
Abstract
Adipose-derived mesenchymal stromal/stem cells (ASCs) represent a commonly used cell source for adipose tissue engineering. In this context, ASCs have routinely been cultured in conventional 2D culture and applied as single cell suspension for seeding onto scaffold materials or direct injection. However, this approach is associated with the loss of their intrinsic 3D microenvironment and leads to impaired regenerative capacity of the cells. Thus, the application of ASCs as self-assembled 3D spheroids with cells residing in their own matrix is an attractive alternative. However, characterization of the structural features and differentiation capacity of the spheroids is necessary to effectively apply them as building blocks in adipose tissue engineering. In this study, we focus on extracellular matrix (ECM) development in ASC spheroids, as well as adipogenic differentiation in comparison to conventional 2D culture using different induction protocols. Reproducible assembly of ASCs into spheroids was achieved within 24 h using the liquid overlay technique. Undifferentiated spheroids displayed a stromal ECM pattern, with fibronectin, collagen V, and VI as the main components. In the course of adipogenesis, a dynamic shift in the ECM composition toward an adipogenic phenotype was observed, associated with enhanced expression of laminin, collagen I, IV, V, and VI, similar to native fat. Furthermore, adipogenic differentiation was enhanced in spheroids as compared with 2D cultured cells, with the spheroids needing a distinctly shorter adipogenic stimulus to sustain adipogenesis, which was demonstrated based on analysis of triglyceride content and adipogenic marker gene expression. In summary, culturing ASCs as spheroids can enhance their adipogenic capacity and generate adipose-like microtissues, which may be a promising cell delivery strategy for adipose tissue engineering approaches.
Impact statement
Adipose-derived mesenchymal stromal/stem cells (ASCs) as a widely used cell source for adipose tissue engineering have been shown to be limited in their regenerative capacity when applied as single cells. As an alternative approach, the delivery as spheroids, consisting of cells in a 3D context, may be favorable. However, insights into extracellular matrix (ECM) development and efficient adipogenic differentiation are required for their effective application. In this study, we show that differentiated ASC spheroids develop an ECM, resembling native adipose tissue. Furthermore, the ASC spheroids exhibited a superior differentiation capacity as compared with conventional 2D culture, and required only a short adipogenic induction stimulus. Our results identify ASC-derived spheroids as an attractive cell delivery method for adipose tissue engineering approaches.
Introduction
Multipotent mesenchymal stromal/stem cells (ASCs) isolated from adipose tissue are widely used in tissue engineering applications due to their multilineage potential and easy harvest. ASCs have extensively been studied for adipose tissue regeneration to address the tremendous clinical need for the development of new strategies to repair soft tissue defects resulting from traumatic injury, tumor resection, and congenital defects.1–7 In current ASC research, traditional 2D culture is widely used, which involves separating cells from their original microenvironment. In addition, for tissue engineering, cells expanded in 2D cultures are commonly seeded onto biomaterial-based scaffolds, which in many cases, at least initially, also represent a 2D environment, as the cells largely grow in 2D on the scaffold surfaces with limited cell-cell contacts. 8 As innate properties such as the differentiation capability of ASCs are greatly influenced by their microenvironment with its spatially arranged cell-cell and cell-matrix interactions, cells in a 2D environment may be limited in their regenerative capacity.9,10
In contrast, the self-assembly and culturing of cells in a 3D context has been shown to create a more in vivo-like microenvironment allowing cells to interact with each other within their secreted extracellular matrix (ECM) rather than with an artificial surface. 11 Culturing and delivery of various cell types as spheroids has been demonstrated to lead to an enhancement of their regenerative potential, associated with superior differentiation ability, augmented secretion of angiogenic and anti-inflammatory cytokines, and enhanced integration and retention after transplantation in vivo.12–14 ASCs form multicellular 3D aggregates, when cultured in suspension or in a nonadhesive environment and can be cultivated as spheroids over prolonged periods of time. 8 Through the implementation of tissue-specific cultivation methods, for example, adipogenic induction, the generation of histotypic microtissues is feasible. 15 In this context, the development of a tissue-specific ECM is an intrinsic feature, as in addition to providing physical support, the ECM actively contributes to the establishment and maintenance of differentiated tissues and is part of the complex communication network of cells in their microenvironment. 16 As such, the proper development of a tissue-specific ECM must be considered as an important requirement in tissue engineering approaches. However, although several studies were done on spheroids consisting of adipogenically differentiated ASCs or 3T3-L1-preadipocytes, they have not addressed the ECM composition and remodeling during the adipogenic differentiation process in these 3D cultures.17–20
In general, few studies have investigated ASC-derived spheroids for use in adipose tissue engineering.21,22 In a study conducted by Zhang et al., ASC-derived spheroids, generated in poly(lactic-co-glycolic acid) (PLGA) scaffolds, were shown to trigger adipose tissue formation in vivo. In that study, the spheroids were cultured in adipogenic induction medium before implantation employing a standard long-term induction protocol, which is often used for 2D cultures. 21 This underlines, in principle, the potential beneficial effects of adipogenic induction and precultivation of ASCs in vitro for adipose tissue formation in vivo. 1
For the effective application of ASC-derived spheroids for adipose tissue engineering, there is a need for further insights into ECM development and the establishment of methods for efficient adipogenic induction. Therefore, in the present study, we investigated the ECM composition of ASC-derived spheroids and the remodeling during the adipogenic differentiation process. The ECM composition of differentiated spheroids was compared with native human fat to demonstrate the resemblance between the ECM of the two tissues. Furthermore, adipogenic induction in 3D spheroids was investigated in comparison to conventional 2D culture. Specifically, different induction protocols were evaluated, showing that ASCs in 3D spheroids needed a distinctly shorter adipogenic stimulus to effectively differentiate into adipocytes as compared with 2D cultured cells. The results of this study may further the use of ASC-derived spheroids for soft tissue engineering.
