
Abstract
Cell-based therapy has been studied as an attractive strategy for therapeutic angiogenesis. However, obtaining a stable vascular structure remains a challenge due to the poor interaction of transplanted cells with native tissue and the difficulty in selecting the optimal cell source. In this study, we developed a cell patch of cocultured human umbilical vein endothelial cells (HUVECs) and smooth muscle cells (SMCs) using thermosensitive hydrogels for regeneration of mature vasculatures. In vitro characterization of HUVECs in the cocultured group revealed the formation of a mesh-like morphology over 5 days of culture. Vascular endothelial growth factor expression was also upregulated in the cocultured group compared with HUVECs only. The cell patch seeded with HUVECs, SMCs, or both cell type was prepared on the synthetic thermosensitive and cell interactive hydrogels, and readily detached from the hydrogel within 10 min by expansion of the hydrogel when the temperature was decreased to 4°C. We then investigated the therapeutic effect of the cell patch using a hind limb ischemic model of an athymic mouse. Overall, the group that received a cell patch of cocultured HUVECs and SMCs had a significantly retarded rate of necrosis with a significant increase in the number of arterioles and capillaries for 4 weeks compared with the groups transplanted with only HUVECs or SMCs. Dual staining of smooth muscle alpha actin and human nuclear antigen showed that the implanted cell patch was partially involved in vessel formation. In summary, the simple transplantation of a cocultured cell patch using a hydrogel system could enhance therapeutic angiogenesis through the regeneration of matured vascular structures.
Introduction
P
Cell-based therapy has been viewed as one of the most promising approaches for revascularization of damaged blood vessel within ischemic tissues. 5 Conventionally, cells are isolated from different sources, expanded in vitro, and subsequently injected as a suspension via intramuscular or intra-arterial routes.6,7 Despite ongoing arguments regarding the ideal cell type and appropriate number of cells, this method has been proven to be effective for therapeutic angiogenesis in vivo and in clinical study. 8 However, use of enzymes to detach cells from culture plates disrupts cell/ECM and cell/cell integration, which causes low retention rates (<10%) and reduced engraftment efficiency of transplanted cells. 1 Cell sheet engineering is an alternative way to overcome these limitations. The harvested cell sheet with intact cell–ECM and cell–cell junctions has been reported to enhance the cellular function and therapeutic efficacy after transplantation. 9 Previously, our group developed a temperature-sensitive Tetronic®-tyramine hydrogel system as a platform to harvest the cell sheet.10,11 Specifically, cells are confluently cultured on thermally expandable and cell-interactive hydrogels, and a monolayer cell sheet of myoblasts can then be harvested and transferred to the target site by a change in temperature from 37 to 4°C.12,13
During the endogenously stimulated angiogenic processes following ischemic injury, endothelial cells (ECs) and endothelial progenitor cells (EPCs) play a vital role in neo-vessel formation. EPCs are involved in the secretion of preangiogenic factors to promote sprouting of new vessels. They are also differentiated into endothelial lineages for vessel formation and recruitment of hematopoietic stem cells and mesenchymal stem cells from bone marrow to ischemic sites to promote angiogenesis.5,14–16
In addition, smooth muscle cells (SMCs) have been reported to serve an important role in the maturation of new blood vessels. SMCs are one of the major cellular constituents of the blood vessel wall and are responsible for expression of contractile ECM proteins, maintenance of mechanical integrity, and signaling through ion channels. Within blood vessels, ECs, and SMCs are divided by basement membrane but interact closely with each other via soluble mediators such as platelet-derived growth factor (PDGF) and transforming growth factor beta. 17 More importantly, SMCs are involved in repair and wound healing processes via enhancement of their proliferation, migration, and ECM production in case of vascular injury with a critical role in the maturation of newly formed blood vessels.18,19 When ECs and SMCs were cultured within the proximity of each other, there was a significant change in the gene expression pattern and morphology.20–22 However, no study using both cell types together has been performed from the perspective of developing angiogenic therapeutics in combination with cell sheet technology.
In this study, we hypothesized that (1) coculture of ECs and SMCs may modulate expression of proangiogenic proteins and thus, (2) delivery of cocultured ECs and SMCs as a cell sheet (herein referred to as a cell patch) to ischemic tissue could enhance the angiogenic effect and retard ischemia-mediated tissue loss. To test our hypotheses, we first investigated the effect of cocultured ECs and SMCs on in vitro cell–cell communication, expression of vascular endothelial growth factor (VEGF), and tubule formation. We then fabricated temperature-sensitive hydrogels and optimized conditions for formation of cell patches of cocultured ECs and SMCs. Lastly, we analyzed the effect of transplanted cell patches on necrotic limb loss using a mouse hind limb ischemia model by using various immunohistological techniques.
Materials and Methods
Materials
Tetronic-tyramine was synthesized as previously described. 11 Synthetic cell-adhesive peptide (GRGDGGGGGY) was customized from AnyGen (Kwangju, Korea). EGM™-2 Bulletkit™ and SmGM™-2 BulletKit were purchased from Lonza (Allendale, NJ). Trypsin/ethylenediaminetetraacetic acid and penicillin-streptomycin (p/s) were purchased from Gibco BRL (Carlsbad, CA). Phosphate-buffered saline (PBS) was purchased from Wisent (St. Bruno, QC, Canada). Peroxidase from horseradish (HRP), hydrogen peroxide (H2O2), biotin-conjugated goat anti-rabbit IgG, and biotin-conjugated goat anti-mouse IgG were purchased from Sigma-Aldrich (St. Louis, MO). Vybrant®DiO and DiD cell-labeling solution and rhodamine-phalloidin were purchased from Molecular Probes (Eugene, OR). Mouse anti-cluster of differentiation 31 (CD31) antibody was purchased from Cell Signaling (Danvers, MA). Fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse IgG and rhodamine-conjugated anti-rabbit IgG were purchased from Proteintech (Chicago, IL). Rabbit anti-alpha smooth muscle actin (SMA) antibody and DyLight® 488-conjugated goat anti-rabbit IgG were purchased from Abcam (Cambridge, United Kingdom). Streptavidin-FITC was purchased from ebioscience (San Diego, CA). Anti-human nuclear antibody (HNA) was purchased from Millipore (Temecula, CA). Zoletil was purchased from Virbac Laboratories (Carros, France). Rompun was purchased from Bayer (Leverkusen, Germany).
Immunofluorescence staining and real-time polymerase chain reaction
Human umbilical vein endothelial cells (HUVECs) (Lonza, Walkersville, MD) and SMCs (Lonza) were cultured in EGM-2 and SmGM-2, respectively, supplemented with 1% p/s under the standard culture conditions (37°C, 5% CO2) and used at passage numbers from four to six. HUVECs, SMCs, and mixed cells were seeded into 24-well plates at a density of 1 × 105 cells/cm2. The ratio of HUVECs to SMCs of the mixed group was 2:8. For immunofluorescence staining, 1 × 105 cells/cm2 HUVECs, SMCs, and mixed cells were seeded into chamber slides (Thermo Scientific, Rockford, IL) for 5 days. Immunofluorescence staining was performed as previously described. 13
For VEGF-A expression, total RNA from each group of cells cultured for 1 or 5 days was extracted by using Ambion®TRIzol Reagent (Life Technologies, Carlsbad, CA), and the concentration of RNA was estimated by measuring absorbance at 260 nm using a spectrophotometer (Nanodrop 2000; Thermo Scientific, Wilmington, DE). The cDNA was synthesized from 1 μg of RNA using Maxime RT PreMix kit (Intron Biotechnology, Gyeonggi-do, Korea) in a BioRad Thermocycler (Biorad Laboratories, Hercules, CA).
