
Abstract
Damaged articular cartilage has limited healing capacity due to the lack of blood supply. Tissue engineering using the chondrogenic potential of mesenchymal stem cells (MSCs) is a promising approach for cartilage regenerative therapy. Canines are a relevant animal model for cartilage regeneration, as they suffer from spontaneous cartilage injury similar to humans. However, the chondrogenic capacity and optimal conditions for chondrogenesis in canine MSCs have not been documented. In this study, we investigated the effect of preconditioning with fibroblast growth factor-2 (FGF-2) and fetal bovine serum (FBS) in chondrogenic induction medium on canine MSC chondrogenesis. Our results show that treatment with FGF-2 promoted cell proliferation and the expression of SOX2. Biochemical and histological analyses revealed that higher concentrations of FBS decreased chondrogenesis of canine MSCs expanded without FGF-2. However, cells expanded with FGF-2 showed substantial chondrogenic potential and produced abundant cartilage matrix, even on FBS addition. Quantitative polymerase chain reaction showed that chondrogenic genes were highly expressed in spheroids, when cells were expanded with FGF-2 under serum-free conditions. Thus, the combination of FGF-2 preconditioning and serum-free chondrogenic induction medium enabled efficient chondrogenic differentiation of canine MSCs and contributed to cartilage regeneration research.
Impact Statement
Tissue engineering using the chondrogenic potential of mesenchymal stem cells (MSCs) is a promising approach for cartilage regenerative therapy. Although dogs are widely used as an animal model for cartilage regeneration, chondrogenic differentiation of canine MSCs is still challenging. In this study, we aimed at establishing the optimal conditions for canine MSC chondrogenesis. Our results demonstrated that preconditioning with fibroblast growth factor-2 and serum-free induction medium enabled robust chondrogenesis of canine MSCs. These findings will allow effective generation of cartilage tissue from canine MSCs and advance research of cartilage regeneration in both dogs and humans.
Introduction
Articular cartilage consists of water, chondrocytes, and abundant extracellular matrix, including proteoglycan, type II collagen, and hyaluronic acid. 1 Once articular cartilage is severely damaged, it has little self-repair or regenerative capacity due to its avascular structure, causing persistent joint pain. Autologous chondrocyte transplantation is one solution for cartilage defects. 2 However, there are some complications with this treatment, including donor site morbidity and hypertrophy of the transplant.3,4 Moreover, harvested chondrocytes have limited proliferation ability in vitro and undergo dedifferentiation, which results in the loss of their original phenotype, transforming into fibrocartilage.5–7 Therefore, a new cell source is required to regenerate articular cartilage.
Mesenchymal stem cells (MSCs) are easily isolated from adult somatic tissues such as bone marrow and fat with minimally invasive procedures and low cost. MSCs show high proliferative ability and can differentiate into several mesenchymal lineages, including adipocytes, osteoblasts, and chondrocytes. Therefore, MSCs are expected to be a clinically useful cell source for cartilage regeneration in human and veterinary medicine. In addition, many preclinical animal models such as rodents, rabbits, dogs, and pigs are used in cartilage research using MSCs. 8
Dogs suffer from spontaneous cartilage injury associated with joint diseases, which shows similar pathological features to those observed in humans. Therefore, dogs are considered to be a relevant animal model for human cartilage injury. 8 For this reason, many researchers have tried to isolate and chondrogenically differentiate canine MSCs. Unfortunately, most reports have shown that canine MSCs have poor chondrogenic capacity and produce only small amounts of glycosaminoglycan (GAG) in a pellet culture.9–13 Thus, further studies should be conducted to verify the critical factors for the optimal chondrogenic condition of canine MSCs.
Fetal bovine serum (FBS) is widely used as a medium supplement to aid cell proliferation and differentiation because it contains several types of growth factors, including both cytokines and hormones. Previous evidence suggests that adding FBS to chondrogenic induction medium inhibits MSC chondrogenesis and increases apoptosis.14,15 Conversely, Diekman et al. reported that 10% FBS promoted chondrogenic differentiation of human adipose tissue-derived MSCs, whereas the opposite was observed for human bone marrow MSCs (BMMSCs). 16 To our knowledge, no study has investigated the impact of FBS in chondrogenic induction medium for canine MSCs, and it remains unclear at what concentration FBS should be added.
Fibroblast growth factor-2 (FGF-2), also known as basic FGF, is a member of the FGF family and is a strong mitogen for several cell types. FGF-2 also plays an important role in embryogenesis, cell differentiation, and maintenance of stemness in pluripotent stem cells. 17 Previous studies have indicated that FGF-2 promoted the proliferation of human MSCs in monolayer culture and subsequent chondrogenic differentiation.18–21 Since similar results have been reported as well in canine BMMSCs,22,23 FGF-2 preconditioning will be necessary to optimize the chondrogenic condition of canine MSCs.
We have previously developed novel canine MSCs, known as bone marrow peri-adipocyte cells (BM-PACs), derived from cells adhering to adipocytes in bone marrow. 24 Since BM-PACs showed superior proliferation and differentiation ability compared with that of BMMSCs derived from bone marrow mononuclear cells, higher chondrogenic potential could be expected under optimal conditions. In this study, we aimed at establishing the optimal conditions for canine BM-PAC chondrogenesis for two approaches: FGF-2 preconditioning and FBS concentration in a chondrogenic medium. First, canine BM-PACs were expanded with or without FGF-2 and their proliferation ability and undifferentiated state were evaluated. Subsequently, these cells were cultured in chondrogenic medium containing 0%, 1%, and 10% FBS to determine the effect of FBS concentration on chondrogenic differentiation of canine MSCs.
Materials and Methods
Animals
Six healthy young beagles (four male and two female) were used in this study to obtain bone marrow. Bone marrow samples were harvested under general anesthesia. Anesthesia was induced with propofol and maintained with isoflurane (2.0%) in oxygen. All animal experiments were approved by the Animal Care Committee of the Graduate School of Agricultural and Life Sciences at the University of Tokyo.
