
Abstract
Structural extracellular matrix molecules gain increasing attention as scaffolds for cartilage tissue engineering owing to their natural role as a growth factor repository. We recently observed that a collagen-type I/III (Col-I/III) matrix, human recombinant transforming growth factor-beta (TGF-β) protein, and fibrin hydrogel (FG) combined to a biphasic construct provided sufficient long-term TGF-β support to drive in vitro chondrogenesis of human mesenchymal stem cells (hMSC). Here we ask whether FG and Col-I/III can both retain TGF-β, describe the influence of cell seeding on TGF-β release, and compare the molecular path of hMSC chondrogenic differentiation under soluble versus local TGF-β supply. Release of growth factor from scaffolds augmented with increasing amounts of TGF-β was analyzed over 7 days and chondrogenesis was assessed over 42 days. Low TGF-β release rates from Col-I/III as opposed to higher release from FG indicated that both molecules retained TGF-β, with Col-I/III being the superior storage component. Cell seeding enhanced TGF-β retention in FG by about threefold and almost stopped release beyond 24 h. TGF-β remained bioactive and supported MSC chondrogenesis without impairing the amount of proteoglycan and collagen-type II deposition per cell and per construct compared to standard scaffold-free MSC pellets supplied with soluble TGF-β. Local TGF-β, however, mediated lower cell content, less collagen-type X relative to collagen-type II deposition and no matrix metalloproteinase-13 up-regulation. In conclusion, cells quickly halted release of local TGF-β from FG, turning FG and Col-I/III into attractive TGF-β repositories capable to drive full hMSC chondrogenesis, but via a modulated differentiation pathway. Since only part of the changes was reproduced by transient soluble TGF-β supply, release kinetics alone could not explain the molecular differences, suggesting that local TGF-β acts distinct from its soluble counterpart.
Introduction
Major challenge in biomaterial research is that each individual tissue demands specific scaffold characteristics. In the case of cartilage tissue engineering, cartilage avascularity and poor cellularization restrict intrinsic regeneration capacity so that cell-delivery per se is imperative. Moreover, the regenerative tissue has to survive low oxygen and nutrient supply, while for integration into the host tissue, synthesis and stabilization of a cartilaginous matrix are main features in addition to resistance to the mechanical loads cartilage in general is subjected to.
In vivo, cytokines including growth factors are the main regulators of such cellular processes with the extracellular matrix (ECM) functioning as reservoir and modulator of growth factors.4–6 Indeed, many structural molecules in cartilage including collagens have growth factor binding properties.7–10 Natural ECM molecules are thus attractive to generate functionalized bioactive scaffold materials by augmentation with growth factors.
In cartilage tissue engineering, hydrogels composed of ECM proteins like fibrin 11 or formed by polymers like poly-(ethylene glycol)12,13 are frequently used scaffolds since uniform cell distribution makes them ideal cell carriers14–16 and bioactive molecules can easily be entrapped during polymerization of the gels.13,17–20 Fibrin-based biomaterials are biocompatible, exhibit controlled biodegradability,21,22 and support tissue repair 23 ; moreover, allogeneic fibrin sealants are already Food and Drug Administration (FDA)-approved and in clinical use.
For improved mechanical stability, fibrin has frequently been combined with solid biomaterials, in the clinical treatment of cartilage defects particularly with collagen-type I/III (Col-I/III) matrix, which is known to maintain the chondrocyte phenotype in vitro and in vivo. 24
Unless bound by structural matrix molecules, growth factors in general are small enough molecules to diffuse freely in a hydrogel network and might consequently be lost into the surrounding media too fast to work efficiently. Various growth factor delivery systems have been used for continuous controlled release of growth factors from hydrogels, including entrapment within liposomes 19 or nano-/microparticels,25–27 covalent conjugation,28,29 or indirect binding via short peptide sequences known to bind specific growth factors.12,30 Interestingly, our previous results implicated that a biphasic scaffold composed of a Col-I/III carrier and fibrin hydrogel (FG) maintained fibrin-entrapped transforming growth factor-beta (TGF-β) bioactivity over several weeks in culture. 31 Human mesenchymal stem cells (hMSCs) seeded into the bicomposites differentiated into cells with a chondrocyte phenotype in the absence of TGF-β medium supplementation although chondrogenic in vitro differentiation of hMSCs is highly sensitive and requires a sustained high TGF-β level for several weeks. 32
Aim of this study was to characterize local versus soluble TGF-β supply by biomaterials composed of distinct structural ECM molecules. We asked whether Col-I/III and FG represent an adequate TGF-β reservoir, investigated dose-related release kinetics in the presence and absence of MSCs, and determined whether local TGF-β supply would alter the quantity or the molecular route of chondrogenic differentiation compared to continuous medium supply.
Materials and Methods
Preparation of TGF-β-augmented FG biomaterials
For preparation of TGF-β1-augmented FG (Tissucol Duo S™, Baxter), the fibrinogen component was diluted 10-fold with phosphate-buffered saline (PBS) and mixed with TGF-β1 (PeproTech; 6 ng/mL) in a 2:1 ratio. About 17.5 μL of this TGF-β1-fibrinogen solution (final fibrinogen concentration 3 mg/mL, 35 ng TGF-β1) was mixed with 17.5 μL thrombin (5 IU/mL final).
Round disks (diameter 4 mm) from a Col-I/III matrix (Chondro-Gide®, Geistlich Pharma AG) were prepared with biopsy punches. Ten microliters of TGF-β1 (3.5 ng/μL) was mounted onto each disk and incubated for 1 h at 37°C in humidified atmosphere to allow growth factor attachment. Thirty-five nanograms of TGF-β was chosen since this corresponded to the cumulative soluble TGF-β dose applied to cell-seeded constructs over 6 weeks in later experiments (18×200 μL).
Bicomposites were prepared by quickly mixing 9 μL fibrinogen solution containing 35 ng TGF-β with 9 μL thrombin and immediate transfer onto 4-mm Col-I/III membranes for polymerization. FG and bicomposites were incubated at 37°C for 30 min to allow stabilization of the hydrogel.
