
Abstract
We have shown that human mesenchymal stem cells (hMSCs) have the potential to differentiate into bone when seeded within three-dimensional (3-D) fibrin constructs. Proteins endogenous to the fibrin construct and those secreted by cells in the 3-D constructs provide cues that can promote differentiation of hMSCs along with mechanical support for cell growth and migration. In this study, we decided to take a step back and examine the effect different extracellular matrix (ECM) proteins—fibrinogen, fibronectin, and collagen type I—had on hMSC osteogenic differentiation on two-dimensional (2-D) monolayer cultures. Briefly, 24-well tissue culture plates pre-coated with either fibrinogen (10 mg/mL), fibronectin (10 μg/mL), or collagen type I (1 mg/mL) were seeded with 25,000 cells/well and cultured in normal growth medium or in osteogenic induction medium. At days 1, 7, 14, 21, and 30, cultures were assessed for cell growth using alamarBlue® and osteogenic indicators using alkaline phosphatase and Von Kossa staining. The results show that collagen type I stained positive for calcium deposition the greatest by day 30 in both osteogenic medium and standard culture medium. However, fibrinogen had the greatest staining in osteogenic medium at day 21 and fibronectin was the only substrate to promote calcium deposition in standard culture medium at day 21. These results suggest that the osteogenic differentiation of hMSCs is influenced by both culturing conditions and substrate and that together they have a synergistic effect. By knowing the effect ECM proteins in 3-D fibrin construct have on promoting osteogenic differentiation of hMSCs, the fabrication of complex, biomimetic models designed to manipulate hMSC differentiation toward an osteoblastic lineage will be improved.
Introduction
Bone marrow-derived human mesenchymal stem cells (hMSCs) have the potential to differentiate into a wide variety of cell lineages, including bone, cartilage, fat, tendon, muscle, and nervous tissue. 7 Additionally, they are a source of self-renewing cells, thereby making them an attractive prospect for treating damaged tissue. However, there still exist multiple challenges that limit clinical application including directing lineage-specific differentiation. As such, a variety of synthetic polymers and biopolymers have been explored as substrate materials for maintaining and directing stem cell differentiation. 8 Previous work from this lab has found that hMSCs behave differently when cultured in 3-D fibrin constructs. Specifically, the proliferation rates vary with fibrin concentration, and lower fibrinogen concentrations result in higher hMSC proliferation rates. 9 However, it was also observed that hMSCs undergo osteogenic differentiation when seeded in fibrin containing high fibrinogen complex concentration. 10 To better understand the role proteins endogenous to the 3-D fibrin constructs had in the observed results, we decided to take a step back and examine the contribution of each component in a 2-D environment to gain an understanding of the role selected proteins play in directing the osteogenic differentiation of hMSCs.
The aim of this study is to examine the cell growth and osteogenic differentiation of 2-D monolayer cultures of hMSCs on collagen type I, fibronectin, and fibrinogen. Each of these extracellular matrix (ECM) proteins are commonly found in the 3-D fibrin constructs fabricated in our lab—fibrinogen as a precursor to fibrin, and collagen type I and fibronectin as ECM proteins excreted by the cells seeded within the 3-D fibrin construct. We precoated the culture surface with adsorbed proteins and examined their differentiation and cell growth at days 1, 7, 14, 21, and 30 to examine the effect the ECM proteins had on cell behavior. The fabrication of future biomimetic scaffolds for bone tissue engineering will be aided by identifying the effect potential scaffold materials have on promoting osteogenic differentiation of hMSCs.
Materials and Methods
Cell culture and experimental setup
hMSCs (Poietics™; Lonza Group Ltd., Basel, Switzerland) were cultured at 37°C in 5% carbon dioxide (CO2) up to passage 7. Human foreskin fibroblasts (HFFs) [American Type Culture Collection (ATCC), Manassas, VA] were used as negative control and MC3T3-E1 Subclone 14 Pre-osteoblasts (ATCC) were used as positive control for Von Kossa staining.
Twenty-four-well tissue culture plates were precoated with one of the following proteins: fibrinogen, fibronectin, or collagen type I. The fibrinogen complex (or sealer protein) component of Tisseel™ fibrin sealant kit (Baxter Healthcare Corp., BioScience, Westlake Village, CA) was reconstituted with 1×Tris buffered saline (TBS) and diluted to 10 mg/mL. This fibrinogen complex component contains different ingredients, possibly including some growth factors that could be osteogenic, such as transforming growth factor-b1 and basic fibroblast growth factor. Bovine collagen type I (PureCol®; Advanced BioMatrix Inc., San Diego, CA) was diluted with TBS to 1 mg/mL. Human fibronectin (BD™ Biosciences, Franklin Lakes, NJ) was diluted with TBS to 10 μg/mL. Uncoated polystyrene was used as a negative substrate control. One milliliter of the diluted protein solution was added to the wells of the tissue culture plates and the proteins were allowed to adsorb to the surface for 30 min. Excess protein solution was aspirated off and the protein-coated wells were washed with warm 1×Dulbecco's phosphate-buffered saline (DPBS) twice prior to cell seeding.
