
Abstract
Currently, autografts still represent the gold standard treatment for the repair of large bone defects. However, these are associated with donor-site morbidity and increased pain, cost, and recovery time. The ideal therapy would use biomaterials combined with bone growth factors to induce and instruct bone defect repair without the need to harvest patient tissue. In this line, bone morphogenetic proteins (BMPs) have been the most extensively used agents for clinical bone repair, but at supraphysiological doses that are not without risk. Because of the need to eliminate the risks of BMP2 use in vivo, we assessed the ability of three putative osteogenic factors, nel-like molecule type 1 (NELL-1), high mobility group box 1 (HMGB1), and CCN2, to enhance the essential processes for bone defect repair in vitro and compared them to BMP2. Although it has been reported that NELL-1, HMGB1, and CCN2 play a role in bone formation, less is known about the contribution of these proteins to the different events involved, such as cell migration, osteogenesis, and vasculogenesis. In this study, we investigated the effects of different doses of NELL-1, HMGB, CCN2, and BMP2 on these three processes as a model for the recruitment and differentiation of resident cells in the in vivo bone defect repair situation, using cells of human origin. Our data demonstrated that NELL-1, HMGB1, and CCN2 significantly induced mesenchymal stem cell migration (from 1.58-fold increase compared to control), but BMP2 did not. Interestingly, only BMP2 increased osteogenesis in marrow stromal cells, whereas it inhibited osteogenesis in preosteoblasts. Moreover, the four proteins studied promoted significantly endothelial cell migration, reaching a maximum of 2.4-fold increase compared to control, and induced formation of tube-like structures. NELL-1, HMGB1, and CCN2 had these effects at relatively low doses compared to BMP2. This work indicates that NELL-1, HMGB1, and CCN2 might enhance bone defect healing via the recruitment of endogenous cells and induction of vascularization and act via different processes than BMP2.
Introduction
C
Bone morphogenetic protein 2 (BMP2) and BMP7 are the most studied bone forming proteins. Moreover, currently BMP2 is the only BMP approved as a bone graft substitute. 4 However, despite the therapeutic potential of BMP2 in bone repair, large doses (up to 40 mg) are needed to produce a significant osteogenic effect, 5 and this can result in undesired ectopic bone formation, soft tissue swelling, bone resorption, and tumor growth enhancement through angiogenesis stimulation.6–8
Accordingly, alternative proteins are being assessed for their osteogenic induction capacity. Three of such proteins are nel-like molecule type 1 (NELL-1), connective tissue growth factor (also known as CCN2), and high mobility group box 1 (HMGB1). Bone regeneration is a complex process that involves a series of well-orchestrated biological events. At the cellular level, migration, proliferation, angiogenesis, osteogenic differentiation, and subsequent mineralization are essential processes to enable bone formation and repair to occur. 9 To optimize in vivo bone defect repair using biologicals, more knowledge of the effect of these factors on the different processes is necessary.
NELL-1 is a secretory protein and its overexpression induces altered bone formation and it is considered an essential factor for human craniosynostosis. 10 It has been shown to promote bone formation on mice in vivo,11,12 and in a comparison study, NELL-1-based bone grafts were comparable to BMP2-based grafts in rat spinal fusion. 13 Recent studies performed on osteoporotic mice and sheep have shown that NELL-1 could potentially be used as a therapy for osteoporotic bone loss.14,15 Furthermore, NELL-1 has been shown to increase osteogenic differentiation in vitro. 16 However, whether NELL-1 is able to recruit osteoprogenitor cells is not known yet. Moreover, several genes affected by NELL-1 appear to promote angiogenesis at early stages of bone regeneration, 17 but whether it is able to promote endothelial cell (EC) recruitment and differentiation is still unknown.
CCN2 has been used mostly in rats and has been shown to promote bone and cartilage regeneration.18,19 It is one of the six CCN matricellular proteins characterized by their conserved modular structure 20 and interacts with integrins, enhancing cellular processes such as adhesion, extracellular matrix (ECM) synthesis, cytoskeleton reorganization, and survival.21–23 In vitro studies have shown that overexpressing cells with CCN2 promote mesenchymal cell proliferation, migration, and aggregation.24,25 In addition, it has been shown that CCN2 expression was increased in mesenchymal stem cells (MSCs) and osteoblasts during formation of new bone, suggesting its role in bone development, 20 but its effect when added to either human osteoprogenitor cells or osteoblasts has not been investigated yet. Furthermore, CCN2 is a necessary mediator for the production of vascular basement membranes, 26 however, its effect on angiogenesis remains unclear.27–30
HMGB1, unlike NELL-1 and CCN2, has never been used as growth factor-based therapy, but knowledge about the potency of this factor in the different processes involved in bone repair makes it an attractive candidate. HMGB1 is a chromatin protein mainly known due to its role as an alarmin.31,32 Consequently, many studies showed that HMGB1 is a potent proinflammatory cytokine secreted by monocytes and macrophages able to regulate migration of mesangioblasts, smooth muscle cells, or myoblasts among others.33–36 Recently, it has been shown that HMGB1 has an osteomodulatory action, 37 being chemotactic to MSCs 38 as well as osteoblasts and osteoclasts during endochondral ossification.31,39 Although HMGB1 is known to be able to upregulate osteogenic markers when added to MSC culture in vitro, 40 its direct effect on osteoprogenitor and preosteoblast mineralization has not been studied. In addition, various studies have identified it as a putative proangiogenic factor and its overexpression is related with an increased ability to develop blood vessels.34,41,42
NELL-1, HMGB1, and CCN2 are indicated to play a role in bone formation. However, their effect on major cell types and specific processes involved in bone formation is barely studied, and they were never directly compared. The main goal of this study is to compare side by side NELL-1, HMGB1, and CCN2 to BMP2 and assess whether they are able to induce cell migration, osteogenic differentiation, and neovascularization, indispensable processes needed for bone formation. This was performed using cells from human origin: two types of osteoprogenitors (human MSCs and fetal osteoblasts) and human umbilical vein endothelial cells (HUVECs).
