
Abstract
Background:
Platelet-rich and platelet-poor plasma (PRP and PPP) are autologous preparations from peripheral blood and contain several growth factors and cytokines involved in tissue repair. Although their neuroprotective and neuroregenerative properties have been already described, little is known about their effects in the inner ear. We, therefore, examined the effects of PRP and PPP on spiral ganglion neurons (SGN) in vitro.
Results:
For all experiments, spiral ganglia were isolated from neonatal rats and were cultured in serum-free medium. PRP from human venous blood was added to dissociated SGN. Treatment with PRP (1:10, 1:50) significantly increased the neuronal survival and the neuronal outgrowth of SGN. This effect was completely reversed by the addition of Bay 11 (nuclear factor kappa B-inhibitor) and SB203580 (p38 mitogen-activated protein kinase [p38MAPK]-inhibitor). Furthermore, PPP was used as a cell-free matrix for the attachment of spiral ganglion explants. Coating with activated PPP improved the adhesion and neurite outgrowth of spiral ganglia explants. Therefore, activated PPP is a promising alternative for poly
Conclusion:
Preparations from autologous whole blood (PRP and PPP, respectively) present an interesting alternative for pharmacological intervention to the inner ear since they contain a balanced and natural composition of trophic factors.
Background
S
The neurotrophic effect of NTF treatment can be enhanced by combined electrical stimulation in deafened guinea pigs,8–10 making NTF application together with the implantation an attractive option to enhance the performance of the cochlear implant.
Combined application of several growth factors seems more efficient to promote an increased survival effect on SGN in vitro. 11 A cocktail of different factors may be a promising approach to improve nerve–electrode interaction by neuroprotection and neurite regeneration in the inner ear. In a previous study, we have shown that recombinant brain-derived neurotrophic factor (BDNF) differs in its potency when compared with cell-derived BDNF. 12 Cell-based delivery of growth factors is more potent and could provide an adequate source of growth factors in a tempered concentration. Platelet-rich plasma (PRP) may be one option for cell-based growth factor delivery since it contains a plethora of bioactive factors important for wound healing and tissue repair. Its preparation from peripheral blood is easy and cost effective. 13
Platelets are anucleate cell elements that are built by fractionation of bone marrow megakaryocytes. Thus, they are composed of a cell membrane and contain the cytoplasm of megakaryocytes filled with granules, mitochondria, and mRNA. 14 Although they lack genomic DNA and were usually not classified as cells, recent evidence accumulates to the notion of platelets as metabolically active cells. 15 Platelet-poor plasma (PPP) also contains several growth factors, but lacks the burst release of bioactives from platelets upon their activation.
Being in clinical use for decades without any adverse events or postoperative complications,16–18 autologous plasma-derived products, such as PRP and PPP, are an attractive source of high levels of a natural composition of growth factors. Upon stimulation, platelets release the contents of alpha, dense and lysosomal granules. 19 Besides growth factors, other substances, molecules, and ions are also secreted to the surrounding tissue exerting fundamental effects on wound healing and tissue repair. 20
Autologous plasma-derived products were able to enhance regeneration of peripheral and cranial nerves as demonstrated in several experimental studies.21–25 A recent review of the literature 26 revealed that reports on the clinical use of plasma-derived products for the treatment of neuropathies are only few.27–31
Hitherto, not a single report exists on the effects of plasma-derived products on the inner ear. Thus, the effects of PRP and PPP on cultured SGN isolated from neonatal rats were investigated in the present study.
Materials and Methods
SGN were dissected from neonatal Sprague Dawley rats (postnatal day 3–5) of both sexes. This procedure was approved and registered (no. 2013/44) by the local authorities (Zentrales Tierlaboratorium, Laboratory Science, Hannover Medical School, including an institutional animal care and use committee) and is reported on a regular basis as demanded by law. The study was conducted in accordance with the German “Law on Protecting Animals” (Tierschutzgesetz) and with the European Directive 2010/63/EU for the protection of animals used for experimental purposes. No further approval is needed if no other treatment than tissue analysis is applied (German Law on Protecting Animals).
Preparation of PRP and PPP
The RegenKit-BCT1 (RegenLab, Le Mont-sur-Lausanne, Switzerland) system was used for autologous PRP and PPP preparation. The tube was filled with venous blood freshly drawn from the basilic vein of three authors (J.S., A.R., and A.W., all female). For each independent experiment, the tubes were filled with about 8 mL of venous blood. Immediate centrifugation (80-2C, RegenLab, Le Mont-sur-Lausanne, Switzerland) for 5 min at 3000 rpm led to a separation of red blood cells from the plasma which contained the mononuclear cells and platelets. Thereafter, 2.5 mL of the PPP was removed from the resulting 5 mL of the plasmatic fraction and mononuclear cells were suspended with the remaining 2.5 mL of the plasma through careful shaking.
