
Abstract
The immunogenicity of tendon-derived stem cells (TDSCs) has implications for their clinical use for the promotion of tendon repair. The immunogenicity and escape mechanisms of rat patellar TDSCs were examined after allogeneic transplantation. Our results showed that TDSCs exhibited low immunogenicity as evidenced by the following: (i) the incubation of target TDSCs with immunized serum did not show antibody recognition and did not induce the complement-dependent cytotoxicity; (ii) target TDSCs elicited a very low level of lymphocyte proliferation and did not exhibit host lymphocyte-mediated cytotoxicity; and (iii) target TDSCs dose dependently suppressed the phorbol 12-myristate 13-acetate (PMA)- and ionomycin-induced host lymphocyte proliferation. For the mechanistic studies, TDSCs expressed major histocompatibility complex (MHC)-I but a very low level of MHC-II, CD86 and CD80 for the induction of T-cell response. Also, TDSCs were found to express intracellular Fas and FasL. γ-IFN pretreatment did not increase the level of MHC-II and CD86 for the upregulation of immune response. Moreover, the immunosuppressive mediators indoleamine 2,3-dioxygenase (IDO) and transforming growth factor-beta 1 (TGF-β1) were found not to be involved in the escape mechanism of target TDSCs from host lymphocyte attack. In conclusion, allogeneic TDSCs exhibited low immunogenicity. Allogeneic TDSCs might be used for transplantation.
Introduction
I
Many previous studies have shown that bone marrow-derived stem cells (BMSCs) could avoid immuno-rejection and hence an allogeneic cell source could be used for transplantation.3–11 However, there has been no study on the immunogenicity of TDSCs. The immunogenicity of TDSCs has implications for their clinical use for the promotion of tendon repair as the use of an autologous cell source may induce donor site morbidity.
In part II of our study, we aimed to examine the immunogenicity and escape mechanisms of rat patellar TDSCs. We hypothesized that TDSCs are immuno-privileged cells that are suitable for allogeneic transplantation. Information from this study would provide information about the immunogenicity and escape mechanisms of TDSCs after allogeneic transplantation and insights on the molecular mechanisms of suppression of infiltration of inflammatory cells after TDSC transplantation. The T- and B-cell responses of TDSCs using lymphocytes and serum collected from the immunized animals were examined. Mechanistically, the expression of immuno-phenotypic markers including major histocompatibility complex (MHC)-I, MHC-II, CD0, CD86, Fas, FasL, with and without pretreatment with γ-IFN, in TDSCs was examined by flow cytometry. The escape mechanism of TDSCs from lymphocyte attack was explored by measuring the release of immunosuppressive mediators indoleamine 2,3-dioxygenase (IDO) and transforming growth factor-beta 1 (TGF-β1) and immuno-stimulatory mediator tumor necrosis factor-alpha (TNF-α) in the co-culture supernatant.
Materials and Methods
Isolation and culture of rat patellar TDSCs
All the experiments were approved by the Animal Research Ethics Committee, the Chinese University of Hong Kong. Four- to six-week-old male outbred wild-type Sprague-Dawley (SD) rats (150–220 g) were used in this study. The procedures for the isolation of patellar TDSCs have been established. 12 The clonogenicity and multi-lineage differentiation potential of these cells were confirmed before being used for the experiments in this study using standard assays as described previously. 12 TDSCs at P5 were used in all experiments.
Animal surgery
Twenty-four wild-type SD rats (7–8 weeks, body weight of 250–320 g) were used in this study. To create the tendon defect, the central portion of the patellar tendon (∼1 mm in width) was removed from the distal apex of the patella to the insertion of the tibial tuberosity without damaging the fibrocartilage zone with three stacked sharp blades with the middle blade moved up to create a gap between the two outer blades as described in our previous study. 13
For the experiments to investigate the T- and B-cell responses to TDSCs, fibrin constructs with wild-type TDSCs were transplanted into 24 animals. Wild-type TDSCs were used because the fluorescence signal of GFP might affect the calcein signal in the lymphocyte-mediated TDSC lysis assay and the complement-dependent cytotoxicity (CDC) assay. At week 0, 2, 6, and 8, the spleens from six animals were harvested and weighed. The weights of spleens of the fibrin glue-only control group from part I of the study published in the same issue were also measured and served as controls. Lymphocytes were isolated and tested for their effects to recognize the target TDSCs using the lymphocyte proliferation and lymphocyte-mediated TDSC lysis assays. The escape mechanism of TDSCs from lymphocyte attack was investigated by measuring the level of positive and negative immuno-modulatory factors such as TNF-α, IDO, and TGF-β1 in the supernatant of TDSC-lymphocyte co-culture. At pre-operation, week 1, 2, 4, and 8 for the week 8 group (n=6 for each time point), serum was collected longitudinally by eye puncture and was used for testing the B-cell responses against TDSCs by the CDC and antigen-antibody binding assays. Supplementary Figure S1 (Supplementary Data are available online at www.liebertpub.com/tea) showed a schematic diagram of the study design. Week 8 post-operation was chosen as the longest time for investigating the B- and T-cell responses as our previous study showed that allogeneic transplantation of TDSCs promoted tendon repair at week 2 and 4. 13 Hence, the memory B- and T-cell clones against TDSCs, if any, should be developed by week 8 post-operation. This was also confirmed by the stimulation of B-cell responses as early as week 1 after injection of sheep red blood cells (SRBCs) into the animals in the positive control (see description below).
The TDSC-fibrin constructs were prepared for in vivo transplantation as described previously. 13 Each construct contained 4×105 cells in 20 μL fibrin glue. The fibrin glue construct with TDSCs was placed in the tendon defect horizontally with the length and width of the construct fitted into the length and width of the wound, respectively. The wound was then closed in layers. The animals were allowed to have free-cage activity until euthanasia.
Wild-type SD rats are outbred animals. The transplantation of TDSCs isolated from wild-type SD rats to other wild-type SD rats is an allogeneic transplantation because outbred animals are genetically different individuals.
Lymphocyte proliferation assay
Lymphocytes were isolated from six spleens of immunized animals at each time point using Lymphoprep™ according to the manufacturer's instructions (Axis-Shield PoC, Oslo, Norway) and cultured in RPMI 1640 supplemented with 10% (v/v) fetal bovine serum, 100 U/mL penicillin G, and 0.1 mg/mL streptomycin (Invitrogen, Carlsbad, CA).
