
Abstract
In the present study, we present a comparative analysis among the outputs of porcine follicle isolation using either mechanical technique alone or in combination with enzymes, proposing an optimized protocol useful for all further applications related to follicle in vitro growth and reproductive tissue engineering. The porcine follicles were isolated using mechanical technique alone (hand blender and scalpels) or in combination with collagenase or Liberase Dispase High (DH) at different doses applying different protocols. Finally, the number, morphology, and stage of isolated follicles were compared between the protocols. Moreover, the follicle viability (live/dead assay) and morphology (rhodamine phalloidin and 4′,6-diamidino-2-phenylindole staining and scanning electron microscopy analysis) were evaluated after 10 days of culture. We found an optimum protocol for intact follicle isolation using the mechanical technique in combination with enzymes at a concentration of 0.5 mg/mL. However, the number of total isolated follicles and primordial follicles was significantly higher when collagenase was used compared to Liberase DH (p < 0.05), while Liberase DH could isolate a significantly higher percentage of preantral follicles. After 10 days of culture, the morphology and health status of follicles were statistically higher when Liberase DH was used in comparison with collagenase. Moreover, on the follicles extracted with Liberase DH, it was possible to observe theca cells covering part of the follicle surface. In conclusion, we demonstrated that the intact primary or secondary follicles could not be obtained using only mechanical methods, which led to the isolation of denuded oocytes and dramatically damaged follicles. We concluded that the collagenase-based follicle isolation could negatively affect the morphology and developmental potential of the follicles. Moreover, the incubation of ovarian cortex tissues with Liberase DH solution is an optimized protocol for porcine ovarian follicle isolation with developmental competence.
Impact statement
Isolation and in vitro maturation of follicles can pave the way for activities on reproductive tissue engineering (REPROTEN) and developing an artificial ovary. In this regard, the standardization and optimization of the extraction methods are pivotal for the design of experiment of follicle in vitro growth. In the present study, we provided a comparative analysis among the outputs of porcine follicle isolation using either mechanical technique alone or in combination with collagenase or Liberase DH, proposing an optimized protocol useful for all further applications related to follicles' in vitro growth and REPROTEN.
Introduction
Isolation and in vitro culture of ovarian follicles can be used as an excellent research tool for folliculogenesis and toxicological studies.1–5 Moreover, in the last decade, isolation and in vitro maturation of follicles have been considered as a potential alternative to restore fertility in a female who has abnormal oogenesis and in cancer patients who undergo gonadotoxic chemotherapy.6–9 Moreover, in the recent years the scientific community started to focus its research activities on reproductive tissue engineering (REPROTEN), as defined by Amorim. 10 In this framework, there is the need of the definition of standard optimized protocols for the follicle extraction and also a matrix to support the ovarian follicle growth in vitro.11,12
Most of the previous studies established the follicle isolation protocol for mice and rats,13–16 and less information is available regarding the isolation of follicles from large animals like pig. Furthermore, it has been reported that pig is a good comparative animal model to study ovarian development and follicles in large animals and women, due to similarities in folliculogenesis and mechanism for dominant follicle selection.17,18
Different mechanical and enzymatic methods, as well as their combination, have been used to isolate ovarian follicles, regardless of the species. For example, Wu et al. 19 isolated porcine preantral follicles by dissection of ovarian tissues into small pieces using forceps dissection. In the case of enzymatic isolation, different types of collagenase (Ia, II, IX, XI), which could isolate more follicles in comparison with mechanical methods, have been used in previous studies.20–22 In this regard, Choi et al. 23 demonstrated that the collagenase IV treatment with agitation could effectively isolate porcine preantral follicles; however, they did not evaluate the retrieved follicles' viability or development potential. Moreover, studies documented that collagenase could severely damage the follicles and so negatively affect culture and grafting outcomes. 24
Liberase, a purified mammalian tissue-free enzyme, has been recently introduced for the isolation of pancreatic islets. 25 In this regard, Nikolic and colleagues have indicated that this enzyme could isolate and preserve viable pancreatic islets better compared with the collagenase. Furthermore, Liberase DH has been used to isolate human follicles, and the results revealed that the follicles isolated with Liberase DH had high developmental competence. 26
Given the lack of a unified protocol to isolate a large number of porcine follicles with proper morphology, vitality, and development potential, in the present study, we present a comparative analysis among the outputs of porcine follicle isolation using either mechanical technique alone or in combination with collagenase or Liberase DH, proposing an optimized protocol useful for all further applications related to follicles' in vitro growth and REPROTEN.