Real-time reverse transcription polymerase chain reaction (RT-PCR) was conducted in a StepOnePlus™ instrument (Life Technologies). Two microliters of cDNA solution (1/2 diluted) was mixed with 10 μL of SYBR® Premix Ex Taq™ (2×) (TliRNaseH Plus) (Takara, Japan), 0.4 μL of 10 pmol primer pairs, 0.4 μL of ROX reference dye (50×), and 6.8 μL of DEPC-treated water. The amplification reaction was performed with denaturation at 95°C for 10 min, followed by annealing at 95°C for 15 s, and extension at 60°C for 1 min for 40 cycles. The melting curve stage was performed from 60.0°C to 95.0°C in increments of 0.5°C per 5 s. All reactions were conducted in triplicate. The sequences of the primers utilized were as follows: GAPDH (Fw: 5′-CAA GGC TGT GGG CAA GCT-3′, Rv: 5′-GCA AGG CCA TGC CAG TGA-3′) and VEGF-AA (Fw: 5′–TCA CAG GTA CAG GGA TGA GGA CAC–3′, Rv: 5′-CAA AGC ACA GCA ATG TCC TGA AG–3′). The expression level of VEGF-A was analyzed by the comparative Ct method and presented as fold-change relative to Ct value of the human housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH).
Analysis of VEGF secretion
VEGF enzyme-linked immunosorbent assay (ELISA) was conducted to detect VEGF secreted from HUVECs, SMCs, and mixed cells. About 5 × 104 cells/cm2 of HUVECs, SMCs, and mixed cells were cultured in EGM-2. The cultured medium of each group was collected 1, 3, and 5 days after incubation and was analyzed by using a Human VEGF ELISA kit (Boster; Pleasanton, CA) according to the manufacturer's instructions.
Preparation and characterization of thermoresponsive hydrogels with RGD peptide
The hydrogels were prepared according to the method described in a previous study. 11 Briefly, we prepared two separate 8% (w/v) Tetronic-tyramine polymer solutions: (1) PBS with H2O2 0.075% (w/v) and (2) PBS with HRP (0.0375 mg/mL) and RGD peptide (2 mg/mL). Each of the polymer solutions was loaded into a dual syringe and injected into the space, which was lined with a Teflon mold (0.5 mm in thickness), between glass plates. After waiting 15 min to allow complete gelation, hydrogels were punched into round shapes (8 mm in diameter) and then washed several times with PBS.
To measure the change in size of the hydrogels, we prepared them with PBS at temperatures of 37°C, 20°C, 10°C, and 4°C and the diameters of the hydrogel discs were measured by using vernier calipers after being maintained in each temperature of PBS for 1 h. To determine the kinetics, hydrogel discs were moved from 37°C to 20°C, 10°C, and 4°C and the diameters were measured every 2 min. To measure the swelling ratio, hydrogel discs were completely dried and maintained in each temperature of PBS for 30 min. The weights of hydrogel disks were measured and compared to the reference disk. The swelling ratio of the hydrogel was measured by the relative swelling of hydrogel in PBS maintained at each temperature for 30 min compared to dried hydrogel.
Preparation of cell patch on the hydrogel and characterization of cell viability and f-actin staining of transferred cell patch
Hydrogel disks were immersed in 70% EtOH for 5 min, and completely rinsed with PBS before performing experiments with cells. HUVECs and SMCs were cultured, and enzymatically lifted. The same density of HUVECs, SMCs, and mixed-cell condition (SMCs:HUVECs −8:2) was seeded onto the hydrogel discs and cultured for 12 h in EGM-2 under the standard culture conditions (37°C, 5% CO2). When cell patch was formed on the hydrogel, the hydrogel disc with a confluent monolayer of each cell type was turned upside down and attached to cover glass, incubated at 4°C for 5 or 10 min, and then the hydrogel disc was removed. The viability of the transferred cell patch was confirmed by the LIVE/DEAD Viability/Cytotoxicity Kit (Molecular Probes) and images were captured with a fluorescence microscope (TE-2000; Nikon Corp., Tokyo, Japan). The relative cell viability was calculated by normalizing the number of live cells to the total number of cells (viability % = live cells/[live cells + dead cells] ×100).
The cell patches transferred onto chamber slides were cultured for 2 days, fixed at each time point (day 2 and 3) with 4% paraformaldehyde for 20 min, and permeabilized with cytoskeleton buffer (pH 6.8, 50 mM NaCl, 150 mM sucrose, 3 mM MgCl2, 50 mM Tris-base, and 0.5% Triton X-100) for 20 min. The samples were then stained with rhodamine phalloidin and counterstained with DAPI. Images were captured using a confocal microscope (Eclipse Ti; Nikon Corp.).
Distribution of SMCs and HUVECs on the patch
To track the distribution of HUVECs and SMCs on the patch, cells were prelabeled with VybrantDiO and DiD cell-labeling solution, respectively. Both cell types were enzymatically lifted and then mixed cells were seeded onto the hydrogel disc. After incubating for 12 h, the cell patch was transferred onto the cover glass and further incubated for 2 and 3 days. The distribution of cells in the cell patch was observed through a confocal microscope and images were captured at 0 h, 2, and 3 days after transfer printing.
Hind limb ischemia model and transplantation of cell patch
Nude mice (BklNbt:BALB/c/nu/nu, female; Nara Biotech, Seoul, Korea) were maintained and used under the guidelines of the Institutional Animal Care and Use Committee (IACUC) of Hanyang University (2014–0029). The hind limb ischemia mice were prepared as previously described. 23 Five-week-old mice were anesthetized with Zoletil, Rompun, and saline solution 9:1:40 (v/v/v). The right femoral artery was ligated and excised from the femoral artery distal to the inguinal ligament. The mice were randomly distributed into four groups (n = 5 for each group). Untreated mice were grouped as the negative control. Mice in treated groups had patches of HUVECs, SMCs, or mixed cells transferred to the target site by placement of a hydrogel surface having a cell monolayer facing the target site by allowing 5 min for stabilization and subsequent application of prechilled normal saline (4°C) on the hydrogel for 10 min. Thereafter, the hydrogel was carefully peeled off, leaving the cell layer on the target site. The mice were monitored and photographed at different time intervals of 3, 5, 7, 14, 21, and 28 days to record the status of the hind limb ischemia. Hind limb ischemia status was quantified as (1) limb loss, (2) foot necrosis, and (3) limb salvage. After 4 weeks, mice underwent a surgical procedure for histological analysis of the limb.
Histological analysis
The tissue from the ischemic limb muscles were retrieved after 4 weeks of treatment and embedded in paraffin. Tissue sections of approximately 4 μm thickness were prepared by using a rotary Microtome (Thermo Scientific). The tissue specimens were stained with hematoxylin and eosin (H&E) to analyze muscle degeneration. Masson's trichrome (MT) staining was performed to determine tissue fibrosis. Tissue fibrosis was quantified by MT staining images by randomly selecting five images from each group using Image J software by calculating the ratio of muscle (red) to fibrosis (blue).
Immunohistochemical analyses were also performed to measure the arteriole and capillary formation. The sectioned samples of each group were deparaffinized, hydrated, and gently washed thrice in TBS-Triton X-100. After washing, the tissue specimens were treated with blocking buffer at 37°C for 1 h. Then, samples were treated with primary antibody rabbit anti-SMA (1:100; Abcam) or mouse anti-CD31 (1:50; Cell Signaling) overnight at 4°C. Subsequently, the samples were treated with secondary antibody, biotin-conjugated goat anti-rabbit IgG or biotin-conjugated goat anti-mouse IgG (1:200; Sigma-Aldrich), for 1.5 h at 37°C followed by application of streptavidin-FITC for 2 h at 37°C. Finally, all samples were counterstained with DAPI. Fluorescent images of each group were captured through a fluorescent microscope. The number of arterioles and capillaries were counted from five images that were randomly selected from each group and quantified using NIS-elements AR software.
