
Abstract
One of the most challenging issues in liver tissue engineering is the incorporation of a functional bile duct (BD) network in the hepatic tissue. In this study, we investigated the feasibility of a three-dimensional (3D) coculture of adult rat primary biliary epithelial cells (ABECs) and rat fetal liver cells (FLCs) to form functional BD structures. 3D hetero-spheroids containing various ratios of the two cell types were successfully obtained in low-adherence, round-bottom, 96-well plates. Histological analyses and functional analysis using fluorescein diacetate demonstrated that the ABECs critically contributed to the reconstruction of continuous ductular networks. Metabolized fluorescein was transported toward cystic structures in a time- and ABEC ratio-dependent manner. In particular, the duct-like structures containing fluorescein accumulations interlinked the large cystic accumulations. Furthermore, transplantation experiments demonstrated that ABECs, but not FLCs, enabled the establishment of BD networks in vivo. This study provided important insights into the development of transplantable liver tissues with bile excretion functionality.
Introduction
Despite the increasing evidence that heterotopic transplantation of liver tissue engineered with adult and fetal hepatocytes is therapeutically beneficial,3–6 there are still hurdles to overcome. These hurdles are derived mainly from the difficulty of recapitulating the complex architecture of the liver, including not only the hepatic vasculature but also the bile duct (BD) network. 7 Because bile is toxic to hepatocytes, 8 biliary tract dysfunction could cause hepatocellular necrosis. In the normal liver, bile is synthesized by hepatocytes, secreted into the BDs through the bile canaliculi (BCs), and excreted from the liver into the duodenum. While the BCs are formed by adjacent hepatocytes, the BDs are formed by biliary epithelial cells (BECs). Therefore, the importance of establishing BDs using BECs must be emphasized to engineer a transplantable liver tissue.
Despite some evidence for the possibility of in vitro BD reconstruction, to our knowledge, there are no reports on the incorporation of functional BDs into three-dimensional (3D) hepatic organoids. It has been demonstrated that suspensions of mouse fetal liver cells (FLCs) or rat neonatal liver cells formed hepatic spheroids possessing BD-like luminal structures.9,10 However, these studies did not demonstrate that the BD-like structures could excrete, transport, or store bile. Moreover, primary cultures of human or rat adult BECs (ABECs) grown in the collagen-gel configuration were capable of forming BD-like networks that excreted and stored bile.11,12 This finding is significant, but this insight has not yet been applied to the development of 3D hepatic organoids possessing functional BDs. Improvements in the strategies are thus, necessary to introduce functional BDs into 3D hepatic organoids.
In this study, we investigated the ability of ABECs to form functional BDs in reconstructed hepatic tissues. We generated 3D hepatic hetero-spheroids using suspension cocultures of rat FLCs and rat ABECs. We chose FLCs as the hepatocyte source because we observed that adult hepatocytes did not participate in hetero-spheroid formation in our preliminary study. We determined the functionality of the established BD networks by observing the localization of a fluorescent probe that mimicked bile. Furthermore, transplantation experiments demonstrated that in vitro-generated hetero-spheroids maintained their BD networks in vivo.
Materials and Methods
Isolation of ABECs and FLCs
The E17.5 FLCs and ABECs were isolated from female Wistar rats (Fig. 1). These two types of cells were isolated in parallel from one pregnant rat for each experiment. The ABECs were isolated following the protocol previously described with a minor modification. 12 Briefly, BD trees were isolated from the maternal liver after two-step liver perfusion and cut with scissors into small fragments. The fragments were digested by shaking at 37°C for 30 min in Leibovitz L-15 medium (Invitrogen) supplemented with 400 U/mL of collagenase (Wako), 700 U/mL of hyaluronidase (Sigma-Aldrich), 10−7 M insulin, and 10−7 M dexamethasone (DEX). After filtration with a 40-μm cell strainer (BD Biosciences), the cells were collected by centrifugation at 800 g for 10 min. Then the pellet was resuspended in Dulbecco's modified Eagle's medium (DMEM; Invitrogen) supplemented with 10% fetal bovine serum (FBS; Invitrogen) and centrifuged twice at 800 g for 5 min. Finally, the isolated ABECs were suspended at a concentration of 1×106 cells/mL in ABEC culture medium (DMEM supplemented with Insulin-Transferrin-Selenium supplement (Invitrogen), 10−7 M DEX, 10 mM nicotinamide (Sigma-Aldrich), 1 mM ascorbic acid (Wako), 10 ng/mL epidermal growth factor (EGF; Sigma-Aldrich), 10 ng/mL of hepatocyte growth factor (HGF; Peprotech), 10% FBS, and antibiotic/antimycotic solution (100 U/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B) (Invitrogen).