Culture of canine BM-PACs
Canine BM-PACs were isolated according to our previously published method. 24 Briefly, bone marrow was aspirated from the proximal humerus by using a sterilized 15-gauge bone marrow biopsy needle (Angiotech Pharmaceuticals, Inc., Vancouver, Canada). After density gradient centrifugation with Ficoll-Paque (GE Healthcare, Little Chalfont, United Kingdom), the top adipose layer containing mature adipocytes was collected, washed with Dulbecco's modified Eagle's medium (DMEM) (Invitrogen, Carlsbad, CA), and supplemented with 10% FBS (Lot no. 1526235; Gibco/Invitrogen) and 1% penicillin–streptomycin/amphotericin-B (Wako). After a second wash, the adipose layer was placed in 25 cm2 flasks filled with DMEM supplemented with 20% FBS and 1% antibiotics. Cells were then subjected to ceiling culture at 37°C in a humidified atmosphere containing 5% CO2 for 7 days. If cells did not grow to 80% confluence within 7 days, the flask was inverted and the culture was maintained with medium containing 10% FBS.
Cell proliferation assay
Passage 0 BM-PACs were plated at a density of 1 × 104 cells/cm2 and expanded in growth medium supplemented with or without 10 ng/mL of human recombinant FGF-2 (Peprotech, Rocky Hill, NJ). The confluent cells were detached with 0.25% trypsin/1 mM EDTA solution (Wako), and the number of viable cells was counted by using a trypan blue exclusion test. Doubling time (DT) was calculated according to the following formula: DT = T × log2/(log Nt – log N0), where T is the incubation period (days), and N0 and Nt indicate the initial seeding cell number and the final harvesting cell number, respectively.
Chondrogenic induction in spheroid culture
Cells expanded with or without FGF-2 were plated in low-adhesive 96-multi-well plates (Sumitomo Bakelite, Tokyo) to achieve spheroid formation at a density of 3 × 104 cells per well. For chondrogenic differentiation, spheroids were cultured in chondrogenic induction medium consisting of DMEM, 4.5 mg/mL
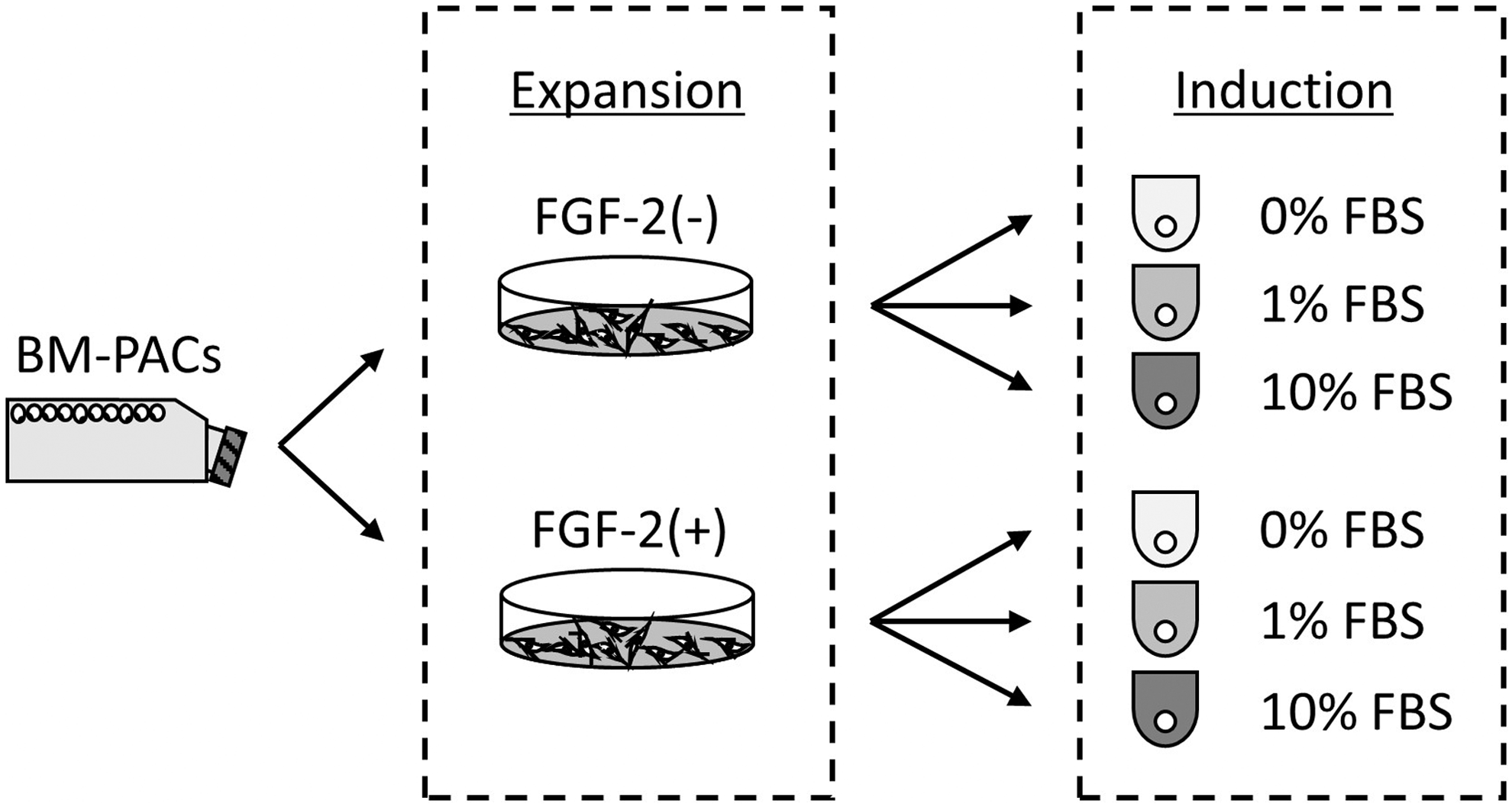
Schematic representation of the experiment. P0 BM-PACs were passaged and expanded with or without FGF-2 in a monolayer culture. Subsequently, cells were subjected to suspension culture to form spheroids in chondrogenic differentiation media containing 0, 1, or 10% FBS. BM-PAC, bone marrow peri-adipocyte cell; FBS, fetal bovine serum; FGF-2, fibroblast growth factor-2.
