day 14 on had no evident inhibitory effect on further cartilage formation in the donor where cartilage matrix was already deposited at day 14 (donor 4). In these pellets abundant collagen II was seen on immunohistochemistry in treated pellets after 35 days of culture, not differing from the control (Fig. 5A, B). In the two other donors with less chondrogenic capacity, no collagen II was present at day 14 and only small spots at day 21. Although, in these cases addition of dorsomorphin inhibited collagen II production compared to the control, the collagen II positive area further increased in time, and at day 35 more collagen II was seen than after the addition of SB-505124. Collagen X production and MMP13 expression were inhibited in dorsomorphin-treated pellets compared to the controls. Most importantly, in the donor where an abundant cartilage matrix was deposited in the presence of dorsomorphin after the onset of chondrogenesis, this collagen II rich matrix was lacking collagen X and MMP13 expression, indicating that this cartilage-like matrix was not terminally differentiated.
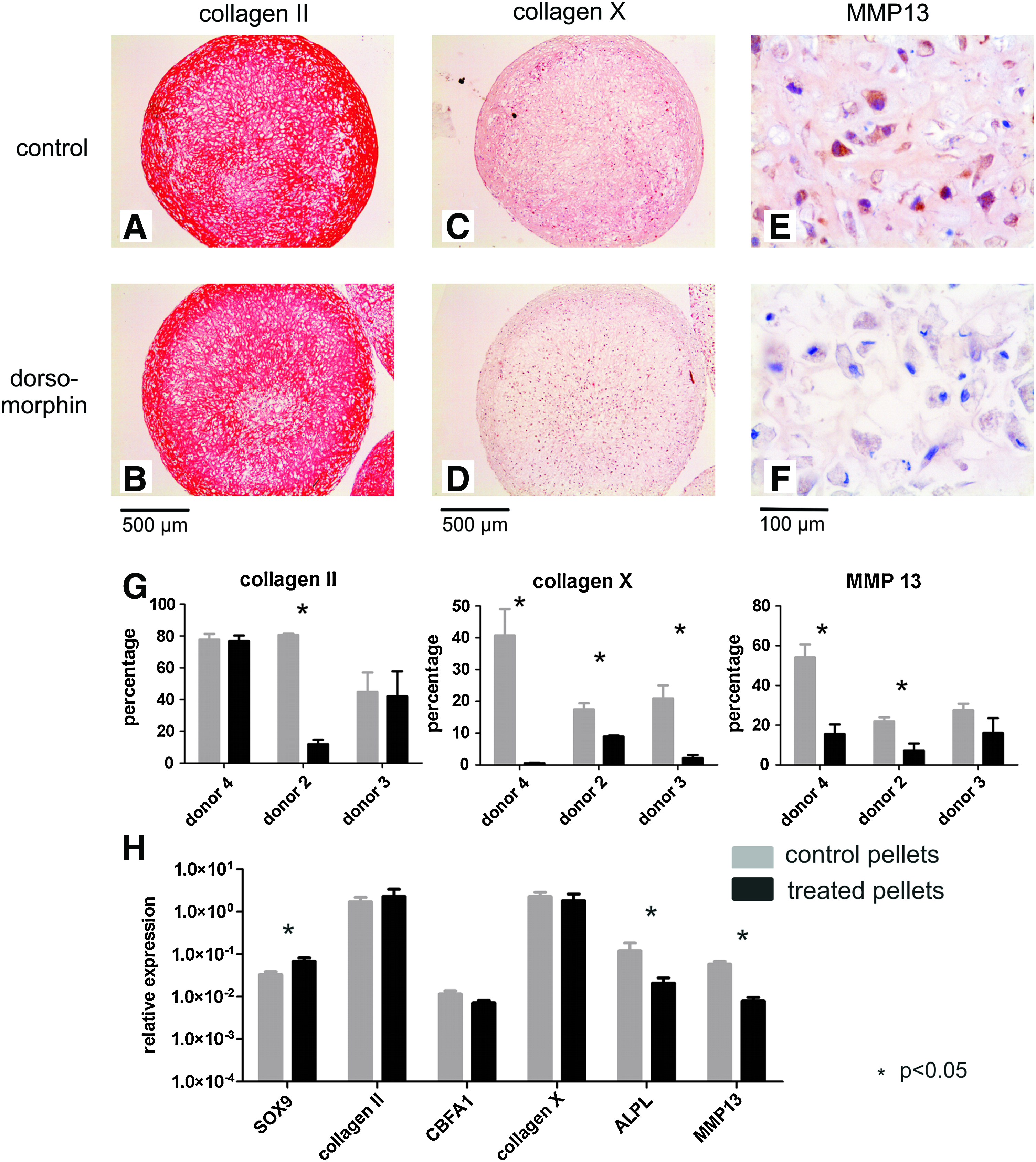
Immunohistochemistry and gene expression of MSC pellets at day 35 of chondrogenic differentiation, with or without blocking Smad1/5/8 phosphorylation on days 14–35. (
This effect of dorsomorphin, inhibiting terminal differentiation while allowing collagen II production, was even more evident when studying gene expression. SOX-9 expression was significantly higher than in the controls (Fig. 5G). Although no significant difference was observed in gene expression of CBFA1 or collagen X at day 35, the intensity of the collagen X staining was clearly diminished in the treated pellets compared to the control (Fig. 5C, D). Gene expression of ALPL was somewhat reduced when dorsomorphin was added to the medium, though not significantly (p = 0.13) (Fig. 5G). Moreover, both gene expression and immunohistochemistry for MMP13 was significantly reduced in treated pellets compared to the control (Fig. 5E–G). Therefore, inhibiting Smad1/5/8 phosphorylation by addition of dorsomorphin after initial chondrogenic differentiation allowed further cartilage production and inhibited terminal differentiation.