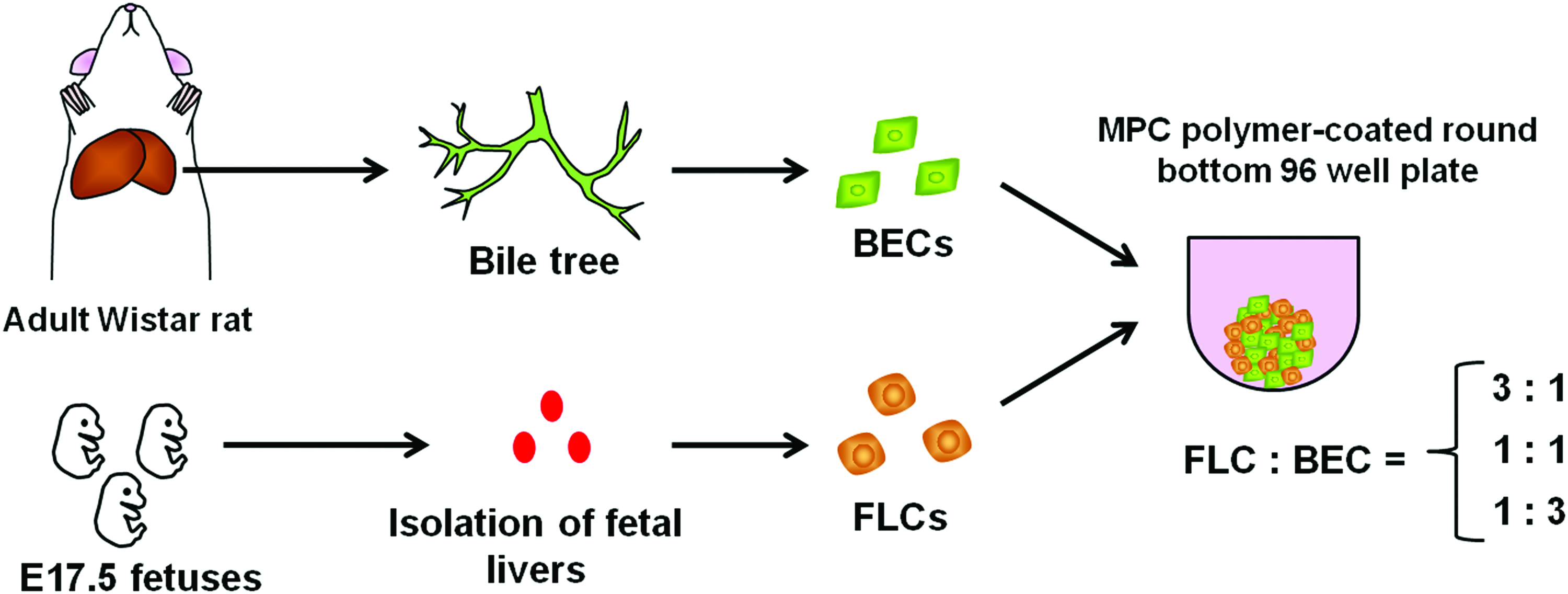
Schematic representation of the method for hetero-spheroid formation. The tree structure containing the BD was extracted from adult Wistar rats after collagenase perfusion, and ABECs were isolated. Meanwhile, fetal livers were harvested from E17.5 fetuses, and FLCs were isolated. ABECs and FLCs were mixed at various ratios and inoculated to low-adherence 96-well plates. ABECs, adult rat primary biliary epithelial cells; BD, bile duct; FLCs, fetal liver cells. Color images available online at www.liebertpub.com/tea
A small portion of the cell suspension was seeded on collagen-coated plates (Asahi Technoglass) or on plates coated with type I collagen gel (Nitta Gelatin) to confirm the ability of the isolated cells to form BD networks. The medium was subsequently changed every other day. When the ABECs that were cultured on gel reached 80%–100% confluency, they were overlaid with collagen gel. Approximately 2 weeks after inoculation, a fluorescein diacetate (FD) assay was performed to confirm BD function.
FLCs were isolated from E17.5 fetuses using the collagenase digestion method described previously. 13 The isolated FLCs were suspended at a concentration of 1×106 cells/mL in FLC culture medium, namely Williams' medium E (Invitrogen) supplemented with 2 mM Glutamax (Invitrogen), 10−6 M DEX, 10 ng/mL of EGF, 20 ng/mL of HGF, 20 ng/mL of fibroblast growth factor (FGF)-1 (Peprotech), 20 ng/mL of FGF-4 (Peprotech), 10 ng/mL of oncostatin M (Wako), 10−7 M insulin, 10−8 M glucagon (Sigma-Aldrich), 0.5 mM ascorbic acid, 10 mM nicotinamide, antibiotic/antimycotic solution, and 10% FBS.
Generation of 3D hepatic hetero-spheroids
For hetero-spheroid formation, we used a 96-well, round-bottom plate coated with 2-methacryloyloxyethyl phosphorylcholine (MPC; NOF Corporation), a phospholipid biocompatible polymer that prevents cells adhesion to the surface of the culture dish. 14 Heterogeneous cell suspensions containing FLCs and ABECs were inoculated at 2×105 cells/well with various ratios of FLC: ABEC=“3: 1,” “1: 1,” and “1: 3.” To investigate the cellular origin of the reconstructed BD structures, we generated hetero-spheroids using FLCs and fluorescent probe-labeled ABECs (see the section “PKH labeling of ABECs”) with the ratios of FLC: ABEC=“10: 1,” “1: 1,” and “1: 10.” The medium was replaced with fresh FLC culture medium every day.
Histochemical and cytochemical staining
Cytokeratin (CK) 19 immunocytochemistry was performed on day 3 to confirm the characteristics of the cells isolated from the intrahepatic tree structures. The cells were fixed in 4% paraformaldehyde (Sigma-Aldrich) in PBS, and treated with 0.2% Triton-X 100 for 5 min to permeabilize the cell membranes. After blocking with 1% bovine serum albumin (Sigma-Aldrich), the cells were incubated with mouse anti-CK19 antibody (Novocastra) at 1: 50 for 1 h at room temperature. Thereafter, the cells were incubated with Alexa Fluor 594-conjugated donkey anti-mouse IgG antibody (Invitrogen) at 1: 500 for 30 min.