Materials and Methods
Cell culture
Human MSCs were obtained from leftover material from iliac crest biopsies of four donors (age 9–12 years; three males and one female) undergoing cleft palate reconstruction surgery (Erasmus MC Ethics Committee number MEC-2014-106) with implicit consent. Cells were seeded at a density of 50,000 cells/cm2 in α-MEM (Gibco, BRL), supplemented with 10% fetal calf serum (FCS), 1 ng/mL fibroblast growth factor (FGF2), 25 μg/mL ascorbic acid-2-phosphate, 1.5 μg/mL fungizone, and 50 μg/mL gentamicin. After 24 h, nonadherent cells were washed out and adherent cells were expanded in the abovementioned medium. The medium was renewed twice per week until MSCs neared confluency. Then, cells were used for migration or osteogenic differentiation assays.
Simian virus-immortalized human fetal osteoblast (SV-HFO) cells 43 were expanded and cultured as previously described. 44
HUVECs (Lonza) were cultured at a density of 5000 cells/cm2 in endothelial growth medium (EGM-2 with SingleQuots; Lonza). Medium was renewed every 2–3 days. When cells neared confluency, they were used for migration or tube formation assays.
Migration assay
MSC and HUVEC migration was assessed using modified Boyden chambers (polyethylene terephthalate cell culture inserts, pore size: 8 μm in diameter, Millipore-Merck). Briefly, to analyze MSC migration, α-MEM containing NELL-1 (0.07–3.5 nM [10–500 ng/mL], R&D Systems), CCN2 (0.89–8.9 nM [10–100 ng/mL]; Abnova), HMGB1 (0.4–4 nM [10–100 ng/mL]; R&D Systems), and BMP2 (0.38–38 nM [10–1000 ng/mL]; kindly provided by Dr. Joachim Nickel, Fraunhofer IGB) was added to the lower chamber of a 24-well plate. Platelet-derived growth factor-AB (PDGF-AB) (20 ng/mL) or 10% FCS was used as positive controls. 6 × 103 MSCs suspended in a volume of 200 μL α-MEM were added into the upper chamber. The plates were incubated at 37°C for 17 h. To test the chemotactic effect of NELL-1 (10–500 ng/mL), CCN2 (10–100 ng/mL), HMGB1 (10–100 ng/mL), and BMP2 (10–1000 ng/mL) on HUVECs, the endothelial cell basal medium (EBM-2, Lonza) containing NELL-1, CCN2, HMGB1, or BMP2 was added in the lower chamber. EGM-2 was used as positive control. 5 × 104 HUVECs were added into the upper chamber. The plates were incubated at 37°C for 10 h. In both cases, the membrane was then washed and the cells remaining on the upper surface of the chambers were mechanically removed with a cotton swab. Those that had migrated to the lower surface were fixed with 4% formalin, stained with 4′,6-diamidino-2-phenylindole (DAPI) (100 ng/mL) in the dark for 5 min, and imaged using fluorescence microscopy (Zeiss Axiovert 200 M Fluorescence Imaging) in five random fields for each membrane and counted using ImageJ software.
Osteogenic differentiation
Osteogenic differentiation assays were performed on MSCs and SV-HFOs. 3000 Cells/cm2 (MSCs) or 9000 cells/cm2 (SV-HFOs) were seeded with α-MEM in a 12-well plate. For MSCs, after 24 h, the medium was replaced with the complete osteogenic medium: DMEM high glucose (Gibco) with 10% FCS, 1.5 μg/mL fungizone, 50 μg/mL gentamicin, 1 ng/mL FGF2, 25 μg/mL ascorbic acid-2-phosphate, 10 mM β-glycerophosphate, and 0.1 μM dexamethasone. For SV-HFOs, the osteogenic differentiation medium consisted of phenol red-free α-MEM (Gibco), pH 7.5, supplemented with 20 mM HEPES (Sigma), streptavidin/penicillin, 1.8 mM CaCl2·2H2O (Sigma), 2% heat-inactivated charcoal-treated FCS, 0.1 μM dexamethasone, and 10 mM β-glycerophosphate. During each medium refreshment, medium was supplemented with proteins of interest: NELL-1 (10–500 ng/mL), CCN2 (10–100 ng/mL), HMGB1 (10–100 ng/mL), and BMP2 (10–1000 ng/mL). Osteogenic differentiation was carried out until onset of mineralization, monitored by measuring calcium concentration in the culture supernatant. Cells were scraped from the culture dish in phosphate-buffered saline (PBS)/0.1% Triton X-100. Cell lysates were sonicated on ice before analysis.
DNA and protein measurements
For DNA measurement, cell lysates were incubated for 30 min at 37°C with 100 μL heparin (8 IU/mL in PBS) and 50 μL RNAse A (50 μg/mL in PBS) solution. Ethidium bromide solution (25 μg/mL) was added and DNA content was measured on the Wallac 1420 Victor2 (PerkinElmer Life and Analytical Science) plate reader using an excitation filter of 340 nm and emission filter of 590 nm. For standards, calf thymus DNA (Sigma) was used.
Protein was measured in cell lysates with BCA protein assay (Pierce™ BCA protein assay; Thermo Scientific) according to the manufacturer's instructions.
Alkaline phosphatase activity
Alkaline phosphatase (ALP) activity was performed as described previously 45 by determining the release of paranitrophenol from paranitrophenyl phosphate (pNPP) in cell lysates. pNPP (20 mM) was added to each sample at 37°C. After exactly 10 min, the reaction was stopped by adding 0.06 M NaOH. Absorption was measured on the Wallac 1420 Victor2 plate reader at 405 nm. For standards, ALP (10 U/mL) from bovine kidney (Sigma) was used.