The neuroprotective effect of PRP was investigated by adding it in different dilutions to primary SGN (Spiral ganglion cultivation). Moreover, PPP has been activated by adding calcium gluconate (2.25 mmol Ca2+; B. Braun, Melsungen, Germany) to get a gelatinous matrix for the adhesion of spiral ganglion explants. In these experiments, the activated PPP replaced the poly
Isolation of the spiral ganglion
The isolation of the cochleae was performed as described previously. 11
Spiral ganglion cultivation
Spiral ganglia were cultured after dissociation and as whole organ explants. The mechanical and enzymatical dissociation of the spiral ganglion as well as the seeding on laminin-coated 96-well plates were conducted as described previously. 11
For cultivation of spiral ganglia explants as whole organ culture, the spiral ganglion of one cochlea was dissected into five pieces of the same length. All five pieces of one cochlea were directly transferred into one well of a 24-well plate coated with activated PPP and 15 cochleae were used for each experimental set-up.
The culture medium (serum free) for SGN consisted of Panserin 401 (PAN BIOTECH, Passau, Germany) supplemented with HEPES (25 mM; Life Technologies, Carlsbad), glucose (6 mg/mL; Braun AG, Melsungen, Germany), penicillin (30 U/mL; Grünenthal GmbH, Aachen), N2-supplement (3 mL/mL; Life Technologies, Carlsbad), and insulin (5 mg/mL; Sigma-Aldrich, St. Louis).
Dissociated cells were maintained at 37°C, 5% CO2 in a humidified atmosphere for 48 h. The SGN were cultivated in a 1:1 mixture of serum-free SGN medium and PRP at four different dilutions (1:4; 1:10; 1:50; 1:100 in serum-free SGN medium) or BDNF (recombinant human BDNF; Invitrogen, Karlsruhe, Germany) in a concentration of 50 ng/mL or without any factors as negative control (NC). Each experimental condition and control was set up in triplets and each experiment was repeated thrice resulting in a total n = 9.
The explants were cultivated for 72 h at 37°C and 5% CO2. At a dilution of 1:2, 1:4, and 1:10, PPP was used as coating of the well after activation with calcium gluconate as described in Preparation of PRP and PPP. Cultivation of explants in uncoated wells served as control. In addition, BDNF at a concentration of 50 ng/mL was added to the PPP coating as further control (1:2 + BDNF). Each experimental condition and control was set up in triplets and each experiment was repeated thrice resulting in a total, n = 9.
The seeding control was fixed after 4 h of incubation in serum-free medium. Fixation of all cells has been performed with a 1:1 mixture of methanol (Roth, Karlsruhe, Germany)/acetone (J.T. Baker, Deventer, Netherlands) for 10 min. For storage or further analysis, cells were washed three times with phosphate buffered saline.
Immunohistochemistry and evaluation
The spiral ganglion contains different cell types, for example, neurons, glial cells, and fibroblasts and the term spiral ganglion cells refers to all these three cell types. To identify neurons in the dissociated mixed spiral ganglion cell culture and in the spiral ganglion explants, methanol/acetone-fixed cells as well as explants were stained immunochemically for the heavy chain 200 kD neurofilament (NF) as described in detail previously using the VECTASTAIN® Elite® ABC Kit (Vector Laboratories, Burlingame) and peroxidase diaminobenzidine substrate (the Peroxidase Substrate Kit DAB; Vector Laboratories, Burlingame). 11 After the neuron-specific staining, the SGN were quantified in the dissociated cell culture using an inverted microscope (Olympus CKX41, Hamburg, Germany) coupled with a CCD-camera (Colorview XS, SIS; Olympus, Münster, Germany) and were analyzed using CellP (SIS; Olympus).
Surviving neurons were defined as 200 kD neurofilament-positive cells with neurites of at least three times the soma diameter in length. The number of surviving neurons in each experimental group is given as percentage of the average number of neurons in the seeding control. To evaluate neuroregenerative effects of the PRP treatment, five neurons per well were measured (neurite length and soma diameter). For analysis of the whole organ cultures (spiral ganglion explants), the number of adhered explant pieces per well was determined. The neurite outgrowth of the spiral ganglion explants was evaluated by counting all neurites sprouting out of each cultivated and attached piece of the SGN explants.
Inhibition of PRP with selected inhibitors
To verify over which intracellular pathway the observed effect of PRP was mediated, five selected inhibitors were used:
• AG490 (#658401—5 mg; Calbiochem, Merck Millipore) inhibiting Janus kinase 2 and a number of other kinase signaling pathways. • STAT3 inhibitor VII (#573103—5 mg; Calbiochem, Merck Millipore) inhibiting the Janus kinase/Signal transducers and activators of transcription 3. • Bay11 7082 (#1968710—10 mg; Calbiochem, Merck Millipore) inhibiting nuclear factor kappa B (NFκB). • SB203580 (#559389—5 mg; Calbiochem, Merck Millipore) inhibits the p38MAPK. • Wortmannin (#12-338; Calbiochem, Merck Millipore) specific inhibitor of the phosphtidylinositol-3-kinase (PI3K).
The inhibitors were diluted in dimethyl sulfoxide (AppliChem, Darmstadt, Germany) to final concentrations of 100 nM/mL AG490, 170 nM/mL STAT3 inhibitor, 10 μM/mL Bay11, 100 nM SB203580, and 5 nM/mL Wortmannin. For these inhibitor experiments, SGN were treated with PRP (1:10) and an inhibitor to reverse the effect of PRP.