DNA and GAG contents
After 7 and 14 days of chondrogenic induction, four spheroids in each group were digested with 100 μg/mL papain at 65°C for 4 h. Total spheroid DNA content was determined with Hoechst 33258 dye (Dojindo Molecular Technologies, Kumamoto, Japan). Fluorescence intensity was measured by using the multilabel counter ARVO MX (Perkin Elmer, Waltham, MA) at an excitation wavelength of 355 nm and an emission of 460 nm. A standard curve was generated by serial dilution of calf thymus DNA (Sigma). Total spheroid GAG was then quantified by using a Blyscan Kit (Biocolor, Westbury, NY) according to the manufacturer's instructions. The optical density at 630 nm was read by using a microplate reader (BioRad, Hercules, CA), and total GAG content was normalized to total DNA content. Each experiment was done in duplicate.
Histological staining and immunohistochemistry
After 14 days of chondrogenic induction, spheroids were fixed with 10% formalin neutral buffer solution (Wako) and embedded in paraffin. Specimens were cut into 4-μm sections. To detect proteoglycan, sections were stained with Safranin O/Fast Green. Immunohistochemistry was done to assess the expression of collagen type II, I, and X. Antigen retrieval was carried out with 50 μg/mL proteinase K (Promega, Madison, WI) for 10 min at room temperature. For collagen type II, the additional antigen retrieval step was done by using 25 mg/mL hyaluronidase (Sigma) for 2 h at 37°C. The sections were then incubated with 0.3% hydrogen peroxide in methanol for 30 min to inhibit endogenous peroxidase activity and washed with Tris-buffered saline with 0.1% tween-20 (TBS-T). After blocking with TBS-T containing 10% normal goat serum (Sigma) for 30 min at room temperature, the sections were incubated with rabbit anti-bovine type II collagen antibody (1:200, LB-1297; LSL, Tokyo, Japan), mouse anti-bovine type I collagen antibody (1:1000, ab6308; Abcam, Cambridge, United Kingdom), and mouse anti-porcine type X collagen antibody (1:1000, C7974; Sigma) at 4°C overnight. Slides were washed with TBS-T three times for 5 min each and incubated with HRP-labeled polymer (K4001 and K4003; Dako, Tokyo) for 1 h at room temperature. Finally, sections were washed with TBS-T three times for 5 min each and DAB substrate (Dako) was applied. All slides were counterstained with hematoxylin.
Quantitative real-time reverse transcription polymerase chain reaction
Total RNA was extracted with TRI Reagent (Cosmo Bio, Tokyo, Japan) from cells treated with or without FGF-2 for 2 days, and from spheroids after 14 days of chondrogenic induction. Complementary DNA was synthesized with ReverTra Ace qPCR RT Master Mix with gDNA Remover (Toyobo, Osaka, Japan). Real-time quantitative PCR was done by using real-time monitoring of SYBR Green dye (Thunderbird SYBR qPCR Mix; Toyobo) fluorescence increase on the Step One Plus Real-Time PCR system (Applied Biosystems, Foster City, CA). The mRNA expression of chondrogenic marker gene (SOX9) and pluripotent marker genes (SOX2 and Oct4) was evaluated in cells treated with or without FGF-2. The expression levels of mRNA were normalized to the level of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) expression. The mRNA expression of SOX9, ACAN, COL2, COL1, COL10, and HPRT1 was evaluated in chondrogenic spheroids. HPRT1 was used as an endogenous control gene. The primers used for PCR are listed in Table 1. The PCR cycling conditions used were one cycle at 95°C for 10 min, 40 cycles at 95°C for 15 s and at 60°C for 1 min. Each experiment was done in triplicate.
Primers Used for Quantitative Real-Time Polymerase Chain Reaction
Statistical analysis
All data were expressed as mean ± standard deviation. Comparisons were made by using the Student's t-test between two unpaired groups. One-way analysis of variance followed by Tukey's multiple-comparisons test was used to detect any statistical differences between multiple unpaired groups. Statistical significance was accepted at p < 0.05. All statistical analyses were performed with EZR (Saitama Medical Center, Jichi Medical University, Saitama, Japan), which is a graphical user interface for R (The R Foundation for Statistical Computing, Vienna, Austria). 25
Results
The effect of FGF-2 on BM-PACs in monolayer expansion
BM-PACs expanded with FGF-2 for 2 days became smaller and more spindle shaped than those expanded without FGF-2 (Fig. 2A). In addition, BM-PACs expanded with FGF-2 reaching confluency in shorter time than cells expanded without FGF-2. The DT of BM-PACs expanded with FGF-2 was significantly shorter (p < 0.0001) than that of cells expanded without FGF-2 (Fig. 2B). The expression of chondrogenic and immature markers was compared after 2 days of FGF-2 stimulation. There was no significant difference in the mRNA expression of SOX9 and Oct4 between the two groups (Fig. 2C). However, significantly higher expression of SOX2 mRNA was observed in BM-PACs expanded with FGF-2 (p < 0.05).