Mineralization
To quantify the calcium content, cell lysates were incubated for 48 h in 0.24 M HCl at 4°C. For analysis of calcium concentration in the culture medium, supernatant was collected from day 9 onward. In both cases, the calcium content was colorimetrically determined after addition of 1 M ethanolamine buffer (pH 10.6), 19.8 mM 8-hydroxyquinoline, and 0.35 mM O-cresolphthalein complexone, at 595 nm on the Wallac 1420 Victor2.
For von Kossa staining, cell cultures were fixed for 15 min in 4% formaldehyde, stained by 5% silver nitrate solution (Sigma; 85228) for 30 min under a 60 W light, and imaged using an inverted microscope (Olympus CKX41).
HUVEC tube formation assay
Growth factor-reduced Matrigel (Corning) was added to a 96-well plate and incubated at 37°C for 1 h. HUVECs were resuspended in EBM supplemented with NELL-1, CCN2, HMGB1, or BMP2. EGM-2 complete medium was used as positive and EBM-2 as negative control. Fifteen thousand cells were seeded on top of the Matrigel, incubated at 37°C, and imaged after 4, 6, and 24 h using an inverted microscope. Three independent experiments in triplicate were performed. The results were analyzed using ImageJ software.
Statistical analysis
Data were analyzed with IBM Statistics 21 (SPSS). A linear mixed model was applied; the different conditions (different doses of the proteins studied) were considered a fixed parameter and the donors (experiments) as a random factor. A descriptive analysis was performed to assess the normal distribution of the data. When multiple comparisons were analyzed, the Bonferroni test was performed. p < 0.05 was considered statistically significant.
Results
NELL-1, CCN2, and HMGB1 stimulated MSC migration, whereas BMP2 did not
To determine whether the proteins could recruit progenitor cells to the site of injury, we assessed their effects on migration of MSCs. The positive control PDGF-AB was chemoattractant showing a 4.6-fold increase compared to negative control (data not shown). NELL-1, CCN2, HMGB1, but not BMP2, were capable of inducing MSC migration (Fig. 1). NELL-1 increased migration 2-, 2.3-, and 2.79-fold compared to control in a dose-dependent manner, although only statistically significant (p < 0.05) with the highest dose. CCN2 significantly increased migration at the lowest and medium dose (2.75- and 2.48-fold, respectively), while the highest dose had no effect compared to control. In contrast, HMGB1 was chemotactic at the highest dose tested, 100 ng/mL, with more than fivefold increase compared to control. Although there was a trend toward a stimulation of migration of BMP2 on MSCs (1.3–1.6-fold increase), this effect was not significant.
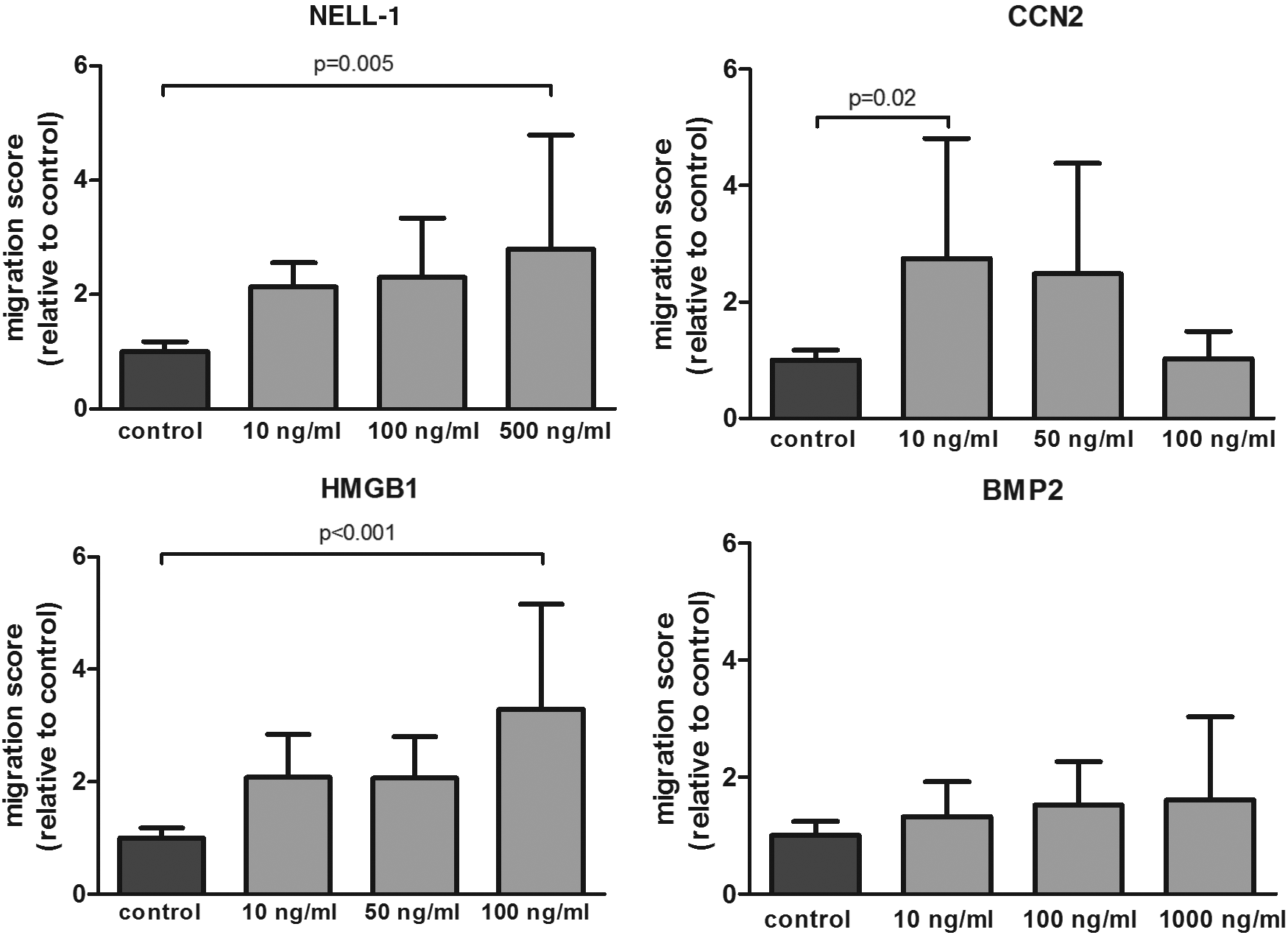
NELL-1, CCN2, and HMGB1 stimulated MSC migration. Average migration of MSCs exposed to several doses of each factor relative to the negative control (n = 4 donors in duplicate). The bars represent the mean ± SD. BMP2, bone morphogenetic protein 2; CCN2, connective tissue growth factor; HMGB1, high mobility group box 1; MSCs, mesenchymal stem cells; NELL-1, nel-like molecule type 1.