We also performed double staining with HNA antibody (Millipore) and anti-SMA antibody for the mixed cells group to evaluate the role of the transplanted cells within newly formed vessels. The same procedure was employed for double staining as was performed for SMA and CD31. The primary antibodies of anti-human nuclei (1:50) and anti-SMA (1:100) were used while secondary antibodies included FITC-conjugated anti-mouse (1:100) and rhodamine-conjugated anti-rabbit (1:100) and were counterstained with DAPI. Fluorescent images were captured by a fluorescence microscope.
Statistical analysis
Quantitative data were obtained from triplicate samples. Data were expressed as the mean ± standard deviation. Statistical analysis was performed using one-way analysis of variance (ANOVA) with Tukey's honest significant difference test and two-way ANOVA (for two variables) with Bonferroni post-test using Graphpad Prism 5 software (La Jolla, CA). p < 0.05 were considered as statistically significant.
Results
Characteristics of cocultured cells
First, we examined the effect of coculture on cell/cell communication and growth factor expression. As shown in Figure 1A, B, no staining for SMA and CD31 for 5 days of in vitro culture was observed in the HUVECs-only and SMCs-only groups, respectively, while the mixed group demonstrated clear staining for both proteins. Interestingly, the mixed group showed a mesh-like structure connected with each other at day 5 with clear positive staining for CD31. In contrast, HUVECs showed dispersed staining of CD31 with no distinct pattern.
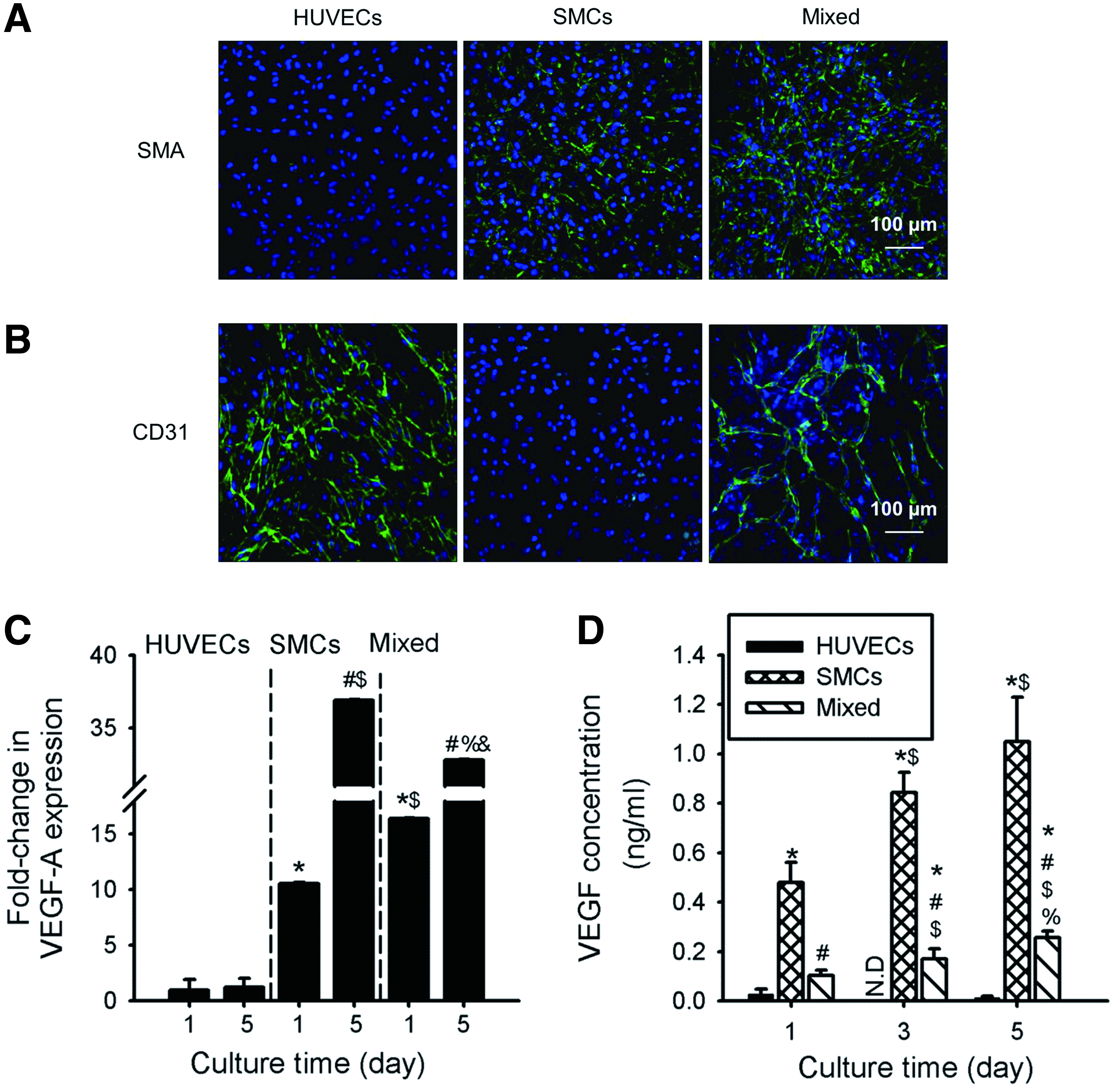
In vitro cell–cell interactions and vascular endothelial growth factor (VEGF) expression of cocultured cells. Immunofluorescent images of cells cultured for 5 days stained for
Figure 1C shows the expression of VEGF-A measured using quantitative reverse transcription polymerase chain reaction (qRT-PCR). We observed a significant increase in the expression of VEGF-A in the mixed culture (16.2-fold) and SMCs (10.6-fold) as compared to HUVECs at day 1. The trend at day 5 was the same as at day 1, however, the expression of VEGF-A was greater in SMCs (37.0-fold) than in the mixed culture (32.9-fold). ELISA was then carried out to measure the secretion of VEGF from cells into the medium. The results revealed an increasing trend of VEGF concentration with time in the SMCs (480.5 ± 81.4 pg/mL–1050.5 ± 179.9 pg/mL) and mixed (103.9 ± 22.1 pg/mL to 256.1 ± 26.6 pg/mL) groups, rather than in HUVECs, with the concentration of VEGF being the greater in the SMCs group than in the mixed group (Fig. 1D).
The effect of coculture on modulation of tubule formation assay was further carried out. As shown in Supplementary Figure S1 (Supplementary Data are available online at www.liebertpub.com/tea), the formation of an extended tubular network was observed in SMCs and in mixed cells for over 5 days, while the tubular structure in HUVECs was degenerated quickly. Quantitative analysis revealed that the tube length in SMCs (8.3 ± 2.5 mm) and mixed group (9.0 ± 1.3 mm) was significantly greater than that in the (−) control and the HUVECs group at day 3. Although a slight reduction in overall tube length was found in all groups after 5 days, the trend was similar to that seen at day 3.
Thermoresponsive characteristics of hydrogels
We next analyzed the thermosensitive characteristics of hydrogels. As shown in Figure 2, hydrogels expanded in response to the temperature change from 37°C to 4°C. Specifically, the relative size of the hydrogel increased by 1.28 ± 0.02, 1.36 ± 0.03, and 1.39 ± 0.04-fold when the temperature was cooled down from 37°C to 20°C, 10°C, and 4°C, respectively. The temperature-dependent expansion was repeatedly achieved for multiple times in which the hydrogel reattained its original size when the temperature was warmed to 37°C (Fig. 2A, B). The time-dependent response of the hydrogel to the reduced temperature demonstrated that the expansion occurred rapidly, within 10 min, and reached a plateau after this stimulation. The characterization of swelling of hydrogels at given temperatures reconfirmed the same trend; the relative swelling ratios were 4.1 ± 0.9, 7.6 ± 1.7, 9.1 ± 2.1, and 10.1 ± 2.3-fold, at 37°C, 20°C, 10°C, and 4°C, respectively (Fig. 2D).