To study whether blocking Smad1/5/8 phosphorylation after initial chondrogenic differentiation not only inhibits terminal differentiation but also prevents mineralization, an additional experiment was performed with cells from the donor with very good chondrogenic capacity. β-Glycerophosphate was added to the medium from days 35–49 to allow mineralization of the matrix. While mineralization was clearly present in control pellets, blocking Smad1/5/8 phosphorylation completely prevented mineralization as was demonstrated by von Kossa and Alizarin Red staining (Fig. 6).
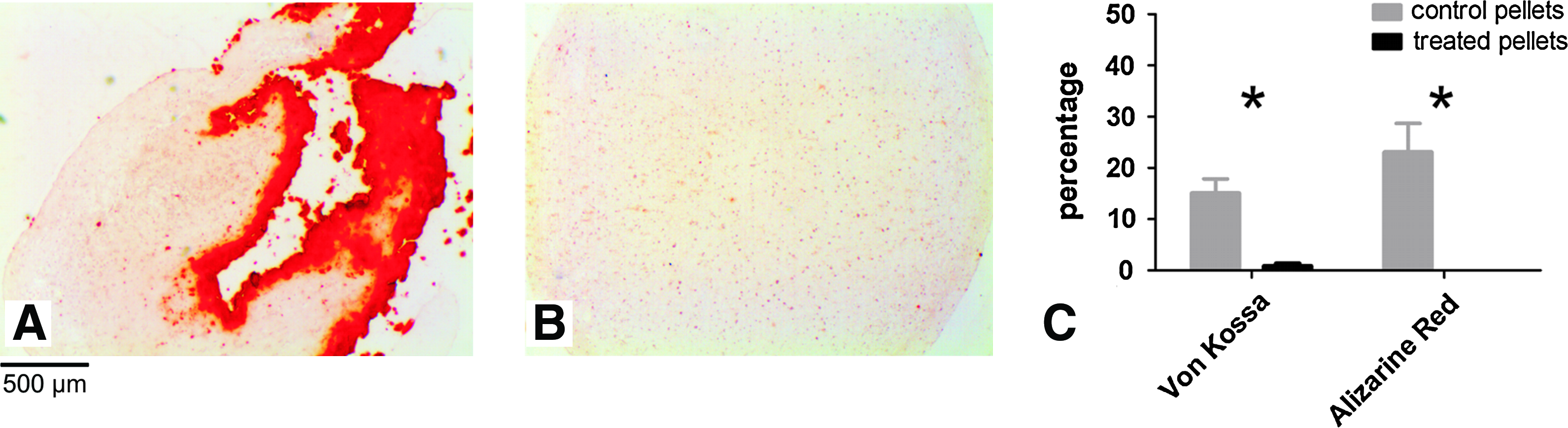
Blocking Smad1/5/8 phosphorylation prevents mineralization. (
Discussion
Generation of stable hyaline cartilage from BMSCs is currently still a challenge. Although chondrogenesis and cartilage formation are achieved, it eventually leads to terminal differentiation of chondrocytes instead of the production of stable hyaline cartilage. Further, the tissue-engineered cartilage construct is not stable when it is implanted in vivo but mineralizes.4,5 The process of in vitro chondrogenic differentiation therefore seems a recapitulation of endochondral bone formation. A better knowledge of mechanisms determining chondrocyte differentiation and terminal differentiation is therefore crucial to control the chondrogenic differentiation of BMSCs. To our knowledge this study is the first to show the possibility to direct chondrogenic differentiation of BMSCs toward a more stable hyaline-like phenotype by targeting Smad signaling stage specifically.