TGF-β release from biomaterials
Monophasic (FG, Col-I/III) and biphasic constructs (triplicates, five independent experiments) were augmented with 35 ng TGF-β1 and incubated with 200 μL growth factor-free chondrogenic basal medium composed of Dulbecco's Modified Eagles Medium (DMEM) high glucose (Gibco, Invitrogen) supplemented with 0.1 μM dexamethasone, 0.17 mM ascorbic-acid 2-phosphate, 5 μg/mL transferrin, 5 ng/mL sodium-selenite, 1 mM sodium-pyruvate, 0.35 mM proline, 1.25 mg/mL bovine serum albumin (BSA) (all from Sigma-Aldrich), 5 μg/mL insulin, and 100 units/mL penicillin/100 μg/mL streptomycin. Composites were incubated at 37°C with 6% CO2 in humidified atmosphere for a total of 7 days. The medium was changed five times a week and the collected supernatants were frozen at −80°C until further analysis.
The TGF-β1 amount released from the composites was detected using an enzyme-linked immunosorbent assay (ELISA, Quantikine® Human TGF-β1; R&D Systems). Standards were prepared with the same TGF-β1 lot used for cell culture in replacement of the standard included in the assay. Cumulative TGF-β release was calculated and values related to day 7 total cumulative release from FG. TGF-β remaining within the composites after 7 days of culture was measured by dissolving FG with urokinase (1 IU/mL in 0.9% NaCl and 1.5% BSA; Medac GmbH) and Col-I/III with collagenase B (1.5 mg/mL in PBS; Roche) until the scaffolds were solubilized completely (2–4 days, changing the enzyme solution daily).
MSC isolation and culture
MSCs were isolated from fresh bone marrow aspirates after informed consent of 14 patients (mean age 51±24 years) undergoing total hip replacement or osteotomy. The study was approved by the local ethics committee. The mononuclear cell fraction was separated by Ficoll density gradient centrifugation, washed, and seeded in the expansion medium (DMEM high glucose w/o
Preparation of MSC-loaded constructs
MSCs were embedded into FG by mixing 350,000 cells with 17.5 μL fibrinogen-TGF-β solution and subsequent polymerization with 17.5 μL thrombin as described above. After incubation at 37°C and 6% CO2 for 1 h, 200 μL chondrogenic basal medium was added to each composite.
TGF-β1-augmented Col-I/III was seeded with 350,000 cells per disc by loading 10 μL cell suspension on top of each membrane. To allow cell attachment, constructs were incubated for 1 h at 37°C, 6% CO2. For bicomposites, growth factor-free Col-I/III was seeded with 10 μL cell suspension and covered with 18 μL TGF-β1-fibrinogen-thrombin solution as described above.
Positive controls comprised cell-seeded, but TGF-β1-free, constructs that were supplied with the chondrogenic medium containing 10, 20, or 30 ng/mL TGF-β1 as indicated. In dose–response groups, 35, 70, or 105 ng was embedded in FG, respectively. One further group of cell-loaded FG constructs received the total amount of 35 ng TGF-β1 supplied via the chondrogenic basal medium only on day 1 of culture, but no TGF-β1 thereafter. Negative controls comprised MSC-loaded FG scaffolds without TGF-β. Transient TGF-β supply was performed with FG-embedded MSCs receiving 30 ng/mL TGF-β for the first 2 weeks, followed by decreasing TGF-β with each medium exchange (20 ng/mL–10 ng/mL–5 ng/mL) within the third week and TGF-β-free culture for the remaining 3 weeks. All constructs were cultured for 6 weeks with the medium changed thrice a week. Culture supernatants of all spheroids of each group were pooled at days 7, 14, 21, 35, and 42 and frozen at −80°C for further analysis of matrix metalloproteinase-13 (MMP-13) secretion. Two constructs per group were prepared for histology. The remaining composites were washed with PBS and frozen at −80°C for analysis of proteoglycan and collagen deposition.
Histology
Constructs were washed with PBS, fixed in 4% formaldehyde, dehydrated, and embedded into paraffin. Five-micrometer sections were stained with Safranin O (0.2% in 1% acetic acid) and counterstained with fast green (0.04% in 0.2% acetic acid). Type II collagen was stained with mouse anti-human monoclonal antibodies (MP Biomedicals/Quartett) according to standard immune histochemistry protocols with fast red counterstaining. 31
Quantification of proteoglycan and collagen deposition
Proteoglycans and collagen II were quantified from the same samples (n=3 per donor, six independent experiments) by fractionated extraction with guanidine hydrochloride. 33 Guanidine-HCl extracts (4 M in 50 mM Tris-HCl, pH 8.5) were prepared from three homogenized constructs per group. Proteoglycans were stained with 3% alcian blue, washed with dimethyl sulfoxide (40% in 0.05 M MgCl2), and resolved in guanidine/propanol buffer (4 M guanidine-HCl, 33% isopropanol, 0.25% Triton X-100). Standards were prepared from chondroitin sulphate. Alcian blue absorption was measured in a microplate reader at 650 nm. Proteoglycans were normalized to DNA content, which was determined fluorimetrically after staining the guanidine-HCl extracts with Hoechst 33258.
The pellets remaining after guanidine-HCl extraction were further processed for collagen extraction. Therefore, pellets were digested with 1.1 mL pepsin (2.5 mg/mL) overnight at 4°C. Pepsin digestives of three parallel pellets were pooled and neutralized, and collagens were extracted with 4.5 M NaCl overnight at 4°C. Collagens were precipitated after addition of 0.4 M NaCl in 0.1 M Tris (pH 7.4) with 1.2 mL ethanol per sample for 4 h at −20°C. Collagen pellets were stored at −20°C before resolving in 100 μL lysis buffer (50 mM Tris, 150 mM NaCl, 1% Triton X-100, pH 8.0) for type II collagen ELISA (type II collagen detection kit; Chondrex, Inc.).
For collagen-type II and X Western blotting, lysates were separated by denaturing sodium dodecyl sulfate–polyacrylamide gel electrophoresis and proteins blotted onto a nitrocellulose membrane. The lower part of the membrane was incubated with mouse anti-human collagen-type X antibody (Quartett) and the upper part with mouse anti-human collagen-type II antibody (MP Biomedicals). Bands were visualized with peroxidase-coupled goat anti-mouse antibody using the ECL detection system (Roche).
Quantification of MMP-13 secretion
MMP-13 secretion was detected in pooled cell culture supernatants of four independent experiments using an ELISA (Quantikine Human pro-MMP-13 Immunoassay; R&D Systems).
Statistical analysis
Statistical analysis of differences between experimental groups was performed with Mann-Whitney U-test for direct comparison of two groups, with a p-value<0.05 considered statistically significant.