NELL-1, CCN2, HMGB1, and BMP2 proteins stimulated EC migration
We explored the role of NELL-1, CCN2, HMGB1, and BMP2 on HUVEC recruitment. The positive control (EGM-2 complete medium) showed a 3.2-fold increase in migration compared to negative control (EBM). Interestingly, similar to MSCs NELL-1 and CCN2 stimulated HUVECs to migrate. Specifically, NELL-1 increased migration in a dose-dependent manner, showing more than twofold increase with the highest dose, while CCN2 had a greater effect at the lowest dose (more than twofold increase compared to control). Although HMGB1 showed less pronounced effect on HUVEC than MSC migration, it had a significant effect on migration in all the tested doses, showing a twofold increase compared to the control. It is noteworthy that, in contrast to the other three proteins that had similar effects on both cell types with regard to dose, BMP2 showed different effects, demonstrating a significantly higher EC migration at the lowest and medium dose supplied with more than 1.5-fold increase compared to control (Fig. 2).
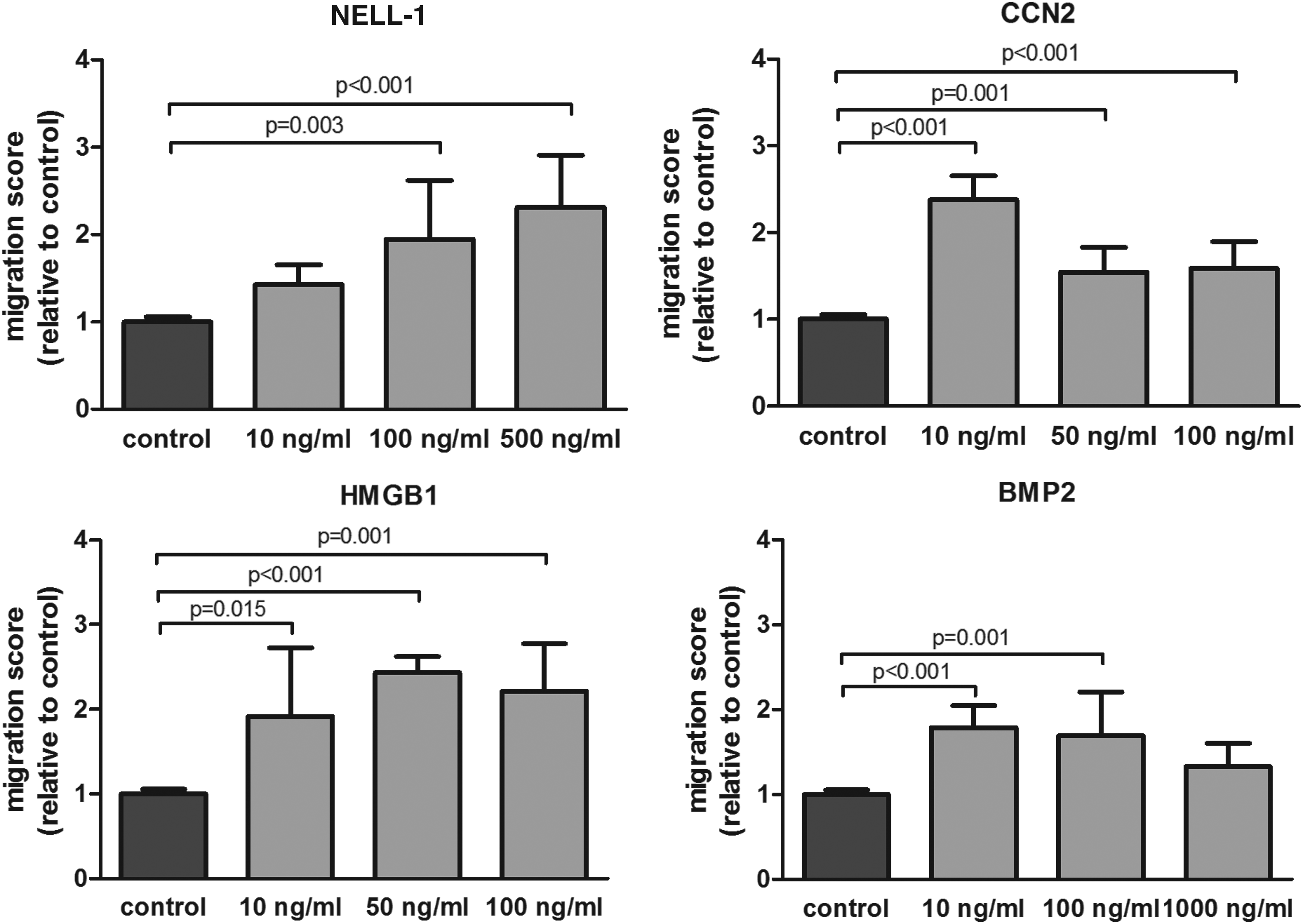
NELL-1, CCN2, HMGB1, and BMP2 stimulated EC migration. Average migration of HUVECs exposed to several doses of each factor relative to the negative control (n = 3 donors in duplicate). The bars represent the mean ± SD. HUVECs, human umbilical vein endothelial cells.