Both Smad2/3 and Smad1/5/8 phosphorylation is required for onset of chondrogenic differentiation as blocking of either one from the start of culture completely prevented chondrogenesis of BMSCs. Our results are supported by a study by Roark and Greer, demonstrating in chick limb mesenchymal cells that both TGF-β and BMP are important for early chondrogenesis. 25 However, in the present study we demonstrate that the different Smad signaling pathways have a very distinct function during later stages of chondrogenesis. By affecting Smad signaling in BMSC pellets from days 14 to 35, it was possible to study the function of both pathways after the onset of chondrogenic differentiation. Blocking Smad2/3 phosphorylation prevented further increase in collagen II expression on both mRNA and histology level, but did not alter MMP13 expression. This confirms our previous findings that expression of ALK5, one of the main activators for Smad2/3 phosphorylation, correlates with collagen II expression in human articular cartilage, but not with MMP13 expression. 19 Smad2/3 phosphorylation has been shown to block terminal differentiation. 15 Thus, besides impairing cartilage formation directly, blocking Smad2/3 might release the block on terminal differentiation and via this way may allow MMP13 expression. This is also suggested by the enhanced terminal differentiation, in addition to the cartilage loss, seen in Smad3-deficient mice, 16 and the fact that reduced TGF-β signaling via Smad2/3, by proteosomal degradation of Smad2 and 3 through enhanced expression of Smurf2, stimulates endochondral ossification. 26 Smad2/3 signaling is important not only for chondrogenesis, but also for the reparative response of cartilage. We have previously shown a reduced ALK5 expression and reduced responses to TGF-β in aged mice. 27 Moreover, reduced Smad2 phosphorylation was accompanied by an impaired repair response of the cartilage.
In contrast to Smad2/3, blocking Smad1/5/8 phosphorylation from day 14 on did allow further cartilage matrix production. Moreover, it prevented expression of both collagen X and MMP13 on histology and inhibited mineralization. Cbfa1 and Smad1 and 5 have been suggested crucial for terminal differentiation.28,29 We found no effect on CBFA1 mRNA level, but cbfa1 requires Smad interaction for induction of an osteogenic signal.29,30 Cbfa1 is required for MMP13 expression and it can be anticipated that the lack of Smad1/5/8-CBFA1 interaction in presence of dorsomorphin is a direct cause for the reduction in MMP13 found in these chondrogenic pellets.31,32
Our findings are in line with the results of Valcourt et al. 33 showing the involvement of Smad1,5,8 in BMP2-induced terminal differentiation of chondrocytes and our previous study that demonstrated that ALK1 correlates with MMP13 expression in articular cartilage, whereas ALK5 does not. 19 Smad1 and 5 are crucial for terminal differentiation during embryonic development, as was previously demonstrated in Smad1/5CKO mutants that had a disorganized growth plate with loss of terminal chondrocytes and very limited collagen X expression. 17
The observed Smad1/5/8 signaling in BMSCs may be due to signaling of the exogenous TGF-β to the ALK1 receptor. However, we have seen in a previous study that chondrogenically differentiated BMSCs express BMP2 (data not published). Therefore, it is possible that the observed Smad signaling is due to not only exogenous but also endogenous signaling involving BMP. The precise mechanism of Smad phosphorylation would require further research.
These experiments confirm the importance of using different donors because of the variability in timing of chondrogenic differentiation. Although gene expression of collagen II did not obviously differ from donor to donor, timing of collagen II gene expression and deposition in the matrix did differ, thereby influencing the results of blocking Smad signaling. The results of inhibition of Smad1/5/8 phosphorylation by dorsomorphin clearly demonstrate the importance of targeting the chondrogenic processes at the exact right moment. BMSCs of a donor that had an excellent chondrogenic capacity and formed cartilage within 2 weeks performed very well with dorsomorphin. Cartilage was formed, terminal differentiation was inhibited, and mineralization was prevented. However, chondrogenesis of BMSCs of another donor that took 3 weeks to form a small amount of cartilage was prevented by adding dorsomorphin from day 14, whereas addition from day 21 did allow further cartilage formation and inhibited terminal differentiation. This variation in cultures highlights the stage specificity of Smad signaling during chondrogenesis. For future studies it may be interesting to study whether a more restricted administration of dorsomorphin upon the onset of terminal differentiation will completely block terminal differentiation or will just merely temporarily inhibit this process, avoiding any adverse effects the continuous administration of dorsomorphin may have. However, the donor-to-donor variation in timing of differentiation will make it difficult to identify during culture the exact timing of the onset of terminal differentiation and the administration of inhibitors.
In conclusion, our data strongly suggest that the role of Smad2/3 and Smad1/5/8 phosphorylation in chondrogenesis by human BMSCs is stage dependent. Based on our experiments, the timing of Smad2/3 and Smad1/5/8 phosphorylation, relative to the expression of known chondrogenic differentiation markers, is schematically demonstrated in Figure 7. Both pathways are crucial for initial chondrogenesis. While Smad2/3 is important for ongoing cartilage matrix production, Smad1/5/8 is crucial for terminal differentiation and mineralization. Most importantly, our data imply that in vitro cartilage tissue engineering might benefit from blocking the Smad1/5/8 route after chondrogenic differentiation of BMSCs is induced, to prevent terminal differentiation and mineralization while sustaining further cartilage–matrix production.