Results
Release kinetics of TGF-β added to biomaterials
The growth factor release kinetics from FG, Col-I/III, and bicomposites (FG plus Col-I/III) was determined after immobilization of 35 ng TGF-β1. All three composites showed an initial burst release of loosely bound or nonadsorbed TGF-β into the chondrogenic basal medium within the first 24 h followed by a slow and declining release (Fig. 1A). Significantly more TGF-β was released from FG than from Col-I/III (p=0.008). Monophasic and biphasic Col-I/III scaffold groups revealed very low release rates with a total release within 7 days of 1.4±0.3 ng (median±standard error of the mean [SEM]) from Col-I/III, and 0.9±0.5 ng from bicomposites compared to 11.7±4.3 ng from FG, suggesting a higher TGF-β retention capacity when Col-I/III is present (Fig. 1B).
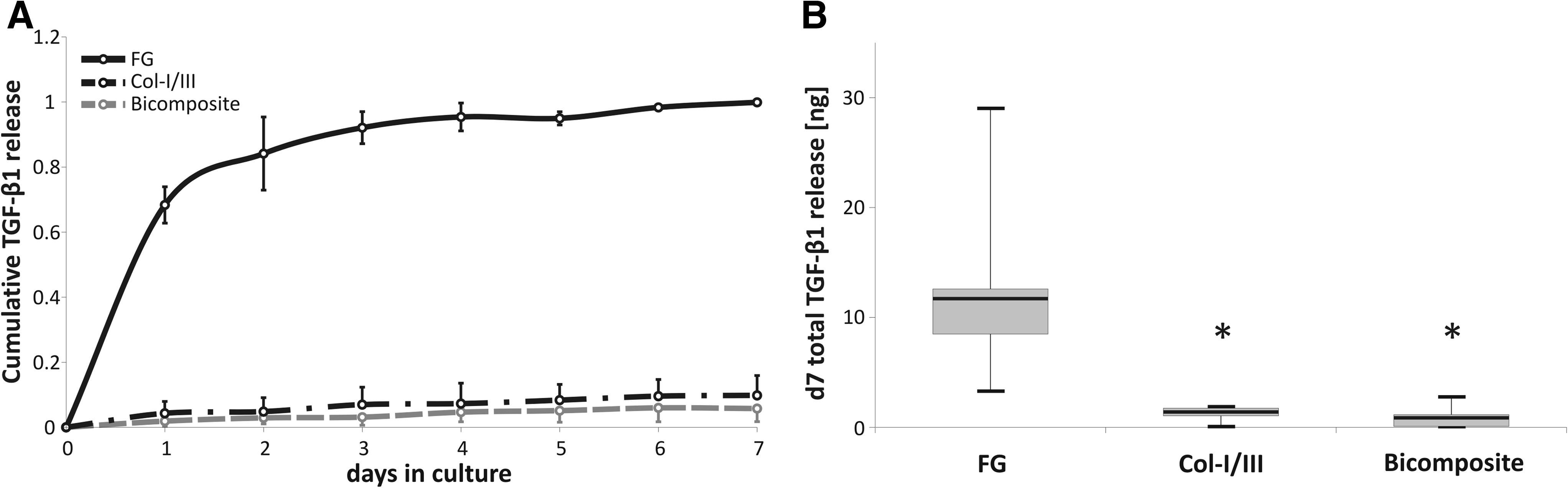
Cumulative release of local transforming growth factor-beta 1 (TGF-β1).
TGF-β remained biologically active within the scaffolds in considerable amounts, and we performed a functional bioassay and tested whether the local growth factor could drive MSC chondrogenesis.
Local TGF-β remains bioactive
Bioactivity of TGF-β1-augmented scaffolds was tested by culturing MSC-seeded constructs in the chondrogenic basal medium (no growth factor) for 6 weeks. Control samples received either full chondrogenic medium (with 10 ng/mL TGF-β, corresponding to a cumulative dose of 36 ng in 6 weeks, positive control) or a single pulse of soluble TGF-β (35 ng soluted in 200 μL medium) for 24 h to mimic a quick release profile. MSCs underwent chondrogenic differentiation under local TGF-β in all scaffold types and deposited a proteoglycan and collagen-type II-rich ECM similar to the biomaterial-free positive control pellets (Fig. 2A). None of the samples in the corresponding 24-h pulse control group showed histological signs of differentiation with one exception. One very small area of collagen-type II deposition was observed at the edge of a Col-I/III 24 h pulse sample (Fig. 2B), indicating that the Col-I/III matrix may have some TGF-β retention capacity even when exposed to lower concentrated TGF-β during the 24-h pulse period.