Statistical analyses
All data were presented as mean ± standard error of mean. Data were analyzed for statistical significance using one-way analysis of variance followed by Bonferroni's multiple comparison test. Statistical analyses were conducted using the software GraphPad Prism 5 (GraphPad, La Jolla). p Values of less than 0.05 were considered to be statistically significant. All data represent the mean of three independent approaches (N), including triplicates of each sample (n) (N = 3, n = 3). Levels of significance are indicated as follows: *p < 0.05; **p < 0.01; and ***p < 0.001.
Results
Effects of PRP on dissociated SGN
An increased neuronal survival was observed after treatment with BDNF and the different concentrations of PRP (Fig. 1A). Experimental cultures treated with concentrated (2:1) and undiluted (1:1) PRP were difficult to analyze due to the high amounts of platelets that obscured the visualization of the stained neurons (data not shown). Thus, only diluted PRP was used for the survival experiments. The highest survival rate was obtained after treatment with PRP at a concentration of 1:10 (27.9% ± 2%) and 1:50 (25.5% ± 3.1%) (Fig. 1A). This effect was statistically significant (p < 0.001) when compared with the NC (4% ± 0.3% survival rate after cultivation in medium without the addition of growth factors) and even to the positive control (10% ± 1.1% after treatment with BDNF at a concentration of 50 ng/mL).
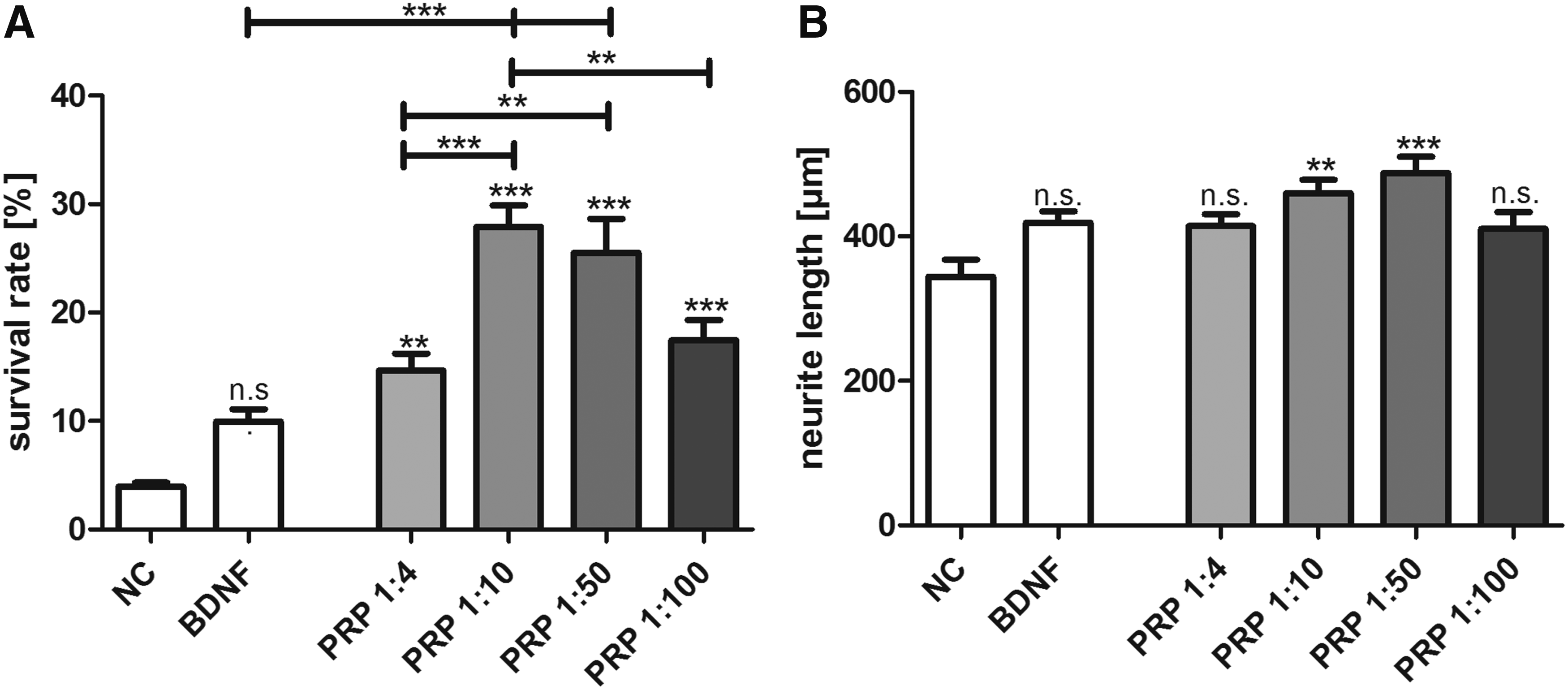
More concentrated PRP (1:4) led to a significantly lower survival of the SGN (14.6% ± 1.6%) when compared with the treatment with PRP at the most efficient concentrations (1:10 and 1:50; p < 0.001 and p < 0.01, respectively). At higher dilutions of PRP (1:100), the neuroprotective effects starts to decline (17.5% ± 1.9%) and is similar to the one achieved with higher PRP concentrations (PRP 1:4 vs. PRP 1:100; n.s.).