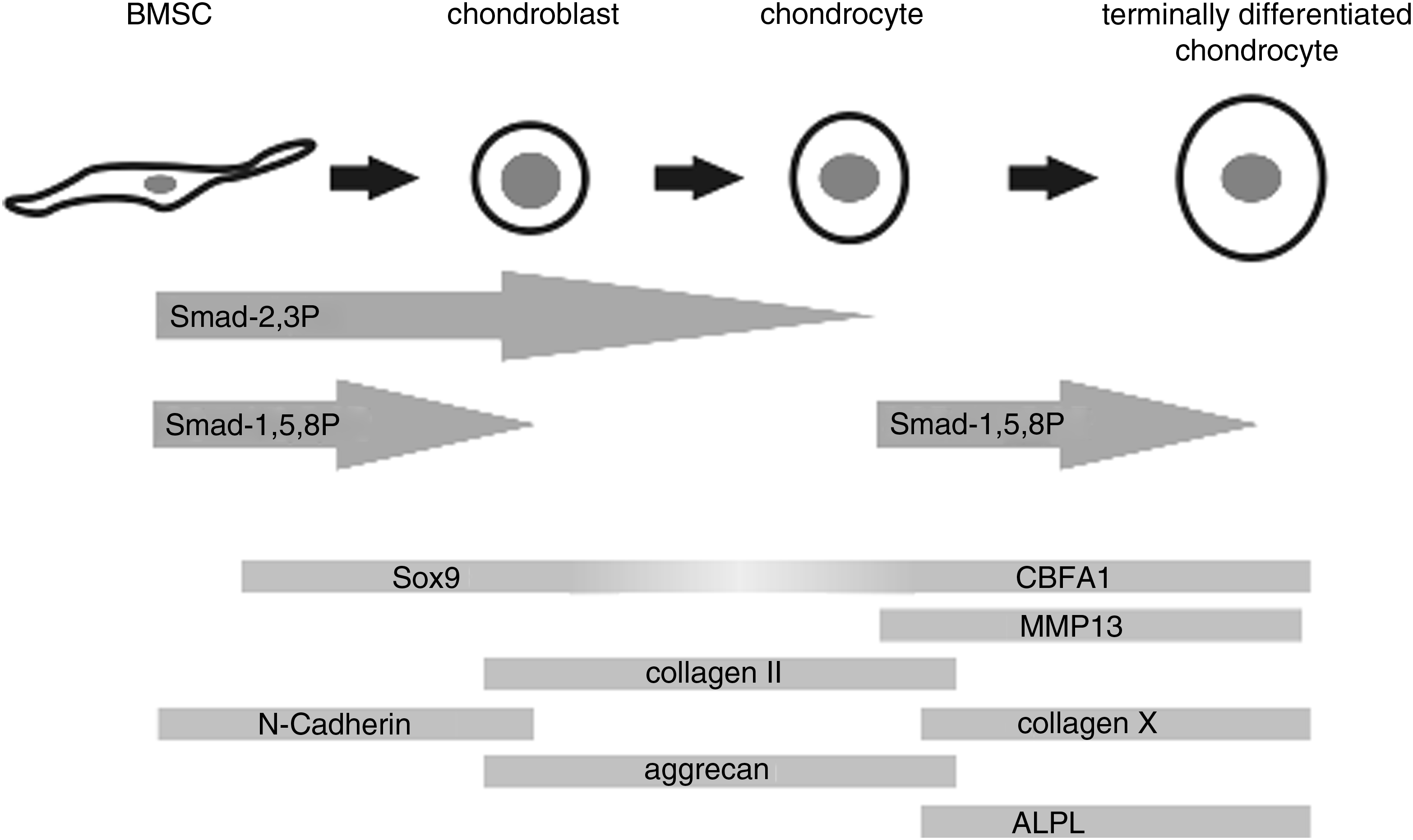
Schematic figure of probable timing of Smad2/3 and Smad1/5/8 signaling during chondrogenic differentiation of BMSCs, relative to some known markers of differentiation.
Footnotes
Acknowledgments
The authors would like to thank Marijn Rutgers (UMC Utrecht, The Netherlands) for isolating articular chondrocytes and Dorothy Frenz and Wei Liu (Albert Einstein College of Medicine, Bronx, NY) for providing us with murine embryonic limbs. This research was financially supported by the Dutch Program for Tissue Engineering.
Disclosure Statement
No competing financial interests exist.