
Abstract
Tendinopathy remains a significant clinical challenge. Although there is some evidence that leukocyte-rich platelet-rich plasma can improve the symptoms of tendinopathy, more efficacious treatments will be required in the future to improve probability of successfully resolving this condition in athletes. Because optimal treatments are not currently available, there is a need to better understand the pathology of tendinopathy from the perspective of tendon progenitor cells (TPCs). TPCs isolated from normal and tendinopathy donors were characterized by their stem cell properties and proliferation capacities, along with their ability to become tenocytes under mechanical loading. The results showed a significant 2.6-fold increase in the viable cell population in tendinopathy versus normal donors. Although the percentage of self-renewing cells was similar, the total number of TPCs in tendinopathy was significantly higher (1.6-fold) than normal TPCs based on the colony formation assays. In contrast, TPCs from tendinopathy tissue showed significantly lower cellular proliferation rate by cumulative population doublings. Next, the expanded TPCs from both tissues successfully demonstrated the trilineage differentiation capabilities with specific gene markers, staining, and biochemical assays. To induce tenogenic differentiation, stretchable silicone wells were designed and fabricated, plus the creation of an adaptor platform used on a syringe pump for mechanical stretch. This economic design provided the adequate cyclic loading to drive tenogenic differentiation. With these devices, the stretch duration was optimized and showed the significant increase in scleraxis (SCX) and tenomodulin (TNMD) expression at 2.60 (fold change) and 3.86 (fold change in logarithm), respectively, by reverse transcription-quantitative polymerase chain reaction in normal TPCs after stretch. This assay also demonstrated the widespread cell reorientation following stretch in normal TPCs. In contrast, the mechanical loading did not increase the SCX gene expression; TNMD expression remained undetectable, and cell realignment was significantly less in tendinopathy TPCs. In addition, western blot analysis confirmed the elevated TNMD protein expression in normal TPCs following stretch and the lack of expression in tendinopathy TPCs. In summary, tendinopathy TPCs were unable to differentiate into tenocytes following mechanical stretch. Future studies may aim to reprogram tendinopathy TPCs to allow tenogenic induction.
Impact Statement
This article presents a model to distinguish between normal and tendinopathy progenitor cell behavior, which reveals insight into the pathophysiology of tendinopathy. With the design of a platform adaptor, mechanical stretch was applied to tendon progenitor cells (TPCs) that promoted tenogenic differentiation. This design provided programmable features for more flexible application with low cost. These devices successfully stimulated tenogenic differentiation of TPCs from normal, but not tendinopathic tendons under cyclic stretch. The scientific method provided in this article will allow testing of biologics, exosomes, and other treatment strategies to derive new, more efficient treatment of tendinopathy in the future.
Introduction
Almost 50% of all sports injuries are tendon related.1,2 Tendinopathy is one of the most common tendon disorders that is characterized by pain, swelling, and focal tendon tenderness at the affected area.3,4 The prevalence rate of lower extremity tendinopathy is ∼11.83 per 1000 person-years. 5
The histological characteristics of tendinopathy range from acute inflammation 6 to chronic degeneration with chondrogenic and osteogenic metaplasia. 7 Once injured, the repair process is sluggish due to the local hypoxic microenvironment, low metabolism, and constant load on the tendon. 8 Currently, the treatment of tendinopathy remains a significant clinical challenge in orthopedic surgery and sports medicine due to the lack of evidence-based management.9–11 Conservative treatments for tendinopathy, such as physical therapy and the use of nonsteroidal anti-inflammatory drugs, tend to produce short-term pain relief, but lack long-term effect. 12 On the other hand, operative management has a higher complication rate compared to other treatment options. Therefore, there is a need to better understand the pathophysiology of tendinopathy to develop more effective treatments.
The analysis of tendon progenitor cells (TPCs) in human and animal tendons13–15 has shown that they possess all features of progenitor cells such as self-renewal, clonogenicity, and multipotency with a strong tendency toward induction into tenocytes under normal physiological conditions. 14 Therefore, TPCs may play a pivotal role in the maintenance of healthy tendons and the repair process after injury.
The pathogenesis of tendinopathy may be due to mechanical strain, which correlates well with its high incidence rate in athletes with excessive mechanical loading.16,17 TPCs have been shown to differentiate into different cell types by responding to mechanical stretch in a load-dependent manner. Under low strain, TPCs primarily differentiate into tenocytes, while they may express genes associated with chondrocyte, osteocyte, and adipocyte development when subjected to high mechanical loading.18,19
Various types of human cells change orientation by responding to cyclic uniaxial stretch.20–22 Tenocytes in intact tendons almost exclusively align themselves with the stretching direction, while cells are more randomly distributed in a healing tendon. In addition, extracellular matrix production of both tendon cells and progenitor cells was low when exposed to a randomly organized niche, compared to an aligned one.23–27 This suggests cell reorientation, followed by extracellular matrix protein deposition may play a critical role during the tendon repair process.
To further understand the pathogenesis of tendinopathy from the viewpoint of the TPCs, we hypothesize that TPCs isolated from tendinopathy may be defective in three possible areas: (i) they are fewer in number and/or have a shorter life-span compared to normal TPCs; (ii) they are less responsive to mechanical loading and fail to align with the applied force, or (iii) they have lost their capacity for tenogenic differentiation. The goal of this study is to compare the behavior of TPCs isolated from normal and tendinopathic tendons by testing these three hypotheses and provide implications for the future treatment of tendinopathy.