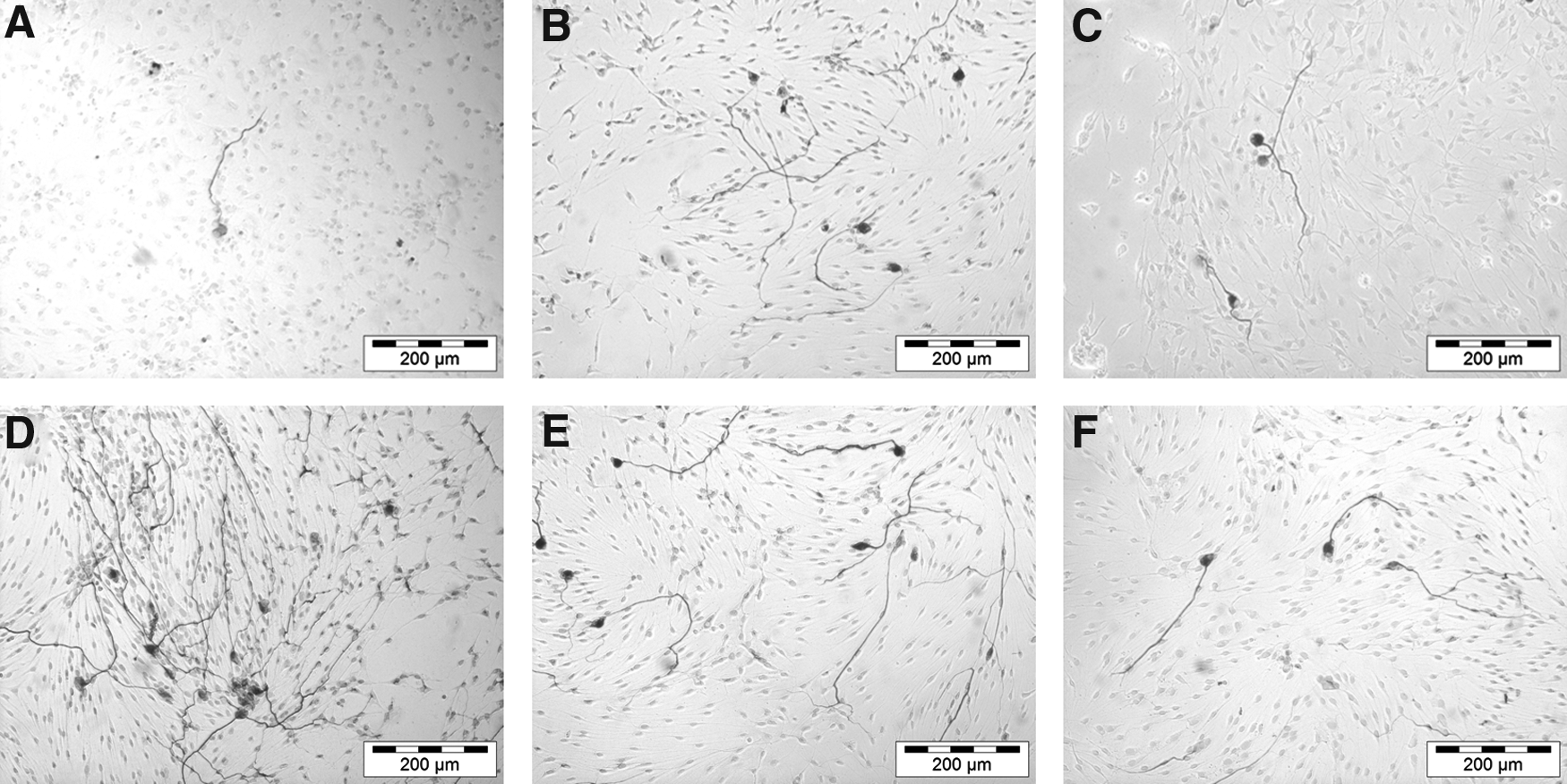
The neurite length was significantly increased for SGN treated with PRP at a concentration of 1:10 (459.8 ± 18.5 μm; p < 0.01) and 1:50 (487.7 ± 22.7 μm; p < 0.001) when compared with the NC (344.2 ± 23.4 μm) (Fig. 1B). Thus, the treatment with PRP increased the neuroregenerative effect of SGN. Representative images are shown in Figure 2. However, the neuroregenerative effect of PRP is much weaker than the neuroprotective effect.