At the time of experiment, 2-D monolayer cultures of hMSCs were plated in the precoated 24-well plates at 25,000 cells/well. hMSCs (Poietics; Lonza Group Ltd.) were cultured in either MSC basal medium (10% fetal bovine serum, 2% L-glutamine, and 0.1% GA-1000) for standard growth medium or in AdvanceSTEM osteogenic differentiation medium (HyClone™; Thermo Fisher Scientific, Inc., Waltham, MA). The growth medium for HFFs (Cellgro®; Mediatech Inc., Manassas, VA) was Dulbecco's modified Eagle's medium (10% fetal bovine serum and 1% antibiotic/antimycotic) and α-minimum essential medium (10% fetal bovine serum and 1% antibiotic/antimycotic) for MC3T3-E1 (Cellgro; Mediatech Inc.). All culture plates were placed at 37°C in 5% CO2 for up to 30 days. Culture medium was replaced every 2–3 days and cell growth and osteogenic differentiation were analyzed at days 1, 7, 14, 21, and 30.
Cell growth
hMSC growth analysis was conducted in parallel with alkaline phosphatase (ALP) analysis. Two-dimensional monolayers were washed with warm DPBS twice. A 10% solution of alamarBlue® (Invitrogen Corp., Carlsbad, CA) in culture medium was added to 2-D monolayer cultures and incubated at 37°C in 5% CO2 for 3 h. Cell growth was monitored by measuring fluorescent intensity of the culture medium with a multiwell plate reader (Infinite® F200; Tecan Group Ltd., Männedorf, Switzerland) at 3 h.
Cell differentiation
Osteogenic differentiation of hMSCs was analyzed using a standard ALP colorimetric assay (Abcam®, Cambridge, MA) as an early maker for osteogenic differentiation and Von Kossa staining (American MasterTech Scientific, Inc., Lodi, CA) for mineral deposition as a late marker for osteogenic differentiation.
ALP assay
Two-dimensional monolayer cultures were washed with warm DPBS twice. The final wash was with 1×assay buffer. 1×assay buffer with 1 mg/mL of bovine serum albumin and Triton X-100 was added to the cultures. A cell scraper was used to free the cells from the well surface. The resulting cell suspension was incubated at 4°C for 10 min with vigorous shaking. Cells were then centrifuged for 10 min at 4°C and 50 μL of the supernatant was added to a 96-well plate and incubated with 50 μL of p-nitrophenol phosphate stock substrate solution at 37°C. The absorbance of p-nitrophenol product formed at 405 nm, which is proportional to the ALP activity, was read at 60 min according to the assay manufacturer's protocol using a multiwell plate reader.
Von Kossa staining
On days 21 and 30, the culture medium was removed from each 2-D monolayer culture. Each well was washed twice with DPBS and then fixed with 10% formalin at room temperature for 30 min and placed at 4°C for later staining. At the time of staining, each well was washed thrice with distilled water. 5% silver nitrate was added for 1 h under UV light. Wells were washed again thrice with distilled water and then 5% thiosulfate was added for 5 min. Each well was washed three additional times with distilled water and counterstained with nuclear fast read for 10 min. Each well was washed five more times with distilled water and photographed with a standard inverted light microscope.
Results
hMSCs were grown as 2-D monolayer cultures on various 2-D substrates. The concentration of collagen type I was 1 mg/mL, fibronectin was 10 μg/mL, and fibrinogen was 10 mg/mL. The concentrations of collagen and fibronectin were selected according to the manufacturer's recommendation for coating 2-D tissue culture plates. Fibrinogen concentration was selected based on the concentration used to fabricate 3-D fibrin constructs in our lab.
Cell growth of 2-D monolayer cultures of hMSCs on different substrates
Analysis of cell growth by alamarBlue showed that the hMSCs were viable and proliferated. Cell growth increased with incubation time for hMSCs cultured on all the 2-D substrates in both standard culture medium and osteogenic medium, as shown by the increase in fluorescent intensity (Fig. 1). Additionally, each culture reached confluence by day 14.