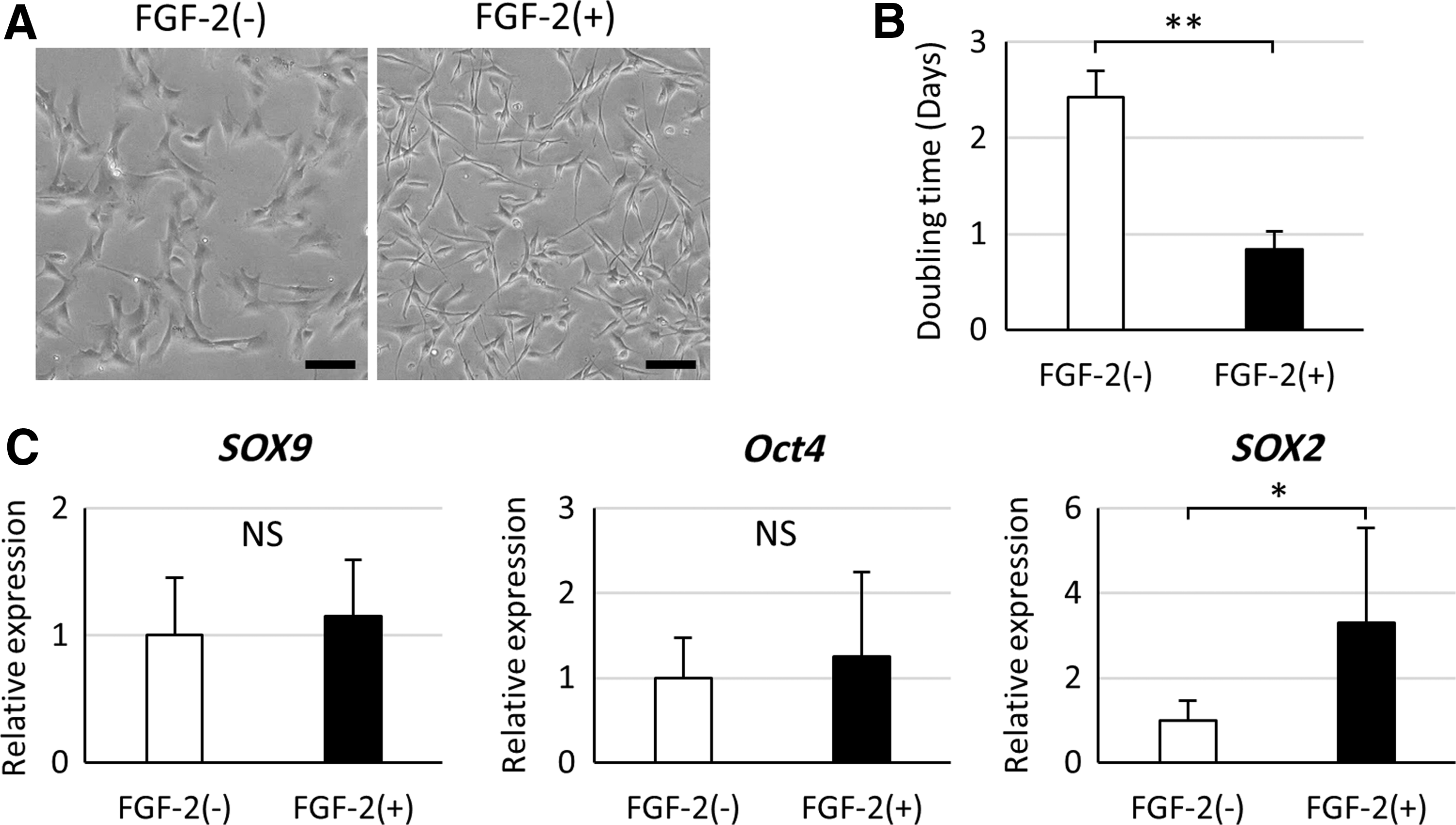
The effect of FGF-2 on BM-PACs in monolayer expansion.
Spheroid diameter
After 7 and 14 days of chondrogenic induction, there was no significant difference in the diameter of spheroids among the FGF-2(−) groups (Fig. 3). However, the diameter of spheroids in FGF-2(+) groups became larger depending on the FBS concentration in the chondrogenic induction medium. In addition, a higher concentration of FBS increased the diameter of spheroids in FGF-2(+) groups from day 7 to day 14.
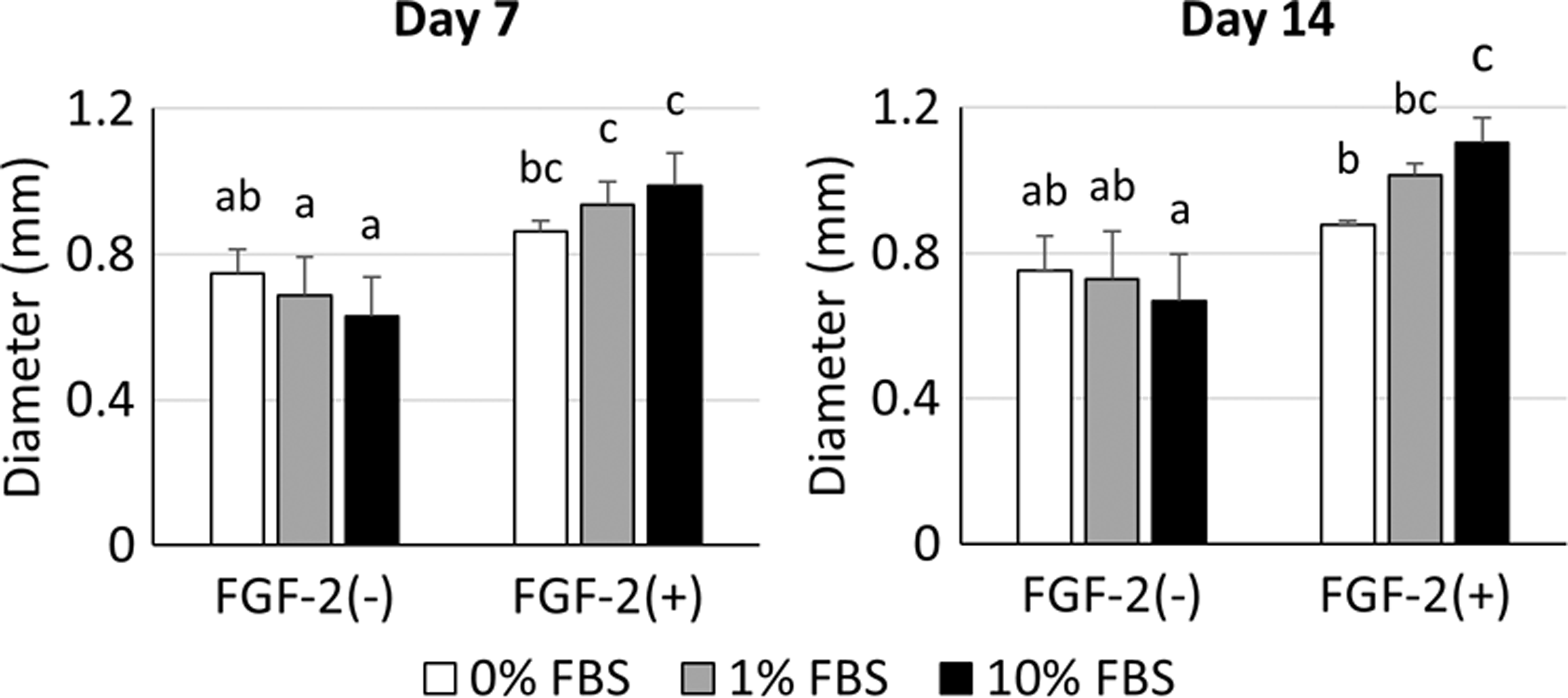
The diameter of spheroids after 7 and 14 days of chondrogenic differentiation. No significant difference was recorded for spheroid diameter among FGF-2(−) groups. Spheroids in FGF-2(+) groups became larger depending on FBS concentration. All groups not sharing common letters are significantly different (p < 0.05).