Hematoxylin-eosin (HE) staining and immunohistochemistry (IHC) were performed using sections of the hetero-spheroids on day 3 or the retrieved transplants 4 weeks after transplantation using the following methods. The hetero-spheroids or retrieved grafts were fixed in 10% formalin neutral buffer solution (Wako), dehydrated, and then embedded in paraffin wax. Sections with a thickness of 4–5 μm were obtained, dewaxed, and stained with HE using standard procedures. For IHC, dewaxed rehydrated sections were autoclaved in a 10 mM citric acid solution (pH 6.0) at 121°C for 10 min or boiled in 1: 200 diluted ImmunoSaver (Nissin EM) for 45 min and then treated with 0.05% Triton-X 100 for permeabilization. After treatment with a blocking reagent (Nacalai Tesque) at 4°C for 30 min, the sections were incubated at 4°C overnight in a solution of goat anti-CK18 polyclonal antibody (Santa Cruz Biotechnology) (1: 200) or mouse anti-CK 19 antibody (1: 50). The sections were stained using an ImmPRESS anti-mouse or anti-goat Ig peroxidase kit (Vector Laboratories) and a metal-enhanced DAB substrate kit (Thermo Scientific), according to the supplier's instructions. After counterstaining with hematoxylin, specimens were dehydrated and mounted. For double immunofluorescence staining, blocked specimens were incubated at 4°C overnight in the mixture of two primary antibodies, goat anti-CK18 (1: 200) and mouse anti-CK19 (1: 50) antibodies or goat anti-CK18 antibody (1: 200) and mouse anti-MRP2 monoclonal antibody (Abcam) (1: 200). Then, the specimens were incubated at room temperature for 1 h with the mixture of Alexa Fluor 488-conjugated donkey anti-mouse IgG antibody (Invitrogen) (1: 1000) and Alexa Fluor 594-conjugated donkey anti-goat IgG antibody (Invitrogen) (1: 1000). Stained sections were mounted in Vectashield mounting medium with DAPI (Vector Laboratories), and observed using confocal laser scanning microscopy (FV10i; Olympus). Periodic acid-Schiff (PAS) staining was performed with or without salivary diastase pretreatment to detect glycogen.
Functional analysis of the excretion, transport, and accumulation of bile by the hetero-spheroids
To investigate the functionality of reconstructed BDs, we used FD (Sigma-Aldrich), which is metabolized by intracellular esterase into fluorescein. FD is routinely used to assess the excretion, transport, and accumulation of bile in vitro and in vivo.12,15,16 By observing the distribution of metabolized fluorescein, we can assess whether a spheroid possessed functional BDs.
To confirm the bile excretion capacity of ABECs cultured in the collagen-gel sandwich configuration, we incubated the cells with culture medium containing 2.5 μg/mL of FD for 20 min in a CO2 incubator. After a wash with fresh medium, the cells were observed using a confocal laser-scanning microscope (LSM; Carl Zeiss).
Hetero-spheroids were cultured in medium containing 2.5 μg/mL of FD for 40 min in a CO2 incubator, and the distribution of the metabolized fluorescein was observed using an All-in-One fluorescence microscope (BZ-9000; Keyence). Subsequently, the medium was replaced, and the spheroids were incubated in the absence of FD for 40 min. Fluorescein distribution in the spheroids was observed using a BZ-9000 or LSM microscope. Sectional images were obtained by deconvolution using the BZ-9000 microscope. 3D reconstructions were created from images taken with the LSM microscope using ZEN 2009 software (Carl Zeiss) and further processed by 3D surface-rendering using Imaris software (Bitplane).
PKH26 labeling of ABECs
To determine whether the fluorescein accumulations in the hetero-spheroids coincided with the cyst- or duct-structures formed by ABECs, we labeled ABECs with a fluorescent dye before inoculation. ABECs isolated from the BD tree were labeled with a PKH26 red fluorescent labeling kit (Sigma-Aldrich), following the manufacturer's instructions.
Transplantation of hetero-spheroids
To investigate whether the in vitro reconstructed hetero-spheroids were able to maintain their BD networks in vivo, we transplanted “1: 1” hetero-spheroids on day 3 of culture into nude mice (Fig. 2). As a control experiment, spheroids composed solely of FLCs (FLC-spheroids) were transplanted into nude mice in the same manner. One day before the transplantation, the recipient nude mice were given an intraperitoneal injection of carbon tetrachloride (10 μL in 100 μL of olive oil/mouse). Polylactic acid (PLA) nonwoven sheets composed of fibers with an average diameter of 50 μm (Unitika) were used for the fabrication of scaffolds. Discs 7 mm in diameter were cut from the PLA nonwoven sheet and further cut into 4 circular sectors. Each circular sector was used as a scaffold. Fifteen to 20 “1: 1” hetero-spheroids or FLC-spheroids present on day 3 of culture were immobilized in each scaffold by applying 10 μL of Matrigel (BD Biosciences) and solidifying it at 37°C. The spheroid-scaffold composites were transplanted onto the mesenteric membrane of the recipient male nude mice at the age of 18 weeks using a technique similar to that described previously. 6 Four weeks after transplantation, the grafts were retrieved and fixed with 10% formalin, and paraffin sections were prepared from each sample.