Materials and Methods
Sample collection
After obtaining Institutional Review Board approval and written informed consent from donors, tendon samples were collected surgically from either patellar tendinopathy patients diagnosed by magnetic resonance imaging (N = 3) or normal patellar tendons in patients undergoing bone-patellar tendon-bone anterior cruciate ligament reconstruction (N = 3). All donors were male with age ranging from 22–33 years for tendinopathy donors (27.00 ± 5.57 years) and 19–33 years for normal donors (27.33 ± 7.37 years). TPCs were isolated from the harvested tendons after the peritendinous connective tissues were completely removed before processing.
Cell isolation and culture
The wet weight of each tissue sample was recorded, minced and processed as previously described. 28 Briefly, tissues were digested in 0.1% type IV collagenase (Sigma-Aldrich, St. Louis, MO)/low-glucose Dulbecco's modified Eagle medium (Invitrogen Corporation, Carlsbad, CA) at 37°C with gentle agitation. After wash with the medium, single-cell suspension was obtained by using a cell strainer (70 μm; Becton Dickinson, Franklin Lakes, NJ). The number of viable cells was determined by NucBlue Live Ready Probes Reagent (Invitrogen Corporation) staining, according to the manufacturer's procedure. The clonogenicity and cell expansion assays were described previously. 29 Passage 2 cells were used for the all experiments, except the clonogenicity assay, which was performed with single-cell suspension isolated from tissues. All progenitor cells harvested from normal or tendinopathic tendons were used for assays in this investigation. No attempt was made to identify the origin of the progenitor cells; however, a portion of the cells could theoretically be from vascular or perivascular origin. For the purposes of this article, all progenitor cells harvested from the surgical tissues will be referred to as TPCs.
Cumulative population doublings
To determine the cell proliferation capacity of TPCs, the cumulative population doublings (CPD) were analyzed.28,30 Cells were seeded at 250 cells/cm2 in 60-mm plates in triplicates and cultured for 7 days with two media changes. Live cell density was determined by trypan blue exclusion assay using a hemocytometer. The process was repeated for a total of 17 weeks. The population doublings was calculated using the formula, (logN2 − logN1)/log2, where N1 is the initial cell number during cell seeding and N2 is the cell number at harvest. The CPD were determined by adding the population doublings for each passage to the population doublings of the previous passages.
Trilineage differentiation
The multipotency assays for progenitor cells toward chondrocytes, adipocytes, and osteocytes have been described previously.29,31,32 All staining materials were obtained from Sigma-Aldrich. After induction, chondrogenic differentiation was stained by Safranin O for the proteoglycan-rich extracellular matrix; bone induction was stained by Alizarin Red for calcium deposition, and lipid droplets from adipogenic differentiation were visualized by Oil Red O staining. To demonstrate tissue-specific mRNA expression, RNA was purified from triplicates, followed by reverse transcription (RT) with random primers (Applied Biosystems, Foster City, CA) and quantitative polymerase chain reaction (qPCR) (Applied Biosystems).29,32 The TaqMan primers (Applied Biosystems) used in this study include aggrecan (ACAN), alkaline phosphatase (ALP), type II collagen, SOX9, peroxisome proliferator-activated receptor gamma (PPAR-γ), octamer-binding transcription factor 4 (Oct4), and 18S RNA as an internal reference for comparison. The analysis of glycosaminoglycans (GAG) synthesized during chondrogenesis was described previously. 32
Cell stretch device and experiments
The mold used to make the silicone stretchable wells (Fig. 1A) was fabricated using Delrin Acetal Resin and stainless steel screws from McMaster-Carr (Santa Fe Springs). Polydimethylsiloxane (PDMS) solution (Sylgard 184 Silicone Elastomer from Dow Corning; Sigma-Aldrich) was prepared at 1:10 mixture for the silicone block and the thin layer according to the manufacturer's instruction. For surface coating, a spin coater from Specialty Coating Systems (Indianapolis, IN) was used. To create the thin layer for cell attachment, a microscope slide (75 × 38 × 1 mm; Fisher Scientific, Pittsburgh, PA) was coated with a sacrifice layer of 10% polyvinyl alcohol (Sigma-Aldrich) solution at 1000 rpm/60 s. Once dried, a thin layer of PDMS solution was coated on top of it at 500 rpm/60 s. The final stretchable culture wells were generated by bonding the well block to the thin layer on glass slide with PDMS solution. Once cured, the assembly was soaked in water to dissolve the sacrifice layer and release the final product. The stretchable culture wells were then cleaned thoroughly and autoclaved. To provide the physical force for the mechanical loading, a stretch platform on a syringe pump (Model NE-1600-U; New Era Pump Systems, Inc.) was designed from a pair of 90° angle aluminum elbows (McMaster-Carr, Santa Fe Springs, CA) attached with parallel pins as well anchors (Fig. 1B). The assembled stretch device is shown in Figure 1C. More details are shown in a close-up (Fig. 1D).