Materials and Methods
The porcine ovaries were collected from the slaughterhouse (registration number DE 09 562 00 38 21; district veterinary office Erlangen-Höchstadt) and washed with cold Dulbecco's phosphate buffered saline (DPBS, D8661; Sigma-Aldrich).
Mechanical isolation
Using hand blender
As the first attempt to isolate the porcine follicles with a mechanical method, we applied a hand blender (Bosch, Germany) as previously described by Nuttinck et al. 27 and Leonel et al. 28 In this method we put at least three ovaries into the hand blender container containing 4° C DPBS buffer. The ovaries were mixed until obtaining a homogenate of ovarian tissues. Then the homogenate was evaluated under an inverted microscope at 100 × and 250 × magnifications. This method was repeated thrice with different ovaries.
Using scalpels
Based on the previous studies, dissecting the ovarian tissue by scalpel could be an easy method to obtain follicles. For this purpose, we isolated the cortex of ovaries in DPBS buffer. The ovaries were bisected into two equal halves through the hilum of ovary using scalpels. Each piece was further trimmed from the more permissive medulla site, leaving only the denser cortex tissue. Then the tissue cortex was divided into three different Petri dishes (labeled as a–c) containing 4°C DPBS.
Using scalpels, the cortex tissues were dissected into pieces with a size of <1 mm3 in dish (a), 1–2 mm3 in dish (b), and 3–5 mm3 in dish (c). Finally, after a mild shaking, each dish was evaluated under an inverted microscope at 100 × and 250 × magnifications. Moreover, in a similar experiment, the contents of dishes (a–c) were separately transferred into 50 mL falcon tubes and incubated for 30 or 60 min in a 37°C water bath in combination with shaking every 10 min for about 10 s. Then, the samples were evaluated under the microscope. All experiments were repeated thrice with different ovaries.
Enzymatic and mechanical isolation
Collagenase type I-A from Clostridium histolyticum (C2674; Sigma-Aldrich) and Liberase DH (05401054001; Sigma-Aldrich) were used to isolate porcine follicles. To find the best enzyme dose, incubation time, as well as the tissue size, we set up a series of pilot tests in which we varied cortex size (1–5 mm3), enzyme dose (0.1–1 mg/mL), and incubation time (30–75 min) with different shaking intervals (every 5–15 min).
After finding the best protocol, the cortex of one ovary was dissected in 4–5 mm3 pieces using scalpels. Then, 4 g of the pieces were transferred into two 50 mL falcon tubes containing 2.5 mL lysis buffer (2 g in each tube). The lysis buffer was prepared by adding either 1.25 mg collagenase (tube a) or Liberase DH (tube b) into 4° C DPBS (final concentration of 0.5 mg/mL). The tubes were incubated in a 37°C water bath for 70 min and were gently shaken for about 20 s every 10 min. After the incubation 10 mL of 4°C DPBS was added into the tubes to stop further enzymatic digestion.
The content of the tube was transferred into a petri dish, and the follicles were analyzed and collected under an inverted microscope with the magnification of 250 × . The follicles of each tube were evaluated based on the count, stage of follicular development, and morphology. The isolation process and follicle evaluation were repeated thrice for both enzymes. In each experiment, we used ovarian samples from the same animal for both enzymatic isolations.
Stage of follicles
The stage of follicles was evaluated based on the follicle diameters and layers of granulosa cells as described by Moniruzzaman et al. 29 and Telfer et al. 30 Based on the previously described properties, the follicles were categorized in four developmental stages, including primordial follicles, with a diameter of <50 μm and one layer of flattened granulosa cells; primary follicles, with a single layer of cuboidal granulosa cells and a diameter of 50–150 μm; preantral follicles, with 150–300 μm in diameter and two or more layers of granulosa; and early antral follicles with >300 μm in diameter.