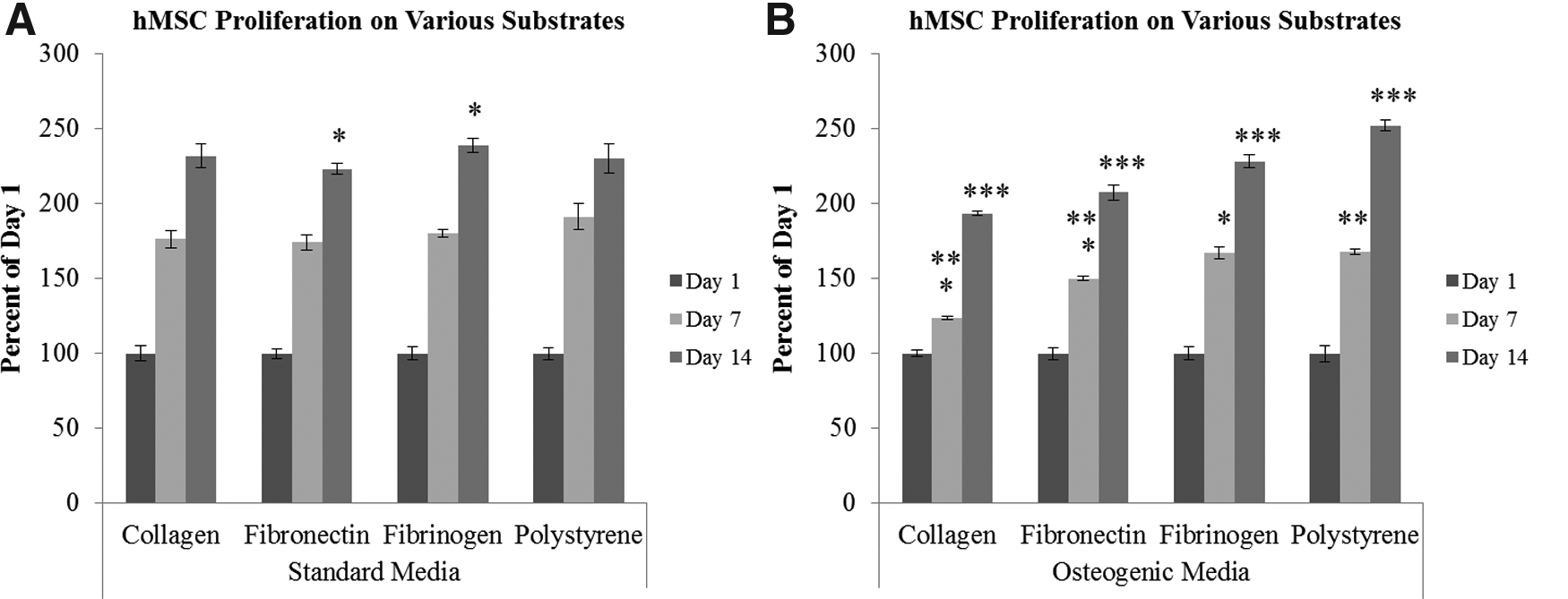
Comparing the cell growth of human mesenchymal stem cells (hMSCs) cultured on two-dimensional (2-D) substrates at days 1, 7, and 14 in standard culture medium versus osteogenic culture medium measured by alamarBlue (n=3). About 25,000 hMSCs were seeded on 24-well plates coated with either collagen type I (1 mg/mL), fibronectin (10 μg/mL), or fibrinogen (10 mg/mL). Uncoated well plates (polystyrene) served as control. The effect of the 2-D substrate on cell growth is compared in standard culture medium
Collagen type I
hMSC growth on collagen type I was greatest when cultured in standard medium compared to osteogenic medium with a statistically significant difference in cell growth at both days 7 and 14 (data not shown). Between days 1, 7, and 14, both standard and osteogenic medium saw statistically significant increases in cell growth with time (data not shown). In osteogenic medium there was a 1.25-fold increase in cell growth at day 7 from day 1 and a greater increase in cell growth between days 7 and 14 with a 1.5-fold increase. In total, there was nearly a 2-fold increase in hMSC growth cultured on collagen type I in osteogenic medium. Standard medium increased hMSC growth 1.75-fold from days 1 to 7 when cultured on collagen type I. There was a 1.3-fold increase in cell growth from days 7 to 14 in standard medium with a total 2.3-fold increase in hMSC growth when cultured on collagen type I in standard medium.
Fibronectin
Standard culture medium had the greatest cell growth of hMSCs cultured on fibronectin compared with osteogenic medium although only day 7 had a statistically significant difference (data not shown). Between days 1, 7, and 14, both standard and osteogenic medium saw statistically significant increases in cell growth with time (data not shown). There was a 1.5-fold increase in cell growth at day 7 from day 1 in osteogenic medium and a 1.4-fold increase from days 7 to 14. In total, there was a 2-fold increase in hMSC growth cultured on fibronectin in osteogenic medium. Standard medium increased hMSC growth 1.75-fold from days 1 to 7 when cultured on fibronectin. There was a 1.25-fold increase in cell growth from days 7 to 14 in standard medium with a total 2.2-fold increase in hMSC growth when cultured on fibronectin in standard medium.