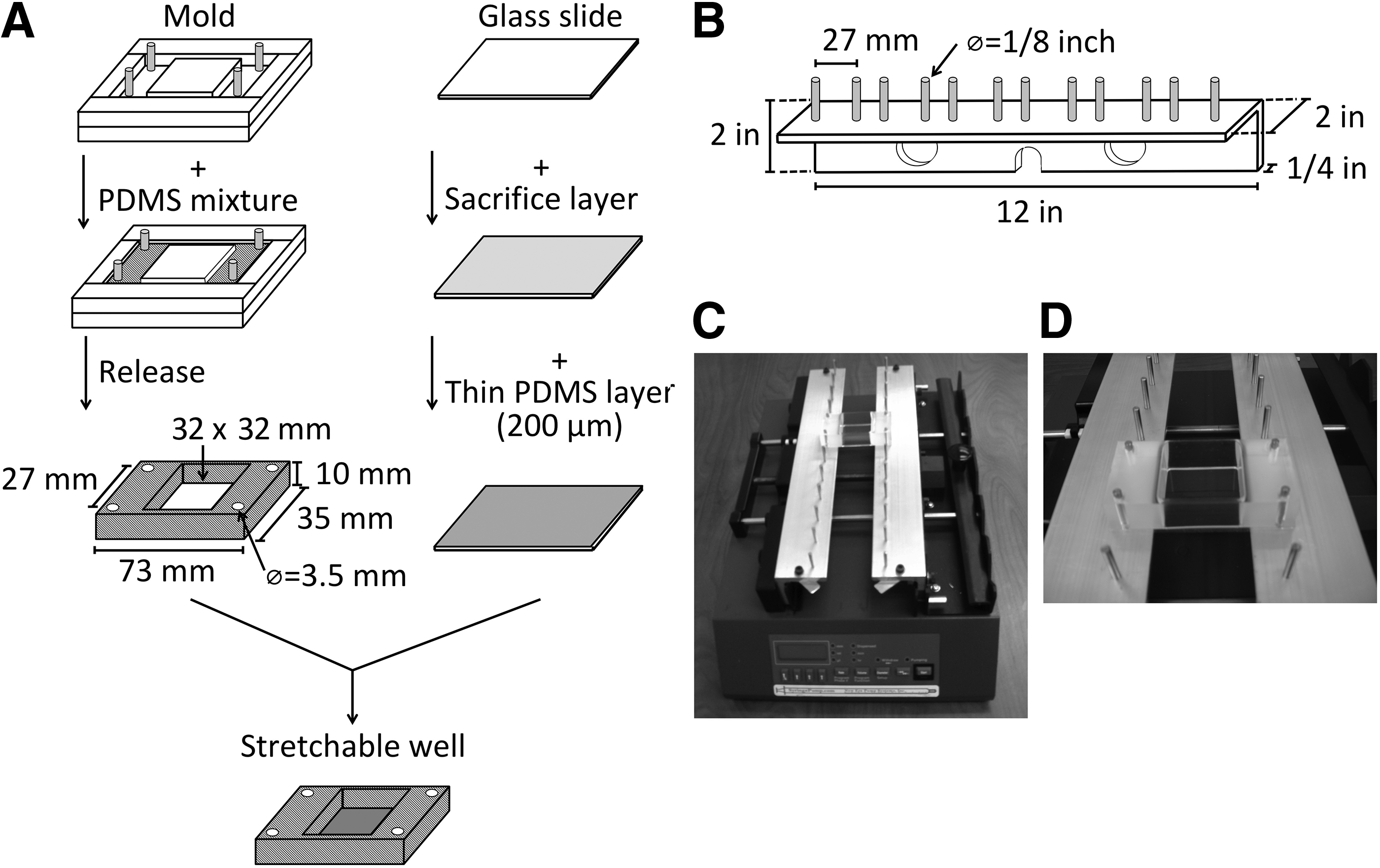
Design and fabrication of cell stretching device. The diagram shows steps to make the stretchable wells in
For cell attachment, sterile silicone wells were coated with 0.4% gelatin (Sigma-Aldrich) and incubated overnight. After drying, cells were seeded at 20 × 103 per cm2 in triplicates and cultured overnight before subjecting to cyclic stretch at moderate strength, 6% clamp-to-clamp, for the designated time period per day for 3 days with 1 day rest before harvest. At endpoints, cells were subjected to RT-qPCR assay for tenogenic differentiation markers, scleraxis (SCX), tenascin-C (TNC), and tenomodulin (TNMD). The comparative ΔΔCt method was applied for data analysis by SDS RQManager 1.2 (Applied Biosystems). Results are presented as relative fold changes to the untreated or resting controls. Note that TNMD expression was nondetectable in resting cells. By default, the last cycle number of qPCR was used as its Ct by the analysis software to calculate the fold change.
Since TNMD is a type II transmembrane glycoprotein, 33 the plasma membrane protein fraction was prepared by using Minute Plasma Membrane Protein Isolation and Cell Fractionation Kit (Invent Biotechnologies, Plymouth, MN) according to the manufacturer's instructions and subjected to western blot analysis with a TNMD antibody (Invitrogen Corporation). The same blot was stripped and reprobed with calnexin antibody (Cell Signaling Technology, Danvers, MA) as an internal control. The results of western blot analysis were presented as fold change relative to the protein levels in resting cells. Due to the limited amount of proteins available from plasma membrane, triplicate wells from each treatment condition were pooled together for the western blot analysis.
Cell orientation analysis
To examine the differences in cell orientation responding to mechanical stretch, photos were taken from nine predetermined positions in each well with Zen-Pro controlled inverted microscope (Carl-Zeiss, Thornwood, NY). The orientation of cells was quantified using the Directionality plugin in ImageJ.34,35 This plugin worked by calculating the angle of the cells relative to the stretch direction using a Local Gradients orientation method. 36 These data were then compiled into a histogram from 0° to 180° in 2° increments, and the mean, standard deviation, and percent in each increment were reported along with a Gaussian distribution as the indication of a good fit (goodness >0.90). The data were combined in 10° increments from 0° to 90° relative to the stretch direction. Then, the difference between nonstretched and stretched wells at each angle range was calculated as the final data points, where the higher number represented the increase of cells oriented in that particular angle range.
Statistical analysis
Unless it is indicated in the text, all experiments were performed in triplicates from three normal and three tendinopathy donor tissues and results are presented as mean ± standard deviation. Statistical analysis was carried out using SAS 9.4 software (SAS Institute). A p-value <0.05 was considered significant. For cell number data comparison between normal and tendinopathy groups, a two-tailed student t test was applied. In the CPD and cell orientation assay, data from each time point or angle range were compared individually between normal and tendinopathy group. qPCR and GAG content results were subjected to normality and homogeneity tests, followed by either Welch's test (osteogenesis and tenogenic differentiation following mechanical loading) and Tukey's test for p-values, or Kruskal-Wallis test (chondrogenesis, adipogenesis, and GAG content) followed by Bonferroni post-hoc adjustment for p-values. The western blot results were analyzed by a two-tailed student t test between the stretched and noninduced groups.