Materials and Methods
2D and 3D cell culture
Human ASCs (hASCs) were purchased from Lonza (Walkersville; human adipose-derived stem cells). For expansion, cells were cultured in basal medium consisting of Dulbecco's modified Eagle's medium/Ham's F-12 (DMEM/F-12) (Invitrogen, Karlsruhe, Germany), supplemented with 10% fetal calf serum (FCS; Invitrogen) and 1% penicillin/streptomycin (Invitrogen) at 37°C and 5% CO2 until confluence. Culture medium was changed every other day. For passaging or harvesting cells for subsequent experiments, 0.25% trypsin-EDTA (Invitrogen) was used. Cells at passage 4 or 5 were used for subsequent experiments.
For 2D culture conditions, ASCs were seeded at a density of 25,000 cells/cm2 and cultured in growth medium consisting of Preadipocyte Basal Medium-2 (Lonza) containing 10% FCS and 1% penicillin/streptomycin. For 3D culture, multicellular spheroids were generated from the ASCs using the liquid overlay technique. More specifically, after coating 96-well plates with 1.5% agarose diluted in DMEM/F-12, ASCs were seeded at 5000 cells/well and cultured in growth medium on an orbital shaker at 50 rpm. To prevent loss of spheroids during medium changes, only half of the medium volume was replaced with each medium change. Cells in 2D cultures were treated the same way for consistency.
The spheroid assembly process was monitored by microscopy at different time points with an Olympus IX51 inverted light microscope equipped with a XC30 digital camera (Olympus, Hamburg, Germany). The diameter of the spheroids was assessed using the Olympus cellSense™ Dimension Microscope Imaging Software.
Adipogenic differentiation of 3D spheroids and 2D monolayers
After 2 days cultivation in growth medium, adipogenic differentiation was induced (start of induction, designated day 0) by exchanging half of the medium with adipogenic differentiation medium (DM), consisting of growth medium with the hormonal inducers insulin (final concentration 1.7 μM; PromoCell, Heidelberg, Germany), dexamethasone (1 μM; Sigma-Aldrich, Steinheim, Germany), 3-isobutyl-1-methylxanthine (500 μM; Serva-Electrophoresis, Heidelberg, Germany), and indomethacin (200 μM; Sigma-Aldrich). Two different induction regimes were performed under 2D and under 3D culture conditions. For permanent induction, the ASCs were cultured in DM for 9 days until harvest. For short-term induction, cells were treated with DM for 2 days and then the DM was replaced by maintenance medium (MM), consisting of growth medium supplemented with 1.7 μM insulin. When first adding the MM, half of the medium was exchanged for three times to minimize the concentration of the hormonal inducers. Noninduced control groups were cultured in growth medium during the whole culture time.
Live/dead staining
Cell viability within the 3D spheroids was analyzed using live/dead staining (PromoKine, Heidelberg, Germany) according to the manufacturer's instructions. Images were captured using a TCS-SP2 AOBS Leica confocal laser scanning microscope (CSLM) together with the Leica LCS Lite Software (Leica, Wetzlar, Germany).
Histological analysis of adipogenic differentiation
For visualization of intracellular lipid accumulation within differentiating cells, 2D monolayers and cryosections of spheroids were stained with Oil Red O (Sigma-Aldrich) and cell nuclei were counterstained with Hematoxylin (Bio Optica, Milan, Italy). Microscopical images of stained 3D spheroid sections were taken using a DP71 digital camera (Olympus) attached to an Olympus BX51 microscope. Two-dimensional monolayers were imaged with an Olympus IX51 inverted light microscope equipped with a XC30 digital camera (Olympus).