Fibrinogen
hMSC growth on fibrinogen was statistically greater in standard medium compared with osteogenic medium (data not shown), and, between days 1, 7, and 14, both standard medium and osteogenic medium experienced statistically greater increases in cell growth (data not shown). hMSCs cultured in osteogenic medium experienced a 1.7-fold increase in cell growth from days 1 to 7 and a 1.4-fold increase from days 7 to 14. There was a total 2.3-fold increase from days 1 to 14 for hMSCs cultured on fibrinogen in osteogenic medium. hMSC growth in standard medium experienced a 1.8-fold increase from days 1 to 7 and a 1.3-fold increase from days 7 to 14. In total, there was a 2.4-fold increase in hMSC growth in standard medium cultured on fibrinogen.
Polystyrene
hMSC growth was greatest in standard medium at day 7 but greatest in osteogenic medium at day 14. However, only the difference at day 7 between culture mediums was statistically significant (data not shown). Between days 1, 7, and 14, both osteogenic and standard medium experienced statistically significant increases in cell growth with time (data not shown). The final change in cell growth from days 1 to 14 was a 2.5-fold increase in osteogenic medium and 2.3-fold increase in standard medium. There was a 1.7-fold increase in cell growth from days 1 to 7 and 1.5-fold from days 7 to 14 in osteogenic medium. The standard medium experienced a 1.9-fold increase from days 1 to 7 and a 1.2-fold increase from days 7 to 14.
Comparing substrates
In standard medium, fibrinogen had the greatest cell growth of the substrates examined; however, only at day 14 was the difference statistically significant between fibrinogen and fibronectin (Fig. 1A). In osteogenic medium, fibrinogen had the greatest cell growth at days 7 and 14 when compared with the other two substrates and collagen type I had the least. The difference between the substrates was statistically significant for all time points except for the difference between fibrinogen and polystyrene at day 7 (Fig. 1B).
Early osteogenic activity of 2-D monolayer cultures of hMSCs on different substrates
Collagen type I
hMSCs cultured on collagen type I in standard medium experienced a decreasing trend in ALP activity from days 1 to 14. This difference, however, was not statistically significant (data not shown). hMSCs cultured on collagen type I in osteogenic medium experienced a 19-fold increase in ALP activity from days 1 to 14. There was a 4.7-fold increase from days 1 to 7 and a 4-fold increase from days 7 to 14. Between days 1, 7, and 14, hMSCs experienced statistically significant increases in ALP activity with time (data not shown).
Fibronectin
hMSCs cultured on fibronectin in standard medium experienced a slight decreasing trend in ALP activity from days 1 to 14. This difference was not statistically significant (data not shown). There was a 38-fold increase in ALP activity from days 1 to 14 for hMSCs cultured on fibronectin when cultured in osteogenic medium. From days 1 to 7 there was a 6.5-fold increase in ALP activity and from days 7 to 14 there was a 5.8-fold increase. Between days 1, 7, and 14, hMSCs experienced statistically significant increases in ALP activity with time (data not shown).
Fibrinogen
hMSCs cultured on fibrinogen in standard medium experienced a slight decline in ALP activity from days 1 to 7. This difference was not statistically significant. There was a significant 1.8-fold increase in the ALP activity of hMSCs cultured on fibrinogen from days 7 to 14 (data not shown). hMSCs cultured on fibrinogen in osteogenic medium experienced a 22-fold increase in ALP activity from days 1 to 14. From days 1 to 7 there was a 5.7-fold increase and from days 7 to 14 there was a 3.9-fold increase in ALP activity. Between days 1, 7, and 14, hMSCs experienced statistically significant increases in ALP activity with time (data not shown).
Polystyrene
hMSCs cultured on polystyrene in standard medium experienced an increasing trend in ALP activity from days 1 to 7 and then a slight decrease from days 7 to 14. The change in ALP activity, however, was not statistically difference between any of the time points (data not shown). There was a 53-fold increase in ALP activity from days 1 to 14 for hMSCs cultured on polystyrene when grown in osteogenic medium. From days 1 to 7 there was a 6.6-fold increase in ALP activity and from days 7 to 14 there was an 8-fold increase. Between days 1, 7, and 14, hMSCs experienced statistically significant increases in ALP activity with time (data not shown).