Follicle culturing
About 100 nonruptured follicles with intact morphology from each isolation tube (collagenase or Liberase DH) were collected and transferred into a separate culture well after thrice washing with sterile DPBS. Follicles at different developmental stages (primordial to preantral stages) were equally cultured from both isolation tubes. The culturing protocol was described in our previous article. 12
Briefly, serum-free culture medium (McCoy's 5a medium; Life Technologies) containing 20 mM HEPES buffer (Gibco), 3 mM L-Glutamine (Gibco), 0.1% BSA (Bovine Serum Albumin, fraction V, Carl Roth), 0.1 mg/mL of each Penicillin and Streptomycin (Sigma-Aldrich), 2.5 μg/mL Amphotericin B (Fungizone; Gibco), ITS solution (Sigma-Aldrich), 50 μg/mL Ascorbic acid (Sigma-Aldrich), and 0.272 IE rFSH (Gonal-f; Merck) was used as the culture medium. The follicles were seeded in 24-well plates using CellCrown™24 inserts with a 0.4-micron PET membrane (Scaffdex, Finland). The follicles were cultured for 10 days at 38.5°C in humidified air with 5% CO2, and half the medium was removed and replaced every second day. The experiments were conducted thrice for each isolation method (collagenase or Liberase DH).
Live/dead assay
After 10 days of culturing, the viability of the follicles was assessed using LIVE/DEAD™ Viability/Cytotoxicity Kit (Thermo Fisher Scientific, Germany) based on the manufacturer's instructions. Briefly, the LIVE/DEAD working solution was prepared by adding 5 μL of the 2 mM EthD-1 solution and 2.5 μL of the 4 mM calcein-AM solution into 10 mL of sterile DPBS. After washing the samples thrice with DPBS, the working solution was added into the wells containing follicles and left to incubate for 30 min in a dark place at room temperature and then evaluated under a Zeiss fluorescence microscope (IM 35; Zeiss, Oberkochen, Germany). Evaluated follicles were classified in four categories corresponding to the proportion of dead granulosa cells, namely ≤1%, <5%, 5–10%, and >10% dead granulosa cells (Fig. 1). The experiments were conducted thrice for each isolation method (collagenase or Liberase DH).

Evaluation of follicle viability after 10 days of culture. Scale bars represent 100 μm. GCs, granulosa cells. Color images are available online.
Rhodamine phalloidin and 4′,6-diamidino-2-phenylindole staining
The morphology of follicles was evaluated after the 10 days culturing by staining them with rhodamine phalloidin (Thermo Fisher Scientific) and 4′,6-diamidino-2-phenylindole (DAPI; Thermo Fisher Scientific). For this purpose, the follicles were fixed using a fixation solution containing PIPES buffer, ethylene glycol tetraacetic acid, polyethylene glycol, paraformaldehyde, DPBS, and sodium hydroxide (all reagents purchased from Sigma Aldrich, Germany). After washing the samples with DPBS, they were immersed in a permeabilization buffer, constituted by Triton X-100, sucrose, and DPBS (all reagents purchased from Sigma Aldrich), and rhodamine-phalloidin was added to each well (8 μL/mL) and incubated at 37°C for 1 h.
After the removal of the dye, the samples were washed with DPBS, and DAPI (1 μL/mL) was added to each well. After removal of the dye, the samples were washed in DPBS and analyzed with a fluorescent microscope (Axio Scope A1; Zeiss). Follicle morphology was evaluated and categorized into four groups based on previously described method 31 ; M1, spherical form with complete granulosa cell layer; M2, irregular form with complete granulosa cell layer; M3, irregular form with <10% granulosa cell loss; and M4, totally atypical form with 10–50% granulosa cell loss (Fig. 2).