Biochemical analysis
The DNA content of spheroids in FGF-2(−) groups was stable throughout chondrogenic differentiation regardless of FBS concentration. In contrast, DNA content in FGF-2(+) groups increased in an FBS concentration-dependent manner (Fig. 4A). Spheroids in FGF-2(+) groups showed significantly higher GAG deposition (p < 0.05) than those in FGF-2(−) groups after 7 and 14 days of chondrogenic differentiation (Fig. 4B). To evaluate the efficiency of GAG production, total GAG content was normalized to total DNA content (GAG/DNA). Lower FBS concentration in chondrogenic medium tended to increase GAG/DNA ratio, and the highest mean value was observed in the FGF-2(+)-0% FBS group (Fig. 4C).
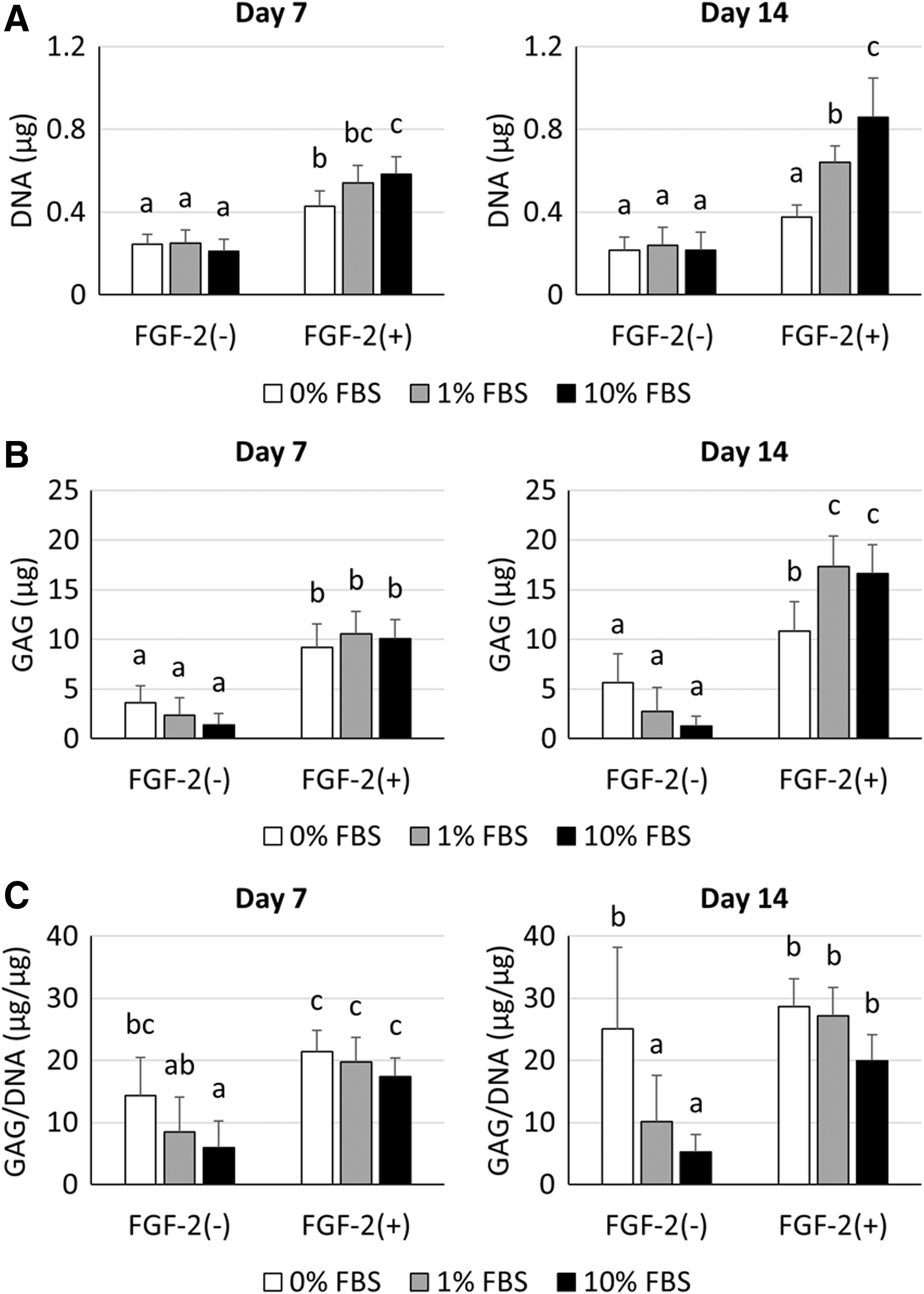
Biochemical analysis of DNA and GAG at days 7 and 14.