Comparing substrates
ALP activity was normalized to cell growth to compare the ALP activity of hMSCs grown on different 2-D substrates. In standard medium, fibrinogen had the greatest ALP activity at day 14; however, this was only statistically greater than ALP activity of hMSCs cultured on collagen type I. At day 7, hMSCs cultured on polystyrene had significantly greater ALP expression than hMSCs grown on fibrinogen and collagen type I (Fig. 2A). In osteogenic medium, fibronectin had the greatest ALP activity compared with the other substrate proteins at day 14. This difference was statistically different. There was no statistically significant difference between collagen and fibrinogen at day 14. Additionally, there was no significant difference in ALP activity at day 7 between any of substrates (Fig. 2B).
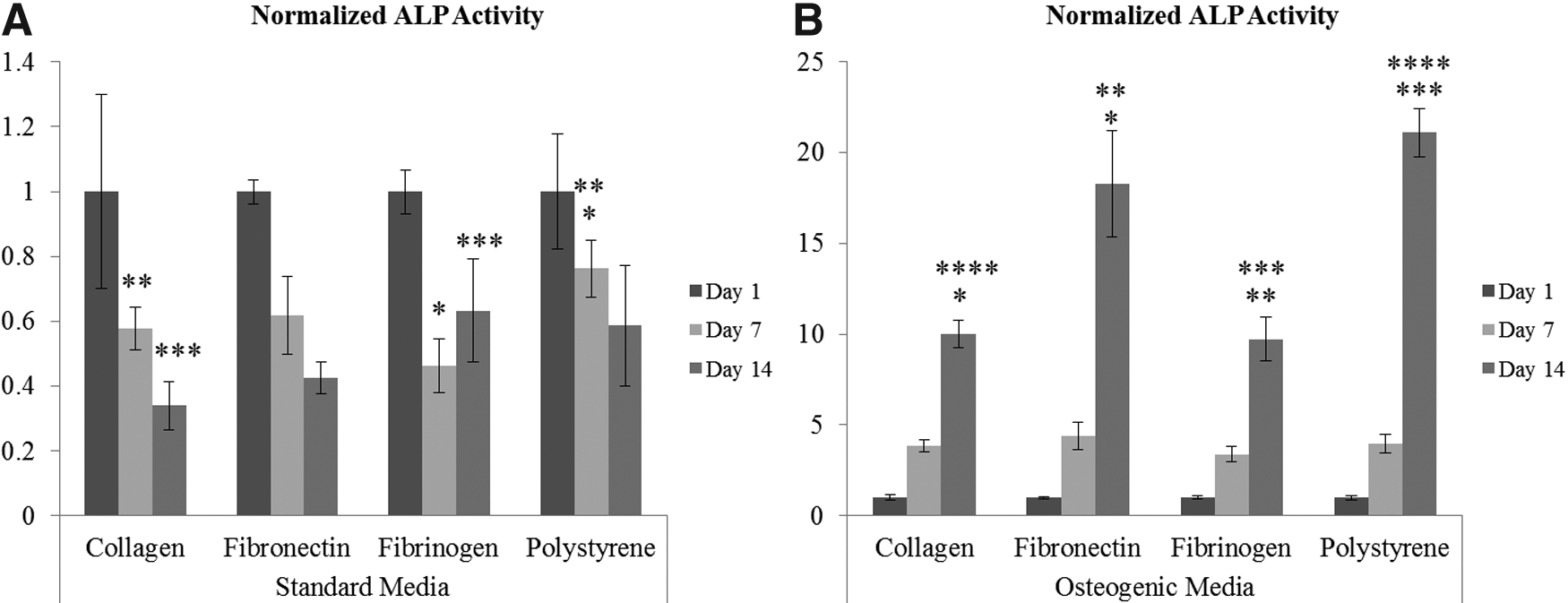
Comparing the effect of the 2-D substrate on alkaline phosphatase (ALP) activity of hMSCs at days 1, 7, and 14 in
Late osteogenic activity of 2-D monolayer cultures of hMSCs on different substrates
Von Kossa staining qualitatively measured late osteogenic activity of hMSCs cultured on all the 2-D substrates in both standard culture medium and osteogenic medium. HFFs and MC3T3-E1 preosteoblasts cultured on polystyrene in their respective standard growth medium were used as negative and positive controls, respectively (Fig. 3).