Results
There are more TPCs in tendinopathic patellar tendon samples than from normal tendons
The weight of tendon tissues was similar between normal (0.61 ± 0.31 g) and tendinopathy (0.65 ± 0.29 g) samples (Table 1). The mean yield of viable cells per gram of tissue from tendinopathy, 14.01 ± 0.81 ( × 105), was more than 2.6-fold higher than normal tissues at 5.33 ± 0.29 ( × 105) with a p < 0.05. The clonogenic capability (Fig. 2A) was similar, with 1.80 ± 0.12% of all viable cells for normal and 1.33 ± 0.62% for tendinopathy samples. On the other hand, the colony yield per gram of normal tendon tissue, 0.96 ± 0.63 ( × 105), was significantly lower than tendinopathic tendons at 1.57 ± 0.59 ( × 105), p < 0.001.
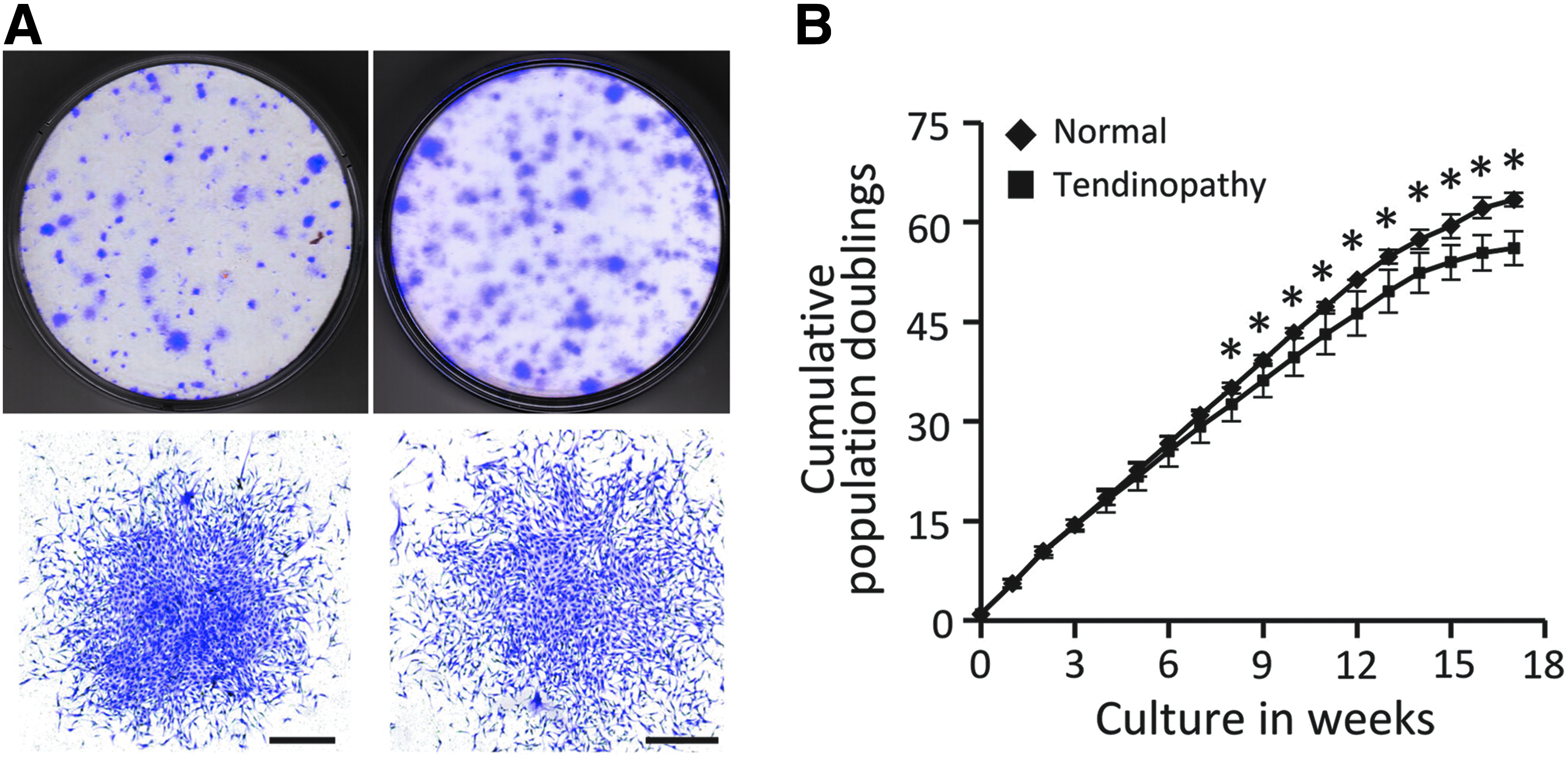
Clonogenicity and replication capacity of TPCs.
Tendon Progenitor Cells from Normal and Tendinopathy Donors
Viable cell yield was determined by cell counts with NucBlue Live Stain assay at isolation.
Colony formation was determined by colony counts with crystal violet staining after 10 days of incubation in complete media.
N.S., not significant.
Tendinopathy TPCs have a lower proliferation capacity
The proliferation rate of TPCs from tendinopathic tendons was significantly decreased starting from week 8 and continued until the end of the assay (week 18, Fig. 2B).