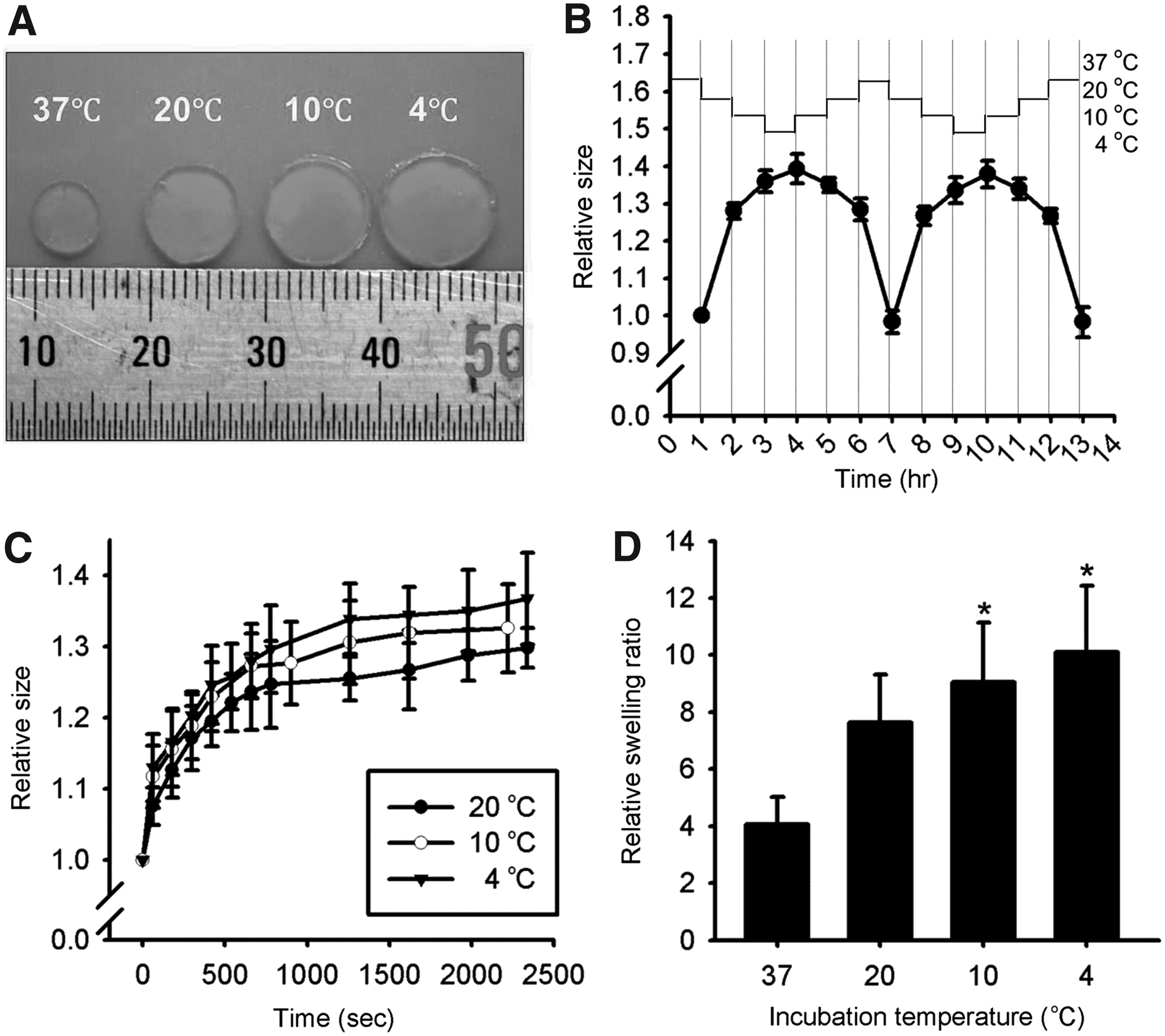
Characteristics of hydrogels.
Preparation of cell patch on the hydrogel and efficient cell patch transfer
To make cell patches on thermally expandable hydrogels, HUVECs, SMCs, and mixed cells were seeded on the hydrogels at a density of 1 × 105 cells/cm2. Under this seeding density, both SMCs and mixed cells were able to attain a monolayer although the HUVECs group appeared to require a greater number of cells for them to be confluently occupied on the hydrogel due to its relatively smaller size compared with SMCs (Supplementary Fig. S2).
We next examined the efficacy of transfer of a cell patch from the hydrogel to a target site and cell viability after the transfer process. The cell patch was transferred to the model surface of a coverglass, and the surface of the hydrogel was observed by a phase contrast microscope following the transfer. It was clear that the confluently formed cell patch was completely transferred to the coverglass after 10 min and there were no remaining cells on the surface of the hydrogel (Fig. 3). A LIVE/DEAD assay showed that the morphology of the cells was maintained after the transfer, and dead signals were scarcely found. It was found that within 5 min time, the viability of HUVECs (80.1% ± 3.1%) was significantly less than that of SMCs (93.4% ± 1.6%) and the mixed (92.7% ± 2.4%) group. However, a longer transfer time (10 min) improved the viability of the HUVECs group, making it similar to those of the SMCs and mixed groups.
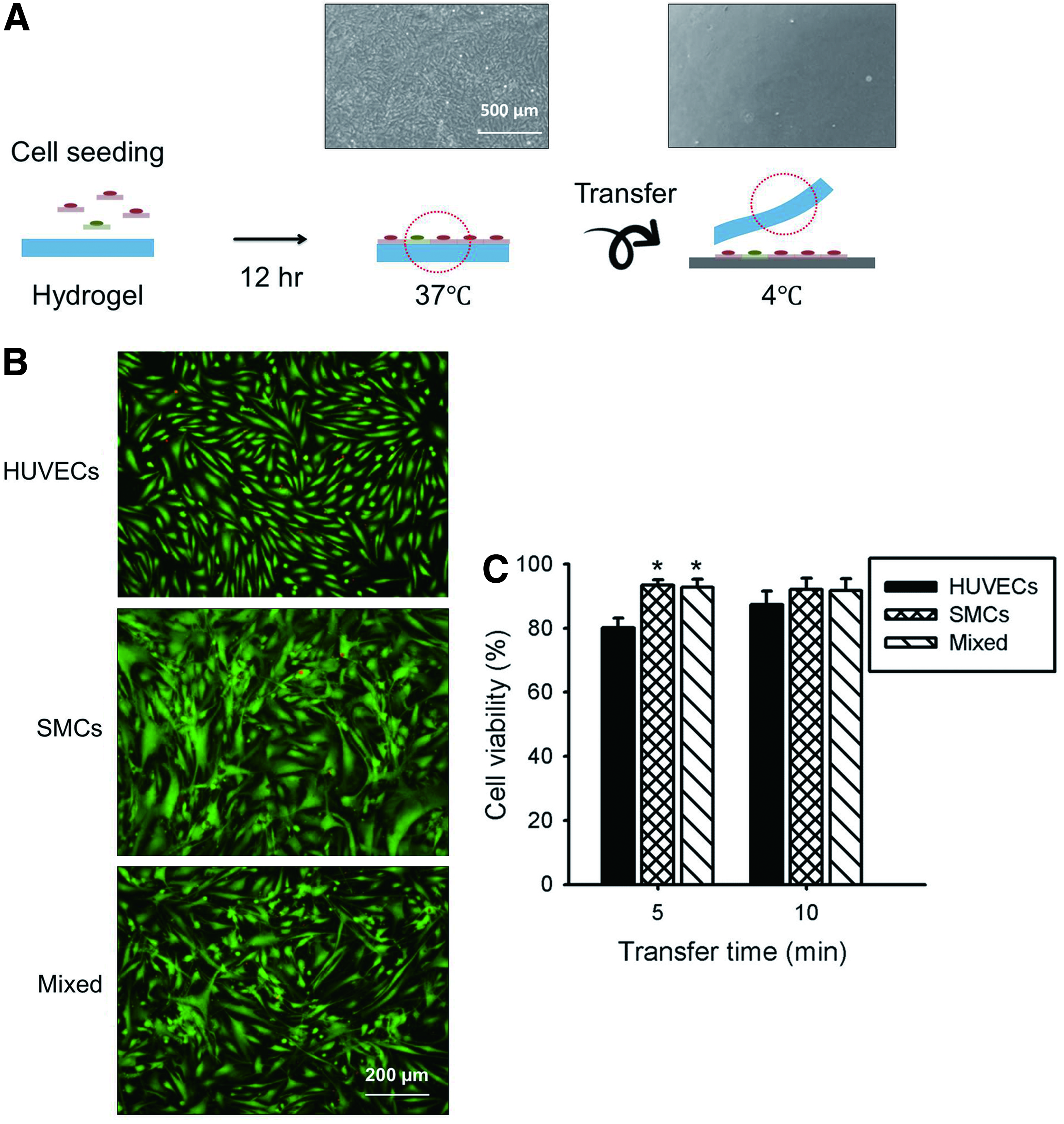
Transfer of cell patch using the hydrogel. The surface of the hydrogel
The F-actin staining of cell patches after 2 days of culture showed robust stress fiber formation with dense and fine texture on all the groups while more elaborated F-actin structure was observed in the mixed group (Fig. 4A). The morphology of the mixture of SMCs and HUVECs appeared to be similar to that of SMCs. In contrast, F-actin staining of the HUVECs patch exhibited a typical