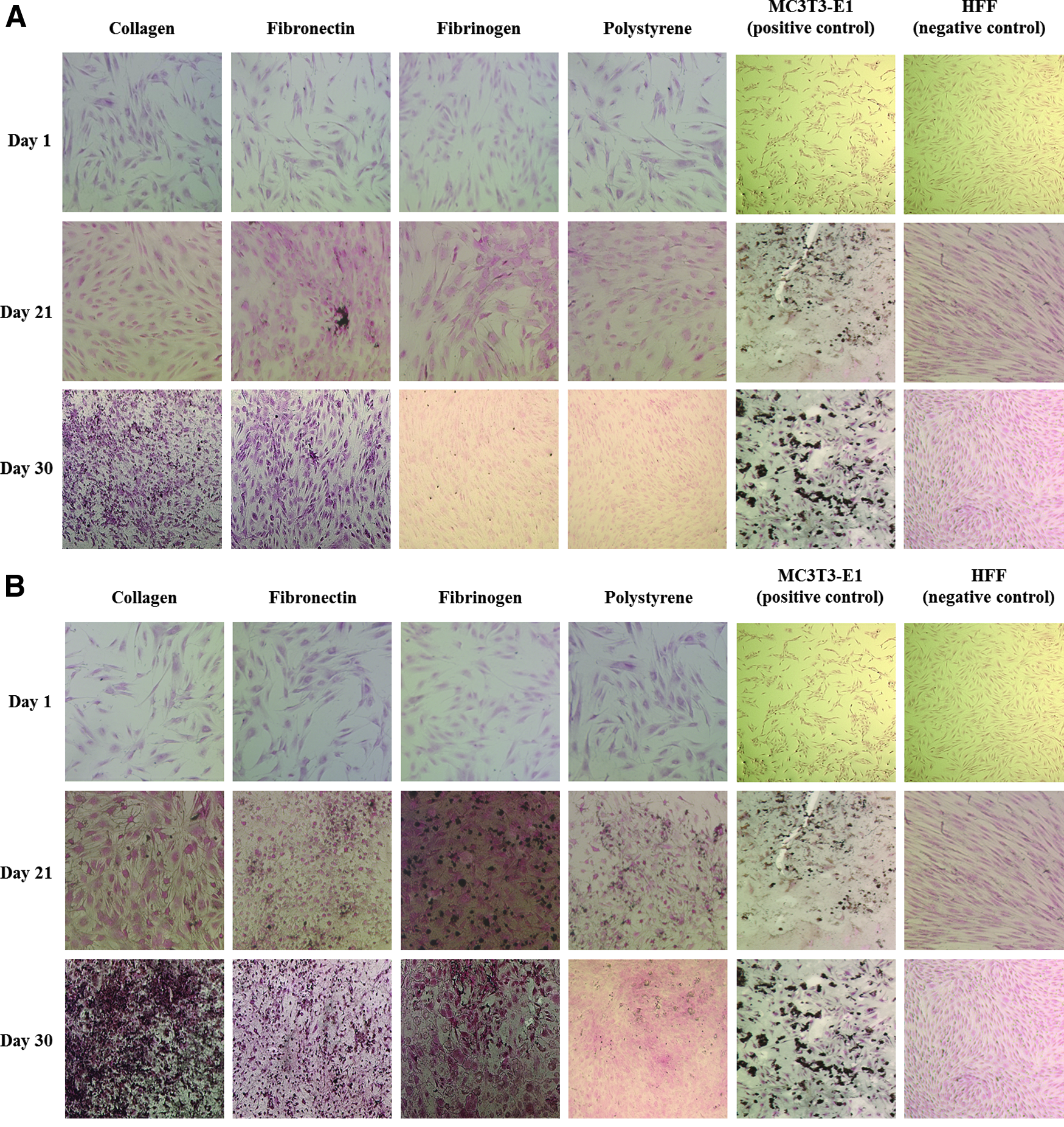
Von Kossa staining of hMSCs cultured on collagen type I (1 mg/mL), fibronectin (10 μg/mL), fibrinogen (10 mg/mL), and uncoated polystyrene at days 1, 21, and 30 in
Collagen type I
hMSCs cultured on collagen type I in standard medium at day 21 did not stain positive for calcium deposition but by day 30 there was positive staining for calcium deposits and it was greater than hMSCs cultured on the other substrates (Fig. 3A). In osteogenic medium, hMSCs cultured on collagen type I stained positive at both days 21 and 30. Calcium deposition was greatest on collagen type I at day 30 when compared to the other substrates examined (Fig. 3B).
Fibronectin
hMSCs cultured on fibronectin in standard medium stained positive for calcium deposition via Von Kossa staining at days 21 and 30. hMSCs cultured on fibronectin were the only samples to stain positive for calcium deposition in standard medium at day 21 (Fig. 3A). In osteogenic medium, hMSCs cultured on fibronectin stained positive for calcium deposition at both days 21 and 30. The extent of positive staining was not as great as collagen type I at day 30 but was greater than fibrinogen at day 30 (Fig. 3B).