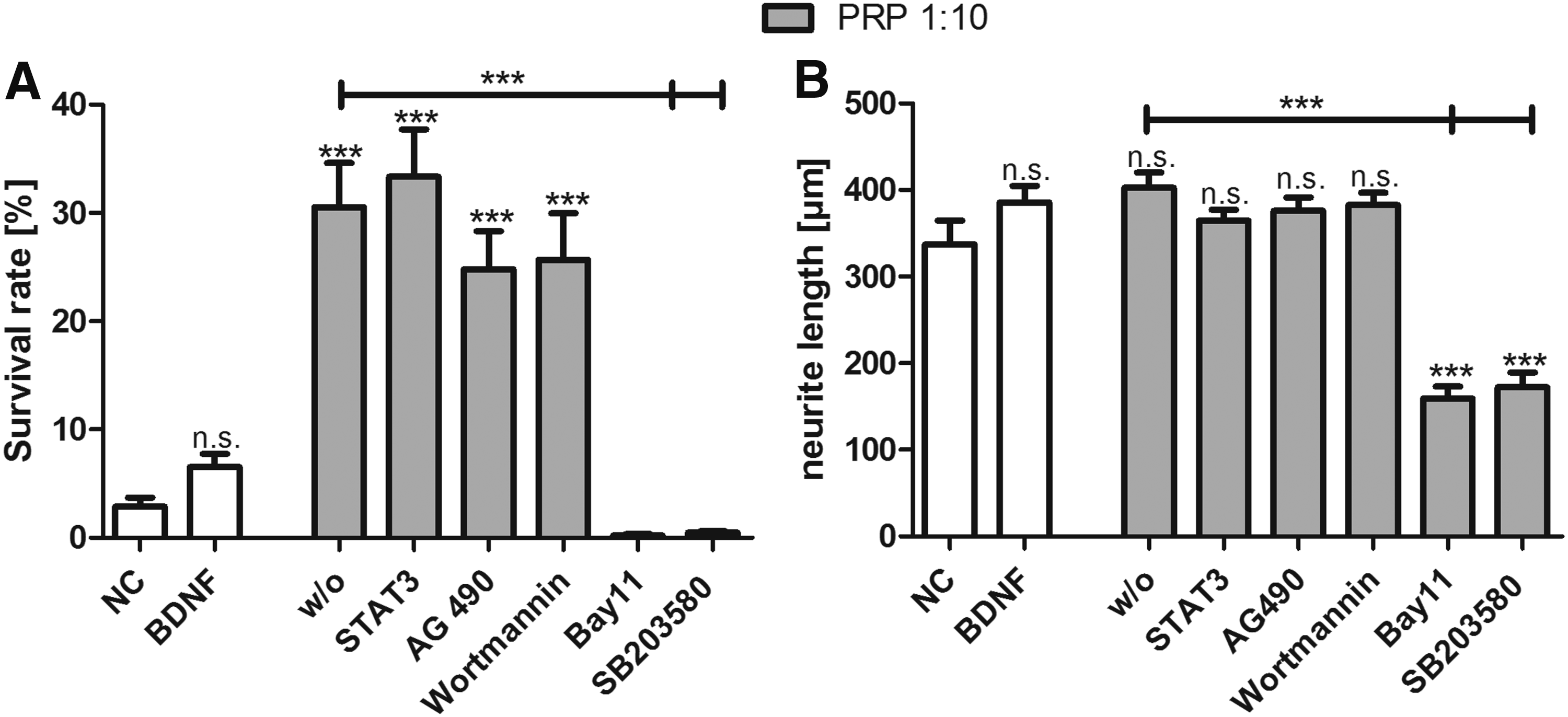
Inhibition of the neuroprotective effect of PRP
The JAK2 inhibitor AG490, the STAT3 inhibitor VII, and the specific inhibitor of the PI3K Wortmannin did not reverse the neuroprotective effect of PRP (1:10) in SGN (Fig. 3A). However, treatment with Bay11 7082 (inhibitor of NFκB) and SB203580 (inhibitor of the p38MAPK) totally reversed the observed neuroprotective effect of PRP (Fig. 3A). The neurite outgrowth was not affected by the inhibitors AG490, STAT3 inhibitor VII and Wortmannin (Fig. 3B). Treatment with Bay11 7082 and SB203580 significantly decreased the neurite length in SGN treated with PRP at a concentration of 1:10 (p < 0.001).
Enhanced adhesion of spiral ganglia explants and enhanced neurite outgrowth due to PPP as adhesion matrix
About one-third of the explants adhered spontaneously to the culture wells that were not coated (w/o, 27.8% ± 6.8%) (Fig. 4). Using PPP at a concentration of 1:10 as cell-free adhesion matrix, the adhesion rate was similar (18.5% ± 7.1%, n.s.). Increasing the concentration of PPP for the coating up to 1:2 and 1:4, the adhesion rates significantly increased up to more than 60% (64.8 ± 5.9 for PPP 1:2 and 63.0 ± 6.1 for PPP 1:4; **p < 0.01).