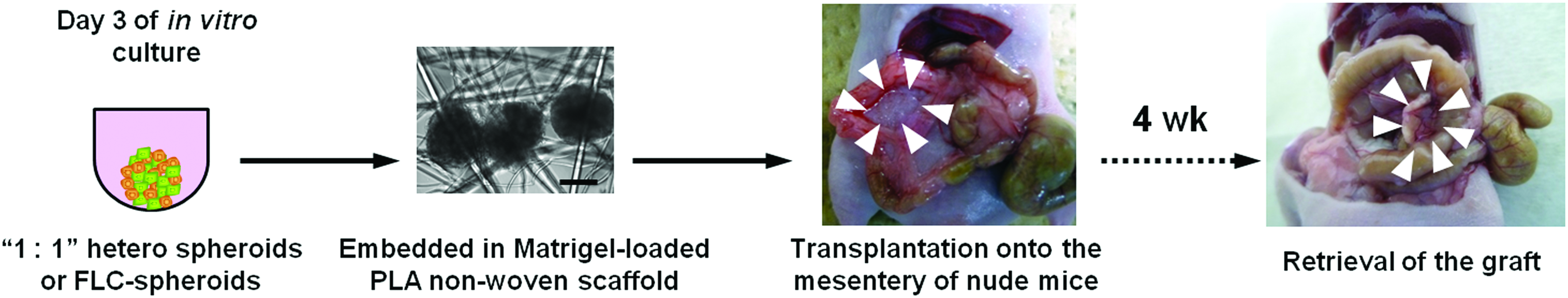
Schematic representation of the method for heterotopic transplantation of hetero-spheroids. On day 3 of culture, “1: 1” hetero-spheroids or spheroids formed by FLCs were embedded in Matrigel-loaded PLA nonwoven scaffold. After solidification of the Matrigel at 37°C, the composites were transplanted onto the mesentery of recipient nude mice. Four weeks after transplantation, grafts were retrieved and fixed in 10% formalin. Arrowheads indicate the location of the grafts. Scale bar: 200 μm. PLA, polylactic acid. Color images available online at www.liebertpub.com/tea
Results
Characterization of FLCs and ABECs in vitro
Before hetero-spheroid formation, we determined the morphological and immunocytochemical characteristics of the isolated FLCs and ABECs. Plated cells isolated from E17.5 fetal rat livers showed the typical morphology of FLCs, as previously described 6 (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea). Cells isolated from the intrahepatic tree structures displayed an epithelial morphology and were positively stained by anti-CK19 antibody (Supplementary Fig. S2). In addition, we confirmed that these cells formed ductular networks in the collagen-gel sandwich configuration, as previously reported (Supplementary Fig. S3A). Accordingly, these structures exhibited the ability to take in FD and excrete the metabolized fluorescein into the luminal spaces (Supplementary Fig. S3B).
In vitro formation of hetero-spheroids
We successfully formed hetero-spheroids containing various ratios of the two cell types by culturing the cells in a 96-well, round-bottom plate coated with MPC polymers. On day 1, cellular adhesion between adjacent cells promoted the formation of loose pancake-shaped aggregates (Supplementary Figs. S4A–C). Within 3 days of culture, the loose aggregates gradually compressed to form compact 3D spheroids (Supplementary Fig. S4D–I).
Histological analysis of hetero-spheroids
HE staining of day 3 hetero-spheroids in each condition revealed that cystic and ductular structures as well as hepatic parenchyma-like structures had formed (Fig. 3A–C). To characterize the cells comprising these structures, we performed IHC for CK19 and CK18. In the liver, CK19 is expressed specifically by the BECs, whereas CK18 is expressed by both the BECs and the hepatocytes. IHC for CK19 clearly demonstrated that BECs had formed cystic and ductular structures (Fig. 3D–F). Of note, the hetero-spheroids formed from cells in a ratio of “1: 3” displayed continuous ductular networks (Fig. 3F), with large cystic structures interlinked by ductular structures. Moreover, IHC for CK18 demonstrated that hepatocytes, which are CK18(+)/CK19(−) cells, composed the parenchyma-like structures that were localized in the outer region (Fig. 3G–I). These results indicated that the ABECs were the cells primarily responsible for the formation of ductular networks.

Histological analyses of hetero-spheroids on day 3. (
We further investigated whether the two cell types were integrated as a tissue structure by double immunostaining for CK18 and CK19. We observed in “1: 1” hetero-spheroids that CK18(+)/CK19(−) hepatocytes and CK18(+)/CK19(+) BECs were in contact with each other (Fig. 4). Moreover, in some regions, CK19(+) ductal structures merged into CK18(+)/CK19(−) parenchymal structures (Fig. 4), suggesting the integration of the two cell types as a tissue structure.