BMP2 enhanced MSC osteogenic differentiation; NELL-1, CCN2, and HMGB1 had no effect
Osteogenic differentiation of MSCs was assessed by calcium deposition. Supplementation with NELL-1, CCN2, or HMGB1 did not cause any increase or decrease in mineralization. However, when BMP2 was added, we observed a dose-dependent stimulatory effect, with a significant increase in nodule formation at the highest dose provided (Fig. 3A). The results were corroborated quantitatively by measuring the calcium content in the cell lysates, showing that addition of BMP2 at the highest dose provided (1000 ng/mL) had an almost twofold increase in mineralization compared to the standard osteogenic differentiation media (Fig. 3B).
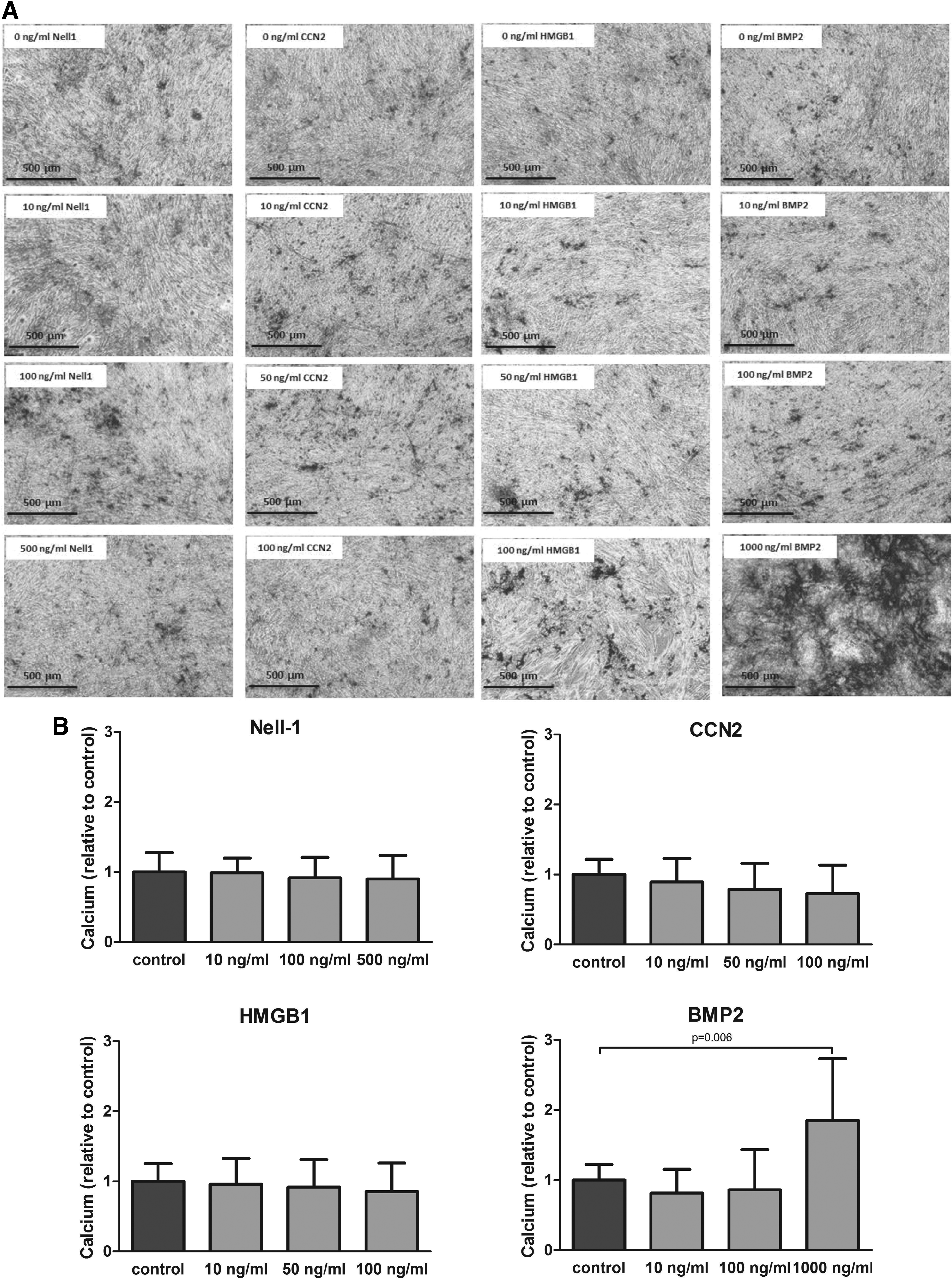
Effect of NELL-1, CCN2, HMGB1, and BMP2 on the MSC mineralization. Human MSCs were induced to mineralize in the absence or continuous presence of NELL-1, CCN2, HMGB1, or BMP2.
BMP2 inhibited preosteoblast differentiation and mineralization; NELL-1, CCN2, and HMGB1 had no effect
The addition of NELL-1, CCN2, and HMGB1 did not increase mineralization or ALP activity at either of the time points studied (Fig. 4A). Interestingly, and in contrast to the effect observed with hMSCs, the addition of BMP2 to the preosteoblast culture had a direct inhibitory effect on differentiation and mineralization. The decrease in ALP activity became significant at day 16. In addition, a reduction in the ECM calcium content at the lowest dose provided was detected (Fig. 4A, B). However, when the highest dose was provided (1 μg/mL), the inhibitory effect was no longer observed and the mineralization reached control levels.