cobblestone shape. Nonetheless, these results demonstrate that cell morphology was maintained during the detachment and reattachment process during the transfer of the cell patch from the hydrogel to the target coverglass. We also observed the distribution of individual cell populations in the mixed group by prelabeling HUVECs and SMCs with DiO (green) and DiD (red), respectively. As shown in Supplementary Figure S3, HUVECs seeded at a lower number were homogeneously distributed throughout the cell patch over time. Interestingly, a distinguishable alteration in the morphology of HUVECs was found in cell patches from the mixed group in which HUVECs presented an elongated and stretched morphology of a tube-like structure (Fig. 4B).
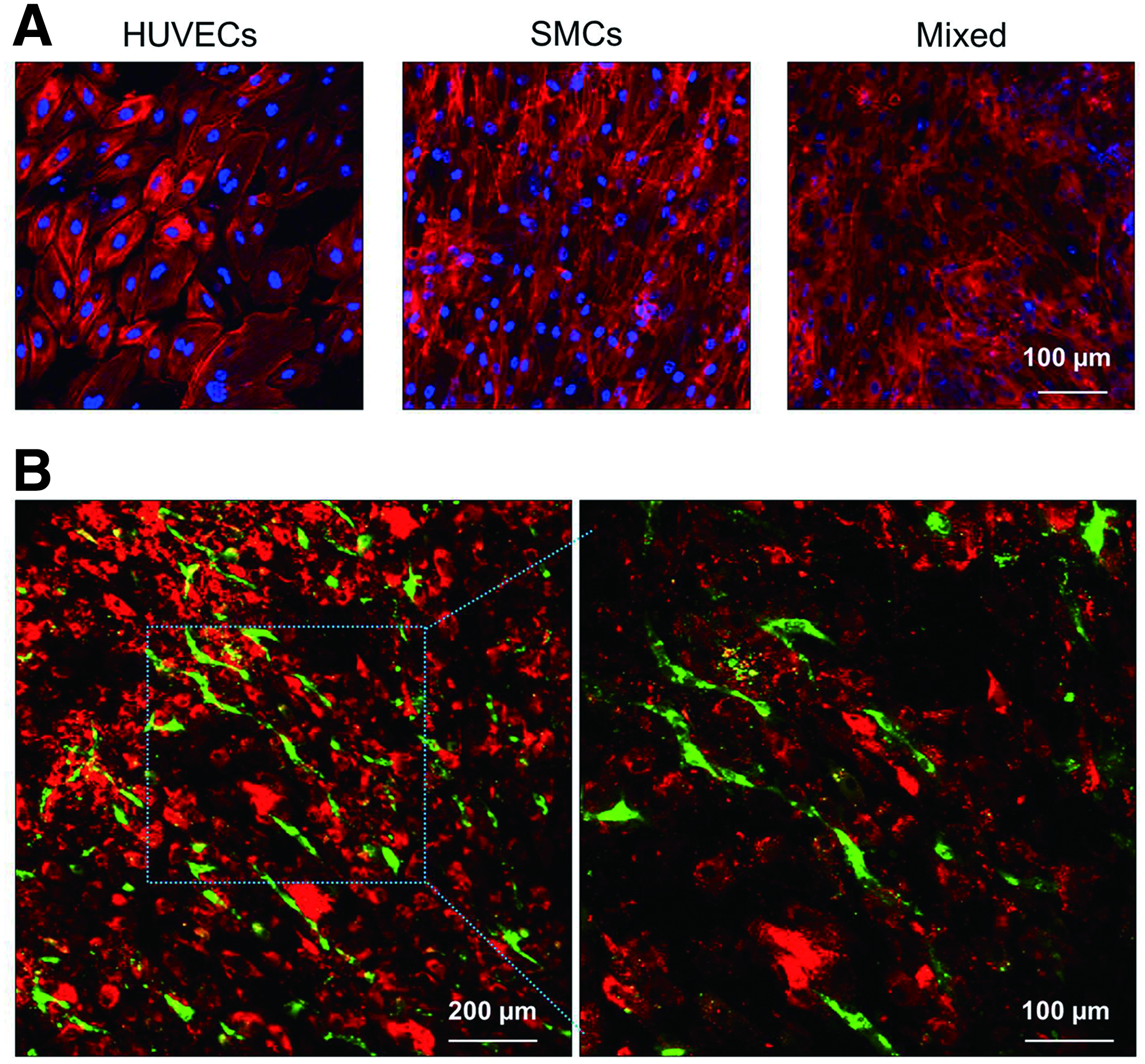
Structure of cells in cell patches after transfer
Effect of co-delivery of HUVECs and SMCs on retardation of necrotic ischemic limb loss
To evaluate the effects of a mixed cell patch on therapeutic angiogenesis, hind limb mouse models were prepared and the severity of muscle necrosis was recorded for 28 days as shown in Figure 5. The physiological status in hind limb showed that necrosis in mice that received a mixed-group cell patch was significantly retarded compared with the other groups. There was no difference among all groups until day 3. while at day 7, the percentage of limb salvage in mice transplanted with HUVECs and SMCs was reduced to 25.0% and 27.3%, respectively, which was significantly lower than that of the mixed group (40.0%) (data not shown). In addition, there was no further limb loss and necrotic change in the mice transplanted with mixed cells for over 28 days, while the percentages of limb loss in mice receiving only HUVECs and SMCs was found to increase from 25.0% and 27.3% at day 7 to 75.0% and 63.6% at day 28, respectively. Another notable finding was that the group that showed the second longest retardation in limb necrosis at day 28 was the group of mice transplanted with the SMCs patch.