Quantification of intracellular triglyceride and DNA content
Quantitative analysis of intracellular lipid accumulation was performed with the Serum Triglyceride Determination Kit from Sigma-Aldrich. Two-dimensional monolayers and 3D spheroids were harvested and sonicated with an ultrasonic homogenizer (Sonopuls; Bandelin electronic, Berlin, Germany) in 0.5% aqueous Thesit solution (0.5% Thesit in H2O; Gepepharm, Hennef, Germany). The assay was carried out according to the manufacturer's instructions and measured with a Microplate ELISA-Reader (Dynatech Laboratories, El Paso) at 570 nm. Calculated triglyceride contents were normalized to the DNA content of the respective samples. For determination of total DNA content cells in 2D monolayers, and 3D spheroids were harvested in phosphate/saline buffer (50 mM phosphate buffer, 2 mM Na2EDTA ·2 H2O, 2 M NaCl, pH 7.4; all obtained from Carl Roth, Karlsruhe, Germany) and sonicated with an ultrasonic homogenizer. DNA content was determined using the intercalating dye Hoechst 33258 (Polysciences, Warrington) by measuring fluorescence intensities at an excitation wavelength of 365 nm and an emission wavelength of 458 nm with a spectrofluorometer (Tecan GENios pro; Tecan, Crailsheim, Germany).
Quantitative reverse transcription polymerase chain reaction
For gene expression analysis, total RNA from 2D cultured cells and 3D spheroids was harvested with TRIzol® reagent (Invitrogen). cDNA was synthesized using the IMProm-II™ Reverse Transcription System from Promega (Mannheim, Germany) followed by real-time PCR using the MESA GREEN qPCR MasterMix Plus MeteorTaq polymerase (Eurogentec; Seraing, Belgium). quantitative real time-polymerase chain reaction was carried out with the corresponding QuantiTect® Primer Assays from Qiagen (Hilden, Germany) for the adipogenic transcription factors peroxisome proliferator-activated receptor gamma (PPARγ) and CCAAT/enhancer-binding protein alpha (C/EBPα), as well as for the adipocyte protein 2/fatty acid-binding protein 4 (aP2/FABP4) using the following cycling conditions: 95°C for 15 min followed by 40 cycles of 95°C for 15 s, 55°C for 30 s, and 72°C for 30 s. A melting curve analysis for PCR product integrity was performed. mRNA expression levels were normalized to the eukaryotic translation elongation factor 1 alpha (EF1α). 23 The 2−ΔΔCT method was used to determine the relative expression levels. The relative expression values were further normalized to the respective day 0 sample of 2D cultures.
Immunofluorescent staining of extracellular matrix components
For immunohistochemical analysis of major ECM components, cryosections of spheroids and native tissue were stained with primary antibodies against collagen I (Col I; ab34710; polyclonal rabbit anti-Col I), collagen IV (Col IV; ab6586; polyclonal rabbit anti-Col IV), collagen V (Col V; ab7046; polyclonal rabbit anti-Col V), collagen VI (Col VI; ab6588; polyclonal rabbit anti-Col VI), laminin (Lam; ab11575; polyclonal rabbit anti-Lam), as well as fibronectin (FN; ab2413; polyclonal rabbit anti-FN), all of which were purchased from Abcam (Cambridge, United Kingdom). After blocking with 1% bovine serum albumin (Sigma-Aldrich) for 20 min at room temperature, sections were incubated with the primary antibodies overnight in appropriate dilutions (Col I 1:800; Col IV 1:150; Col V 1:200; Col VI 1:200; Lam 1:200; FN 1:500). After a washing step with PBS, the secondary antibody was applied for 1 h (Cy3-conjugated AffiniPure goat anti-rabbit, 1:400; Dako, Hamburg, Germany). Spheroid cryosections were optionally stained with BODIPY® 493/503 (0.01 mg/mL in DMSO; Sigma-Aldrich) for lipid droplet visualization. Sample imaging was performed with an Olympus BX51 microscope in combination with the Olympus cellSense Dimension Microscope Imaging Software.
Statistics
Quantitative results are presented as mean value ± standard deviation. Statistical significance was assessed by analysis of variance (one-way or two-way analysis of variance) with subsequent multiple comparisons according to the Bonferroni post hoc test. A value of p < 0.05 was considered statistically significant. Statistical analysis was performed using GraphPad Prism 6 software.
Results
Formation of ASC-derived spheroids
The self-assembly of ASCs was induced by seeding the cells in agarose-coated well plates according to the liquid overlay technique. The nonadhesive surface forced the cells to attach to each other resulting in multicellular aggregates. Using an initial cell number of 5000 cells per well, one single spheroid yielding a densely packed rounded structure that was 320 μm (±8 μm) in diameter was formed per well. Live/dead staining of fully compacted spheroids (48 h after seeding of the cells) demonstrated the viability of cells within the spheroids (Fig. 1A). Thus, the applied technique allows for the generation of viable ASC cell aggregates in a well-defined and highly reproducible manner. To monitor the assembly process, images of assembling spheroids were taken at various time points after seeding (Fig. 1B). Two hours after seeding, the cells formed a loose aggregate, which progressively underwent compaction during the next hours. The assembly process was completed after 24 h, with the spheroids having a round compact shape. This process was reflected by the reduction of the spheroid diameter over time, illustrating a sharp decline during the first 8 h after seeding with no further reduction in spheroid diameter after 24 h (Fig. 1C). The ECM composition of fully compacted spheroids was examined using immunohistochemical staining (48 h after seeding). In this state, the spheroids displayed a stromal ECM pattern with fibronectin, Col I, Col V, and Col VI as the major ECM components (Fig. 1D). Laminin and Col IV as adipose tissue-specific ECM components were not detectable at this time point, which also represented the time point of adipogenic induction in the subsequent studies (Fig. 1D).