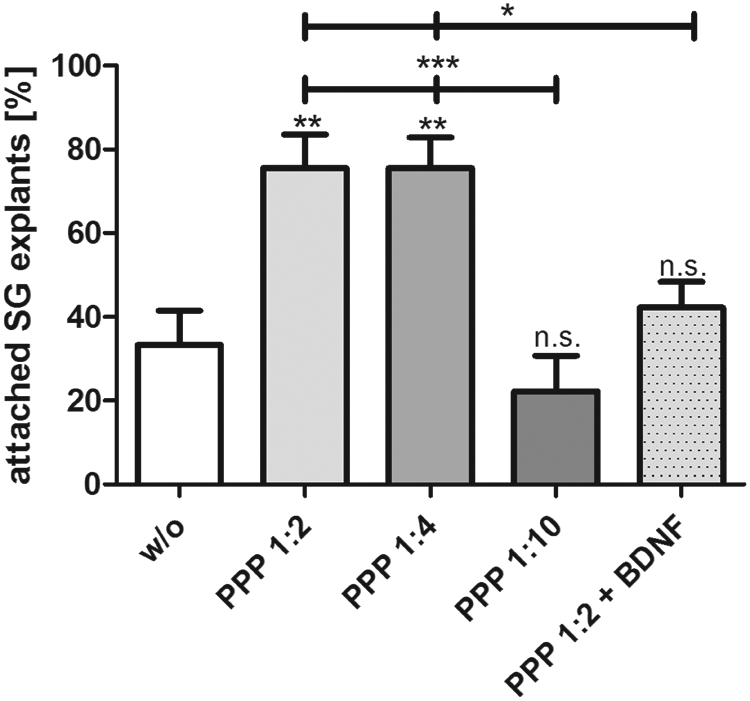
PPP was used as matrix for the adhesion and survival of the spiral ganglion explants. PPP was activated with calcium gluconate. Increased attachment of SG explants was observed when using PPP (1:2 and 1:4 dilution) as matrix for the coating of the wells. This effect could not be further increased by adding BDNF to the PPP. Asterisks over the error bars indicate the significance of the treatment condition compared with the control (w/o = without any coating). Statistical assessment was performed using ANOVA (ns; *p < 0.05, **p < 0.01, ***p < 0.001). PPP, platelet-poor plasma; SG, spiral ganglia.
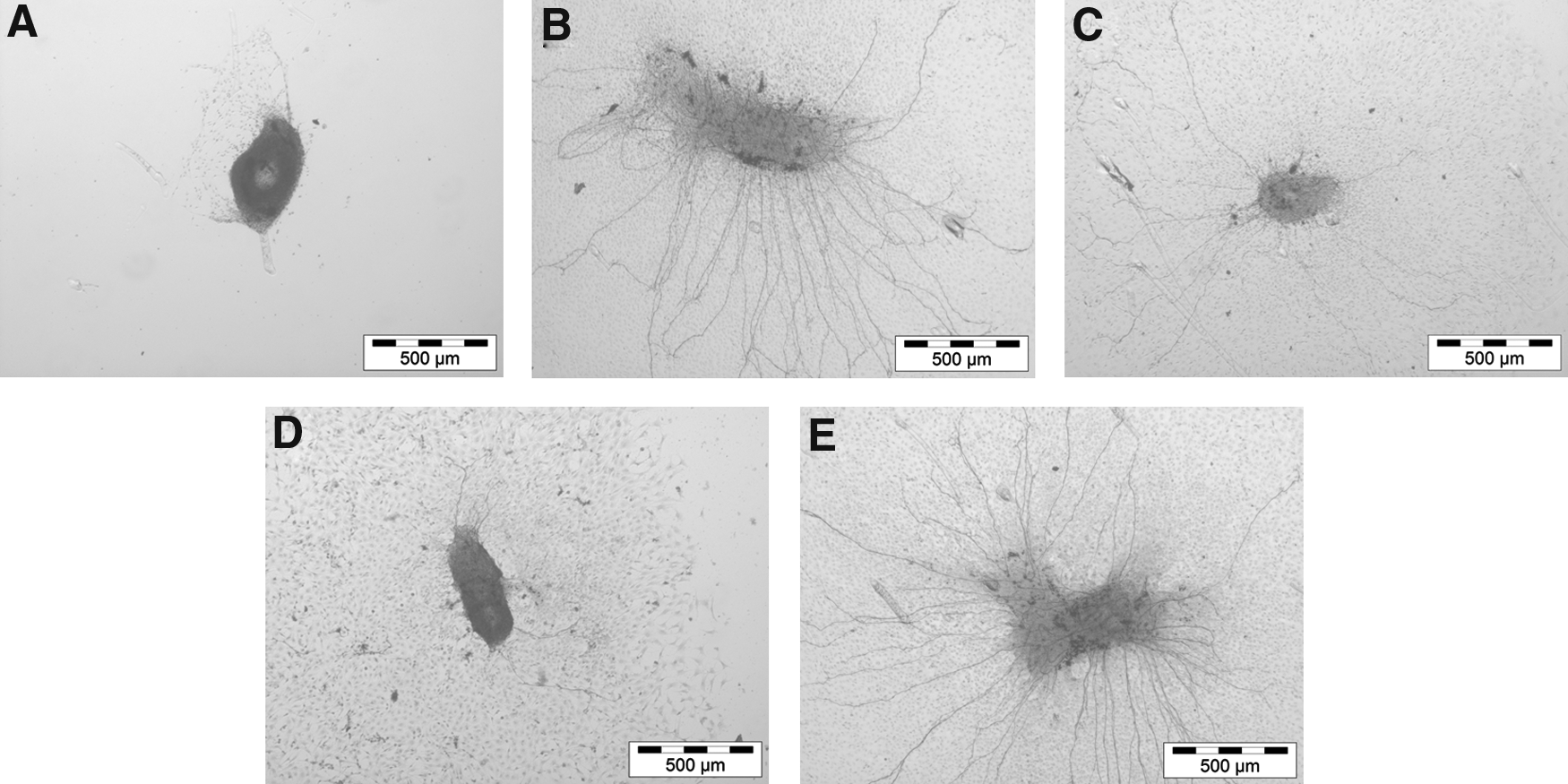
Induction of neurite outgrowth was significantly increased when the highest concentration of PPP (1:2) was used as matrix for adhesion (Figs. 5 and 6). In this condition (i.e., PPP 1:2), the number of neurites per explant was significantly increased when compared with the highest dilution (1:10) and to the uncoated control conditions (Fig. 5; p < 0.01). The highest number of neurite outgrowths was quantified in the SG explants that were treated with a mix of PPP at a concentration of 1:2 and BDNF (50 ng/mL). Representative micrographs are shown in Figure 6.