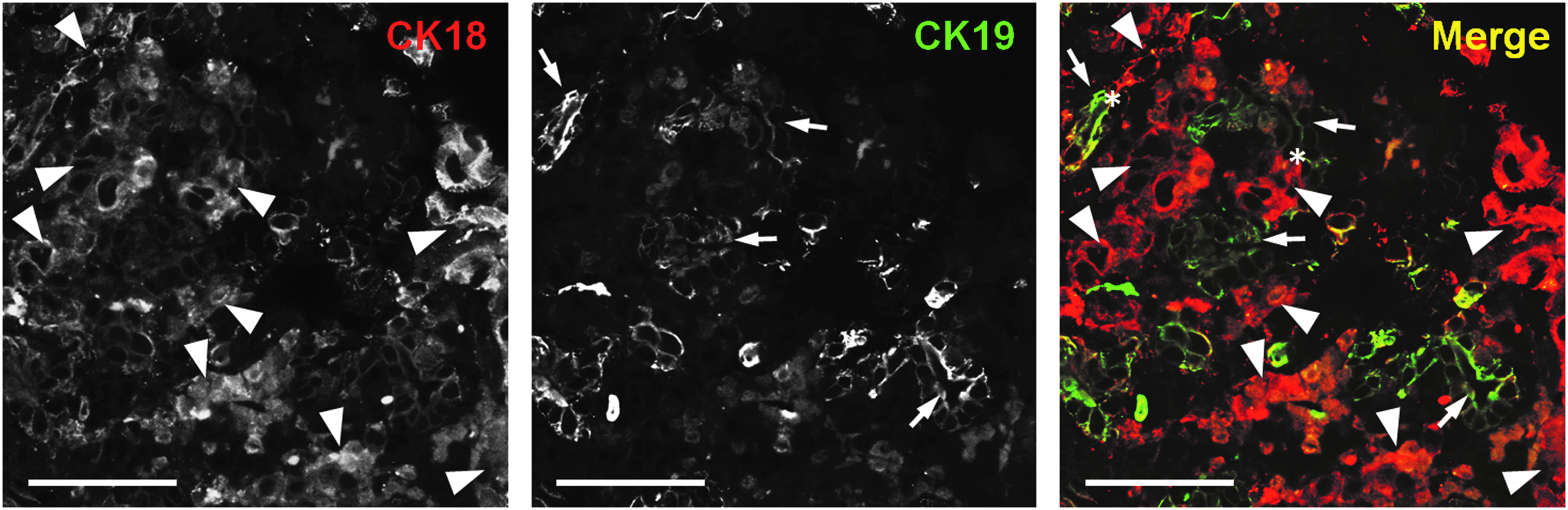
Double immunofluorescent staining for CK18 and CK19. “1: 1” hetero-spheroids on day 3 of culture were stained for CK18 (red) and CK19 (green). Arrowheads indicate CK18(+)/CK19(−) parenchymal structures, whereas arrows indicate CK18(+)/CK19(+) bile ductal structures. Asterisks indicate the junctions that intermediate the parenchymal and ductal structures. Scale bar: 50 μm.
Functional analysis of the BD networks
We examined whether the reconstructed ductular structures could perform the functions of excretion, transport, and accumulation of bile using FD as a marker. On day 4 of culture, the hetero-spheroids were incubated for 40 min with FD and thereafter incubated for 40 min in a FD-free medium. Fluorescein was distributed evenly throughout the “3: 1” hetero-spheroids, indicating that metabolized fluorescein was not excreted to the cystic or ductular structures but instead remained in the cells (Fig. 5A-a, d). In contrast, we observed localized fluorescein distribution in the “1: 1” and “1: 3” spheroids (Fig. 5A-b, c, e, f). In particular, the “1: 3” spheroids displayed large accumulations of fluorescein (Fig. 5A-f). Furthermore, on day 7, the “1: 3” hetero-spheroids exhibited a greater capacity for fluorescein storage: the fluorescein accumulations were 50–100 μm in diameter (Figs. 5B-a, b). In addition, duct-shaped fluorescent areas were also observed between large fluorescein accumulations in the “1: 3” hetero-spheroids (Figs. 5B-d, e). Surface-rendering images suggested that the fluorescein-containing ductular structures interlinked the large fluorescein-containing cystic structures (Fig. 5B-c, f). We also observed that the metabolized fluorescein in the “1: 1” hetero-spheroids localized toward the cystic structures in a time-dependent manner (Fig. 6). To investigate whether the accumulated fluorescein was localized in the cystic structures formed by the ABECs, we labeled ABECs with a red fluorescent dye PKH26 before spheroid formation. Confocal microscopy demonstrated that fluorescein accumulations with a large volume were located in spaces surrounded by the PKH26-labeled ABECs in “1: 1” hetero-spheroids (Fig. 7). Of note, in the “10: 1” hetero-spheroids, no fluorescein accumulations surrounded by PKH-labeled cells were observed (Supplementary Fig. S5A–C). Conversely, in the “1: 10” hetero-spheroids, fluorescein accumulations were observed in the ABEC-surrouned cavities, which were larger than those observed in “1: 1” hetero-spheroids (Supplementary Fig. S5D–F). These results strongly suggest that the cellular origin of the functional duct networks is ABECs, but not FLCs. Taken together, it is demonstrated that ABECs are essential in the establishment of functional BD networks in reconstructed hepatic tissue.

Functional analysis of BD networks in terms of excretion, transport, and accumulation of fluorescent probe.