Same TDSCs at P5 as that were used for transplantation (target TDSCs), at 1×106/mL were treated with 8 μg/mL mitomycin-c (Sigma-Aldrich, St Louis, MO) at 37°C for 1 h. After washing with RPMI twice, 1×104 (i.e., 0.01 M, 1:50) or 5×104 (i.e., 0.05 M, 1:10) mitomycin-c-treated TDSCs were seeded in a 96-well plate and mixed with constant number of lymphocytes isolated from the immunized animals (5×105, i.e., 0.5 M) in a final volume of 250 μL/well and incubated at 37°C, 5% CO2. At 77 h after incubation, the mixture was pulsed with 1 μCi [
3
H]-thymidine (PerkinElmer, Boston, MA) for 17 h, which was the optimal time as determined in a previous study (results not shown). The proliferation of lymphocytes was then measured using a liquid scintillation counter (Packard TopCount NXT™ microplate scintillation and luminescence counter; PerkinElmer). Lymphocytes treated with phorbol 12-myristate 13-acetate (PMA) (80 ng/mL) (Sigma-Aldrich) and ionomycin (0.5 μM) (Merck, Darmstadt, Germany) were used as the positive control in this study.14,15 The effect of TDSCs on the inhibition of PMA- and ionomycin-induced lymphocyte proliferation was investigated by adding TDSCs to the PMA- and ionomycin-treated lymphocytes. Eight μg/mL mitomycin-c was used to inhibit the proliferation of TDSCs because our previous study showed that mitomycin-c at this dose inhibited the DNA synthesis while maintained the metabolic activity of TDSCs (results not shown). Supplementary Figure S2 showed the design of this experiment. The results were presented as follows:
Lymphocyte-mediated TDSC lysis assay
Same TDSCs at P5 as that used for transplantation (target TDSCs), at 1×106/mL were treated with 8 μg/mL mitomycin-c and 1 μM calcein-AM (Molecular Probes, Life Technologies, Carlsbad, CA) at 37°C for 1 h. After washing with RPMI twice, 1×104 (i.e., 0.01 M, 1:50) or 5×104 (i.e., 0.05 M, 1:10) mitomycin-c-treated, labeled TDSCs were seeded in a 96-well plate and mixed with constant number of lymphocytes isolated from the immunized animals (5×105, i.e., 0.5 M) in a final volume of 250 μL/well and incubated at 37°C, 5% CO2. At 4 and 24 h after incubation, the fluorescent intensity of the supernatant was measured by a fluorescent plate reader (Tecan, Männedorf, Switzerland) with an excitation (Ex) and emission (Em) wavelength at 485 and 535 nm, respectively. TDSCs treated with 0.1% Triton X-100 were used to represent total cell lysis. Supplementary Figure S2 showed the design of this experiment. The results were presented as follows:
CDC assay
Same TDSCs at P5 as that were used for transplantation (target TDSCs), at 1×106/mL were loaded with 1 μM calcein-AM in the dark for 1 h at 37°C. The TDSCs were then washed twice and seeded at 2×104/well (i.e., 0.02 M/well) in a 96-well plate overnight. After washing the cells with modified barbital buffer (MBB buffer) with 1% bovine serum albumin (BSA), serum collected from the immunized animals, at 1×and 0.5×(diluted in MBB with 1% BSA) or antibodies against CD73, which is a marker for TDSCs (1:10 or 1:20; BD Biosciences, Franklin Lakes, NJ), were added to the TDSCs for 30 min at 37°C. The supernatant was then removed and guinea pig complement (1:10 diluted in MBB with 1% BSA) (Cedarlane Laboratories USA, Inc., Burlington, NC) was then added and incubated at 37°C for 1 h. The lysis and survival of TDSCs were measured by the release of calcein into the supernatant at Ex 485 nm/Em 535 nm and MTT assay, respectively. The MTT assay was performed according to the instructions of the manufacturer (USB; Affymetrix, Inc., Santa Clara, CA). TDSCs treated with 0.1% Triton X-100 were used to represent total cell lysis. As a positive control, SRBCs were injected at 1×109 in 1 mL phosphate-buffered saline (PBS) i.p. into SD rats. Serum collected longitudinally at preinjection, week 1, 2, 4, and 8 by eye puncture (n=6) and diluted to 1:100 or 1:1000 (at a much higher dilution compared to the serum collected from the animals with TDSC transplantation), was used for the CDC assay the same as that for the TDSCs. SRBCs treated with 0.1% Triton X-100 were used to represent total cell lysis. Hemolysin (antibody against SRBCs, 1:100 diluted in MBB; Serotech Laboratories Ltd., Toronto, Canada) was used as a positive control for the CDC of SRBCs. Supplementary Figure S3 showed the design of this experiment. The results were presented as follows:
Antigen-antibody binding assay
Same TDSCs at P5 as that used for transplantation (target TDSCs) were seeded at 2×104/well in a 96-well plate overnight. After washing the cells with PBS (1% BSA), serum collected from the animals immunized with TDSCs (diluted to 0.5×or 0.25×with PBS (1% BSA), which were at high concentration in immunological studies) or anti-CD73 [1:100 diluted with PBS (1% BSA)] was added and incubated at 37°C for 30 min. After washing, anti-rat antibodies-horseradish peroxidase (HRP) (1:1000 diluted in PBS with 1% BSA) (Abcam, Cambridge, United Kingdom) (for serum groups) or anti-mouse antibodies-HRP (1:1000) (for anti-CD73) (Merck Millipore, Billerica, MA) were added and incubated at 37°C for 1 h. After washing, o-phenylenediamine (OPD) substrate (0.4 mg/mL with 0.012% H2O2 in 0.1 M citrate buffer, pH 5.0; Sigma-Aldrich) was added and the reaction was stopped by adding 4 M H2SO4 after 5 min with shaking at room temperature. The absorbance was measured at optical density (OD) 490 nm. Supplementary Figure S3 showed the design of this experiment. The results were presented as follows:
Flow activated cell sorting analysis
The immuno-phenotype of wild-type TDSCs, with and without pretreatment with γ-IFN (100 U/mL) for 48 h, was examined by flow cytometry (Becton Dickinson, Franklin Lakes, NJ) as described previously. 12 The expression of MHC-I, MHC-II, CD80, CD86, Fas, and FasL in TDSCs were examined. The expression of Fas and FasL has been shown in the previous studies to induce tolerance to promote target cell survival.16–21 γ-IFN regulated the expression of MHC-II and co-stimulatory molecules. 22 It was used to simulate an inflammatory environment after transplantation in vivo and test whether this would affect the immuno-phenotype and hence the immunogenicity of the cells. This method has been well reported.10,22,23
Wild-type TDSCs were fixed with formaldehyde (0.01%, v/v in PBS) and permeabilized with Tween-20 (0.5%, v/v in PBS) for studying the expression of Fas and FasL. Antibodies used included (i) anti-MHC-I (secondary antibody: anti-mouse IgG2b-PE), (ii) anti-MHC-II (secondary antibody: anti-mouse IgG-FITC), (iii) anti-CD80 (secondary antibody: anti-mouse IgG1-PE), (iv) anti-CD86 (secondary antibody: anti-mouse IgG1-PE), (v) anti-Fas (secondary antibody: anti-rabbit IgG-FITC), and (vi) anti-FasL (secondary antibody: anti-mouse IgG2b-PE). All the antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Cells were stained with secondary antibodies alone in the negative controls.