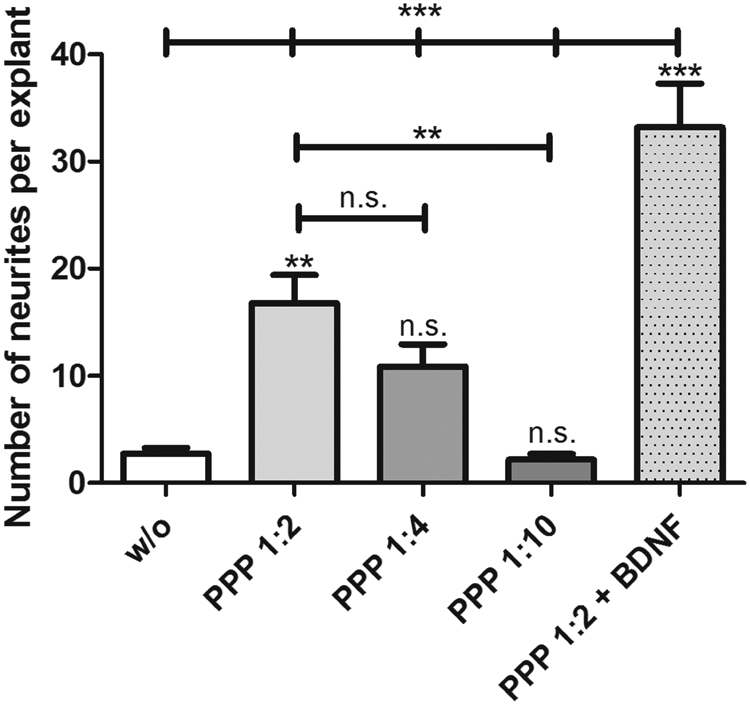
PPP was used as matrix for the adhesion and survival of the spiral ganglion explants. PPP was activated with calcium gluconate. Increased neurite outgrowth was induced from the explants attaching and growing on PPP (1:2 and 1:4) as matrix. At higher dilutions, the support and attachment of SG explants was diminished. The most prominent outgrowth of neurites from the explants occurred when the cells were offered PPP (1:2 dilution) with BDNF (50 ng/mL) as a matrix for adhesion. Asterisks over the error bars indicate the significance to the control (without any coating). Statistical assessment was performed using ANOVA (ns; **p < 0.01, ***p < 0.001).
Discussion
Blood derivatives might present an elegant approach for the delivery of defined cytokines and growth factors to the inner ear. These factors are able to control inflammation and to protect neurons from trauma. Blood derivatives such as PRP and PPP are emerging therapeutics in ophthalmology, in orthopedics, for wound healing, or for facial and bust rejuvenation.32–36 Their use in otology has not been reported thus far.
The herein presented work shows first evidence that the survival of dissociated SGN can be significantly improved due to the treatment with PRP. A significant increase of neurite outgrowth in the dissociated spiral ganglion cultures was also observed, but this was much weaker than the neuroprotective effect. This may be due to the quite high spontaneous neurite outgrowth in surviving neurons without any treatment. Thus, PRP offers protective and regenerative factors to the SGN that are more potent than BDNF. However, the effect was only observed when PRP was highly diluted (1:10). Obviously, too high concentrations of growth factors may lead to adverse effects. The importance of the concentration of PRP for clinical use has been emphasized as well for the survival of mesenchymal stem cells encapsulated in PRP gel. 37
The neurotrophin BDNF exerts neuroprotective but also apoptotic effects by activation of different receptors and the regulation of these different activities are crucial.38,39 The low-affinity receptor p75NTR, responsible for apoptosis and inhibition of neurite outgrowth is upregulated after trauma 38 and strategies to counteract this upregulation would be beneficial in cochlear implantation. It is not clear how exactly PRP exerts the neurotrophic effects observed in our study. One possibility could be the downregulation of p75NTR and the upregulation of receptors in spiral ganglion cells that promote protection and regeneration.
In addition, anti-inflammation exerted by the different immune cells and factors that are also present in PRP 35 might be another possibility. Very likely, the cocktail of the different factors that are present in the plasma and that are released from the platelets is highly protective. Similar results have been observed in SGN cultures with a combination of different factors.11,40 The neuroregenerative effects of PRP and PPP have been successfully used to improve peripheral nerve regeneration by providing acellular nerve allografts loaded with PRP as scaffold for tissue engineering in an animal model of nerve injury. 41 Similar results can be achieved for the auditory nerve. However, even minimal amounts of blood applied to the inner ear have been shown to lead to detrimental effects on hearing in guinea pigs. 42
The adverse effects of blood in the inner ear may rather be related to the presence of erythrocytes. These, however, are not present in PPP and PRP. If advanced toward clinical application in otology, thorough in vivo investigations of PRP and PPP concerning their putative toxicity and the concentration to be applied are mandatory. In addition, the impact of PRP and PPP on the cochlear micromechanics needs thorough consideration.