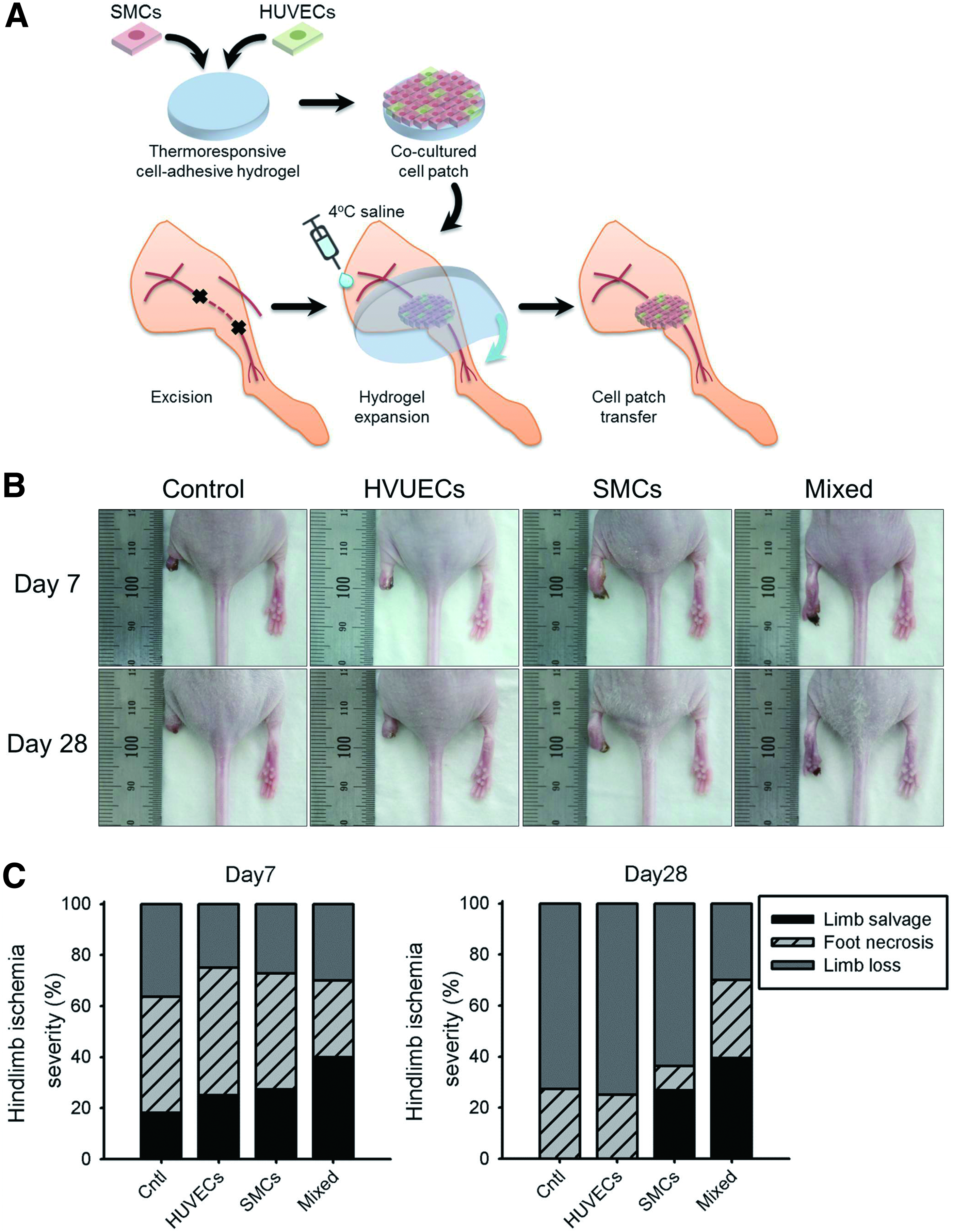
Therapeutic effect of cell patch.
To confirm the intergration of transplanted cell patch we deliver a fluorescently prelabbeled cell patch to hind limb of mouse. As shown in Supplementary Figure S4, the cell suspension group were observed for only 5 days while our cell patch group signals were observed until 10 days. The tissues from ischemic hind limb muscles were retrieved after 28 days of treatment to use for histological analysis. H&E staining showed muscle degeneration was higher in the control groups than those in the HUVECs, SMCs, and mixed groups (Fig. 6A). The percentage of fibrosis (blue) analyzed from MT staining was significantly reduced in the mixed group, as shown in Figure 6B. For example, fibrosis was reduced from 22.6% ± 8.2% for (−) control to 14.5% ± 2.8%, 11.8% ± 4.5%, and 5.3% ± 1.8% for the HUVECs, SMCs, and mixed groups, respectively.
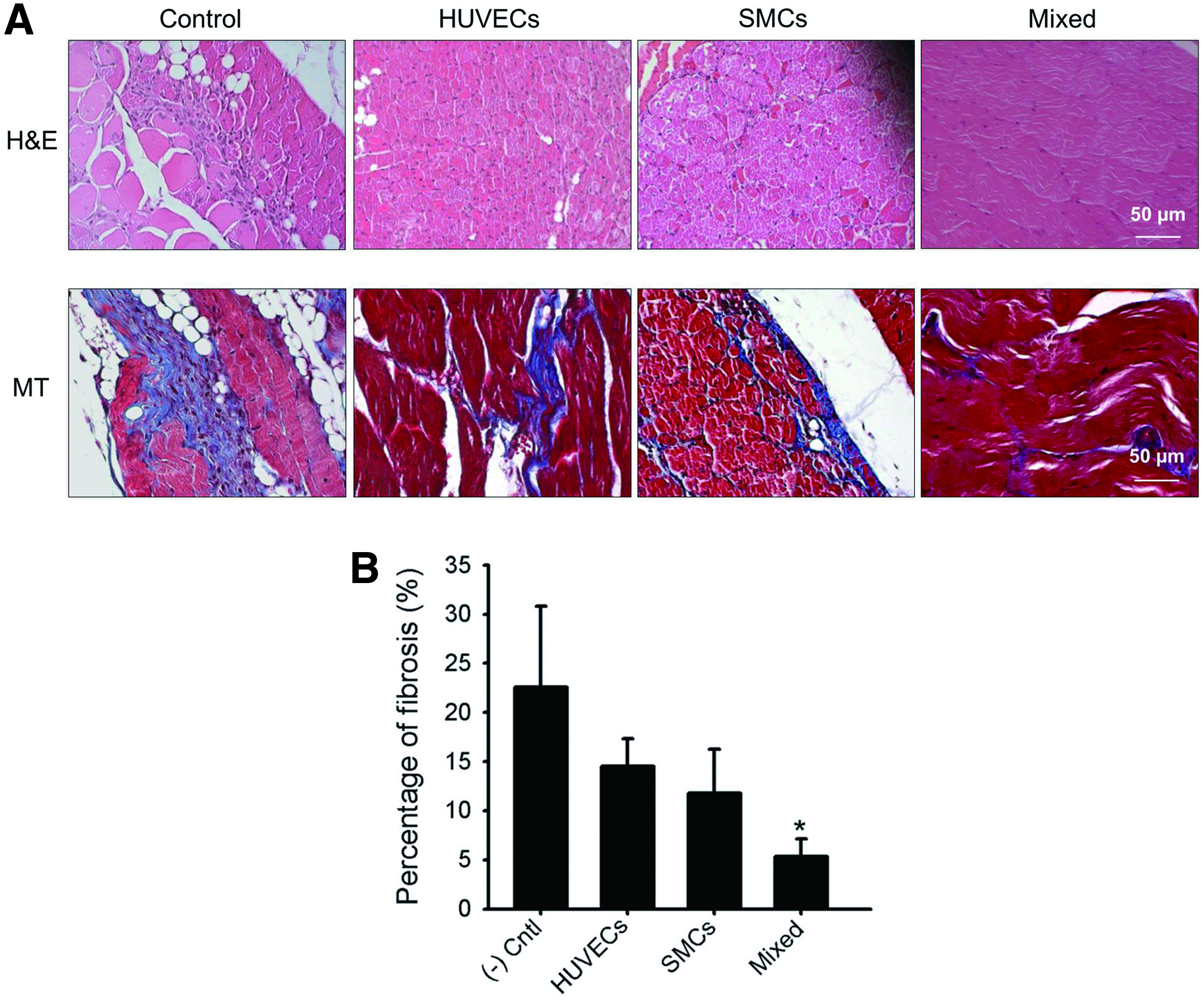
Reduced inflammation and fibrosis.