Expanded TPCs are multipotent
After induction, tissue-specific staining assays showed positive results for both normal and tendinopathy TPCs (Fig. 3A), suggesting a similar multipotency for tissue differentiation. In addition, GAG contents were increased significantly in both normal at 0.71 ± 0.14 (fold in logarithm, same below) and tendinopathy (1.11 ± 0.60-fold) tendons compared to uninduced controls (p < 0.05), which supports successful chondrogenic differentiation (Fig. 3B). The expression of the chondrogenic marker ACAN was dramatically increased (p < 0.005) in both induced samples. In addition, gene expression of SOX9 significantly increased by 1.30 ± 0.21-fold in normal and 1.25 ± 0.25-fold in tendinopathy samples compared to controls (p < 0.05). Type II collagen expression was also significantly elevated in normal (3.06 ± 0.98-fold) and tendinopathy (4.07 ± 0.77-fold) samples (p < 0.005). The expression of osteogenic marker, ALP, was highly expressed (p < 0.001) after bone induction in both groups. As a marker for adipogenesis, the expression of PPAR-γ was significantly elevated (p < 0.05) after induction in both groups. In contrast, the stemness marker, Oct4, was significantly reduced (p < 0.05) in all differentiation conditions in both groups. These results indicated that expanded TPCs from both normal and tendinopathic tendons were similar in multipotent, stemness properties.
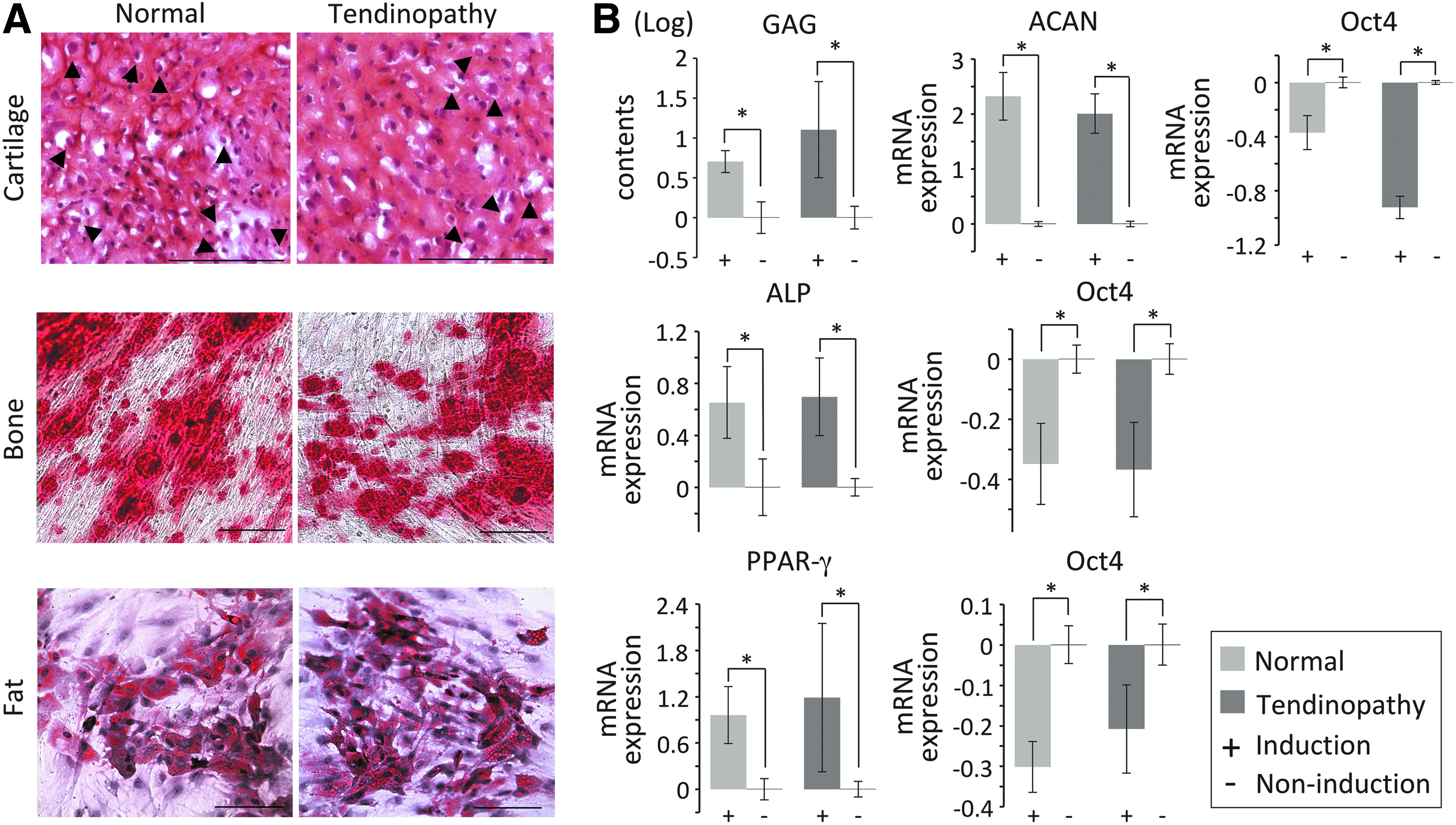
Trilineage differentiation of TPCs. Histological staining
Optimize mechanical loading for cell differentiation
In an initial pilot experiment to optimize the mechanical loading assay, one pair of normal and tendinopathy TPCs was included with 2, 4, 8, and 12 h of cyclic stretch per day. After stretch, normal TPCs showed orientation changes distinct from tendinopathy TPCs (Supplementary Fig. S1). Normal TPCs realigned themselves in a roughly perpendicular angle to the stretch direction, and such patterns increased in size along with the stretching time; however, this behavior was less evident in tendinopathy TPCs.