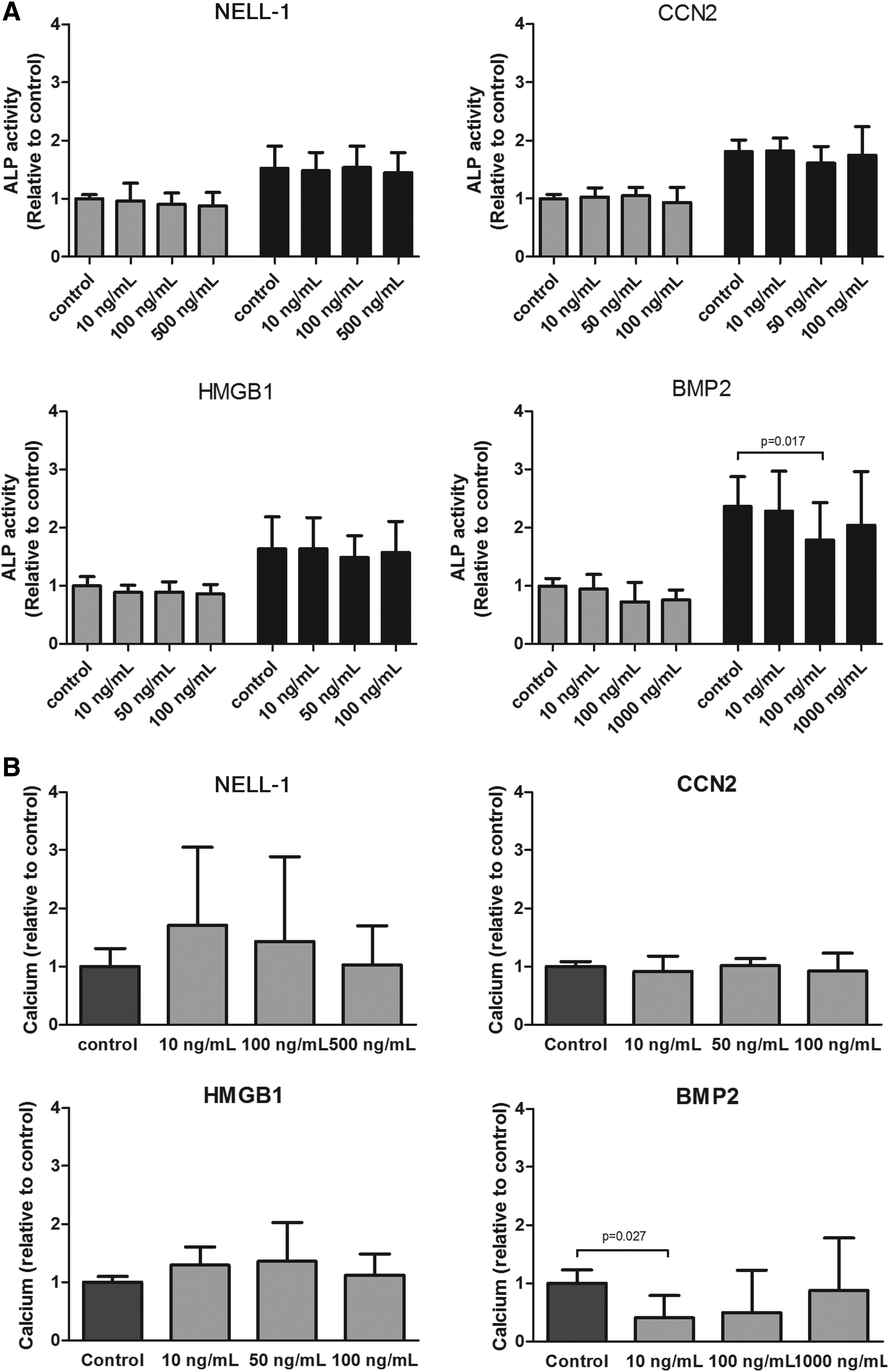
Effect of NELL-1, CCN, HMGB1, and BMP2 on the osteoblast differentiation measured by ALP activity and mineralization.
NELL-1, CCN2, HMGB1, and BMP2 stimulated neovascularization
We have determined the capability of the factors to promote formation of tubular-like structures by ECs in vitro. There was a significant improvement in tube formation at the medium dose of NELL-1 added (100 ng/mL), Furthermore, CCN2 and HMGB1 were able to enhance tube formation at the lowest doses supplied. BMP2 also enhanced tube formation in a dose-dependent manner, resulting in a significant effect on tube formation when the highest dose was added (Fig. 5B).