Measurement of soluble immuno-modulatory factors
Same TDSCs at P5 as that used for transplantation (target TDSCs) at 1×106/mL were treated with 8 μg/mL mitomycin-c at 37°C for 1 h. After washing with RPMI twice, 6×104 (i.e., 0.06 M, 1:50) mitomycin-c-treated TDSCs were seeded in a 24-well plate and mixed with constant number of lymphocytes isolated from the immunized animals (3×106, i.e., 3 M) in a final volume of 1.5 mL/well and incubated at 37°C, 5% CO2 for 24 h. The supernatant was collected and stored at−80°C until used for the measurement of IDO (USCN Life Sciences, Wuhan, China), TNF-α (Abcam), and TGF-β1 (Abcam) by commercially available ELISA kits. Supplementary Figure S2 showed the design of this experiment.
Data analysis
Quantitative data were shown in bar chart with mean±standard deviation. The difference in weight of spleens between the control and the TDSC groups at each time point was compared using Mann–Whitney U test while the difference in weight of spleens across different time points in the control and the TDSC groups was compared using Kruskal–Wallis test followed by post hoc pairwise comparison using Mann–Whitney U test. The comparisons among different treatment groups in each T- and B-cell assay and the comparison among serum collected longitudinally at different time points in the B-cell assay were done by Friedman test, followed by post hoc pair-wise comparison using Wilcoxon Signed Rank test. Y-axes representing 100% TDSC lysis or survival were shown for the lymphocyte-mediated TDSC lysis assay and the CDC assay to better illustrate the clinical significance of any statistically significant difference which is influenced by the sample size and sample variation. All the data analysis was done using SPSS analysis software (version 16.0; SPSS, Inc., Chicago, IL). p≤0.050 was regarded as statistically significant.
Results
Changes in spleen weight
The change in weight of secondary lymphoid organ spleen is regarded as a gross indicator of the immuno-modulatory activity of TDSCs after transplantation. As shown in Figure 1, there was a transient increase in spleen weight at week 2 after TDSC transplantation when compared with that of the control group (p=0.004). There was no significant difference in spleen weight in the TDSC group compared to the control group at week 8 (p=0.253).
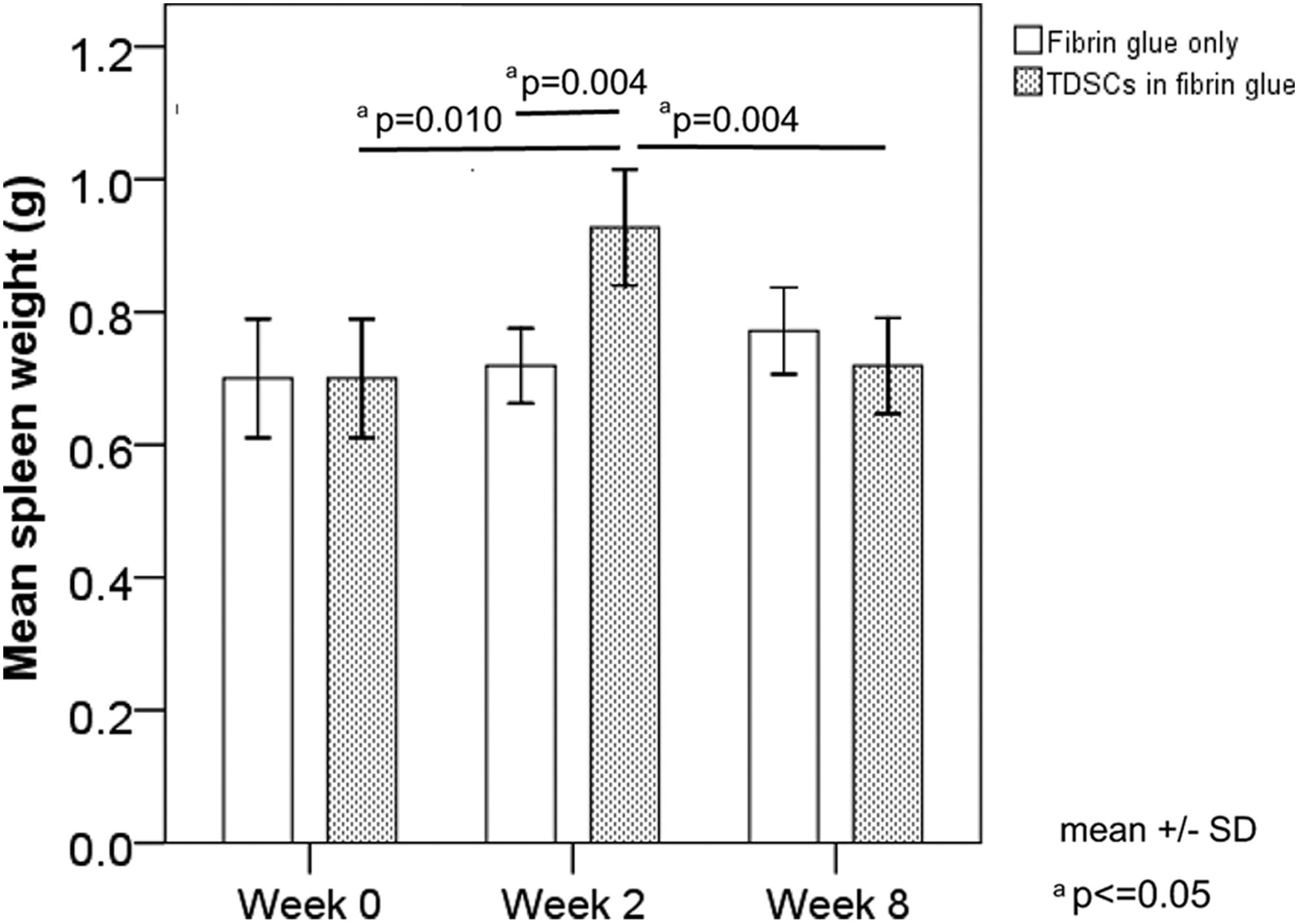
Graph showing the spleen weight of rats transplanted with or without tendon-derived stem cells (TDSCs). n=6/group/time point.