To assess its effect on tenocyte differentiation, we performed RT-qPCR for tenogenic markers on normal TPCs that underwent the stretching protocol versus the resting condition. The mRNA expression levels of SCX was increased from early time points, 1.54 ± 0.47-fold at 4 h, 1.29 ± 0.42-fold at 8 h, and up to 3.05 ± 1.04-fold at 12 h (Supplementary Table S1). In addition, TNMD expression increased along with the stretch time from 2.63 ± 2.38 (fold in logarithm) at 2 h, 3.20 ± 2.46 at 4 h, 2.72 ± 2.32 at 8 h, and up to 4.93 ± 0.35 at 12 h. These results suggested that a 12-h stretch duration resulted in a higher fold increase in both SCX and TNMD expression, and smaller standard deviation, at least for TNMD.
To rule out the possibility that this strain magnitude stimulated unfavorable cell differentiation, such as chondrogenesis or osteogenesis, SOX9 and ALP expression were analyzed. Along with the different time points, the expression of osteogenic marker, ALP (up to 1.49 ± 0.36-fold), and chondrogenic marker, SOX9 (up to 1.71 ± 0.41-fold), was similar to basal levels, which indicated there was no adverse cell differentiation under all conditions tested.
The same assay was then applied to tendinopathy TPCs. The expression levels of SCX in stretched samples were from 0.64 ± 0.16-fold at 2 h to 0.85 ± 0.42-fold at 12 h, and the mRNA of TNMD levels were all nondetectable, suggesting there was no tenogenic differentiation in tendinopathy TPCs after stretch. The expression of ALP (0.71 ± 0.38-fold) and SOX9 (0.85 ± 0.20-fold) at 12 h was near basal level. In summary, these results suggest that the mechanical loading assay stimulated cell rearrangement, and successfully induced tenogenic gene expression in normal TPCs in a stretch duration-dependent manner, and the 12-h time point provided the most consistent results throughout normal TPCs. Therefore, we decided to use the 12-h stretch per day for the subsequent experiments.
Tendinopathy TPCs lost capacity for tenogenic differentiation
Next, the tenogenic differentiation potential of all TPC samples was assessed by the mechanical loading assay. All normal TPCs showed widespread realignment (Fig. 4A) with significantly increased (p < 0.05) percentage of the cell population perpendicular to the stretch direction, along with a profound decline (p < 0.05) in cell numbers parallel to the direction of stretch (Fig. 4B). In contrast, TPCs from tendinopathy displayed significantly less cell rearrangement and appeared to have decreased ability to respond to the mechanical stress provided (p < 0.05, Fig. 4A, B).
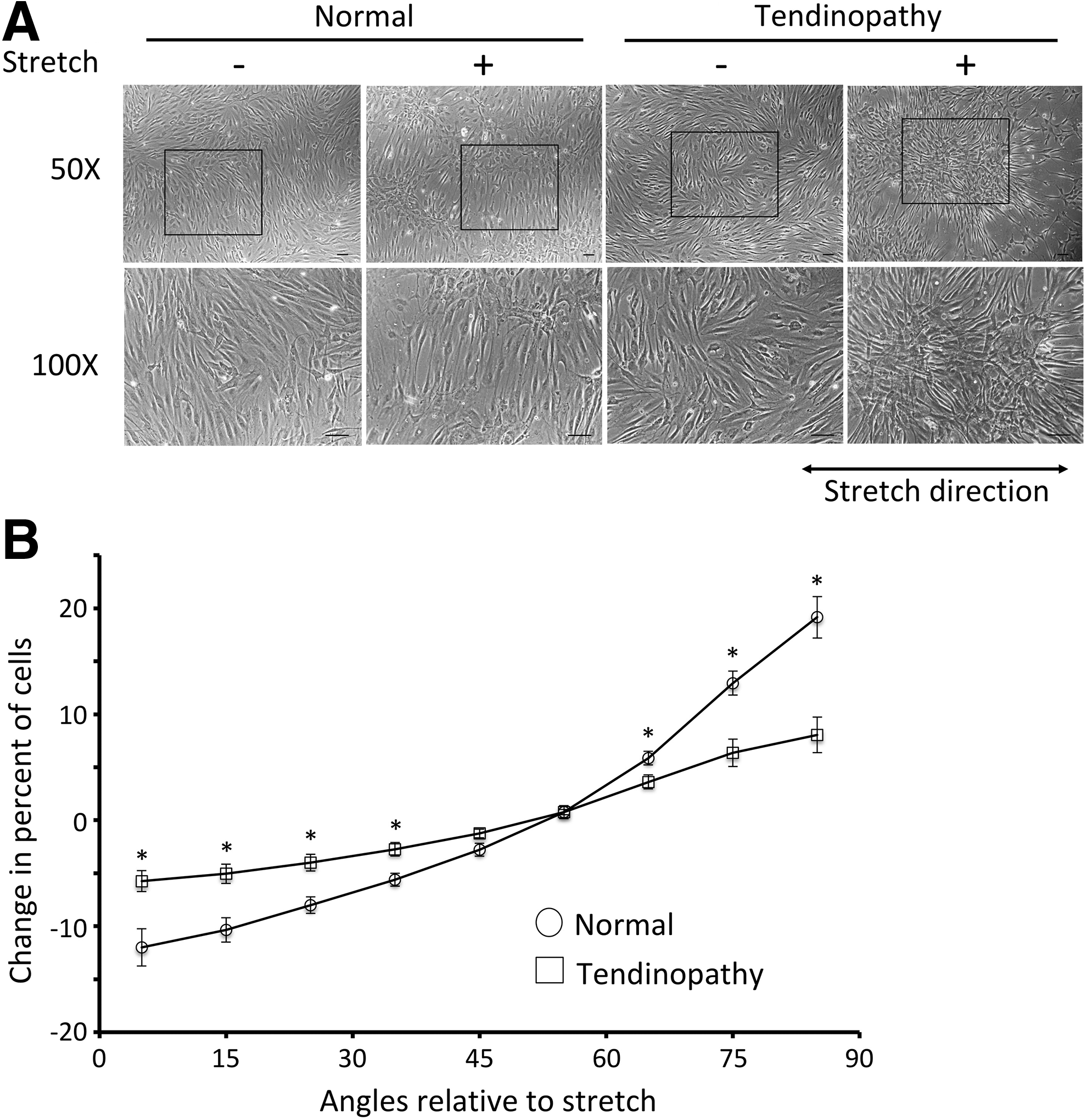
Cell reorientation after mechanical stretch.