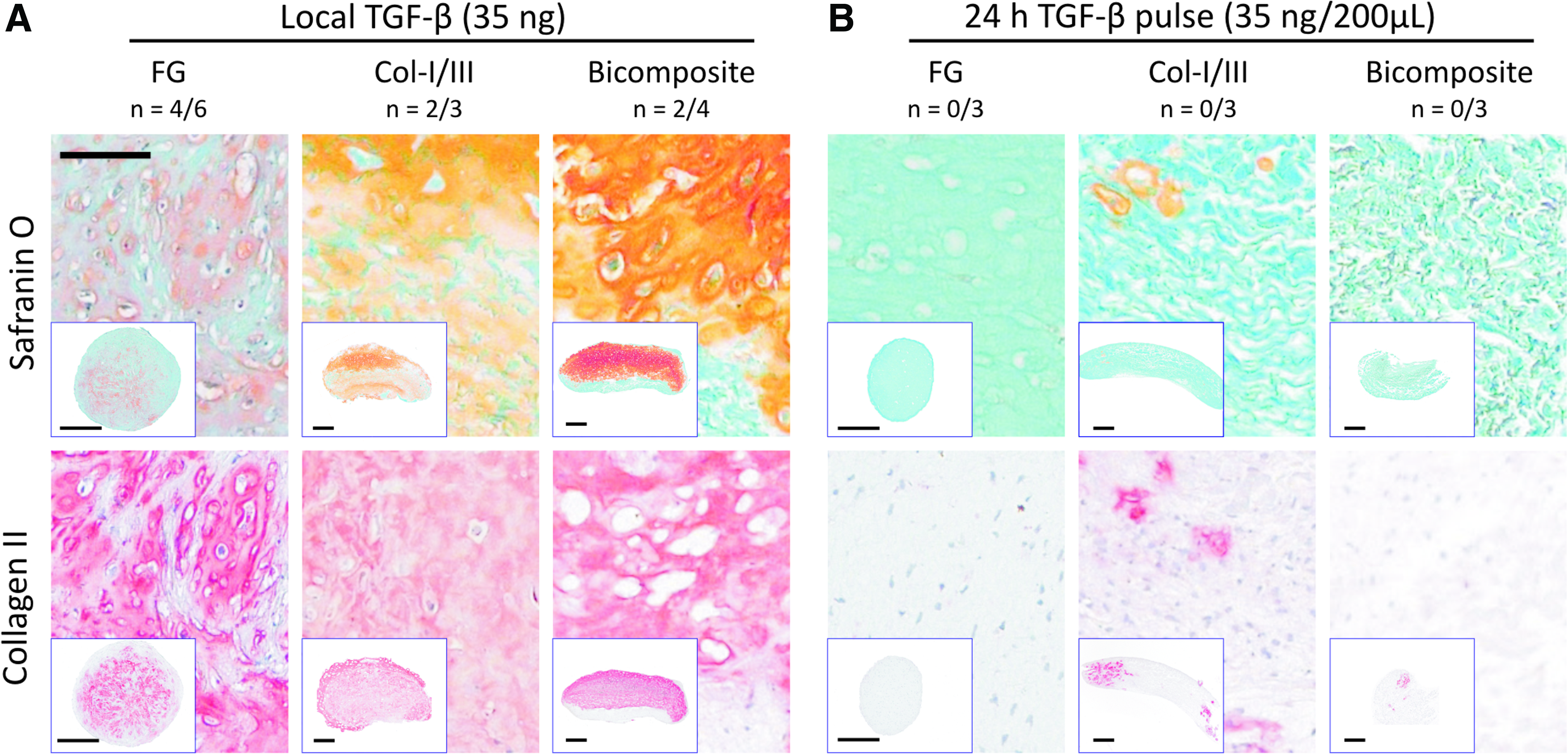
Deposition of proteoglycans and collagen-type II in TGF-β-augmented constructs. Safranin O and collagen-type II stainings of FG composites, Col-I/III membranes, and bicomposites each seeded with 350,000 human mesenchymal stem cells (hMSCs) and either augmented with 35 ng TGF-β1
Thus, FG as well as Col-I/III and bicomposites retained sufficient bioactive TGF-β to drive chondrogenesis similar to positive control pellets over 6 weeks although considerable TGF-β amounts escaped immune detection in recovery experiments.
Reduced TGF-β release from cell-seeded FG
We next asked how embedding of 350,000 hMSCs might affect growth factor release rates into the chondrogenic basal medium and selected FG as a carrier since little release was obvious for the Col-I/III and the bicomposite groups in Figure 1. A significantly lower cumulative growth factor release was seen after 7 days for the cell-loaded group (total release of 5.9±1.4 ng; median±SEM; p=0.001) compared to the cell-free constructs (23.5±3.8 ng) (Fig. 3). Cells particularly affected the secondary phase of low and steady release from day 2 on during which 32% of total released TGF-β was deliberated from cell-free FG, but only 8% from hMSC-loaded gels (p=0.0006). This argues in favor of a cell-mediated retention, metabolization, or degradation of TGF-β. Alternatively, the encapsulated cells themselves might act as an extra crosslink within the hydrogel, thereby increasing scaffold crosslink density and lowering mesh size, which can significantly lower growth factor release kinetics.34,35
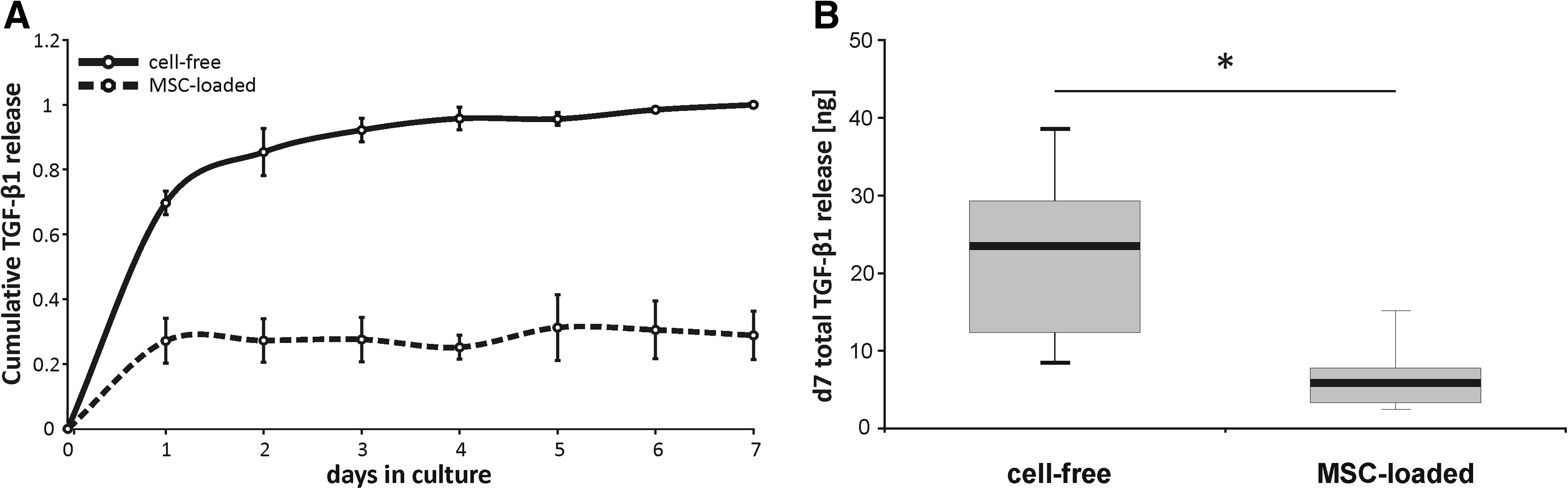
Cumulative TGF-β1 release from cell-free and hMSC-loaded constructs.
Effect of TGF-β dosage on release kinetics and cellular activity
When the amount of TGF-β1 entrapped in FG was enhanced from 35 ng (1× TGF) to 70 ng (2× TGF), and 105 ng TGF-β1 (3× TGF) per construct, release kinetics were similar, but proportionally enhanced depending on the initially immobilized TGF-β amount (Fig. 4). Again, embedding of hMSCs reduced the release significantly by about threefold independent of dosage.