Lymphocyte proliferation assay
Lymphocytes collected at weeks 0, 2, 6, and 8 after TDSC transplantation were used. We reasoned that if the host generated a strong immune response against the TDSCs after transplantation, mixing the mitomycin-c-treated target TDSCs (unable to undergo cell proliferation) with the host lymphocytes would elicit a strong cell proliferation for clonal expansion as reflected by an increase in the [ 3 H]-thymidine incorporation. To our surprise, co-culture of the lymphocytes from the hosts with transplantation of the target TDSCs at different ratios only produced a minimal increase in lymphocyte proliferation at different time points (0.08–3-folds relative to the corresponding lymphocyte-only group) (Fig. 2). To validate our assay system, PMA and ionomycin were used as a positive control to generate [ 3 H]-thymidine incorporation. As can be seen in Figure 2, PMA and ionomycin induced a significant increase in lymphocyte proliferation (8–36-folds relative to the corresponding lymphocyte-only group). Interestingly, when the TDSCs were incubated with the host lymphocytes treated with PMA and ionomycin, they significantly inhibited the PMA- and ionomycin-induced lymphocyte proliferation (0.2–14.9-folds relative to the corresponding lymphocyte-only group, representing 30–98% reduction compared with the corresponding PMA- and ionomycin-treated group). This indicated that TDSCs might carry some suppressive factors for the induction of lymphocyte proliferation mediated by PMA and ionomycin. Also, this suppressive effect was TDSC dose-dependent (Fig. 2).
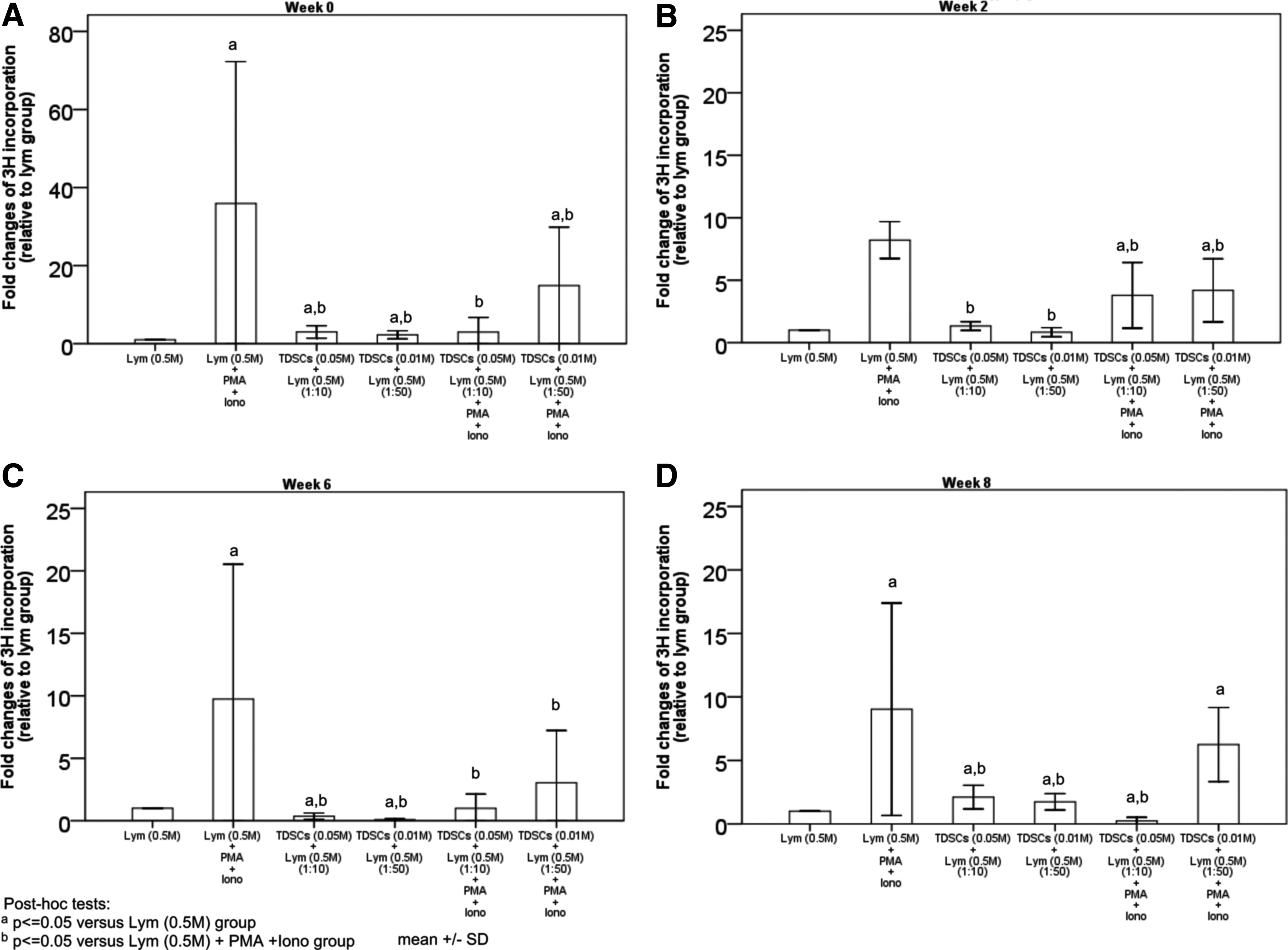
Lymphocyte proliferation assay. Graphs showing the fold changes of lymphocyte proliferation in vitro with lymphocytes collected at
Lymphocyte-mediated TDSC lysis assay
Next, we evaluated whether the host lymphocytes acquired the target cell killing activity. Lymphocytes collected at week 0, 2, 6, and 8 after TDSC transplantation were mixed with the mitomycin-c-treated and calcein-loaded target TDSCs with different effector/target cell ratio to determine the degree of killing by measuring the calcein fluorescence released in the supernatant. As shown in Figure 3 and Supplementary Figure S4, lymphocytes isolated from the hosts with TDSCs transplantation did not induce the death of target TDSCs while Triton X-100 induced the total lysis of TDSCs after incubation for 4 and 24 h, respectively, when compared with the spontaneous release of calcein from the control. These results indicated that the transplantation of TDSCs did not generate a strong T-cell immune response against the target TDSCs.