Histology
Safranin O staining was done to evaluate the deposition of proteoglycan in chondrogenic spheroids (Fig. 5). In FGF-2(−) groups, spheroids showed stronger staining when cultured in lower concentrations of FBS. However, all spheroids in FGF-2(+) groups were strongly stained with Safranin O, even when cultured with 10% FBS. The central region of spheroids showed stronger staining compared with the peripheral region. Higher concentrations of FBS formed a thicker Safranin O-negative layer consisting of fibroblast-like cells on the outer surface of spheroids.
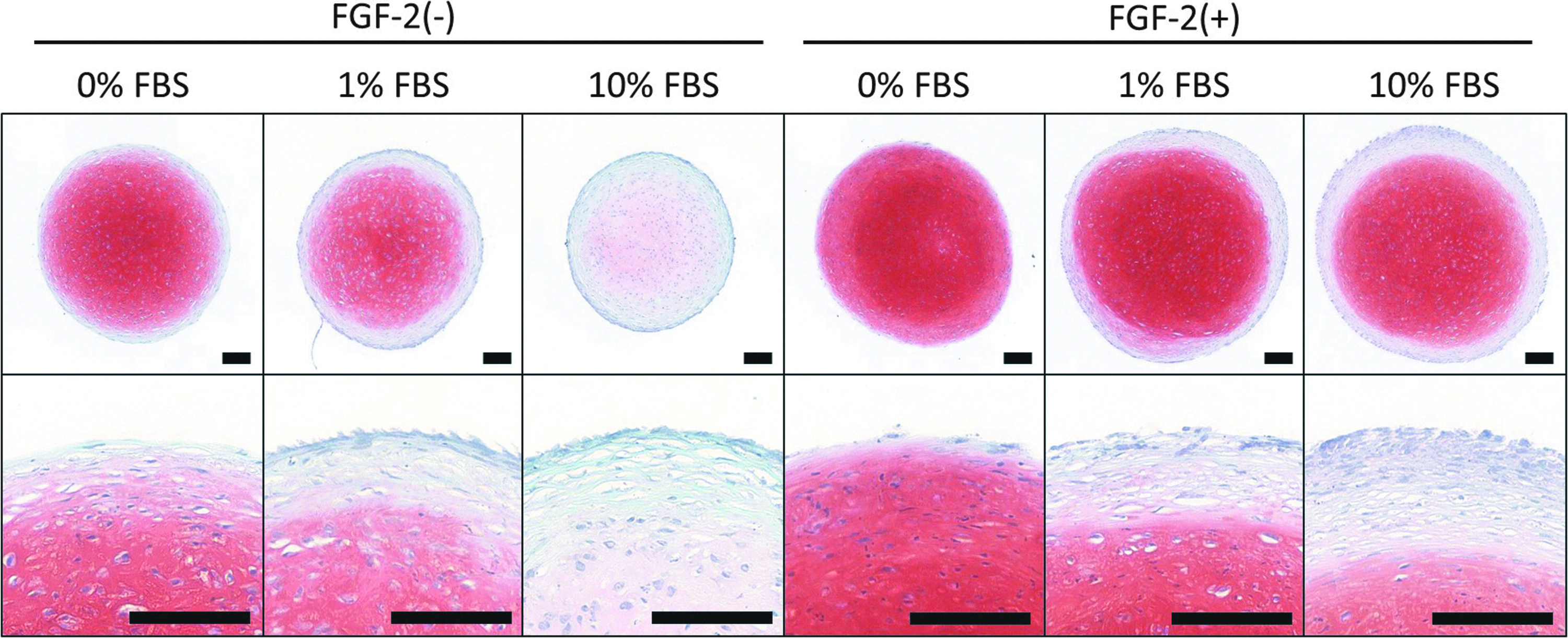
Safranin O staining of spheroids after 14 days of chondrogenic differentiation. In FGF-2(−) groups, spheroids cultured with a lower concentration of FBS showed stronger staining than those cultured at higher concentrations. All spheroids showed strong staining with Safranin O in FGF-2(+) groups. Safranin O-negative outer layers consisted of fibroblast-like cells. Higher concentrations of FBS formed a thicker Safranin O-negative layer. All scale bars indicate 100 μm.
Immunohistochemistry results after 14 days of chondrogenic induction are shown in Figure 6. The expression of type II collagen was detected in all culture conditions, and there was a tendency toward greater type II collagen expression in spheroids cultured with lower concentrations of FBS. In addition, the central region of spheroids tended to express more type II collagen compared with the peripheral region. In FGF-2(−) groups, type I collagen, a marker expressed by fibrocartilage, was homogenously detected over the entire spheroid. In FGF-2(+) groups, type I collagen distribution was significant in the middle to outer surface of the spheroid, and type I collagen accumulation was not apparent in the center of spheroids. The expression of type X collagen, a marker for hypertrophic cartilage, was not observed for any conditions.

Immunohistochemistry for collagen type II, I, and X of spheroids at day 14. Spheroids cultured with a lower concentration of FBS tended to express more type II collagen. Type I collagen was uniformly detected over the spheroid in FGF-2(−) groups; whereas in FGF-2(+) groups, its distribution was more pronounced in the middle to outer surface. Type X collagen was not detected under any conditions. All scale bars indicate 200 μm.