We next performed immunohistochemistry for SMA (a marker of arterioles) and CD31 (a marker of capillaries) to evaluate the therapeutic angiogenesis achieved by cell patches after 28 days of implantation. Consistent with the physiological status of hind limb, the number of arterioles was significantly greater in the mixed group (7.2 ± 1.1 arterioles/mm2) than in the control (3.2 ± 0.7 arterioles/mm2), HUVECs (4.3 ± 1.1 arterioles/mm2), and SMCs groups (5.7 ± 1.4 arterioles/mm2) (Fig. 7A, B). When the number of capillaries was evaluated, the mixed group was again found to have a significantly greater amount (92.2 ± 14.4 capillaries/mm2) than groups of other experimental conditions (32.3 ± 11.1 capillaries/mm2, 55.0 ± 6.8 capillaries/mm2, and 73.2 ± 8.4 capillaries/mm2 for the control, HUVECs, and SMCs groups, respectively) (Fig. 7C, D). To confirm the involvement of implanted cells in arteriole formation, the specimen was double-stained for human nuclei and SMA. As shown in Figure 7E, we found that both signals coexisted in newly formed blood vessel, implying that cells within the implanted cell patch were partially involved in vessel formation.
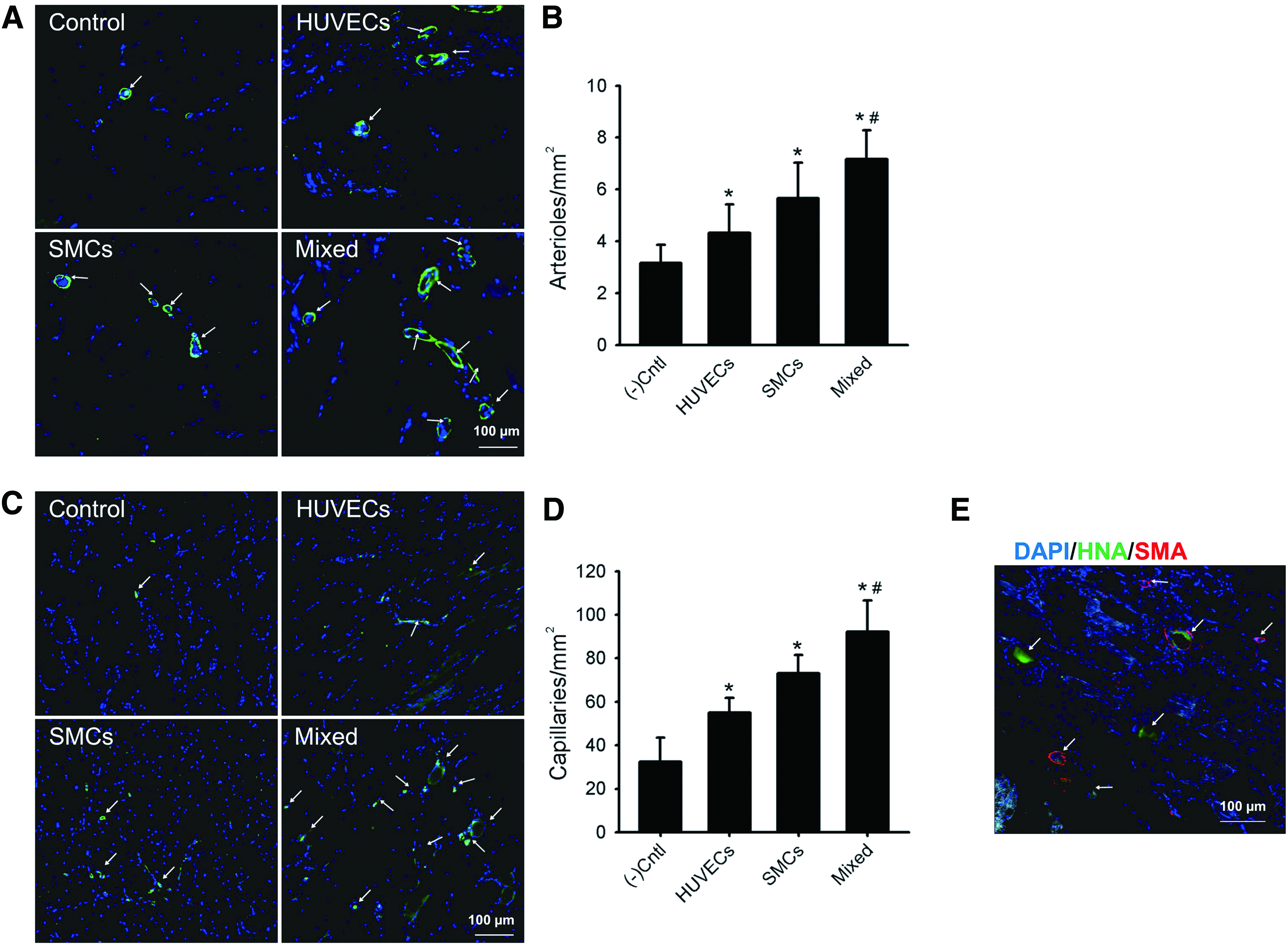
Effect of cell patches on neovessel formation.
Discussion
PAD is a common circulatory disease caused by atherosclerotic occlusion of the arteries to limbs. One of the major risks of PAD is ageing. Over one-third of patients have typical claudication. The severity of PAD is closely associated with the risk of ischemic stroke and myocardial infarction.24,25 An approach involving administration of various types of cells, such as ECs and stem cells, has been developed for therapeutic angiogenesis.26,27 Here, we propose using a cell patch of cocultured HUVECs and SMCs that can be delivered to the ischemic lesion by a thermally expandable hydrogel, and we investigated its effect on the angiogenic process and retardation of ischemic necrosis.
Interactions between ECs and SMCs are known to be essential for formation and maintenance of arterial structure. 12 Co-engraftment of HUVECs and SMCs in a PGA scaffold supported with collagen/fibronectin protein gels accelerated, stabilized, and promoted remodeling of tissue-engineered microvessels. 28 In addition, it was reported that in vitro coculture of E20Cs and SMCs influenced the expression of many growth factors implicated in angiogenesis, such as basic fibroblast growth factor (bFGF) and PDGF-BB at the mRNA and protein levels. 17 We observed CD31-positive staining with polygonal (mesh-like) morphology and no tubule formation in HUVECs cultured alone, while a spontaneous self-assembled tubular network formed by positively stained HUVECs was clearly found under coculture with SMCs, as shown in Figure 1B. These results suggest that the presence of SMCs accelerates tubular structure formation of HUVECs.
Additionally, we found significantly greater gene expression of VEGF-A in the mixed group after 1 day, compared with HUVECs alone, which is consistent with the findings of a previous study showing that coculture of ECs and SMCs exhibited significant gene expression of VEGF for up to 72 h.17,29 Upregulation and downregulation of some proteins or signaling factors in coculture are dependent on culture time, which may be due to stimulation of inhibitors secreted by each cell type reversely affecting the expression of growth factors in later culture times. 17 Our results demonstrated that the expression level of VEGF mRNA in SMCs became greater than in other groups at day 5, indicating that interactions between two cell types are a temporally variable process.