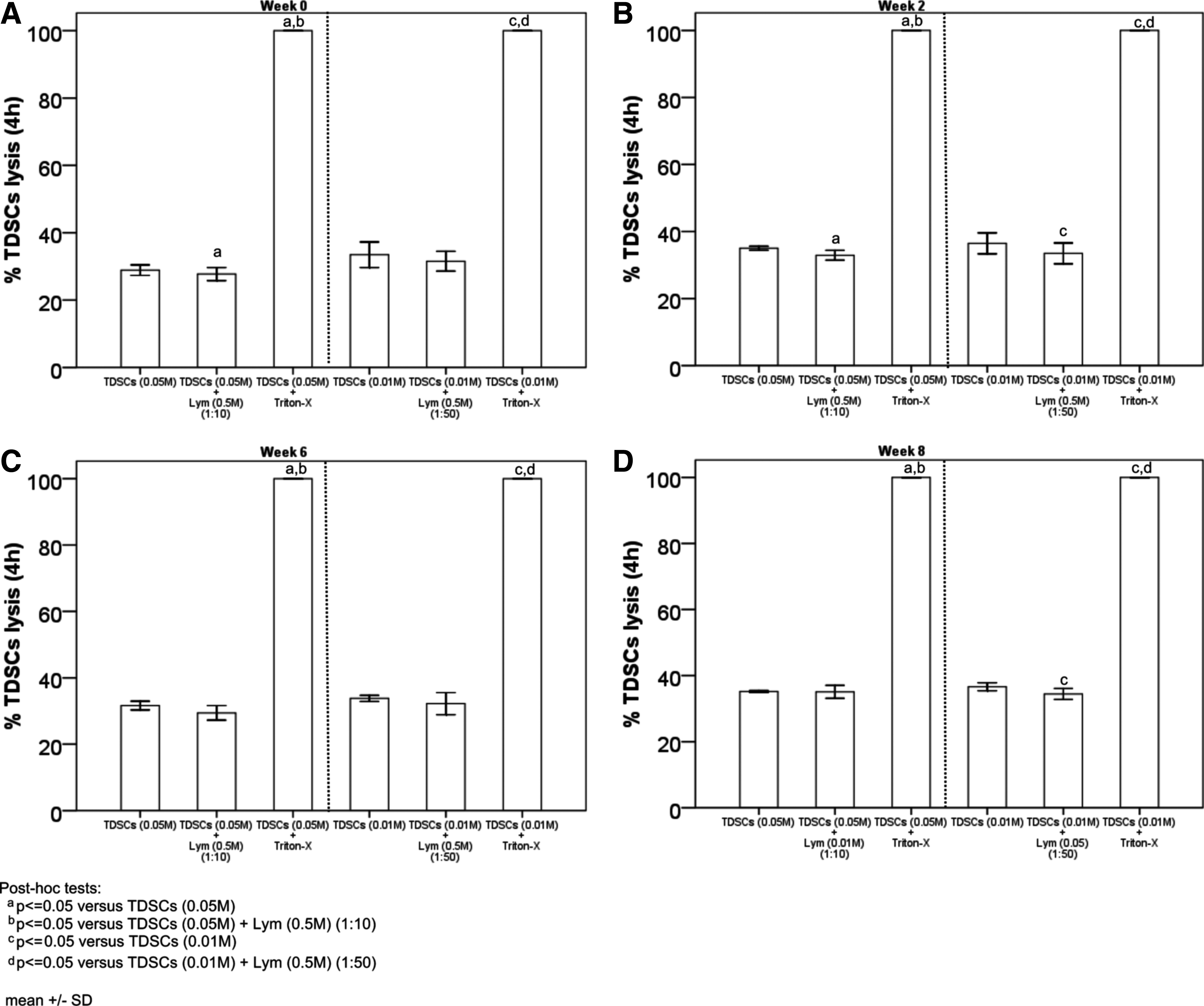
Lymphocyte-mediated TDSC lysis assay. Graphs showing the percentage of lymphocyte-mediated TDSC lysis at 4 h in vitro with lymphocytes collected at
Complement-dependent cell lysis assay
One of the mechanisms to remove the foreign transplant is mediated by the CDC. The results in Figure 4B and E indicated that complement alone did not induce the death of target TDSCs. Also, serum collected from the TDSCs-immunized animals at different times (week 0, 1, 2, 4, and 8) and at different dilutions did not induce clinically significant lysis of target TDSCs in the presence or absence of complement (Fig. 4A, D). Interestingly, antibody against TDSC marker CD73 was unable to induce the complement-dependent lysis of TDSCs (Fig. 4C, F).