Quantitative reverse transcription polymerase chain reaction
Spheroids cultured in chondrogenic induction medium containing 0%, 1%, or 10% FBS were collected at day 14 to do quantitative reverse transcription polymerase chain reaction. These results are shown in Figure 7 and Supplementary Table S1. The expression of the chondrogenic genes, SOX9, ACAN, and COL2, was significantly upregulated in the FGF-2(+)-0% FBS group (p < 0.05). Lower FBS concentration in chondrogenic medium tended to increase the expression of chondrogenic genes. There were no remarkable changes in the expression of COL1 and COL10 among the groups.
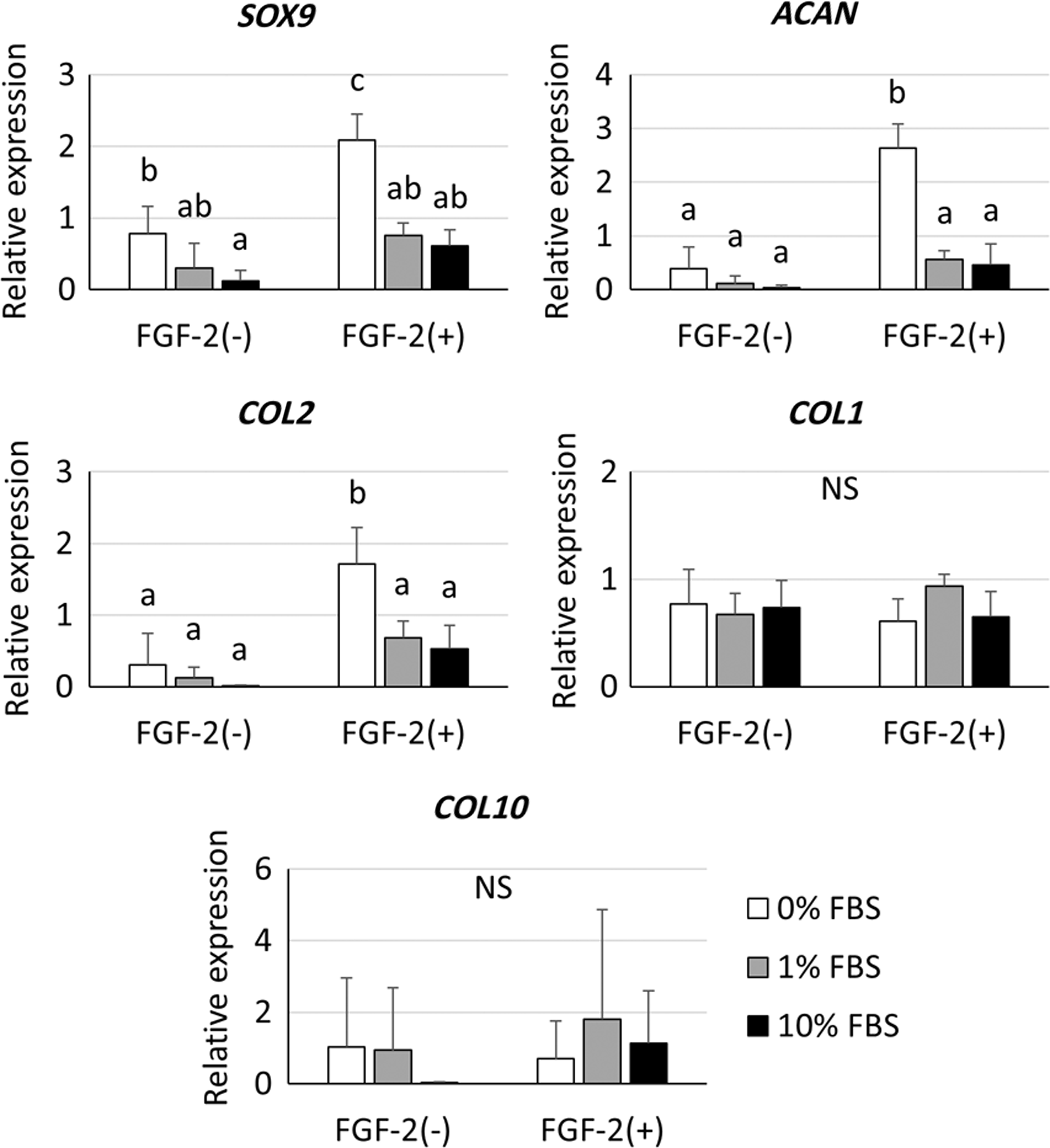
Quantitative RT-PCR was done to assess mRNA expression levels in chondrogenic spheroids. The expression of SOX9, ACAN, and COL2 was significantly upregulated in the FGF-2(+)-0% FBS group. There were no remarkable expression changes observed for COL1 or COL10. All groups not sharing common letters are significantly different (p < 0.05). NS, no significance; RT-PCR, reverse transcription polymerase chain reaction.
Discussion
In this study, BM-PACs became smaller and proliferated rapidly when treated with FGF-2 during monolayer expansion. This result is consistent with those in a previous study, which showed that FGF-2 enhanced the mitotic potential of human and canine BMMSCs.21,23 After treatment with FGF-2, there was no change in the expression of chondrogenic genes, suggesting that the stimulation with FGF-2 did not lead to cell differentiation into a chondrogenic lineage at this point. In addition, we showed that mRNA expression of SOX2 was significantly higher in cells treated with FGF-2. SOX2 is one of the transcription factors essential for the maintenance of the undifferentiated state of embryonic stem cells and induced pluripotent stem cells. Yoon et al. demonstrated that SOX2 was also important for the maintenance of proliferation and multipotency in human BMMSCs. 26
Moreover, it has been reported that FGF-2 upregulated the expression of SOX2 in human apical papilla-derived MSCs, resulting in enhanced proliferation ability and differentiation potential. 27 However, the expression of Oct4 was not changed by treatment with FGF-2. SOX2 and Oct4 are generally believed to work together to sustain an undifferentiated state. However, a previous report has indicated that Oct4 is not required for self-renewal and multipotency in mouse BMMSCs. 28 As mentioned earlier, it is suggested that FGF-2 stimulation plays an important role in SOX2-dependent enhancement of canine BM-PAC proliferation while maintaining an undifferentiated state.