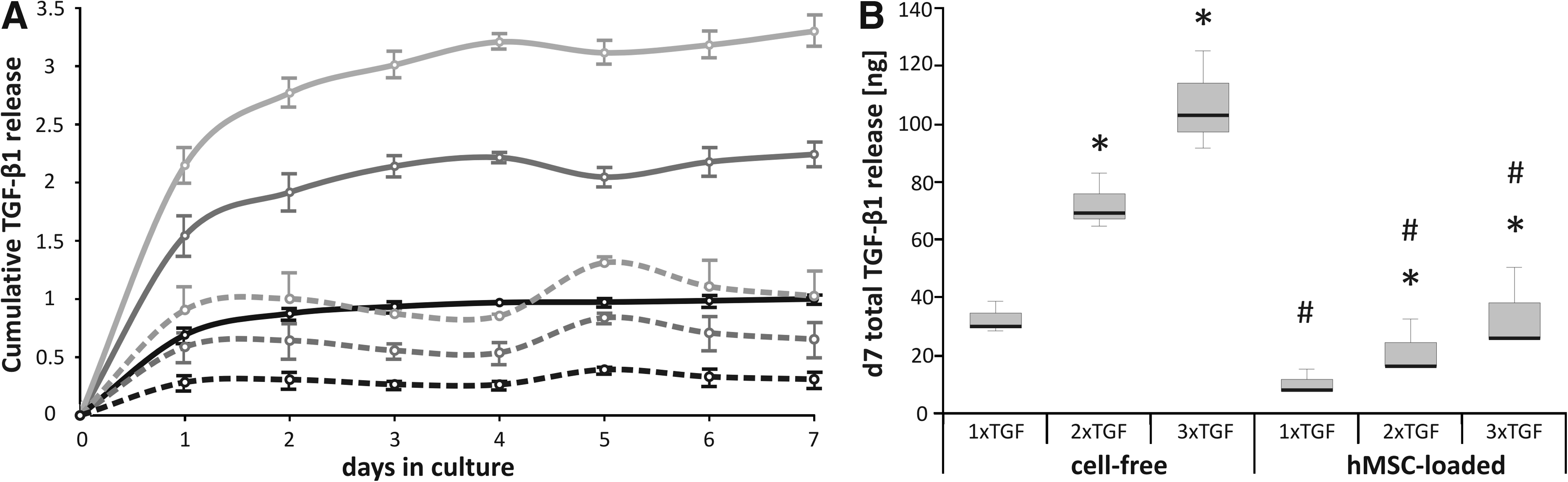
Reduced cumulative TGF-β1 release from hMSC-seeded FG.
Enhanced local TGF-β dosage (n=6 donors) showed surprisingly little effects on differentiation of hMSCs according to histology (Fig. 5A) although one of two suboptimal MSC donor populations, which failed to deposit a cartilaginous matrix with 1× TGF turned positive at 3× TGF (data not shown). All MSC donor populations differentiated successfully in the FG-soluble TGF-β groups, which were prepared with normal, twofold, and threefold higher TGF-β medium concentrations, respectively.
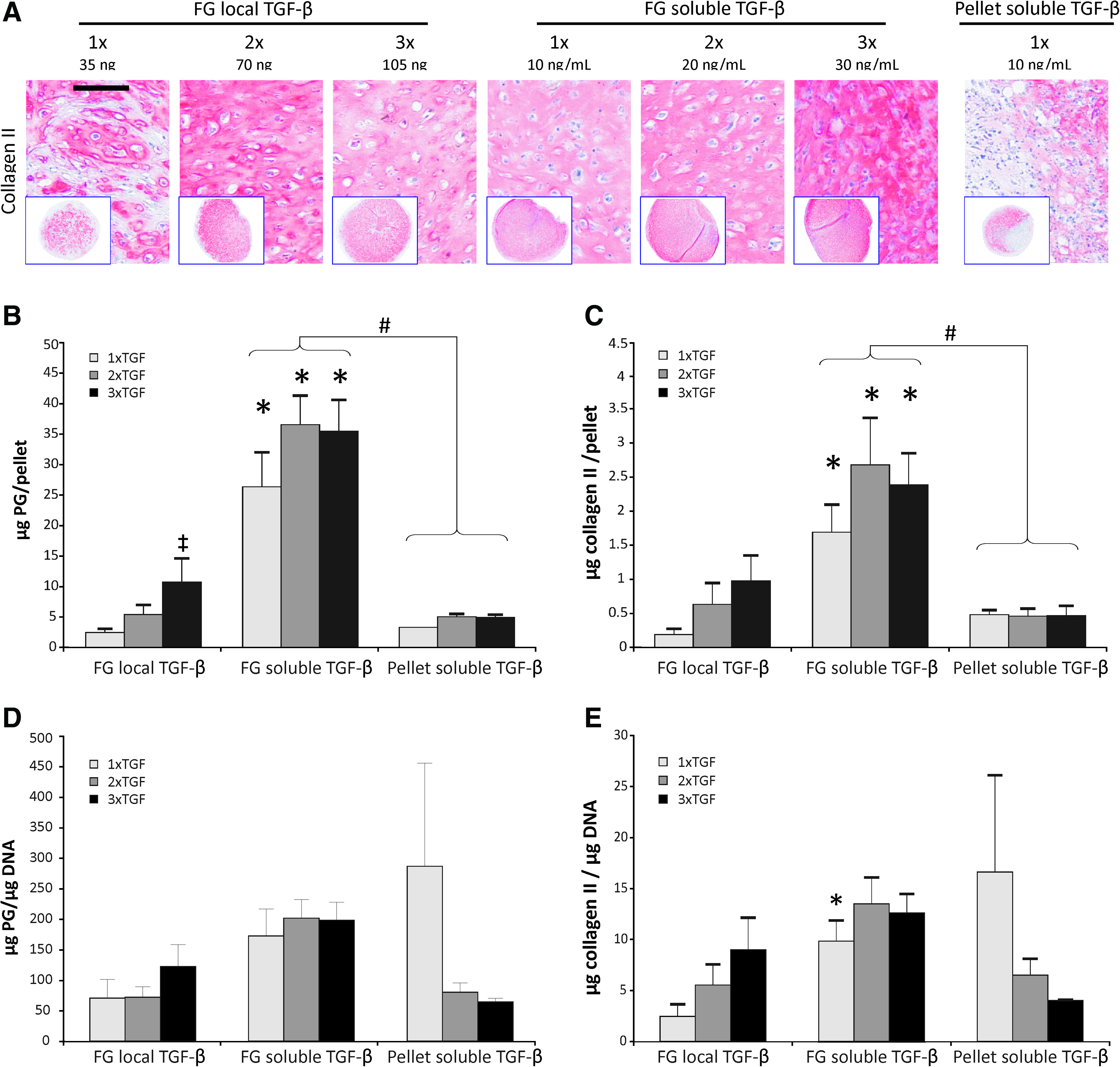
Chondrogenic differentiation under local versus soluble TGF-β.