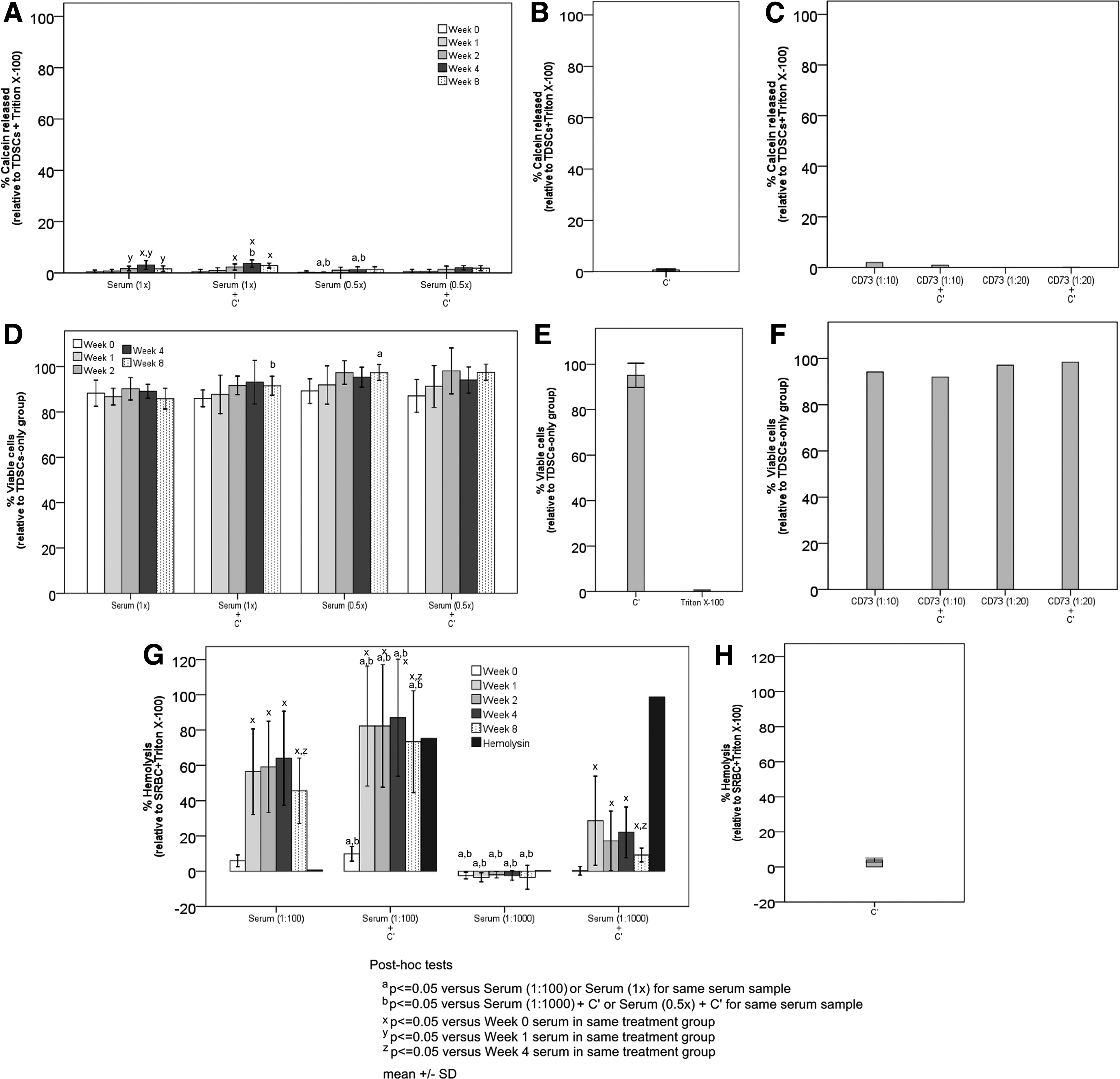
Complement-dependent cytotoxicity assay.
On the contrary, serum collected from the animals immunized with SRBCs at different times (week 0, 1, 2, 4, and 8) significantly induced higher percentage of hemolysis of target SRBCs in the presence of additional complement or endogenous complement in serum compared with that from unimmunized animals [overall p-values for the comparison of different time points for each treatment=0.004, 0.003, 0.856, and 0.026 for Serum (1:100), Serum (1:100)+C′, Serum (1:1000), and Serum (1:1000)+C′, respectively] (Fig. 4G). Also, the hemolysis of target SRBCs occurred at much lower serum concentrations compared with the serum concentrations for testing the lysis of target TDSCs (1:100 or 1:1000 vs. 1:1 or 1:2). There was significant reduction in the hemolysis of target SRBCs treated with serum collected at week 8 compared with the treatment of target SRBCs with serum collected at week 4 [post hoc p-values=0.028, 0.028, and 0.046 for Serum (1:100), Serum (1:100)+C′, and Serum (1:1000)+C′, respectively] (Fig. 4G). Both higher serum concentration and the addition of complement induced higher percentage of hemolysis of target SRBCs (overall p-values for the comparison of different treatments of serum collected at each time point: all p<0.001) (Fig. 4G). Hemolysin induced the lysis of SRBCs in the presence of additional complement (Fig. 4G) while the complement alone did not induce the death of SRBCs (Fig. 4H). Taken together, our results indicated that the complement-mediated lysis system was functional for SRBC while the same was not observed in TDSCs.
Antigen-antibody binding assay
Serum collected from the TDSC-immunized animals at different times (week 0, 1, 2, 4, and 8) and at different dilutions did not induce clinically significant TDSC binding as the absorbance at 490 nm was very low in all groups (about 0.1 arbitrary unit) and the serum concentrations tested were high. On the other hand, CD73 induced binding of TDSCs (Fig. 5).
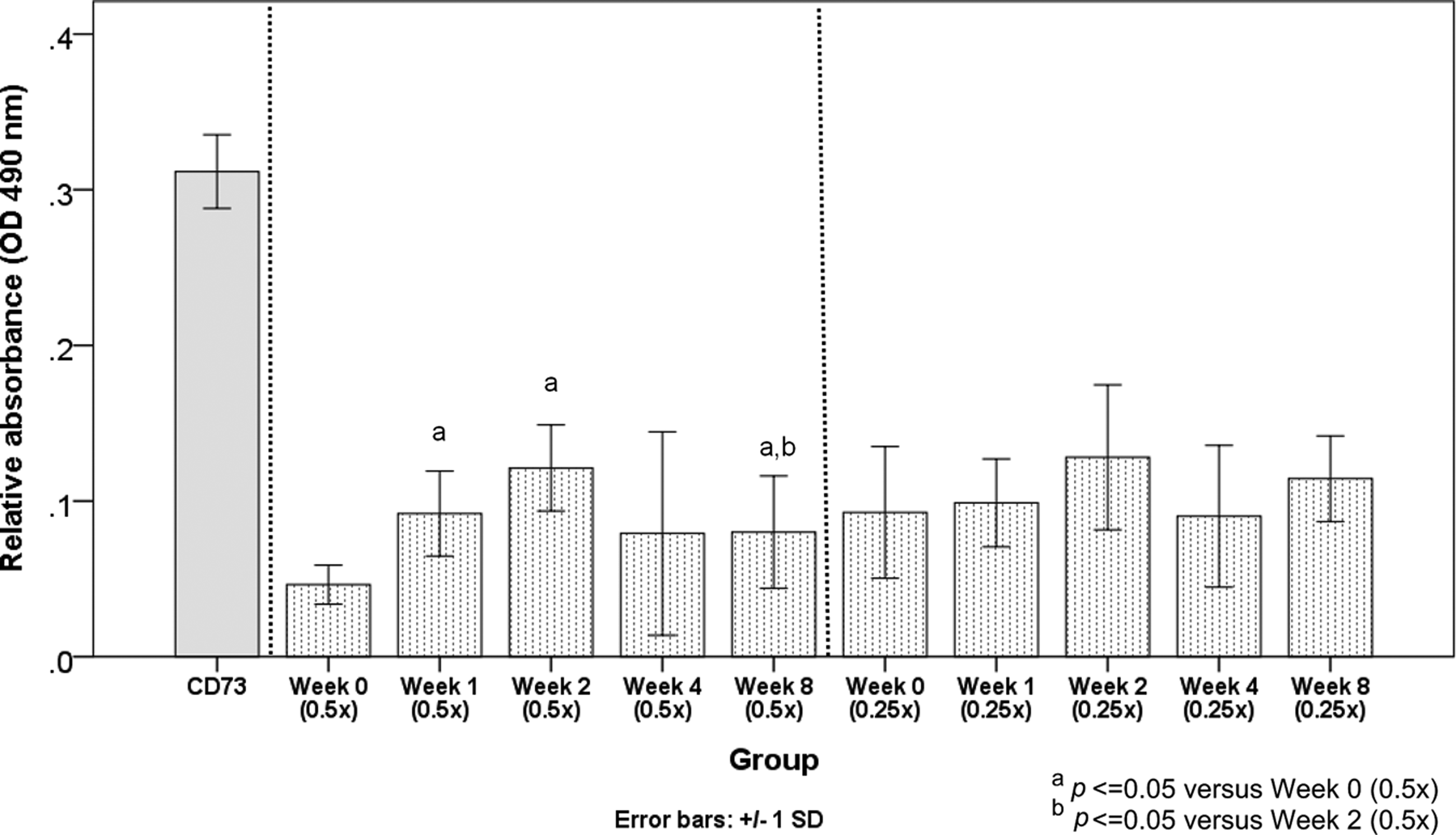
Antigen-antibody binding assay. Graph showing the binding of target TDSCs treated with different concentrations of serum collected from immunized animals at different times (week 0, 1, 2, 4, and 8) after TDSC transplantation. Anti-CD73 was used as a positive control. n=6/group.