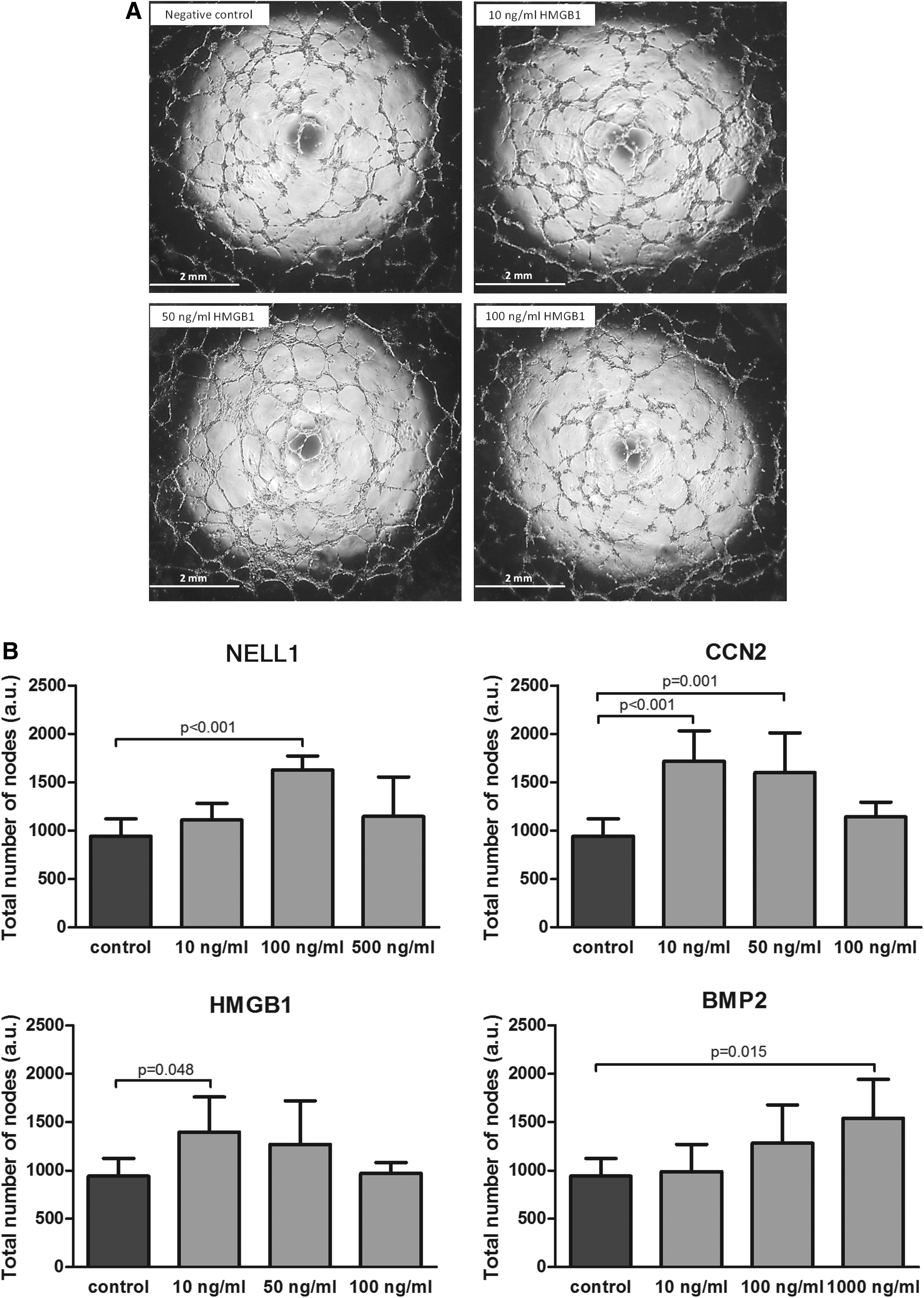
Effect of NELL-1, CCN2, HMGB1, and BMP2 HUVEC on tube formation on Matrigel.
Discussion
We directly compared four proteins that are acknowledged for their apparent ability to stimulate osteogenesis in vitro or in vivo.20,31,46,47 This is the first study to directly compare these proteins for their ability to induce recruitment, osteogenic differentiation, and neovascularization using human cells. Our results demonstrate the ability of NELL-1, CCN2, and HMGB1 to attract both MSCs and ECs even at low doses compared to BMP2. Since new blood vessel formation is crucial for bone development, we assessed neovascularization in an in vitro model, demonstrating that all four tested proteins were able to stimulate formation of tube-like structures. Interestingly, in contrast to BMP2, NELL-1, CCN2, and HMGB1 did not significantly increase the osteogenic differentiation capacity of MSCs above that induced by dexamethasone. Also of note, we observed that BMP2 negatively impacted preosteoblast mineralization, whereas the others had no effect.
Effects on cell migration
It is generally thought that BMPs are mediators of osteoprogenitor cell recruitment. However, it had been published that BMP2 had no effect in the recruitment of MSCs, neither in the cultured MSCs nor in the marrow ablation tissues. 48 In our study, we did not see any increase in MSC migration by BMP2 either. The BMP2 chemotactic effect seems to vary between EC subtypes; studies performed with ECs showed that BMP2 was not able to stimulate human aortic EC migration, 6 but it stimulated HUVEC migration. 49 Our data confirmed that BMP2 is able to stimulate migration of HUVEC when doses between 10 and 100 ng/mL are supplied.
Where BMP2 had no effect on MSC migration and less than doubled the migration of HUVEC, we observed up to a threefold increase in MSC and HUVEC migration when HMGB1, CCN2, or NELL-1 was added to the culture. HMGB1, as a signal of tissue damage, is known to be capable of inducing migration in many different cell types, including inflammatory cells.36,50,51 Fewer articles exist investigating the effects of HMGB1 on osteogenic cells, but those that do indicate it to be important in cell recruitment. Meng et al. demonstrated that HMGB1 promotes MSC migration up to 2.5-fold compared to control. 38 Our results confirm these findings, showing that HMGB1 has a highly significant chemotactic effect on MSCs above doses of 50 ng/mL. To date, there is very little reported about HMGB1s chemotaxis on primary ECs. Low doses of HMGB1 are able to promote endothelial progenitor cell migration using transwells coated with fibronectin or fibrinogen. 52 Bauer et al. have performed a wound healing migration assay and have shown that 1 μg/mL of HMGB1 is able to induce migration of HUVEC. 53 We show that HMGB1 is highly chemoattractant to HUVECs, and even a hundred times lower concentration than 1 μg/mL is sufficient to induce more than a twofold increase compared to control. In addition, from 10 to 100 ng/mL, we observed a similar chemoattractant effect, suggesting a low dose is sufficient to stimulate EC migration.
CCN2 has been shown to promote migration of human MSCs and human dermal microvascular ECs when transfected with pCCN2.24,25 Nonetheless, the effect of the addition of the protein CCN2 to MSCs or ECs had not been studied before. We demonstrated that the addition of low dose (10 ng/mL = 0.89 nM) of CCN2 to the media is sufficient to obtain a significant migratory response from MSCs and HUVECs, reaching more than a fourfold increase compared to the control. The fact that CCN2 showed an inverse dose response is potentially interesting for endogenous cell recruitment though slow release systems.
NELL-1 has been shown to inhibit renal carcinoma cell migration and adhesion. 54 However, we did not find published data about the effect of NELL-1 on MSC and EC migration. We demonstrated in this study its ability to promote migration of both cell types. In fact, it was able to promote EC migration at lower doses than BMP2 (BMP2, 100 ng/mL = 3.8 nM; NELL-1, 100 ng/mL = 0.7 nM) and MSC migration when BMP2 did not.