Fibrinogen
hMSCs cultured on fibrinogen in standard medium did not stain positive for calcium deposition at day 21 and lightly stained positive at day 30 (Fig. 3A). However, hMSCs cultured on fibrinogen in osteogenic induction medium stained positive for calcium deposition via Von Kossa staining at day 21 to a greater extent than the ECM proteins examined, but by day 30 the extent had not significantly increased as compared with collagen type I and fibronectin (Fig. 3B).
Polystyrene
hMSCs cultured on polystyrene in standard medium did not stain positive for calcium deposition until day 30. In osteogenic medium, hMSCs cultured on polystyrene stained positive for calcium deposition at both days 21 and 30. In both culture conditions, polystyrene exhibited weak staining for calcium deposition when compared with hMSCs grown on the ECM proteins examined (Fig. 3A, B).
Discussion
Three-dimensional scaffolds more closely imitate the structural environment of tissues. As such, there has been a growing body of work that aims to create biomimetic scaffolds that recapitulate the microenvironments of various tissue types. 11 Ultimately, the goal is to fabricate better in vitro models and to apply them clinically. hMSCs have been commonly employed in these models because they are a self-renewing cell population with the capacity to differentiate into multiple cell lineages, including osteoblasts. Accordingly, biomimetic scaffolds have been used to direct the differentiation of hMSCs down the osteoblastic lineage and work has been done to study the osteogenic potential of various scaffold materials.9,10,12 Proteins native to the ECM of bone tissue are attractive materials for this application, either as the 3-D scaffolding material themselves or for the functionalization of synthetic materials with their integrin-binding peptide sequence motifs. 13 While comparative investigations into the effect ECM proteins in a 2-D environment have on hMSCs is not new,13–17 here we presented an extended study (30 days) of hMSCs cultured on collagen type I, fibronectin, and fibrinogen and the osteogenic response due to both cell–scaffold interactions, and culturing conditions and cell–scaffold interactions, summarized in Table 1.
The present study demonstrated that hMSC behavior in 2-D monolayer cultures depended on the substrate and culture medium. Collagen type I, for both culture conditions, had the greatest staining for calcium deposition by day 30 and collagen coupled with osteogenic culture medium exhibited the greatest degree of osteogenic differentiation of the conditions studied. This finding seems fitting as collagen type I is the main organic constituent in healthy bone tissue. 18 Additionally, it has been previously shown that integrin–collagen interaction promotes osteogenic differentiation. MC3T3-E1 preosteoblasts differentiate via mitogen-activated protein kinase signaling pathway initiated by α2β1integrin–collagen interaction, 19 and Ode et al. found hMSCs strongly express α2β1 integrins. 13 Further, endogenous expression of bone morphogenetic proteins (BMPs), specifically BMP-2 and BMP-4, promote osteogenic differentiation of preosteoblasts if the cells have a collagenous ECM. 20
Though not specific for osteogenesis, another marker used to measure osteogenic differentiation of hMSCs in different culture conditions was ALP activity, an intracellular enzyme necessary for mineralization and an early marker for osteogenic differentiation.14,21 hMSCs grown on fibronectin in osteogenic culture medium had the greatest degree of ALP activity at day 14. Additionally, for both culture conditions, fibronectin followed collagen in the most staining for calcium deposition by day 30. hMSC interaction with fibronectin is facilitated by α5β1 integrin.13,22 It has been shown that activation of α5 integrin was sufficient to promote osteogenic differentiation of human mesenchymal stromal cells. 22 Additionally, the β1 integrin subunit has demonstrated a role in the formation of new bone tissue.16,23 While fibronectin has the potential to initiate the differentiation of hMSCs toward an osteoblastic lineage, the results suggest that signaling from fibronectin alone is not sufficient for complete differentiation and that additional signaling from scaffolding materials, and molecular signals will be required.
hMSC growth in osteogenic medium was lower when compared with hMSCs grown in standard culture medium. It has been shown that terminally differentiated cell populations usually do not proliferate or have slowed cell growth whereas stem cells do. 24 Collagen type I in osteogenic medium had significantly lower growth rates at both days 7 and 14 when compared with fibronectin and fibrinogen while fibrinogen substrates had the greatest cell growth in osteogenic culture medium. Consistent with work previously done by our lab and others, the low concentration of fibrinogen complex promoted the proliferation of hMSCs.9,25 Though ALP activity and staining for calcium deposition was lower for fibrinogen than collagen and fibronectin, the results show that fibrinogen was able to initiate differentiation toward an osteoblastic cell lineage. Fibrinogen has an integrin-binding motif for α5β1, 13 and we have previously reported osteogenic differentiation of hMSCs cultured in fibrin constructs fabricated with high concentrations of fibrinogen complex. 10 The results presented here suggest that, similar to fibronectin, additional signaling from different scaffold materials and soluble factors will be required for complete osteogenic differentiation.