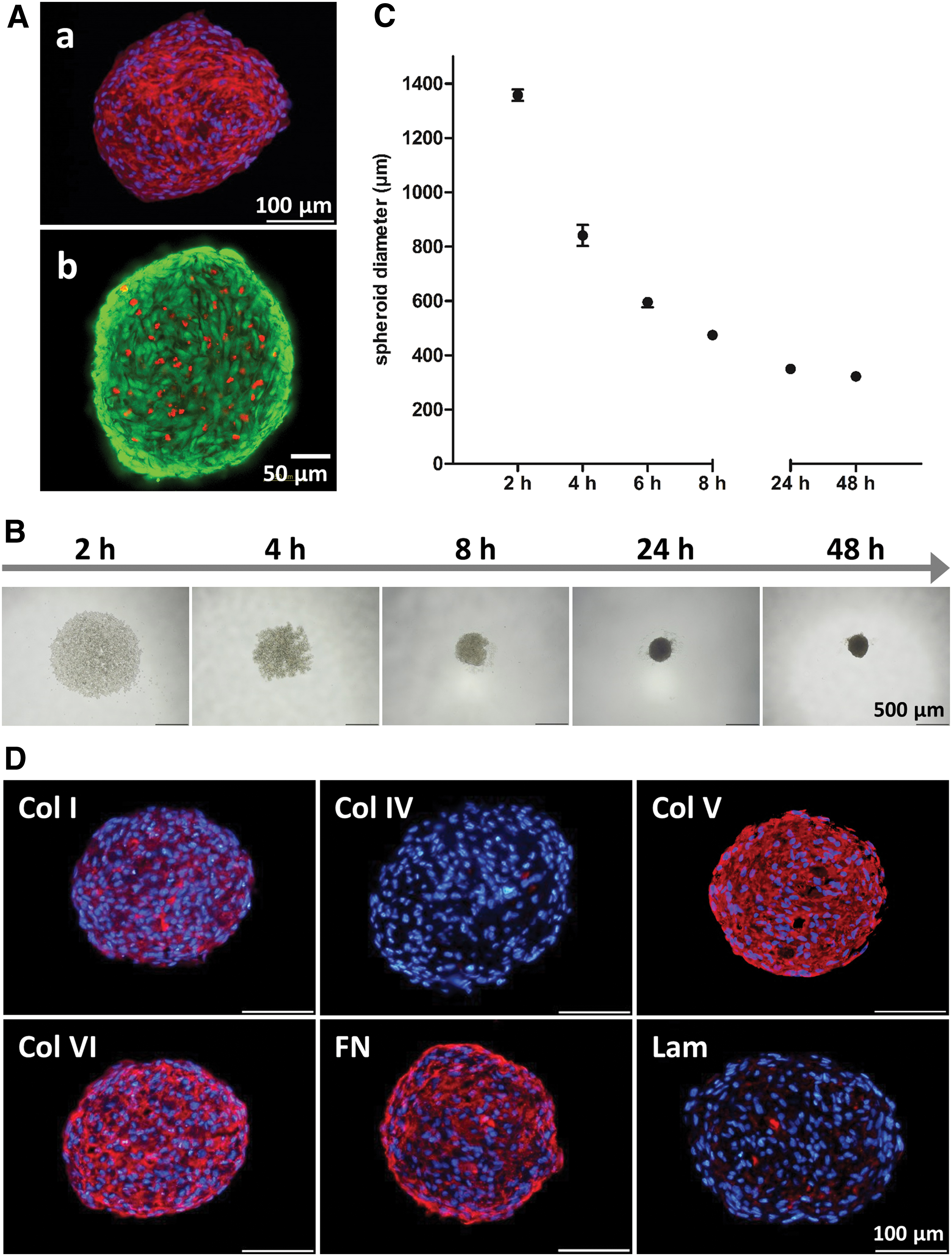
Characterization of human ASC spheroids.