We found that SMCs and mixed cells secreted significantly higher levels of VEGF in media than did HUVECs. However, the secretion of VEGF was also significantly higher in the SMCs group than in the mixed cells. This pattern has been observed before.17,29 Regarding the tubule formation assay, SMCs and mixed cells showed significantly greater tube length than did the control and HUVECs group after 5 days, supporting our hypothesis that coculture of ECs and SMCs would modulate expression of proangiogenic proteins and resulting tubule formation. It has been reported that cocultures of ECs and SMCs secreted significantly higher levels of bFGF, PDGF-BB, and other proangiogenic cytokines while VEGF secretion was significantly lower, compared with the SMCs-only group.17,30 Therefore, this study supports our mixed group matrigel analysis data, in which the reduced secretion of VEGF into media may have been compensated for by high levels of bFGF and PDGF-BB secretion for tubule formation and enhanced angiogenesis.
Various attempts have been performed involving co-injection of different types of cells for neovascularization in ischemia. Combined injection of human embryonic stem cell-derived ECs and mural cells or early endothelial progenitors and late outgrowth ECs augmented therapeutic angiogensis in a hind limb ischemia model.31,32 However, cell delivery through injection is limited due to low retention rate and viability, which often necessitates large amounts of cells for transplantation to achieve a therapeutic outcome. 1 Problems associated with retention and viability of transplanted cells can be addressed by transplantation of cell aggregates or a cell sheet. Injectable cell aggregates showed better retention, viability, and enhanced vessel formation in a hind limb ischemia model.27,33,34 However, fabrication and maintenance of uniformly sized spheroids remains challenging. 35
Cell sheet engineering has enabled the construction of uniformly sized monolayers of therapeutic cells for both single and multiple cell types. 36 Although numerous works have been published on the employment of this approach in functional recovery of damaged tissue, construction of cell sheets on a PNIPAAm-grafted surface has several limitations. Adhesion and reversible detachment of the cell sheet is sensitive to the density and thickness of PNIPAAm-grafted chains and the process requires expensive and complicated facilities for PNIPAAm grafting. 37
Our system to harvest cell patches is based on the use of thermally responsive hydrogels in situ polymerized with cell-adhesive peptides that can rapidly expand when the temperature is reduced from 37°C to 4°C. As a result, binding between the cells and cell adhesive peptides is disrupted while cells maintain their cell–cell junctions and are harvested as a cell sheet. Unlike PNIPAAm-grafted culture dishes, our hydrogel can be easily handled and manipulated to the target site for transplantation of a cell sheet. We have developed hydrogel-based cell patch harvest techniques for many cell types including fibroblasts, ECs, and myoblasts.13,38,39 To prepare a cell patch, it is important to control the interactions between hydrogels and target cells. Our results demonstrated that hydrogels with the RGD peptide sequence incorporated had improved cell attachment and allowed formation of a stable cell monolayer on the hydrogel. Previously, RGD peptide has been incorporated into many types of hydrogels prepared from modified alginate and PEG monoacrylate hydrogel for enhanced cell adhesion of HUVECs and SMCs.40,41
We confirmed the viability of the cell patch after transfer to the model substrate after 10 min at 4°C. The cell viability was very high although cells were exposed to a low temperature. Low exposure time is indeed beneficial for minimizing the detrimental effects on cells. However, too short a time could potentially damage the cell patch since the attachment to the target may not yet be stably formed, which would lead to mechanical disruption of the cell-hydrogel binding. Nonetheless, cell patches were successfully transferred to the target within a relatively short time and therefore, we believe that the function of the cells and cell–cell interactions were also preserved without any adverse effects. The observation of matured F-actin structure also supports these results, as shown in Figure 4A. The mixed group showed maintenance of the F-actin structure of HUVECs and SMCs after transfer. In addition, prelabeled HUVECs exhibited a unique morphology and distribution in coculture that was not observed when HUVECs were cultured alone. These characteristics of HUVECs were also preserved after transfer. Therefore, we expect that HUVECs in a mixed group may participate in neo-vessel formation.
Overall, the in vivo ischemia model and its histological studies showed a significant increase in vessel formation and less fibrotic response in the group transplanted with cell patches of mixed HUVECs and SMCs. Previous studies confirmed cell therapy as a promising approach for therapeutic angiogenesis as transplanted cells may either secrete angiogenic cytokines to stimulate endogenous vessel forming cells or directly participate in vessel regeneration.4,42,43 A variety of cell sources including ECs, EPCs, hematopoietic cells, hMSCs, and SMCs have been transplanted and reported to be involved in secretion of angiogenic cytokines and improvement of tissue perfusion, thereby resulting in improved angiogenesis.5,14,15,44,45 Although ECs are one of the major constituents of blood vessels, the vascular network produced by implantation of ECs was not stable for a long time. For stable vascular network formation, functional interactions between ECs and mural cells, such as vascular SMCs, are critical during vascular development.46–50 For that reason, we selected coculture of HUVECs with SMCs for making a stable vascular network after transplanting to the ischemia model. Our results suggest that the presence of SMCs contributes synergistically to the retardation of necrotic injury of ischemic tissue.
For successful angiogenesis, it is well known that a relatively large number of cells must be injected to compensate for the low engraftment efficiency and reduced survival of transplanted cells due to anoikis. For example, it was reported that the significant enhancement of therapeutic angiogenesis through cell injection required over 5 × 106 cells per mouse. 1 In this study, it should be noted that the total number of cells occupying a patch for transplantation was 0.5 × 104 cells per mouse, which was found to be sufficient to retard necrosis and to enhance angiogenesis. We also evaluated the retention of our cell patch as compared to injection of cell suspension (Supplementary Fig. S4). Our cell patch group signals were pertained over 10 days while cell suspension group lost its signals after 5–7 days.
We then examined the role of SMCs in these observed effects. As demonstrated from our in vitro study data, it would be anticipated that SMCs may contribute to paracrine effects by increasing expression of angiogenic factors. In addition, dual staining of the specimen with antibodies for HNA and SMA revealed that transplanted cells from the mixed cell patch were integrated into native tissues and were found to be involved in the generation of mature blood vessels. Collectively, our results demonstrated that mixed cells in patches not only exerted a paracrine effect, but were also directly involved in the maturation of neo-vessels, a finding that is consistent with those of a previous study. 51 Nonetheless, a detailed study on the role of implanted cells in angiogenesis will be necessary in the future to examine this process with a more molecular biological approach.
Conclusions
In this study, we developed cell patches of cocultured HUVECs and SMCs for use in therapeutic angiogenesis. In vitro characterization of HUVECs from the cocultured group revealed distinct cell–cell interactions and exhibition of a unique interconnected mesh-like morphology, which was absent when HUVECs were cultured alone. In addition, the conditioned media collected from the cocultured group more significantly promoted an extended network of tubules relative to other groups during an in vitro tubule formation assay. Furthermore, we fabricated transferrable mixed cell patches on the thermoresponsive hydrogel incorporating the RGD peptide. The cell patches were simply transferred to a target substrate by expansion of the hydrogel in response to a low temperature (4°C) without any adverse effect. Moreover, we confirmed that mixed-cell patches maintained their unique interconnected tubule-like morphology of HUVECs following transfer. Finally, we evaluated the therapeutic angiogenesis achieved with the mixed-cell patch when transplanted in a hind limb ischemia mouse model. Over 4 weeks of postsurgery observation, we found that tissue necrosis was significantly retarded in the group receiving the cocultured cell patch. We found that our transplanted cocultured cell patches enhanced neovascularization with evidence of the transplanted cells being partially incorporated within newly formed vessels. We propose that our hydrogel-based platform for delivery of cocultured cell patches could hold great promise for regeneration of matured vascular structures, which have previously been limited in cell-based therapies.
Footnotes
Acknowledgments
This work was supported by a grant of National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT & Future Planning (NRF-2013R1A2A2A03067809) and by the Technology Innovation Program (10050526). Development of disposable diaper based on biomass-oriented biodegradable super absorbent polymers, funded by the Ministry of Trade, Industry & Energy.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.