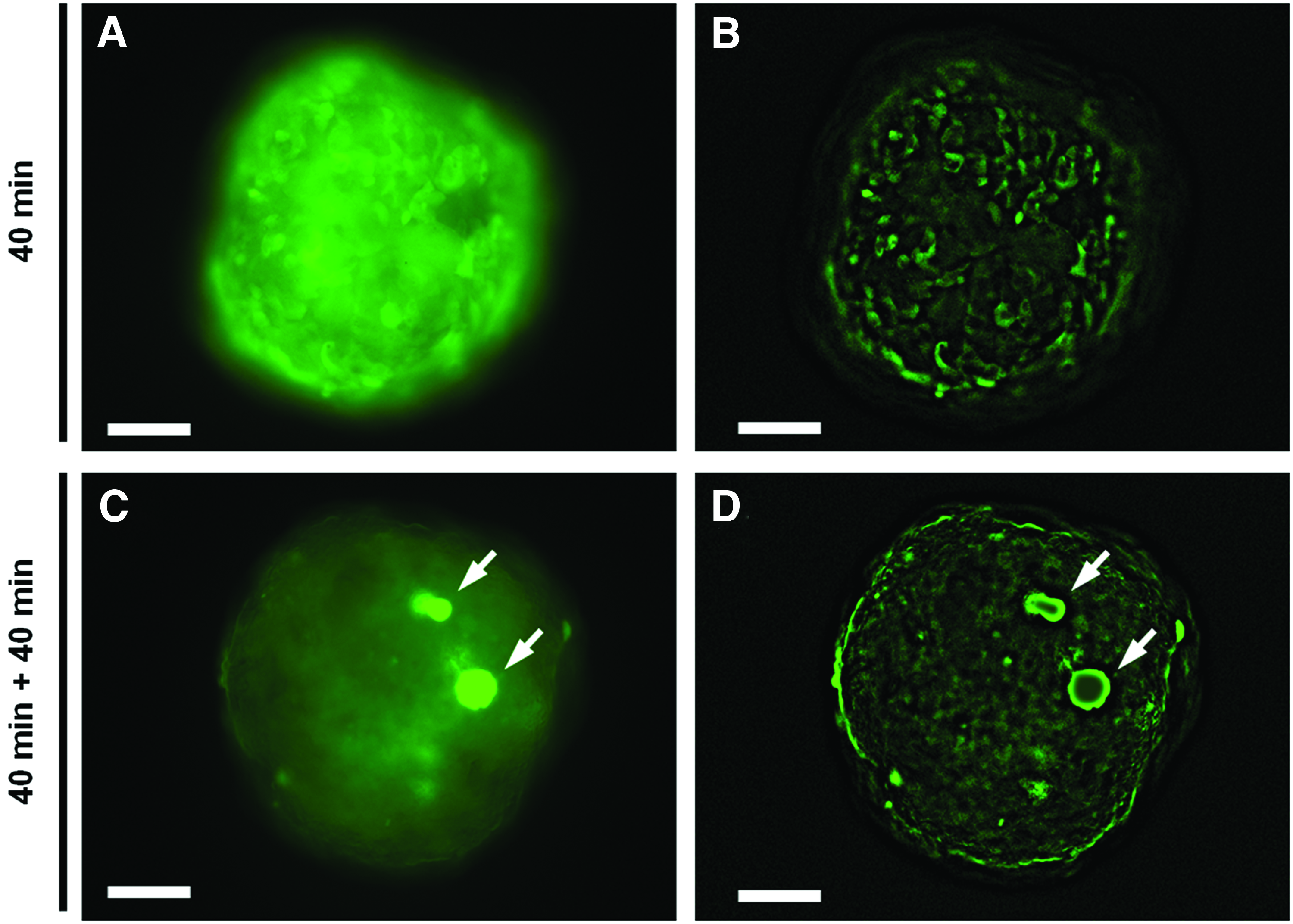
Time course of the localization of metabolized fluorescein in hetero-spheroids. On day 3 of culture, “1: 1” hetero spheroids were incubated with FD for 40 min
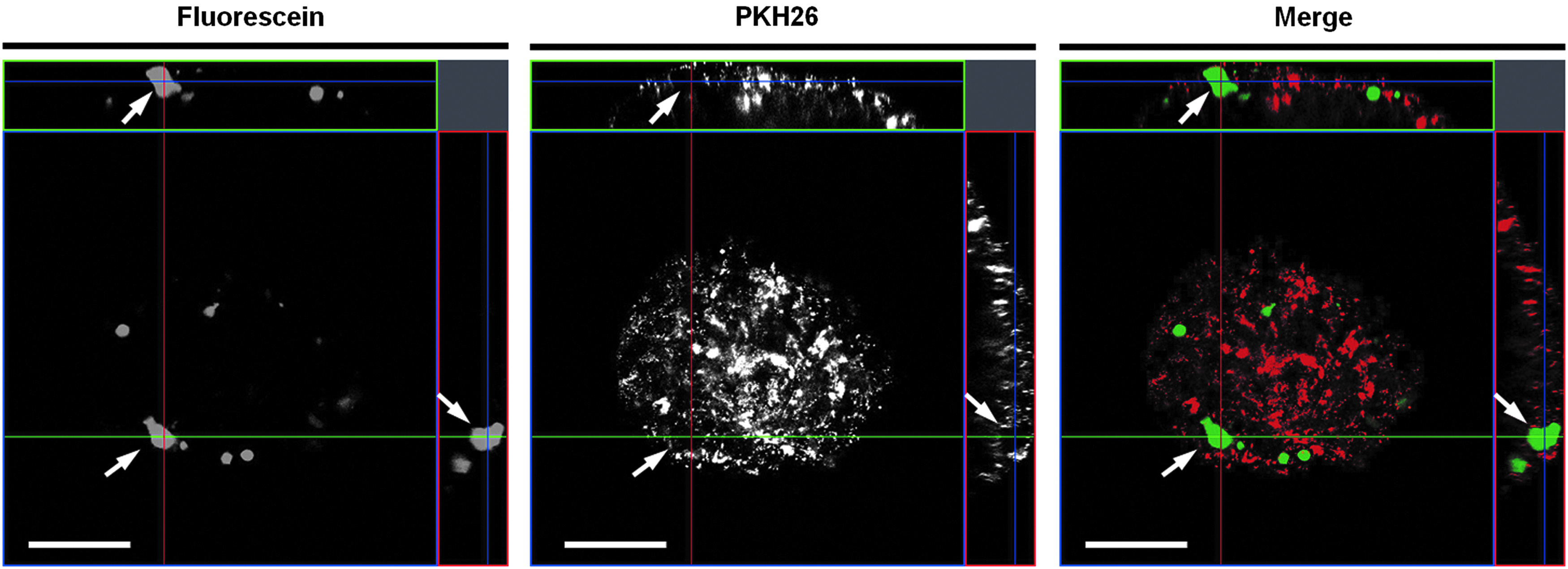
Identification of the positional relation between BECs and accumulated fluorescein. Before hetero-spheroid formation, isolated ABECs were labeled with PKH26, which is a red fluorescent cell linker. “1: 1” hetero spheroids (on day 6) were incubated with FD for 40 min, incubated with fresh medium for further 40 min, and imaged by confocal microscopy. Arrows indicate the fluorescein accumulation surrounded by PKH-labeled ABECs. Scale bar: 100 μm.