After mechanical stretch, the TNMD expression in normal TPCs was significantly elevated at 3.86 ± 0.91 (fold change in logarithm, p < 0.001), while tendinopathy TPCs had no detectable expression (Table 2). The mRNA levels of SCX were increased in normal (2.60 ± 1.58-fold), but decreased in tendinopathy (0.58 ± 0.41-fold, p < 0.05) TPCs after stretch, while TNC content in both populations was about the basal level for normal (1.71 ± 1.01-fold) and tendinopathy (1.39 ± 0.83-fold). After stretch, the change of ALP expression was 0.90 ± 0.70-fold (normal) and 0.48 ± 0.21-fold (tendinopathy) compared to untreated cells, and chondrocyte marker SOX9 expression was 1.39 ± 0.52-fold (normal) and 0.92 ± 0.64-fold (tendinopathy). Altogether, the gene expression results revealed that normal TPCs differentiate into tenogenic lineage with profound cell realignment to external stress, while tendinopathy TPCs lacked the capacity for tenogenic differentiation with much less cell rearrangement to external stress. In both TPCs, unfavorable differentiation markers were similar to basal expression, suggesting stretching conditions were appropriate for tenogenic differentiation without adverse effects.
Relative Changes in Gene Expression Levels after Stretch
The TNMD expression levels of both normal and tendinopathy tendon progenitor cells without stretch were not detectable. The last amplification cycle number of quantitative polymerase chain reaction was used as Ct for the ΔΔCt method.
N.D., not detectable.
To further confirm the increase of TNMD expression, protein levels were analyzed following the aforementioned mechanical loading protocol. The expression of TNMD protein levels was very low in negative control (nonstretched) cells (Fig. 5A). TNMD expression was significantly elevated at 5.98 ± 1.68-fold (p < 0.01) in normal TPCs (Fig. 5B), but remained similar to baseline level in tendinopathy TPCs (0.82 ± 0.15-fold) following stretch. This result further supported the tenogenic differentiation response from normal TPCs following mechanical stretch and the lack of differentiation in the tendinopathy TPCs.
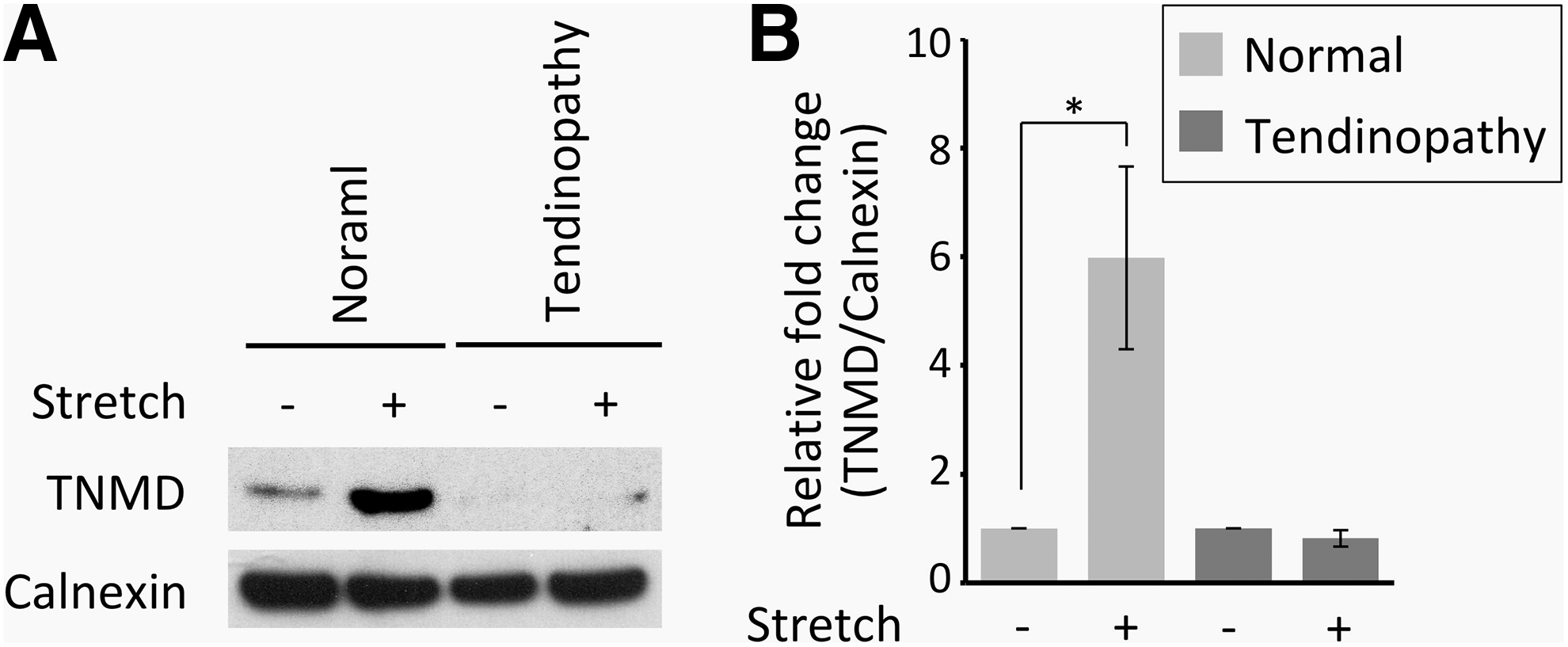
Representative results of TNMD western blot analysis from normal and tendinopathy TPCs in
Discussion
After injury, tendons initiate the repair process by mobilizing TPCs from local niches. The TPCs then migrate to the wound area and differentiate into tenocytes. Our data appear to indicate that there is no deficiency in the quantity of tendinopathy TPCs, as our tissue was harvested from surgical tendinopathy patients, and there were 1.6-fold more TPCs than in normal tendons. However, the proliferation capacity of tendinopathy TPCs was significantly lower than in normal TPCs, which may add to the tendon's insufficient healing response. The main finding of the study was the inability of the tendinopathic TPCs to differentiate into tenocytes, with no detectable expression of TNMD and low levels of SCX after mechanical loading.