Higher concentrations of FBS in the chondrogenic medium significantly decreased GAG/DNA ratio in FGF-2(−) groups. FBS is known to contain several growth factors, including transforming growth factor beta (TGF-β), epidermal growth factor (EGF), and vascular endothelial growth factor (VEGF). 29 Among these, EGF and VEGF have been reported to inhibit chondrogenesis of MSCs.30,31 Based on these reports, it is assumed that these FBS growth factors had an inhibitory effect on chondrogenic differentiation of canine BM-PACs in this study. Although FBS has lot-to-lot variations, there is a possibility that some lots of FBS contain small amounts of growth factors that inhibit chondrogenesis. Therefore, FBS should not be added to induction medium in chondrogenesis of canine MSCs.
In contrast, the GAG/DNA ratio of spheroids in FGF-2(+) groups was not significantly decreased by FBS addition. Therefore, we hypothesize that FGF-2 preconditioning makes spheroids impervious to the inhibitory effect of FBS during chondrogenic differentiation. Moreover, spheroids in FGF-2(+) groups became larger and contained more GAG and DNA than those in FGF-2(−) groups. These findings indicate that cells treated with FGF-2 proliferated in spheroids while maintaining chondrogenic capacity, resulting in increased spheroid size.
Consistent with biochemical analysis, spheroids in FGF-2(−) groups had stronger Safranin O staining at lower concentrations of FBS, and all spheroids had strong Safranin O staining in FGF-2(+) groups. However, fibroblast-like cells were observed on the outer surface of spheroids and a thicker Safranin O-negative layer was formed in an FBS concentration-dependent manner.
Generally, chondrogenically induced MSC pellets form a superficial cell layer consisting of fibroblast-like cells expressing type I collagen, even in the absence of FBS. 32 Similarly, the fibroblastic layer that showed slight type I collagen expression and no cartilage matrix was also observed in 0% FBS groups. It is believed that cells on the outer surface of spheroids are prevented from differentiating into chondrocytes due to direct exposure to inhibitory factors in FBS. Histological staining showed that FGF-2(+) groups produced more proteoglycan and less type I collagen compared with that in FGF-2(−) groups. A previous study on human BMMSCs also demonstrated that FGF-2 preconditioning enhanced GAG production and suppressed the expression of type I collagen. 33 Ito et al. also reported that FGF-2 preconditioning inactivated both insulin-like growth factor-I and TGF-β pathways that were spontaneously upregulated by long-term culture, and it enhanced the chondrogenic potential of human MSCs. 34 Although the underlying mechanism remains unknown, FGF-2 treatment is considered useful for the production of hyaline-like cartilage tissue using MSCs. Consistent with the previous studies,35–37 the central region of spheroids tended to express more cartilaginous ECMs. This is probably because spheroids have the gradient of oxygen concentration in their microenvironment and cells in the center are exposed to a hypoxic condition that enhances chondrogenesis.38–41
Immunostaining detected no expression of type X collagen. Hypertrophic chondrocytes are known to contribute to the formation of bone tissue both in vitro and in vivo. 42 Therefore, avoiding hypertrophy of cartilage tissue is an important issue for cartilage regeneration. However, the expression of COL10 mRNA was observed in all groups. Sekiya et al. have shown that the mRNA expression of COL10 was present before the findings of significant hypertrophy and increased over time during in vitro chondrogenic differentiation of human MSCs. 43 Since there is a possibility that immunostaining failed to detect slight expression of type X collagen, further studies will be needed to observe long-term outcome.
Finally, we did quantitative real-time PCR after chondrogenic induction for 14 days. The mRNA expression of chondrogenic genes was significantly upregulated in the FGF-2(+)-0% FBS group. These results are consistent with biochemical analysis and histological evaluation. In summary, the combination of FGF-2 preconditioning and serum-free chondrogenic induction medium is an efficient method for chondrogenesis in canine BM-PACs.
To our knowledge, this is the first article that has allowed efficient chondrogenic differentiation of canine MSCs and the generation of ECM-rich cartilage tissue with strong Safranin O staining. Reich et al. reported the chondrogenic induction of canine adipose-derived MSCs using a chondrogenic medium containing 1% FBS; however, downregulation of SOX9 expression and poor GAG deposition were observed. 44 In the context of our results, this was probably due to the addition of FBS into the chondrogenic medium. Further, it was reported that chondrogenesis of canine fat, synovium, and bone marrow-derived MSCs was unsuccessful even after induction with serum-free chondrogenic medium.9,13 It is believed that FGF-2 preconditioning may improve chondrogenesis of these types of canine MSCs. Although a previous study demonstrated robust chondrogenesis of canine BMMSCs expanded with FGF-2, high donor variability was observed in GAG/DNA. 21 In this study, BM-PACs cultured under the optimal chondrogenic condition showed high GAG/DNA with low donor variability, perhaps because of their superior chondrogenic potential compared with BMMSCs. 24 Even though the results of our study may be applicable only to BM-PACs, further studies should be designed to elucidate whether FBS and FGF-2 affect the chondrogenic differentiation of canine MSCs derived from other tissues. Nevertheless, BM-PACs could be a promising cell source for canine cartilage regenerative medicine.
Conclusion
FGF-2 enhanced the proliferative ability and mRNA expression of SOX2 in canine BM-PACs. During chondrogenic induction, cells preconditioned with FGF-2 could produce abundant cartilage matrix with low type I collagen expression. In addition, higher FBS concentrations inhibited chondrogenic differentiation, particularly at the outer surface of spheroids. The combination of FGF-2 preconditioning and serum-free chondrogenic induction medium efficiently promoted chondrogenesis of canine BM-PACs. These results will be useful for the abundant production of high-quality cartilage tissue from canine MSCs and contribute to the development of research for cartilage injury or joint diseases in both dogs and humans.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.