In vivo establishment of connected BD networks using hetero-spheroids
We investigated whether hetero-spheroids possessing BD structures would enable the in vivo reconstruction of hepatic tissue equipped with connected BD networks. We transplanted “1: 1” hetero-spheroids embedded in Matrigel-loaded PLA nonwoven scaffolds onto the mesenteric membrane of nude mice (Fig. 2). The purpose of this experiment was to compare the in vivo development of transplanted in vitro-generated hepatic spheroids formed with or without ABECs. Thus, we focused on the “1: 1” hetero-spheroids. Spheroids formed with only FLCs were used as controls. Four weeks after transplantation, the grafts were retrieved and histologically analyzed. HE staining showed that both the “1: 1” hetero-spheroid-derived grafts (“1: 1”-grafts) and the control FLC spheroid-derived grafts (FLC-grafts) possessed liver parenchyma-like structures (Fig. 8A, B). Some of the hepatocytes composing these structures were binucleated, indicating that these cells were well-differentiated compared to those observed before implantation (Fig. 4). Accordingly, PAS staining revealed glycogen stored in parenchyma-like structures (Fig. 8C–F). Meanwhile, the “1: 1”-grafts, but not the FLC-grafts, possessed cystic and ductular structures resembling BDs (Fig. 8A, B). IHC confirmed that both the parenchyma-like structures and BD-like structures were CK18(+) (Fig. 8G, H). In the “1: 1”-grafts, CK19(+) ABECs formed cystic and ductular structures alongside the parenchyma-like structures (Fig. 8I). Moreover, in some areas of the “1: 1”-grafts, CK19(+) ABECs had developed into continuous BD networks, where large cysts were connected to one another by tubular structures (Fig. 8K). In contrast, most of the CK18(+) regions in the FLC-grafts were parenchyma-like structures (Fig. 8H). Although CK19(+) cells were scattered throughout the FLC-grafts, no cystic or ductular structures were observed (Fig. 8L). These results indicated that the FLCs differentiated into both hepatocytes and BECs but did not establish continuous BD networks in vivo. Collectively, these results suggested that ABECs play an important role in the in vivo establishment of BD networks.

Histological analyses of the hetero-spheroid-scaffold composites 4 weeks after transplantation.
Discussion
As noted in the Introduction, the goal of this study was to investigate the ability of ABECs to establish functional BDs in hepatic tissue. We showed that the low-adherence, round-bottom, 96-well plates provided a platform for the formation of hepatic hetero-spheroids composed of FLCs and ABECs. Most importantly, we found that the ABECs effectively contributed to the in vitro establishment of continuous BD networks that performed the functions of excretion, transport, and accumulation of a fluorescent probe. Furthermore, we showed that ABECs were also indispensable to the establishment of continuous BD networks in vivo. We believe that these results provide important insights into the development of transplantable liver tissue that is able to excrete bile.
Our results highlighted the essential roles of ABECs in the reconstruction of functional BD networks in vitro. Conventional approaches using hepatocytes as a cell source have not established BD networks, indicating the necessity of using other cell types, such as hepatic progenitor cells or adult ABECs. Several reports showed that a mixed population of hepatic progenitor cells were capable of forming BD-like structures,9,10 although BD functions were not well-documented. We also observed BD-like structures in hetero-spheroids that contained a relatively small ratio of ABECs (“1: 3”) (Fig. 3A, D, G). However, their BD-like structures did not exhibit BD functionality (Fig. 5). Instead, these hetero-spheroids exhibited the ability to accumulate fluorescein in an ABEC ratio-dependent manner (Fig. 5). Our results suggested that a cell mixture, in which more than half of the cells are ABECs is favorable for the establishment of a functional BD network in hetero-spheroids. In addition, the presence of ABECs allowed BD formation within 3 days of culture, as the spheroids generated with only hepatic progenitors required 8–10 days in culture to develop such structures.9,10 These results demonstrated that ABECs strongly support the in vitro reconstruction of functional BDs.