Immuno-phenotypes of TDSCs
A better understanding of the action mechanism of the low immunogenicity of TDSCs and their suppressive effects against the infiltration of inflammatory cells are critical to promote their use for tendon repair. In this connection, the surface antigens for T-cell response and rejection were examined. Our results showed that TDSCs lacked surface expression of MHC-II, CD80, and CD86 (Fig. 6). However, TDSCs showed surface expression of MHC-I (Fig. 6). FasL and Fas have been shown to induce tolerance to promote cell survival. In this connection, our findings indicated that fixed and permeabilized TDSCs expressed FasL and Fas (Fig. 6). γ-IFN was used to simulate an inflammatory environment after TDSC transplantation in vivo. Results from our study reveal that γ-IFN pretreatment did not increase the expression of MHC-II, CD86, Fas, and FasL but slightly and strongly increased the expression of CD80 and MHC-I, respectively (Fig. 6). These results suggested that TDSCs were immune privileged.
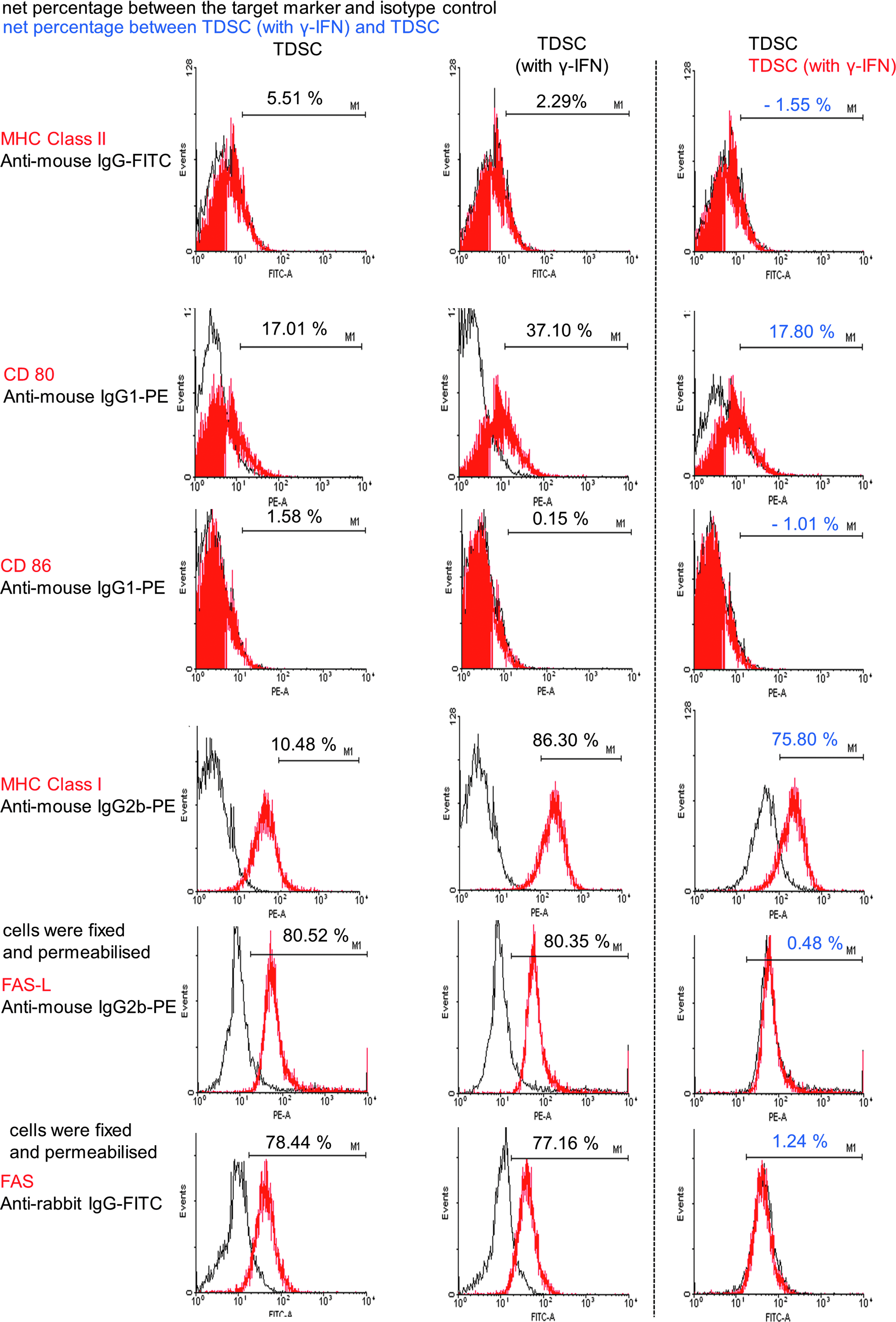
Immuno-phenotype of TDSC. Histograms showing marker expression on TDSCs with or without pretreatment with γ-IFN (red line). Black line: isotype control; percentage in black: net percentage between the target marker and the isotype control; percentage in blue: net percentage between groups with and without γ-IFN pretreatment; Representative results of three independent experiments.