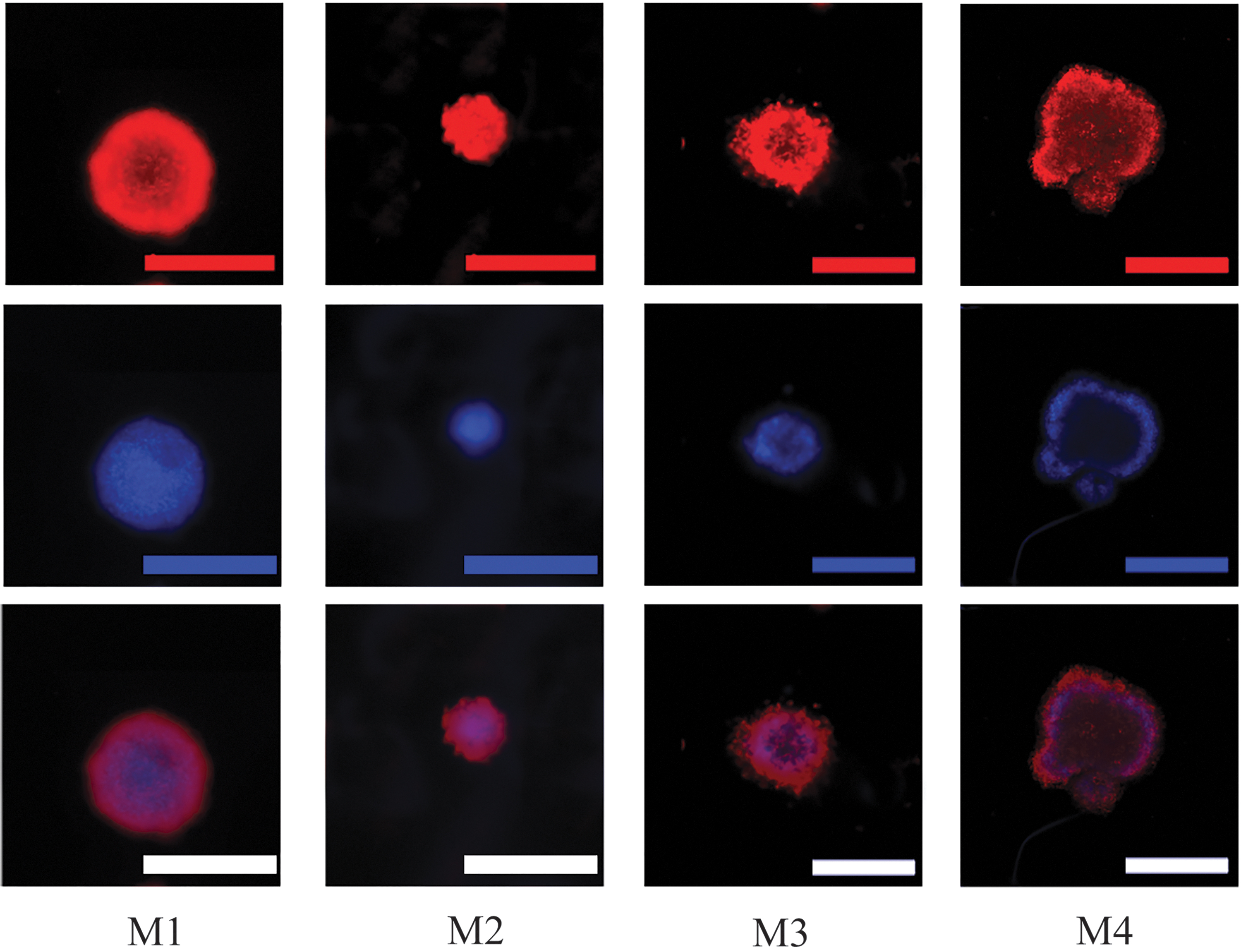
Ovarian follicle morphology categorization after 10 days of culture: rhodamine phalloidin (red) and 4′,6-diamidino-2-phenylindole (blue) staining. M1, spherical form with complete GC layer; M2, irregular form with complete GC layer; M3, irregular form with <10% GC loss; M4, totally atypical form with 10–50% GC loss; scale bars represent 200 μm. Color images are available online.
Scanning electron microscopy analysis
For sample fixation, the follicles in the inserts were immersed in a fixation solution containing glutaraldehyde, paraformaldehyde, sucrose, and sodium cacodylate trihydrate (all reagents from Sigma Aldrich). The samples were then dehydrated after immersion in a series of ethanol solutions and dried using a critical point drier (Leica EM CPD 300; Leica, Germany). Before scanning electron microscopy (SEM) analysis, they were gold sputtered (Sputter Coater Q150T; Quorum Technologies, Germany).
Statistical analysis
After performing the Kolmogorov–Smirnov test, the t-test was used for comparison of follicle factors between two isolation methods. p-Values <0.05 were considered significant. SPSS V.16 software was used for statistical analysis.
Results
Number of isolated follicles using mechanical methods
Our results demonstrated that with both methodologies (hand blender and scalpels, without incubation) it was not possible to isolate primary or secondary follicles. In fact, we found denuded oocytes, as well as dramatically damaged follicles with more than 50% granulosa cell loss (Fig. 3).