It was demonstrated that ABECs were also indispensable to the establishment of BD networks in vivo, at least when in vitro-engineered hepatic tissue was used. We previously reported that transplanted FLCs repopulated hepatic tissue in vivo, forming densely packed aggregates within it. 6 However, there were no BD-like structures in those grafts. Likewise, in the present study, we did not observe BD development in the FLC-grafts, strongly suggesting the limited potential of FLCs to reorganize the whole hepatic architecture in vivo. In contrast, the “1: 1” hetero-spheroids contained continuous and branched BD networks. Ogawa et al. also observed that hepatic sheets fabricated with adult liver progenitor cells and nonparenchymal cells could develop BD structures. 17 Taken together, the results demonstrated that the inclusion of ABECs is crucial to the establishment of BD networks not only in vitro but also in vivo.
Our results suggest that the creation of BD networks by dispersed hepatic cells is likely to occur through a mechanism different from that underlying embryonic BD development. Theoretically, FLCs should be able to reconstitute the whole hepatic architecture because they can differentiate into both hepatocytes and BECs. However, we observed that spheroids generated with only FLCs and those containing a relatively small ratio of ABECs (“3: 1”) did not possess well-developed BD networks in vitro (Figs. 3–6) or in vivo (Fig. 8). One possible explanation for this paradox is that the mechanism underlying duct formation by isolated and dispersed cells is different from that underlying embryonic BD development. Embryonic BD formation occurs in association with the commitment of hepatoblasts to the BEC lineage, where hepatoblasts differentiate into BECs and form a monolayered ring of BECs, the so-called ductal plate, which encircles the periportal mesenchyme. 18 Focal areas of the ductal plate become bilayered, and lumens appear between the two cell layers; thus, forming primitive ductal structures. These facts indicate the importance of contact between the BECs and the portal mesenchyme in embryonic duct formation and might provide an explanation for the insufficient BD formation by only FLCs despite their bipotentiality. Namely, isolated FLCs lose their interaction with the portal mesenchyme and miss the cues for duct formation as well as for differentiation into BECs. In contrast to the bipotent FLCs, ABECs, which are already terminally differentiated, do not require inductive signals for commitment to the BEC lineage. Thus, it seems beneficial to employ ABECs for BD reconstruction. Considering all of the results, it appears that the ABECs, rather than the bipotent FLCs, play a central role in the duct formation in the hetero-spheroids.
We believe that the present study will contribute to the development of a transplantable hepatic tissue equivalent, though there are still problems to solve. We propose that the challenge regarding the establishment of a bile clearance system can be broken down into the following 4 elements: (1) the establishment of functional BCs, (2) the establishment of functional BD networks, (3) interlinking the BCs with BDs and enabling bile transport from the hepatocytes to the BD networks, and (4) connecting the BDs established in the grafts to the host duodenum. The formation of functional BCs is relatively well-studied,19–21 and recent reports have shown that the 3D formation of hetero-spheroids allows the establishment of well-developed BCs.22,23 Meanwhile, BD formation has not been the focus of liver tissue engineering. Here we provide important insights into BD formation. We have also provided the data showing that the ductal structures merged into parenchymal structures, which suggest the possibility of the interlinkage between of BCs with BDs. The next challenge is to demonstrate that bile is transported from the hepatocytes to the BDs via BCs.
Despite the successful reconstruction of functional BDs, our method did not allow the formation of well-developed BCs. This is likely because the formation of well-developed BCs requires that the hepatocytes be densely packed, which did not occur in the hetero-spheroids with the higher ABEC content. In fact, we confirmed the potential of the FLCs to form BCs at the densely-packed parenchymal regions in the hetero-spheroids. Hepatocytes composing the “1: 1” hetero-spheroids partly expressed MRP2, an important transporter for biliary excretion expressed in BCs (Supplementary Fig. S6). One possible strategy to establish BC-BD connections would be to produce hepatic tissue by combining hepatocyte-rich spheroids possessing BCs22,23 with hetero-spheroids possessing BD networks. In addition to these immediate goals, the ultimate goal of transplanted hepatic tissue that is connected to the host duodenum via BDs will require a breakthrough that is beyond the present study. Possible strategies for meeting this challenge are developing biomaterials, such as artificial blood vessels to connect the BDs in the grafts to the duodenum. For this approach, it will be important to select the optimal transplantation site that supports connections between the grafts and the duodenum. Although this final issue requires further investigation, it should be noted that the present study has provided important insights into the potential development of a transplantable liver tissue equivalent.
Conclusions
We demonstrated that ABECs played an important role in the reconstruction of functional BD networks in vitro. We also showed that ABECs were indispensable to the establishment of BD networks in vivo. We believe that this study has provided important insights into the development of a transplantable liver tissue equivalent that would resolve the bile clearance problem.
Footnotes
Acknowledgments
The authors thank Ms. Kayoko Suenaga (Carl Zeiss) for assisting with the image processing using the IMARIS software, and Dr. Masaki Kawamata (National Cancer Center) for preparing pregnant rats and his technical advices. This work was supported by a Grant-in-Aid from the Japan Society for the Promotion of Science (JSPS) Fellows, a Grant-in-Aid for Challenging Exploratory Research from JSPS, the CREST program of the Japan Science and Technology Agency, a Grant-in-Aid for the Third-Term Comprehensive 10-Year Strategy for Cancer Control in Japan, a Grant-in-Aid for Scientific Research on Priority Areas Cancer from the Japanese Ministry of Education, Culture, Sports, Science and Technology, a Grant-in-Aid for cancer research promotion from the National Cancer Center of Japan, and a Grant-in-Aid for Scientific Research on Innovative Areas (‘‘functional machinery for noncoding RNAs'’) from the Japanese Ministry of Education, Culture, Sports, Science, and Technology.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.