Production of IDO, TNF-α, and TGF-β1 in lymphocyte-TDSC co-culture
The mechanism that TDSCs escaped from the lymphocyte attack was examined by testing the levels of soluble immuno-modulators in the supernatant of the target TDSC-lymphocyte co-culture. As indicated in Supplementary Figure S5, the expression levels of IDO, TNF-α, and TGF-β1 in the culture supernatant of the mitomycin-c-treated TDSCs and host lymphocytes after TDSCs transplantation were low. The absorbance at 450 nm from the ELISA assay was only about 0.1 (arbitrary unit) with low practical significance. Co-culture of the mitomycin-c-treated TDSCs with the lymphocytes collected at week 0 and 8 after TDSC transplantation at different ratios did not increase the production of these cytokines and enzyme. These results suggested that the mitomycin-c-treated TDSCs did not induce the lymphocytes to release the immunosuppressive mediators IDO and TGF-β1, nor suppressed the immuno-stimulatory mediator TNF-α.
Discussion
This was part II of a two-part study to examine the healing effects, healing mechanisms, and immunogenicity of TDSCs after allogeneic transplantation. The immunogenicity and escape mechanisms of rat patellar TDSCs were examined in this study.
Data from the previous in vitro cell culture model showed that mesenchymal stem cells (MSCs) suppressed the proliferation/function of naïve T-3–10 and B-lymphocytes. 24 These observations were used as a mechanism to explain how the MSCs avoid immuno-rejection. In our study, we addressed the immunogenicity of TDSCs with a better design using lymphocytes and serum collected from the animals with transplantation of TDSCs. Our results showed that target allogeneic TDSCs elicited a weak lymphocyte proliferation and they could escape from the lymphocyte-mediated cytotoxicity. Furthermore, TDSCs suppressed the PMA- and ionomycin-induced lymphocyte proliferation and the effect was TDSC dose-dependent. Moreover, incubation of the target TDSCs with serum collected from the immunized animals did not induce antibody recognition and CDC. Our result also showed that while anti-CD73 recognized TDSCs as shown by flow cytometry (result not shown), no significant complement-dependent TDSC lysis was observed. This was consistent with two recent reports that MSCs could inhibit complement activation.25,26 The production of factor H 26 and the absence of HLA-B 25 were suggested to protect MSC from complement-dependent cell lysis.
Several different mechanisms have been proposed to account for MSC-mediated suppression of lymphocyte proliferation. These include cell cycle arrest of activated T-cells by inhibition of cyclin D2 expression, 27 induction of regulatory T-cells (Treg), 1 interference of antigen presenting cells maturation, 28 and inhibition of cytotoxic T-cell formation. 29 A previous study demonstrated that Treg significantly increased in mixed lymphocytes reaction when MSCs were present compared with that in the controls. 1 There was increase in the generation of Treg in the spleens of mice after oral administration of Lactobacillus planarum WCFS1, which contained teichoic acid, a probiotic. 30 We did not analyze the proportion of different lymphocyte subsets in the spleen and in vitro co-culture. Whether the increase in Treg might account for the transient increase in spleen weight after TDSC transplantation needs further research. Future studies should assess the proportion of different lymphocyte subsets in the spleen after allogeneic TDSC transplantation and in vitro co-culture.
Our results showed that TDSCs expressed MHC-I. However, TDSCs lacked the expression of surface markers such as MHC-II, CD80, and CD86 that are required for T-cell activation and transplant rejection. Our data corroborated with the previous findings in BMSCs,7,31,32 umbilical cord blood-derived MSCs (UCB-MSCs)3,10 and adipose tissue-derived MSCs. 9 The expression of MHC-I by MSCs might protect the cells from natural killer cells-mediated cytotoxicity. 33 On the other hand, the absence of MHC-II, CD80, and CD86 might allow the MSCs to escape from recognition by allo-reactive T-cells. In the absence of co-stimulation, T-cell engagement might result in anergy that contributes to tolerance rather than allogeneic responses. However, another study showed that the immunosuppressive effect of MSCs was independent of MHC expression.6,10 Similar cases might occur in the TDSCs. Also, the expression of Fas and FasL in target cell was found to be one of the protective mechanisms that led to immuno-unresponsiveness to promote target cell survival.16–20 Data in our study did indicate that TDSCs expressed intracellular Fas and FasL.
Contrary to the findings from BMSCs7,23,31 and UCB-MSCs 23 that γ-IFN increased the expression of MHC-II and co-stimulatory molecules, γ-IFN pretreatment did not increase the expression of MHC-II, CD86, Fas, and FasL but slightly and strongly increased the expression of CD80 and MHC-I, respectively, in TDSCs. The discrepancies were likely to be due to different types and isolation protocols of stem cells. In fact, some studies showed that γ-IFN treatment did not reverse the failure of MSC to elicit a proliferative response when co-cultured with allogeneic peripheral blood mononuclear cells 32 and did not ablate the suppressive effect of MSC on alloantigen-driven lymphocyte proliferation. 34 Moreover, other study showed that γ-IFN-treated MSC promoted immunosuppressive capacity of MSCs. 35 γ-IFN pretreatment of MSCs was reported to upregulate the expression of immunosuppressive cytokines such as IDO.34–36
Our results showed that IDO, TNF-α, and TGF-β1 were not involved in the escape mechanisms of target TDSCs from lymphocyte attack. This was in contrast to the previous studies, which reported that BMSCs promoted immunosuppression by producing suppressive molecules such as TGF-β 5 and IDO, 36 and reducing the secretion of inflammatory molecule TNF-α. 2 Further study is required to elucidate the mechanisms by which TDSCs escaped from lymphocyte attack.
Conclusions
In conclusion, allogeneic TDSCs exhibited low immunogenicity. Allogeneic TDSCs might be used for transplantation.
Footnotes
Acknowledgments
The authors would like to thank Dr. Yun Feng Rui for preparing the TDSC culture for immunophenotyping and Mr. Qi Tan for assisting in the animal work. This work was supported by equipment/resources donated by the Hong Kong Jockey Club Charities Trust and the General Research Fund (project number: 471411) of the University Grant Council.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.