Biochemical analysis revealed a significantly higher DNA content for FG-soluble TGF-β constructs (181.3 ng) compared to FG-local TGF-β constructs (82.9 ng, 2.2-fold, p=0.015) and pellet controls (99.9 ng, 1.82-fold, p=0.041). This demonstrated that more cells survived chondrogenic differentiation in FG if TGF-β was supplied with each medium exchange with little effect of TGF-β dosage. In line with higher cell numbers, significantly more proteoglycan was deposited per construct in the FG-soluble TGF-β group compared to the biomaterial-free pellet control group (7.5-fold, p=0.002) and the FG-local TGF-β group (6.9-fold, p=0.002) (Fig. 5B), respectively. Similar results were obtained for collagen-type II deposition per construct according to ELISA (Fig. 5C). When the proteoglycan and collagen-type II content were related to the DNA of each construct and thus determined on a per cell basis (Fig. 5D, E), no differences were apparent between groups except for a slightly higher collagen-type II deposition in 1×FG-soluble TGF-β compared to 1×FG-local TGF-β (p=0.015). This demonstrated that rather the number of cells undergoing successful chondrogenesis varied between groups than the average production of cartilage-like matrix molecules per differentiating cell. Interestingly, neither the presence of FG alone nor soluble delivery of TGF-β but only the combination of both showed this enhanced survival effect.
Local TGF-β modulates hypertrophic differentiation
Cartilage repair demands production of collagen-type II-rich constructs with an articular chondrocyte-like phenotype and the absence of hypertrophic markers like collagen-type X and MMP-13 reminiscent for endochondral bone formation. To determine whether the undesired hypertrophy of MSCs in standard chondrogenic culture is altered by local supply of TGF-β, the collagen-type X content of samples relative to their collagen-type II content was determined by Western blot analysis. Interestingly, local TGF-β supply reduced the relative collagen-type X levels for all four donors (Fig. 6A). In line with this, MMP-13 was not induced in the FG-local TGF-β group (0.60±0.21 ng/mL) in contrast to the FG-soluble TGF-β group (72.6±31.8 ng/mL; p=0.029) (Fig. 6B). This suggested that the route of chondrogenesis was altered when the same amount of TGF-β was supplied locally via FG instead of soluble at maintained levels over 6 weeks.
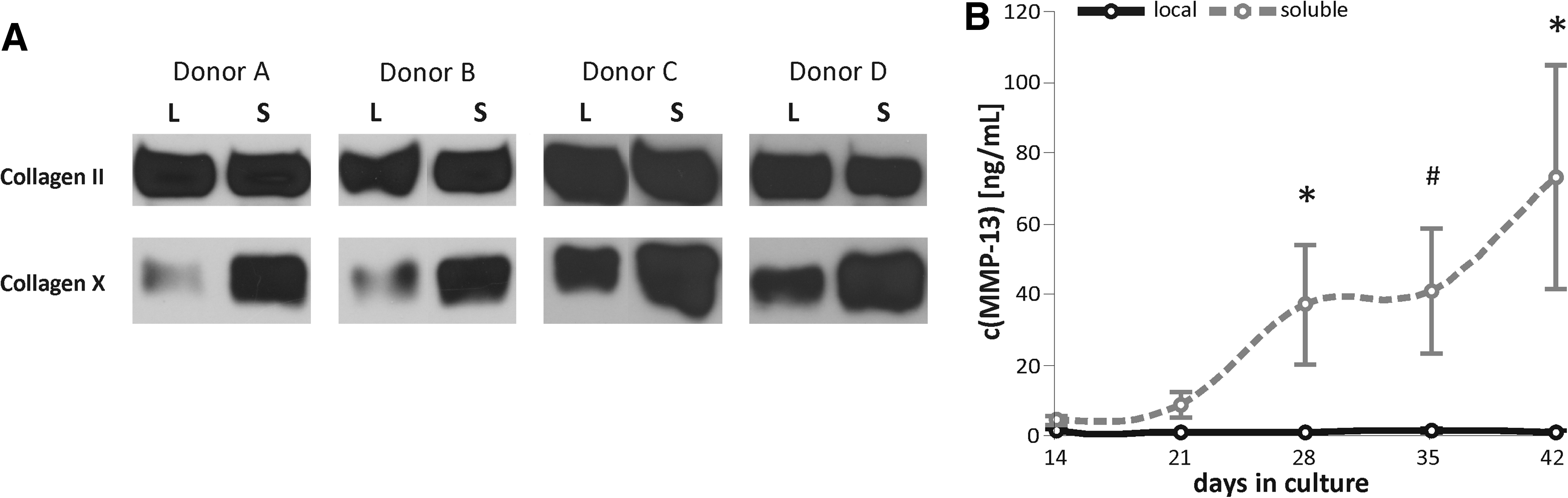
Modulation of hypertrophy by local versus soluble TGF-β supply.
Effect of declining TGF-β supply during chondrogenesis
To determine whether the main effect of local growth factor supply is a lack of TGF-β in the later phase of differentiation culture, hMSC chondrogenesis was compared with an additional group receiving TGF-β transiently. A supply with 30 ng/mL for 2 weeks was followed by a gradual dose reduction with every medium exchange (20 ng/mL–10 ng/mL–5 ng/mL) over 1 week and a subsequent TGF-β-free culture for the remaining 3 weeks.
Like the 3×FG-local TGF-β and the 3×FG-soluble TGF-β constructs, all samples exposed to declining TGF-β (n=9 per group from three donors) deposited a proteoglycan- and collagen-type II-rich ECM (Fig. 7A). The transient and the continuous soluble groups had a higher DNA content compared to local TGF-β supply, which resulted in higher proteoglycan deposition per construct (transient soluble 1.7-fold, p=0.023 and continuous soluble TGF-β 1.8-fold, p=0.019) (Fig. 7B). A similar trend was seen for collagen-type II content (Fig. 7C). Again, no differences in proteoglycan or collagen-type II deposition were evident between groups on a per cell basis (data not shown).