Effects on osteogenesis
BMP2 is the best known osteogenic factor. Next to the capacity to induce bone formation in vivo, the capacity of BMP2 to induce osteogenic differentiation of MSCs in vitro has already been tested.55,56 Our data confirm this as we observed an almost twofold increase when 1 μg/mL of BMP2 was added to the culture. However, little is known about its effect on osteoblasts. In this study, we observed an inhibition of mineralization when BMP2 was added to osteoblast cultures in a concentration range of 10–100 ng/mL. Interestingly, we did not observe that inhibitory effect when the highest dose was added (1 μg/mL). This could be due to the fact that SV-HFOs express activin, 57 which may interfere with BMP2 or simply because BMP2 is not an essential stimulus when the cells are already committed to the osteogenic pathway. Many studies indicated HMGB1's role in the induction of endochondral ossification in vivo and that it may act as a bone resorption signal. 31 However, HMGB1 has not been used as a therapeutic agent for bone regeneration and whether it has any effect on either osteoprogenitor or osteoblast differentiation and mineralization is barely studied. We did not observe any effect on osteogenic differentiation when HMGB1 was added to the MSC or preosteoblast culture media. Although it has been shown that CCN2 expression is increased in MSCs and osteoblasts during bone formation, 20 the effect of the addition of the protein CCN2 to MSCs or to osteoblasts had not been examined previously. Our study demonstrates that while it promotes migration of osteoprogenitor cells, CCN2 is not able to increase osteogenesis in contrast to a published study by Safadi et al., 58 who used rat primary osteoblasts and showed an increase on calcium deposition when 100 ng/mL of CCN2 was added to the culture beginning at day 11. We cannot exclude that the differences in experimental setup, including addition of dexamethasone and single dosing of CCN2, might explain the results.
With regard to NELL-1, most of the osteogenic differentiation studies have been performed with either human osteosarcoma cell lines or murine osteoblast-like cells.59–61 These studies have shown that, when transfected with NELL-1, these cells increase late osteogenic differentiation markers and ECM mineralization. Recently, Pang et al. have identified a functional transcript of NELL-1 able to induce osteogenic differentiation of a murine MSC line. 62 However, no information on human MSCs or osteoblasts was available. We observed an osteogenic trend when NELL-1 was added at low doses to preosteoblasts, although with high variability between cultures. Even then, comparison on a molar basis suggests that NELL-1 could have an osteogenic inducing effect even at low doses, in contrast to BMP2 that exhibited the reverse effect. Although we did not observe a significant effect of NELL-1 on osteogenic differentiation, a synergistic effect of NELL-1 and BMP2 has been shown on osteogenesis in vitro. 63 Moreover, NELL-1 can suppress BMP2-induced inflammation and enhance BMP2-induced bone formation in vivo.64–66 Therefore, NELL-1 is a potentially interesting candidate to be used alone or with BMP2. NELL-1 and BMP2 might stimulate different stages of osteogenic differentiation; where NELL-1 stimulates osteoprogenitor cell migration and could enhance preosteoblast differentiation, BMP2 might promote differentiation of skeletal stem cells.
Effects on vascularization
BMP2 had been shown to stimulate angiogenesis in developing tumors.6,67 In bone tissue engineering, however, BMP2 is frequently used in combination with angiogenic factors as vascular endothelial growth factor (VEGF) or angiopoietin 1,68–70 suggesting BMP2 does not optimally induce vascularization. In this study, we demonstrated the ability of BMP2 to induce the formation of tube-like structures at a dose of 1 μg/mL, a relatively much higher dose than required for CCN2, HMGB1, or NELL-1. HMGB1 is known to have diverse roles in angiogenesis during disease and tissue repair 42 and has been suggested to induce angiogenesis through a VEGF-dependent mechanism. 71 HMGB1 was shown to induce tube formation by microvascular ECs at doses ranging from 0.1 to 1 μg/mL.72,73 In this study, we show that much lower doses of HMGB1 are sufficient to recruit ECs and to promote tube formation. CCN2 has been shown to act as a proangiogenic factor, coordinating vasculature formation during skeletal development21,74 and also as an antiangiogenic agent inhibiting VEGF-induced angiogenesis, 29 suggesting that CCN2 regulates angiogenesis through direct and indirect mechanisms. 75 We show that CCN2 has proangiogenic properties when it is added directly to ECs, even at much lower doses than are required for BMP2 to induce a similar effect. Therefore, CCN2 might also be an interesting factor to be combined in future studies with a powerful osteoinductive growth factor, due to its chemotactic and angiogenic features. Although some studies have shown that NELL-1 induces VEGF expression in human pericytes and perivascular stem cells,12,76 to our knowledge, there are no previous publications investigating the effects of NELL-1 on vascularization of ECs. We demonstrate that the addition of NELL-1 positively affects EC migration within a tight dose range, producing a significant increase at 100 ng/mL, a larger dose than needed with either HMGB1 or CCN2, but still 10 times lower than needed with BMP2.
Bone defect repair
To accelerate and optimize bone remodeling, using more than one type of growth factor involved in bone formation could be preferable to reduce possible side effects due to high doses used when single-factor therapies are applied. In addition, several studies have shown that not only the dose but also the timing of release of the proteins from the carrier used for bone formation greatly modifies the outcome. 77 Based on the promising properties of NELL-1, HMGB1, CCN2, and BMP2 with regard to their promotion of bone formation-related processes, in vivo analyses using dual release systems should be performed to assess their potency for bone defect repair.
Footnotes
Acknowledgments
The authors thank Dr. Joachim Nickel for his generous gift of BMP2. The research leading to these results has received funding from the European Union Seventh Framework Programme FP7-PEOPLE-2013-ITN under grant agreement no. 607051.
Disclosure Statement
No competing financial interests exist.