While fibrinogen and fibronectin showed less calcium deposits compared with collagen, despite having greater ALP expression, all 2-D substrates may have directed differentiation down a different cell lineage to varying degrees. hMSCs have the potential to differentiate into multiple cell lineages found in bone, smooth muscle, cartilage, tendons, and adipose tissue. 26 Therefore, most hMSC cultures are a heterogeneous population after 14 days of culture. 27 For example, Huang et al. found that fibrin was able to induce mesenchymal stromal cells to express genes associated with vasculogenic, myogenic, neurogenic, and chondrogenic tissues. 25 Differentiation toward a chondrogenic cell lineage, however, could suggest an endochondral bone formation where cartilage is ossified to form bone. 28 Further, it is likely that the hMSC timeline for expression of the early and late osteoblastic differentiation markers is dependent upon the culture conditions, including the substrate. For instance, hMSCs cultured on collagen type I appear to reach terminal differentiation faster than the other substrates, as evident by the high positive staining for calcium deposition coupled with the lower ALP activity in the presence of collagen type I when compared with the other substrates. By contrast, hMSCs cultured on polystyrene fail to stain positive for calcium deposition, indicating no terminal differentiation into osteoblasts, despite the strong ALP activity. This may be due to the cells being blocked at the chondrogenesis step or the preosteoblast differentiation step; however, the expression of cell types other than osteoblasts were not tested in this study and further experiments are needed to evaluate the different phenotypic markers of chondrogenesis, hypertrophy, and osteogenesis.
Previous studies looking at the osteogenic differentiation of hMSCs on these 2-D substrates did not report positive staining for calcium deposition, a late marker for osteogenic differentiation, in contrast with our results. A possible explanation could be differences in the experimental setup, for example, substrate concentration (collagen: 1 mg/mL vs. 20–800 μg/mL13,14,25; fibronectin: 10 μg/mL vs. 20–50 μg/mL,13,14 1 μg/mL 25 ; fibrinogen: 10 mg/mL vs. 22 μg/mL 13 ). Further, previous groups carried their differentiation studies to 3 weeks while our study went to 4 weeks. These findings suggest that the osteogenic effect of the 2-D substrates is both concentration and time dependent.
Our lab has previously studied the behavior of cells, including hMSCs, in 3-D fibrin constructs and both fibronectin and collagen are secreted from hMSCs into the 3-D fibrin constructs. We have previously reported that hMSCs behave differently when cultured in 3-D fibrin constructs and that hMSCs undergo osteogenic differentiation when seeded in fibrin containing high fibrinogen complex concentration. 10 Though not studied here, one possibility based on the results from this study is that high fibrin concentration upregulates collagen production and thereby promotes osteogenic differentiation. Therefore, we anticipate that future 3-D biomimetic scaffolds can be fabricated based on the understanding of how hMSCs grow and differentiate on these 2-D substrates. For example, one of the obstacles facing the therapeutic use of hMSCs is the limited supply and it would be advantageous to fabricate a scaffold that promotes hMSC growth, and osteogenic differentiation. Based on our results, a 3-D fibrin construct fabricated from lower concentrations of fibrinogen complex and mixed with collagen type I could promote both cell growth and osteogenic differentiation. Additionally, the fact that the fibrinogen concentration studied here promoted proliferation and initiated osteogenic differentiation suggests that there may be an optimal fibrinogen concentration that maximizes hMSC growth and osteogenic differentiation. Further, incorporation of osteoinductive growth factors can synergistically enhance the osteogenic response of our 3-D fibrin-based constructs.
In summary, the results presented support previous findings and add to our understanding of the role scaffolding materials play in guiding the osteogenic differentiation of hMSCs while conflicts with results from previous studies may be attributed to differences in experimental setup. These results suggest that the culture medium has a greater influence over the differentiation of hMSCs into osteoblast cells than the substrate alone. However, the combination of substrate with osteogenic culture medium has a synergistic effect and produces a greater osteogenic response of hMSCs than either alone. Additionally, knowing the effect potential scaffold materials have on promoting cell growth and differentiation of hMSCs will aide in the fabrication of complex, biomimetic models designed to manipulate hMSC fate. Specifically, concentrations of collagen type I, fibronectin, and fibrinogen—proteins examined in this study and endogenous to a 3-D fibrin construct—will be optimized for the fabrication of a complex, 3-D biomimetic bone model designed to improve the osteogenic differentiation of hMSCs.
Footnotes
Disclosure Statement
No competing financial interests exist.