Among the plethora of growth factors and cytokines found in PRP, platelet-derived growth factor and insulin-like growth factor-1 (IGF-1) are the main growth factors.43–45 They show strong potency in the protection of the cochlea.46–50 IGF-1 has been successfully applied in the clinic for the treatment of inner ear disorders. 51 These factors and other substances, such as catecholamines, transmitters, ATP, and calcium ions, are involved in the different innate processes of tissue repair 20 and regeneration. 52
From our matrix experiments, we may conclude that PPP offers a favorable environment not only for the adhesion of the spiral ganglia, but also facilitated enhanced neurite outgrowth from the explants. It seems likely that the proteins present in the PPP form—upon activation with calcium gluconate—polymers that cover damaged areas offering smooth surfaces that facilitate wound healing. Generating such smooth matrices on the surface of the cochlear implant by coating procedures may offer a more physiological surface that promotes adhesion of outgrowing neurites without allowing the formation of fibrous scar tissue around the electrode array.
To identify the signaling cascades that are involved in the protection of SGN by PRP, several inhibitors were tested. The two inhibitors AG490 and STAT3 are inhibitors of the Janus kinase/signal transducers and activators of transcription 3 (JAK/STAT3) pathway, and this pathway is mainly activated by erythropoietin (EPO). 53 Since EPO strongly protects neurons against neurotoxicity 54 and may be present in PRP, it seems possible that part of the protective and neuroregenerative effects of PRP is mediated through EPO by the JAK/STAT3 pathway. However, neither the specific inhibitor STAT3 nor the unspecific inhibitor of JAK AG490 shows a significant effect on the cultures treated with PRP. Thus, we may conclude that the JAK/STAT3 pathway is not mainly involved in the neuroprotection of SGN mediated by PRP.
PI3K is one of the key enzymes activated in signaling pathways of growth factors. For example, neurotrophins such as BDNF and NT-3 use the PI3K pathway to maintain target innervation.55,56 Using Wortmannin, a PI3K inhibitor, only slightly but not significantly reverses the effects of PRP on neuronal survival. Thus, PI3K also seems not to be the main pathway for neuroprotection activated by PRP in our cell culture.
The p38 mitogen-activated protein kinase (MAPK) and NFκB are involved in inflammation and are known to mediate proatherogenic effects. 57 However, NFκB is the most widely recognized transcription factor for its quintessential roles in regulating inflammation and immune responses 15 as well as for the upregulation of prosurvival genes such as Bcl-2 in neuronal cells. 58 In addition, the most extensively investigated neurotrophin BDNF exerts its neuroprotective effects also by the upregulation of the telomerase activity. 59 Telomerase can be regulated by its promotor NF-κB and this is a downstream target of MAPK, 59 suggesting a crosstalk between the MAPK pathways and the NF-κB transcriptional cascade. 60 Thus, we assessed their involvement in mediating the neuroprotective effects of PRP.
Inhibition of the p38-MAPK by SB203580 completely reversed the positive effects of PRP on SGN survival. Interestingly, this pathway is not involved in the mediation of NT-3-induced neurite outgrowth as shown by Aletsee et al. 61 The neuroprotective effects of PRP on SGN seem to be depending on NF-κB since the increased survival was blocked by the use of Bay11 7082, an irreversible NF-κB inhibitor. Bay11 7082 not only inhibits NF-κB but also several other pathways, such as the PI3K/Akt, ERK/JNK, as well as the JAK/STAT-1 signaling pathway. 62 By contrast, SB203580 is a specific inhibitor of the alpha and beta isoform of the p38 MAPK. 63 Based on our results, we may conclude that the p38 MAPK pathway seems to be involved in the mediation of the neuroprotection of SGN by PRP. However, other pathways may also be involved that have not been investigated in our study.
One bias of this study may be the use of blood drawn from relatively young females for the generation of PRP and PPP. Thus, future studies should include PRP and PPP generated from blood of male and/or elderly donors to prove if gender- or age-specific differences may influence the potency of PRP and PPP.
Conclusions
Autologous plasma rich in growth factors formulated as PRP or PPP seems very promising for the optimization of the electrode–nerve interaction in cochlear implantation and other neural prostheses by improving neuronal survival and their adhesion to the electrode arrays. Despite their balanced composition of growth factors and other molecules, their applied concentration seems crucial for the inner ear. Further investigations are necessary to verify their potency in vivo and to derive clinically applicable biological therapies.
Footnotes
Acknowledgments
The authors would like to thank RegenLab for providing the centrifuge as well as the RegenKits for generation of PRP and PPP. The technical assistance of Darja Werner is kindly acknowledged. This work was funded by the Cluster of Excellence of the German Research Foundation (DFG; “Deutsche Forschungsgemeinschaft”) “Hearing4all,” grant number EXC 1077/1.
Disclosure Statement
No competing financial interests exist.