ECM remodeling during adipogenic differentiation and comparison to native human adipose tissue
Next, we analyzed the adipogenic differentiation of the ASCs, as well as ECM remodeling during the differentiation process in spheroid culture. To this end, spheroids were transferred to adipogenic DM at 48 h after seeding. After 9 days in culture, the ASCs in the 3D spheroids had markedly differentiated along the adipogenic lineage as confirmed by Oil Red O staining of accumulated triglycerides. In noninduced spheroids, triglyceride accumulation was not detectable (Fig. 2A). The ECM composition of differentiated ASC-derived spheroids and their undifferentiated counterparts was characterized by immunohistochemical staining of major ECM components (Col I, Col IV, Col V, Col VI, fibronectin, and laminin) and compared with the ECM composition of native human fat tissue (Fig. 2B). Spheroids cultured in growth medium (noninduced) retained their baseline ECM composition, consisting predominantly of fibronectin, Col V, and Col VI. In contrast, spheroids cultured in adipogenic DM (induced) developed an ECM pattern associated with high expression of Col IV and laminin, which are integral parts of the basement membrane surrounding adipocytes, while fibronectin was markedly downregulated as compared with the noninduced controls. Col I and Col VI were found to be expressed more prominently in differentiated spheroids as compared with the undifferentiated spheroids. In addition, collagen V was expressed in the induced spheroids. The comparison with the ECM pattern of native human adipose tissue revealed a distinct resemblance with the ECM of differentiated spheroids. Col I, Col IV, Col V, Col VI, and laminin were present in native fat as well as in the differentiated ASC-derived spheroids, while fibronectin was expressed at low levels in both the adipose tissue and in induced spheroids.

Adipogenic differentiation and ECM development during the differentiation process in ASC spheroids and comparison to native human adipose tissue.
To explore a chronological sequence of ECM remodeling and adipogenic differentiation in the spheroids, accumulating triglycerides and laminin as one major component of the adipose-specific ECM were stained at various time points (d0, d2, d5, d7, d9) during the differentiation process (Fig. 3). Accumulating triglycerides, as detected by BODIPY staining, became visible starting at day 7, with an increase in staining intensity observed at day 9. By costaining for laminin at these time points, it became evident that laminin production preceded the deposition of lipid droplets, as a distinct laminin signal could be detected as early as day 2 after adipogenic induction, with a further increase in signal intensity during the ongoing differentiation process.

Development of laminin expression during adipogenic differentiation of 3D spheroids. To visualize progression of adipogenesis, accumulating triglycerides were stained with BODIPY (green) at various time points and laminin deposition was monitored by immunohistochemical staining (red). Nuclei were counterstained with DAPI (blue). Scale bars represent 100 μm. Color images are available online.
Effect of short-term adipogenic induction on adipogenesis in 2D cultures and 3D spheroids
To refine the adipogenic induction protocol for efficient differentiation, that is, reduction of induction time, spheroids were adipogenically induced using different culture regimes. ASCs in fully compacted spheroids (48 h after seeding) were induced to undergo adipogenesis using a standard induction protocol with permanent application of the adipogenic hormonal cocktail in the adipogenic DM during the complete differentiation period (9 days), or alternatively, a short-term induction regime was employed with culture in adipogenic DM for only 2 days followed by culture in MM for 7 days (Fig. 4A).