To stimulate tenogenic differentiation of the TPCs, we designed and fabricated an adapter on a syringe pump to serve as a stretch platform for mechanical loading. By programming the syringe pump, this device provided suitable mechanical loading for the experiments at a fraction of the cost of a similar functioning machine from commercial sources. In addition, this design can be easily customized for flexible use to accommodate different needs.
TPCs have demonstrated response to the cyclic uniaxial stretch represented as “clamp-to-clamp” engineering strains for cell differentiation in vitro.18,37 Using the production of prostaglandin E2 as an inflammation marker, the mechanical loading of 4% on cultured tendon cells appears to be well tolerated versus 8% leads to unfavorable conditions for cells in vivo.37,38 In this study, 6% mechanical strain was used on both normal and tendinopathy TPCs, which was directly between historically known tolerances.
These results showed a significant increase of SCX and TNMD gene expression in normal TPCs after mechanical loading, but not TNC expression. TNC is tightly regulated during embryogenesis and tissue remodeling, suggesting effects from both cell type and local environment. 39 It was shown that TNC expression was increased in cultured fibroblasts or tendon stem cells on three-dimensional scaffolds under mechanical stress,40,41 but there are only a few studies using similar conditions as this study.42–44 In these studies, the increase of TNC gene expression occurred with higher mechanical strains at 8% or 10%, but not at 4%. The magnitude of TNC gene induction was from 1.5- to 1.8-fold in the previous reports, which is similar to our results at 1.71-fold increase in normal TPCs after stretch. Overall, these results support the notion that the expression of TNC may be subject to tight regulation, affected by both cell type and local environment, which may have limited the induction of TNC expression in our mechanical loading protocol.
As the major load-bearing component of tendons, type I collagen expression is highly anticipated along with tenogenic differentiation. In contrast to this, previous studies showed the decrease of type 1A2 collagen expression of mesenchymal stem cells stimulated for tenogenesis by the decellularized tendon scaffold alone, 45 or with cyclic stretching. 46 Therefore, the negative feedback mechanism for collagen expression was raised to separate it from other positive gene markers for tenogenic differentiation in those studies. In this study, gelatin was chosen for the culture surface preparation of stretchable silicone wells due to the easy handling, access, and cost effectiveness. As the mixture of collagen proteins, gelatin may behave similar to decellularized tendon scaffold and impact the expression of type I collagen during mechanical loading-induced tenogenesis. Therefore, the gene expression assay for type I collagen was excluded from this study. Nevertheless, the increase of SCX and TNMD following mechanical loading clearly showed the tenogenic differentiation of normal TPCs. A non-collagen coating agent may be more suitable for this study model in the future to address type I collagen expression in tenogenesis following mechanical loading.
In this study, TPCs from tendinopathy specimens displayed a similar potency for trilineage differentiation as normal TPCs. Therefore, the deficiency for tenocyte differentiation following mechanical loading may well be tendon lineage specific. To identify the affected genes and signaling pathways in tendinopathy TPCs, the comparison of gene profile between normal and tendinopathy TPCs after stretch is currently underway. In addition to mechanical stimulation, recent studies showed multiple factors were involved in the regulation of tenogenic differentiation from various stem or progenitor cells.47,48 These include growth factors (e.g., transforming growth factor-beta, bone morphogenetic protein (BMP)-12, BMP-14, and connective tissue growth factor), biomaterials (scaffolds, aligned collagen fibrils, and decellular matrix from tendon), and OrthoBiologics (bone marrow aspirate concentrate and platelet-rich plasma [PRP]).25,49–56 In particular, the leukocyte-rich PRP treatment for tendinopathy showed some promising potential in few double-blind randomized clinical trials.57–60 It may be possible in the future to prime tendinopathy TPCs with a stretch device and the aforementioned factors to alter gene expression, and once again allow TPCs to be responsive to mechanical loading and be inducible toward tenogenic differentiation.
In summary, TPCs harvested from patellar tendinopathy patients did not demonstrate the capacity for tenogenic differentiation and displayed decreased responsiveness to external mechanical forces compared to normal tendons. The scientific method provided in this article will allow testing of biologics and other treatment strategies to derive new, more efficient treatments for tendinopathy in the future.
Footnotes
Acknowledgments
The authors gratefully acknowledge Anthony Behn and Timothy Thio in the Department of Orthopaedic Surgery, School of Medicine, Stanford University, for help with the fabrication of molds and stretch platforms.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.