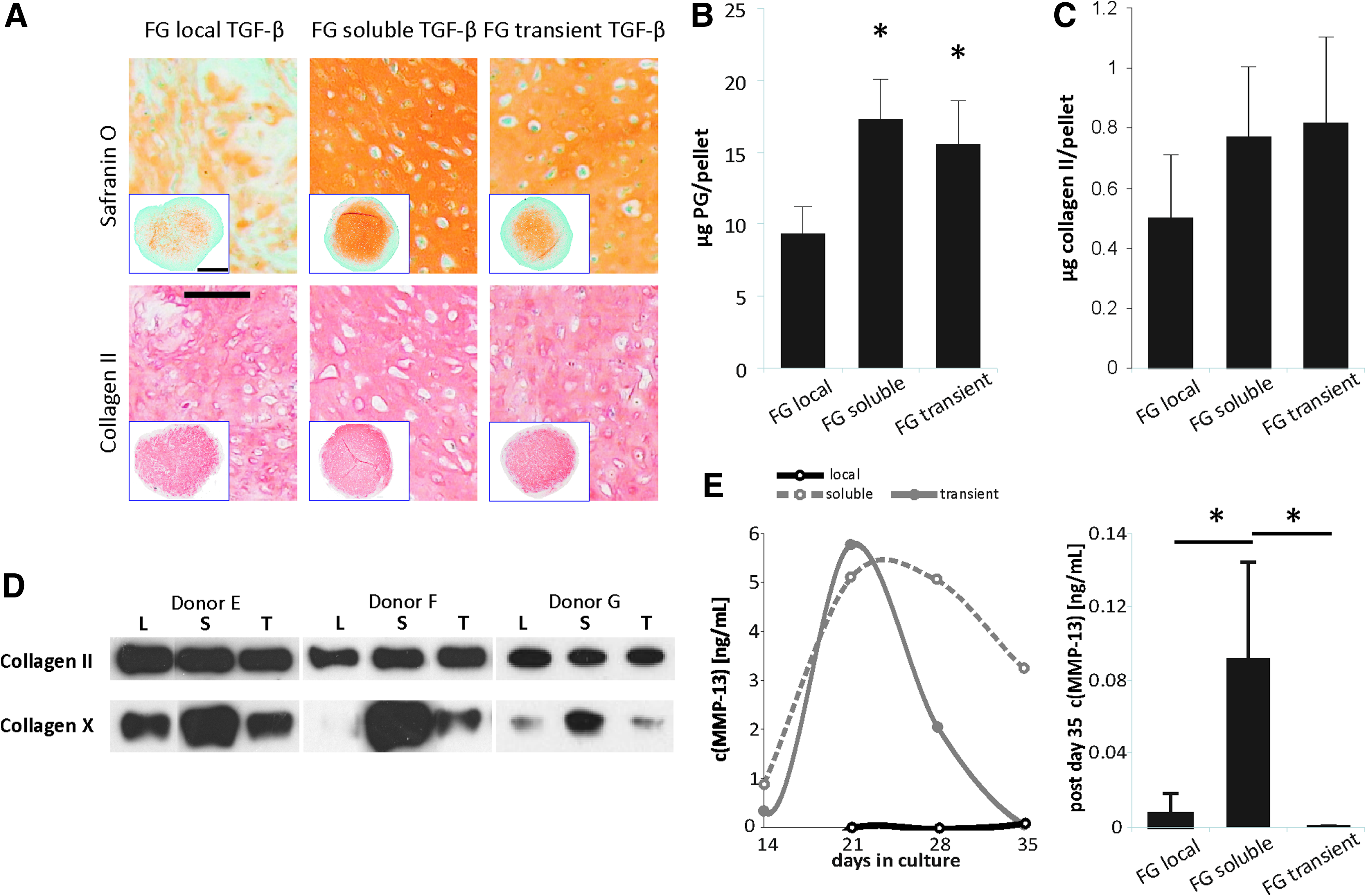
Chondrogenic differentiation under declining TGF-β supply. MSCs from three donors were exposed to chondrogenesis for 6 weeks under local TGF-β (105 ng/construct), constant soluble TGF-β (30 ng/mL), or declining soluble TGF-β supply (30 ng/mL for 2 weeks, followed by a week of gradual dose reduction with every medium exchange: 20 ng/mL–10 ng/mL–5 ng/mL and subsequent TGF-β-free culture for the remaining 3 weeks).
According to collagen-type X deposition in the constructs (Fig. 7D) and MMP-13 secretion into the culture supernatant (Fig. 7E), transient soluble TGF-β supply followed a molecular path of chondrogenesis alike the local TGF-β group. This suggests that the presence of soluble TGF-β within the last 3 weeks of culture stimulates the undesired collagen-type X and MMP13 production. In summary, declining soluble TGF-β supply thus reproduced only part of the features of the local TGF-β group, suggesting that the effects of local TGF-β may not be reduced only to its release kinetics.
Discussion
With the purpose to improve tissue regeneration strategies, innovative and sophisticated systems like liposomes 19 , nano-/microparticels,25–27 covalent conjugation,28,29 or indirect binding via short peptide sequences12,30 have been developed to achieve optimized growth factor release from synthetic biomaterials.36–38 To estimate the value of such advanced approaches compared to technically simpler systems, more knowledge on the limits of natural ECM-derived biomaterials augmented with recombinant growth factor protein is desired. This in vitro study demonstrated that the mere loading of a fibrin hydrogel, or a clinically used Col-I/III carrier, with TGF-β protein allows to drive full long-term in vitro chondrogenesis of hMSCs without impairing the amount of proteoglycan- and collagen-type II-deposition compared to a biomaterial-free standard pellet culture. Nor did local TGF-β supply reduce the deposition rate of cartilaginous matrix in fibrin hydrogel on a per cell basis. Local TGF-β, however, altered the molecular path of chondrogenesis, allowing less cell content and a reduced up-regulation of undesired collagen-type X and MMP-13 compared to a soluble TGF-β approach. Altered TGF-β release kinetics reproduced only part of these features, which suggests, in line with previous reports, that ECM-bound growth factors act distinct from their soluble counterparts. Induction of prolonged receptor activation, 39 facilitation of growth factor receptor integrin crosstalk, 36 and establishment of concentration gradients are suggested mechanisms making natural ECM-derived biomaterials loaded with growth factors especially attractive for tissue engineering approaches.
FG is most promising for tissue engineering applications as vehicle for growth factors20,28,29; a major constraint, however, is the rapid release of entrapped small-sized molecules that are not biochemically bound to the hydrogel components and are thus allowed to diffuse freely within the hydrogel network. In an earlier study we demonstrated that TGF-β-augmented bicomposites composed of FG and Col-I/III were able to drive chondrogenic differentiation of hMSCs. 31 In the current study, we now show that fibrin glue and Col-I/III on their own, respectively, are both able to retain TGF-β and supply it in bioactive form to cells driving their full chondrogenesis. Remarkably, despite high release rates, fibrin augmented with TGF-β provided sufficient support to achieve an extent of chondrogenesis like in standard carrier-free micromass culture with soluble TGF-β supply. Minimal release of TGF-β from Col-I/III carriers compared to 50%–100% release of TGF-β from cell-free FG within 7 days, however, suggests that Col-I/III holds the better retention capacity. Interestingly, the bicomposite behaved like the Col-I/III carrier alone, implicating that the collagen carrier may capture the growth factor diffusing from the FG.