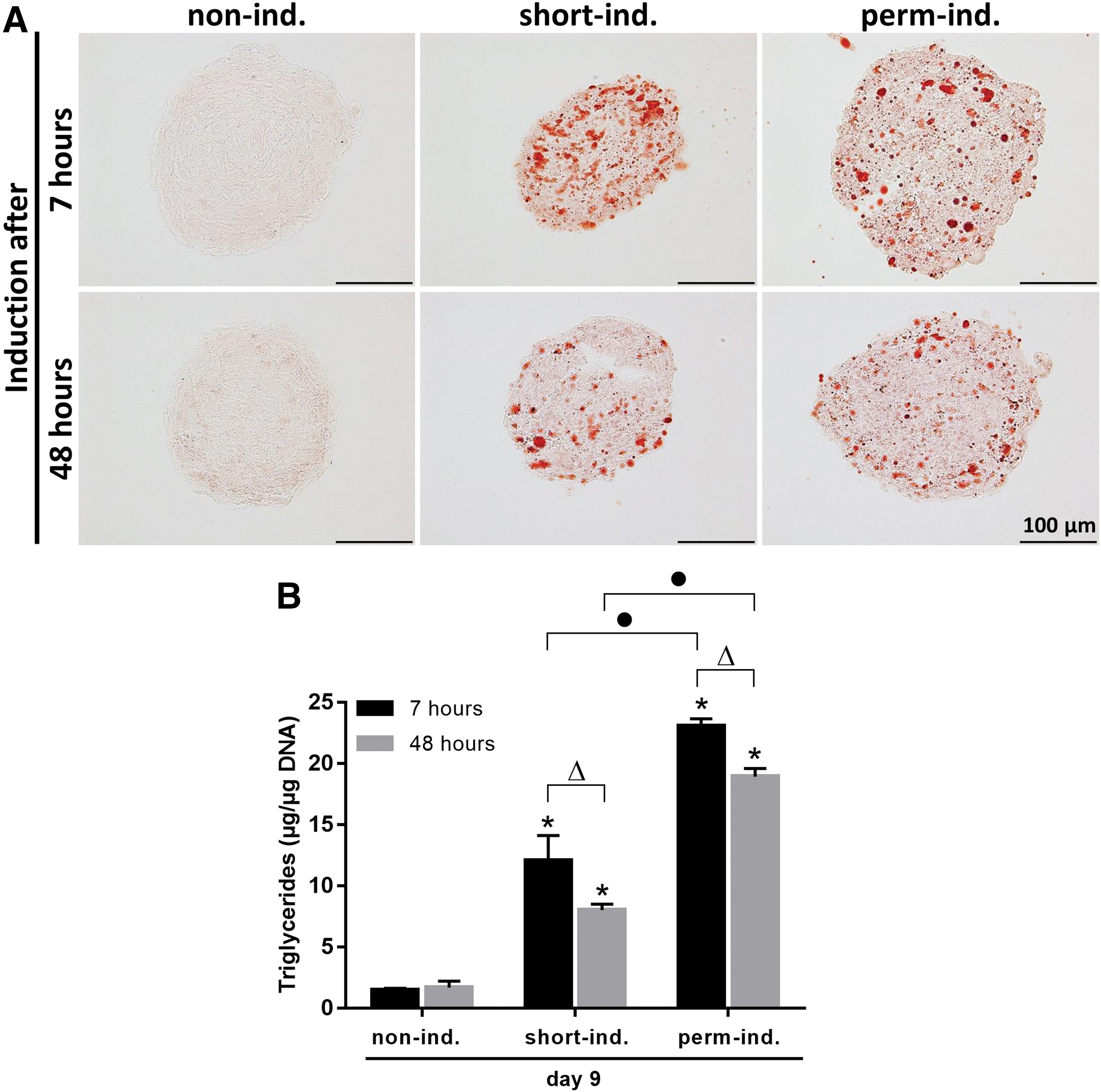
Characterization of adipogenic differentiation of ASCs in 2D monolayer and 3D spheroid culture with different induction protocols (permanent and short-term induction).
Adipogenically induced spheroid cultures were compared with induced conventional 2D cultures, and 3D spheroids and 2D monolayers cultured in growth medium during the whole cultivation period served as noninduced controls. Adipogenesis under the different induction conditions was assessed by quantification of accumulated triglycerides at day 9 (Fig. 4B). With permanent induction, the cells in the 2D cultures and spheroids differentiated along the adipogenic lineage, as shown by significantly elevated triglyceride levels compared with the noninduced controls. ASCs in spheroids displayed enhanced differentiation, reflected by a higher triglyceride content per cell under this condition. With the short-term induction regime, only the ASCs cultured as spheroids underwent adipogenesis, whereas the triglyceride content in the cells in 2D monolayers remained at the same level as the noninduced controls.
Histological analysis confirmed these results (Fig. 4C). Only a minimal number of lipid droplets could be detected in the cells that were cultured in monolayers, whereas in short-term-induced spheroids, triglyceride accumulation was comparable to the accumulation observed in cells cultured in adipogenic DM for 9 days. To investigate whether the observed differences in adipogenic capacity between the 2D and 3D cultures were also reflected in terms of adipogenic gene expression, quantitative reverse transcription PCR (qRT-PCR) was performed to analyze the expression of PPARγ and C/EBPα, major transcription factors in the adipogenic differentiation process, and aP2/FABP4, a marker gene of terminal adipogenic differentiation. In spheroids, expression of PPARγ and C/EBPα was distinctly increased under short-term as well as permanent induction, as compared with 2D monolayers. A similar expression pattern was observed for aP2/FABP4.
Overall, these results demonstrated that ASCs cultured as 3D spheroids required a shorter stimulus to undergo adipogenic differentiation and showed enhanced differentiation as compared with ASCs in conventional 2D monolayer culture.
Adipogenic differentiation of ASC-derived spheroids after early and late onset of induction
To further improve the precultivation regime for the application of spheroids in adipose tissue engineering, we varied the time point for adipogenic induction. For this purpose, cell aggregates were induced during the assembly process (7 h after seeding) or after final compaction (2 days after seeding), with either the short-term induction or the permanent induction regime. The application of the adipogenic hormonal cocktail during assembly did not impact the compaction process of the spheroids (Supplementary Fig. S1). Histological staining of accumulated triglycerides with Oil Red O indicated differentiation along the adipogenic lineage of ASC-derived spheroids both under short-term and permanent induction regardless of the time point of induction (Fig. 5A). The quantification of accumulated triglycerides confirmed the histological observations and indicated that differentiation within the ASC spheroids induced during assembly (after 7 h) was significantly enhanced as compared with the induction of fully compacted spheroids (induced after 2 days) (Fig. 5B).
Adipogenic differentiation of 3D spheroids with different starting time points of induction. Spheroids were induced after final assembly and compaction (48 h after seeding) or during the assembly process (7 h after seeding) and treated with short-term or permanent induction.