Importantly, neither fibrin nor Col-I/III is so far known to possess defined binding sites for TGF-β. However, a binding affinity of TGF-β to fibrinogen was suggested by Catelas et al. 17 and binding of TGF-β to collagen via electrostatic interactions is likely. Since not native collagen molecules but processed Col-I/III fibers composed the scaffolds of this study, the observed interaction might also be influenced by the material fabrication procedure. Studies on additional collagen carriers are demanded to establish the superior TGF-β storage by collagen carriers compared to fibrin as a general phenomenon.
This is to our knowledge the first study analyzing the effect of cell-embedding on release rates of local growth factors from a hydrogel. While in the absence of cells up to 99% of initially entrapped growth factor was released, only about 31% of input TGF-β was recovered from cell-seeded FG, independent of TGF-β dosage. Remarkably, in cell-seeded constructs, release was marginal beyond 24 h. This suggests that after an initial adaptation phase, cells start to metabolize or degrade the growth factor, or synthesize ECM molecules, which bind and store the local TGF-β, and thus reduce total growth factor release resulting in altered release kinetics. That the growth factor must still be available to the cells beyond the 24-h time point, however, can be concluded from our observation that a 24-h pulse of soluble TGF-β was not sufficient to induce chondrogenesis.
In line with Catelas et al., 17 enhanced TGF-β loading had no influence on total cumulative release rates and release kinetics in our study. Surprisingly, enhanced soluble or local TGF-β dosage had little effect on chondrogenesis according to proteoglycan and collagen-type II deposition (Fig. 5). Earlier observations on adipose tissue-derived MSCs showed that up to fivefold increased TGF-β concentrations did not improve the outcome of chondrogenesis. 40 This demonstrates that the total amount of local TGF-β was not rate-limiting for chondrogenesis at the observed release kinetics.
Chondrogenesis of hMSCs is an attractive well-established in vitro model in which TGF-β action could so far not be replaced by any other signaling factor. Hydrogels and scaffolds composed of structural ECM molecules are known to increase cartilaginous matrix deposition under soluble TGF-β supply.31,41 The current data show that one key function of matrix-enhanced chondrogenesis may be to support cell survival during chondrogenesis. Obviously, biomaterial-free pellet culture under soluble TGF-β supply, like FG with local TGF-β provided a less supportive microenvironment during the demanding reprogramming of cells from a fast-proliferating entity in serum-rich two-dimensional culture to a matrix producing cell in high density three-dimensional culture with moderate hypoxia and no serum, during which up to 60% of cells are lost. Neither early TGF-β levels nor a lack of TGF-β from 3 weeks on seemed responsible for the lower DNA content since the transient TGF-β FG group had similar cell content as the continuous soluble TGF-β group. Local TGF-β may not stimulate each and every cell in sufficient quantity to allow survival or an abrupt shift to high TGF-β-fibrin concentrations may represent an additional stress for the cells. Alternatively, the pulsed support with fresh TGF-β three times a week may add an additional quality to soluble FG groups, which cannot be reached even with higher TGF-β doses given only once. Opposite action of continuous versus pulsed application has long been established for the soluble signaling molecule parathyroid hormone (PTH).42,43 The design of biomaterials allowing pulsed release of a biological factor is, however, still awaited as a further step to elaborate growth factor delivery.
One main finding of our study is that local TGF-β supply did not affect the quantity of cartilaginous matrix deposition per cell but influenced proceeding of chondrogenesis into the late stage of differentiation and hypertrophy. So far, there was no evidence in the literature that relative collagen-type X levels remain lower and MMP-13 is not induced when TGF-β treatment is discontinued after the first 2–3 weeks of chondrogenesis. These results might be interpreted such that although the discontinued TGF-β supply was sufficient to initiate chondrogenesis per se, it was insufficient later on to keep matrix production high, and failed to guide terminal differentiation including hypertrophy. This would mean that the phenotype under local supply simply represented a less advanced chondrogenic differentiation. Since hypertrophy is only the first step in endochondral ossification and only matrix calcification is a major restrain for MSC constructs from being clinically useful for cartilage regeneration, these results might be tempting to accept slightly decreased matrix deposition under local TGF-β supply, to keep hypertrophy low. However, we also observed increased alkaline phosphatase (ALP) activity in the medium of the local TGF-β group during the last weeks of differentiation (data not shown). ALP is strongly connected with matrix mineralization,44,45 is known to be induced during terminal chondrogenic differentiation of MSCs, 46 and influenced by TGF-β. Therefore, chondrogenesis under local TGF-β supply cannot simply have been less advanced, but it must rather be concluded that local TGF-β supply altered the molecular path of chondrogenic differentiation, an interesting aspect that is currently followed further on.
How long or short treatment with TGF-β can be to still sufficiently induce hMSC in vitro chondrogenesis is not well covered by the literature. From our experiments we feel that this time frame depends on the donor MSC and the biomaterial chosen to support the culture. Extended experimentation is required to further address this issue to decide whether, in view of the strong retention of TGF-β in the presence of cells, a controlled release or cell-guided covalent linkage systems 47 is of further value for chondrogenesis.
In summary, ECM components from natural sources like fibrin and Col-I/III are highly attractive as bioactive structures displaying inherent binding properties for TGF-β especially when cells are present. In vitro, current limits of this technically simple approach are only a slightly lower cell and cartilaginous matrix content per construct compared to fresh soluble TGF-β supply. In view of the altered molecular path of locally driven chondrogenesis, it is tempting to speculate that TGF-β, when bound to ECM, may act distinct from its soluble counterpart and further studies are warranted to unravel this in more detail.
Footnotes
Acknowledgment
We thank Simone Gantz for statistical advice, Birgit Frey for histological assistance, and Katja Martin for providing Col-I/III matrix. This work was supported in part by DFG Grant Ri707/7-1, the German Ministry of Education and Research (Grants 0313.755 and 0315.579), and the Orthopaedic University Hospital Heidelberg, Germany.
Disclosure Statement
No competing financial interests exist.