Discussion
Spheroids generated from adipose stromal/stem cells appear eminently suited as cell delivery platform for tissue engineering approaches, as they can be implanted as microtissues with cells residing and interacting within their endogenous ECM, which has been reported to enhance cell survival, cell retention, and functional performance after transplantation.24–26 As preassembled, biomimetic 3D structures, ASC spheroids can further serve as building blocks in biofabrication applications. 27 In this context, spheroids have been proposed for the use in microneedle-based (Kenzan) spheroid assembly, 28 bioprinting-assisted spheroid fusion, 29 spheroids combined with direct polymer melt deposition, 30 and caging within 2PP-fabricated “lockyballs”. 31 In a recent study, sheet-like structures were generated using ASCs, which were incorporated as 3D spheroids in tailored melt electrowritten cage-like scaffolds. 32 However, as cells in a 3D context behave differently compared with cells originating from well-explored conventional 2D cultures, a profound characterization of spheroids regarding their structure and cellular behavior is mandatory for their effective application. In the context of adipose tissue engineering, there has been limited research on ASC spheroids and the in-depth characterization of these 3D microtissues has not previously been reported. In this study, we specifically addressed the ECM composition and remodeling in ASC spheroids, investigated different culture regimes for effective adipogenic induction, and demonstrated the suitability of these cell aggregates to build adipogenic microtissues upon a short adipogenic stimulus.
For spheroid production, several models have been described to force cellular aggregation of ASC. Low-adhesive materials such as polydimethylsiloxane (PDMS) 33 or synthetic hydrogels in combination with distinct spatial surface patterns or pores21,34 have been used for the generation of aggregates of defined size and shape. Chitosan films have been reported to promote cellular aggregation of ASCs, however, aggregation on these planar films resulted in considerable size heterogeneity. 17 The generation of cellular aggregates in suspension cultures under dynamic conditions, for example, in spinner flasks 35 or microgravity reactors, 18 is an attractive solution for large-scale production, however, does not allow for any control of size and shape or the assembly process. In this study, the liquid overlay technique36,37 was used to generate ASC-based spheroids. The seeding of defined numbers of cells in microwells provoked the generation of one spheroid per well in a highly controlled manner with regard to the assembly process as well as size and shape of the resulting spheroids, which is favorable for studies on basic characteristics of these cell aggregates.
According to the specific demands on the extracellular environment of differentiating cells, their ECM constantly undergoes dynamic changes and has been recognized to play a critical role for the differentiation process.38,39 To date, the composition and remodeling of the ECM during adipogenic differentiation has been investigated in 2D cultures for bone marrow-derived MSCs, 3T3-L1 preadipocytes, and bovine intramuscular preadipocytes.40–44 We have now sought for the first time to elucidate the ECM composition and its remodeling during adipogenesis in 3D culture within ASC spheroids, which as 3D culture displays the ECM in a more in vivo-like setting compared with conventional 2D culture. In compacted, undifferentiated spheroids, fibronectin, and collagen I, V, and VI were found to be the most prominent ECM components, consistent with a stromal-like composition, with limited expression of adipose-specific components such as laminin or collagen IV.
Upon hormonal induction, ASC spheroids were shown to differentiate effectively along the adipogenic lineage within 9 days. During adipogenesis, ECM remodeling was evident, with the development of typical features of an adipose-specific ECM, with collagen IV and laminin being abundantly expressed in the adipocyte basement membrane.45,46 In addition, enhanced expression of collagen I and VI was shown in the differentiated 3D spheroids as compared with their undifferentiated counterparts. These findings are in accordance with studies in 2D cultures, which report an increase in collagen VI expression during the differentiation process of human mesenchymal stem cells (hMSCs).41,42 Collagen VI is a nonfibrillar collagen, distributed widely in tissues that forms thin microfilaments and binds to various other ECM components, including collagens I, II, and IV, and FN. 45 Collagen I as part of the supporting ECM framework was previously described in adipogenically differentiated hMSCs. 40 Collagen V is a minor fibrillar collagen, which forms heterotypic fibrils with Col I. The observed expression of Col V in noninduced and induced ASC spheroids as well as in native adipose tissue is in accordance with previous reports describing the production of Col V in both undifferentiated and differentiated 3T3-L1 preadipocytes and ASCs in 2D culture, and in native adipose tissue.43,45,47,48 In the present study, fibronectin, which represented an integral component of the ECM in the undifferentiated spheroids, was only expressed at low levels in differentiated spheroids. These data are consistent with studies using 3T3 preadipocytes that reported a degradation of fibronectin during adipogenesis in 2D cultures of these cells. 49
To prove the resemblance of the generated adipogenic 3D microtissues to human native fat, a comparison regarding the expression of the investigated ECM components was conducted using human abdominal fat tissue samples. The similarity in the ECM composition between the native tissue samples and spheroid-based microtissues was evident with collagen I, IV, V, VI, and laminin expression found at high levels in both the native tissue and in vitro generated microtissues. As in native human fat tissue, fibronectin was only expressed at low levels in the ECM of the differentiated spheroids. Overall, these findings emphasize the similarity of the native fat tissue and in vitro fabricated adipogenic microtissues with regard to the tissue-specific ECM composition.
In an attempt to delineate the time course of ECM remodeling during adipogenesis, laminin deposition was investigated in combination with staining for triglyceride accumulation. Laminin is an integral part of the basement membrane surrounding differentiated adipocytes. 50 In the spheroids, laminin deposition was evident during the course of adipogenesis, with increasing signal intensity over time. Interestingly, the onset of laminin production was observed at very early time points in the differentiation process (2 days after induction) and preceded the generation of lipid droplets. Laminin has been reported to promote adipogenic differentiation in 2D culture on laminin-coated substrata as shown with a MSC cell line (D1-Cells) and primary porcine preadipocytes.51,52 Furthermore, laminin alpha 4 null mice exhibited a decreased capacity for adipose tissue expansion and weight gain, underlining the important role of laminin in adipose tissue development and function. 53 Immunohistochemical studies in pig embryos demonstrated an association of laminin expression with the earliest phases of adipocyte development during embryogenesis, indicating a role of laminin in very early stages of the adipogenic differentiation process. 54 Thus, the observed deposition of laminin at the onset of the differentiation process in the ASC spheroids may trigger and sustain adipogenesis and contribute to the superior differentiation capability of the spheroids.
For the use of ASC spheroids in adipose tissue engineering applications, an efficient adipogenic induction leading to effective differentiation is desirable. Similar to other studies of adipogenic differentiation of ASC spheroids,18,33,55 we observed that differentiation was enhanced, when the ASCs were cultured as spheroids as compared with 2D monolayer cultures, with the cells in spheroids having significantly elevated triglyceride content and expression level of adipogenic marker genes. In addition to the enhanced triglyceride storage in the spheroids cultured under permanent induction, we demonstrated that applying a short-term induction protocol involving stimulation with the adipogenic hormonal cocktail for only 2 days led to effective differentiation in the spheroid cultures. In contrast, 2D cultured cells failed to differentiate under these conditions. Thus, in the 3D setting with cells embedded within their own matrix, a shorter stimulus was required to trigger and sustain the adipogenic differentiation cascade as compared with conventional 2D culture.
To further refine the time frame for efficient induction, spheroids were adipogenically induced during assembly (after 7 h) before reaching their final size or after full compaction (after 2 days). Interestingly, the differentiation of spheroids induced during assembly was improved relative to the spheroids induced after compaction. As the compaction process has not been finished after 7 h of spheroid assembly, the spheroids may be more permeable to the hormonal inducers. Using a low-molecular-weight fluorescent dye, Lin et al. demonstrated that the penetration of small molecules to the core of cell aggregates was only possible during early stages of assembly. 56 Overall, priming of cells toward the adipogenic lineage by adipogenic precultivation has been shown to be advantageous for fat formation in vivo.21,57 As ASC spheroids require only a short induction period early in culture (7 h after seeding), the cost-intensive in vitro precultivation time necessary to trigger adipogenic differentiation in vivo may be minimized.
In conclusion, the ECM of ASC spheroids was observed to be remodeled during adipogenic differentiation from a stromal to an adipose-specific pattern, with the ECM remodeling starting at very early time points and preceding the accumulation of lipid droplets. The observed resemblance of the ECM composition to native fat tissue may provide a valuable 3D model to further characterize the functional relationship between ECM composition and cellular behavior in a more in vivo-like context, which could also be beneficial in studies regarding obesity research. For adipose tissue engineering approaches, ASC spheroids appear to be promising prefabricated, material-free modules consisting of cells within their own tissue-specific matrix that could be applied as building blocks for generating larger tissues.
The superior differentiation capacity and the need for only a short induction stimulus to achieve sustained differentiation are attractive features of the ASC spheroids in terms of potential clinical application. Further research should include the combination of ASC spheroids with delivery vehicles such as hydrogels or specifically designed scaffolds. Recent biofabrication concepts, including the automated assembly of preformed microtissues within a 3D plotted polymer scaffold 58 or the seeding of thin-fiber melt electrowritten scaffolds with cellular spheroids generating sheet-like structures 32 may be applied to adipose tissue engineering.
Footnotes
Acknowledgments
The authors thank Sabine Müller-Morath and Julia Bachmann, University of Wuerzburg, Germany, for assistance with the qRT-PCR and image analysis, respectively. Furthermore, the authors thank Lauren Flynn, University of Western Ontario, Canada, for critically reading the manuscript.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the German Research Foundation (DFG), collaborative research center SFB TR225, project number 326998133 (sub-project C02). Furthermore, the authors wish to acknowledge the financial support by the Interdisciplinary Center for Clinical Research Wuerzburg, project number D-321, to K.R. and T.B.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.