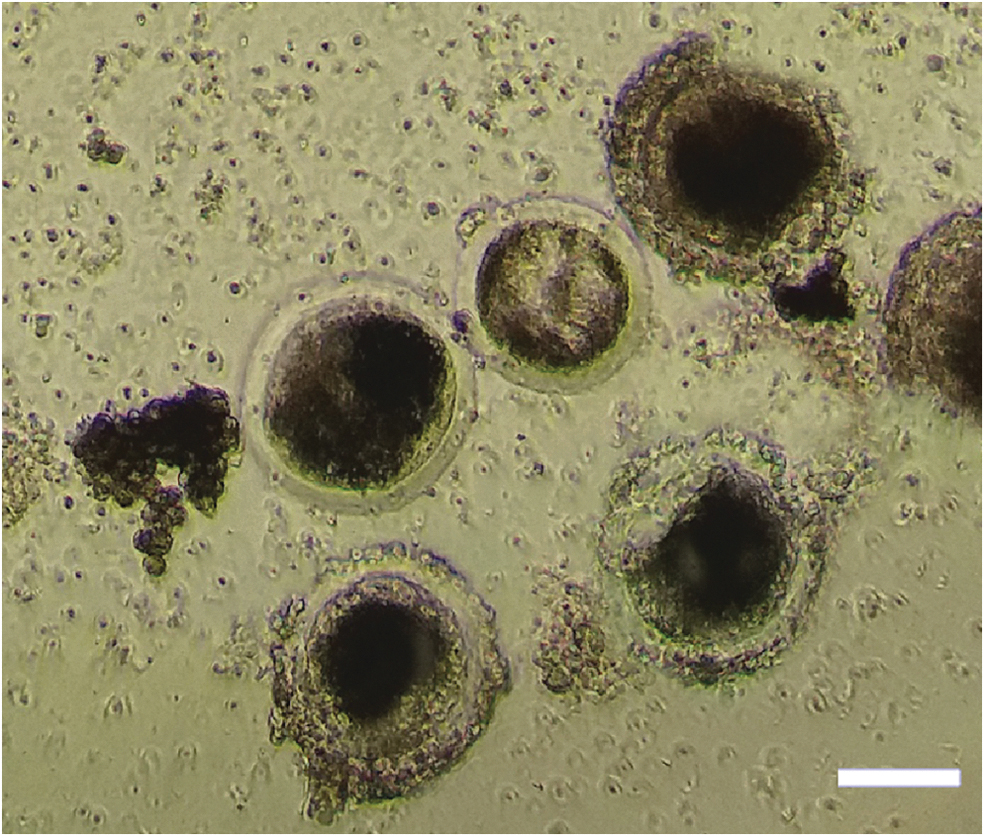
Isolated porcine follicles using mechanical methods. Denuded oocytes, as well as damaged follicles, with loss of GCs were obtained using the mechanical methods. Scale bar represents 50 μm. Color images are available online.
We found that dissection of the ovarian cortex into 4–5 mm3 pieces with subsequent incubation for 60 min in a 37°C water bath with shaking every 10 min resulted in the isolation of <20 intact follicles, as well as lots of damaged ones. However, incubation for a shorter time (30 min) or dissection into smaller pieces (<4 mm3) resulted in lack of intact follicle isolation. Moreover, the number of denuded oocytes increased when we used harsh method for isolation; therefore, the number of collected oocytes was higher in the mechanical method using blender than scalpels. Furthermore, we found a higher number of denuded oocytes when we dissected the tissue into <1 mm3 pieces with scalpels and incubated for 60 min in a 37°C water bath compared to the condition that we used for larger pieces (3–5 mm3) and 30 min incubation time (three oocytes vs. one oocyte in each 250 × high-power field (HPF) of microscope).
Number of isolated follicles using enzymatic and mechanical methods
Our pilot tests to find the best protocol for intact follicle isolation demonstrated that incubation of 2 g tissue fragments (4–5 mm3) in a lysis buffer containing 0.5 mg/mL of enzyme (collagenase or Liberase DH) for 70 min at 37° C could cause isolation of a higher number of intact follicles.
Comparison of isolated follicle numbers by collagenase type I-A and Liberase DH using the best protocol demonstrated that collagenase isolated a significantly higher number of follicles than Liberase DH (888 ± 132 vs. 570 ± 121 follicles per 2 g ovarian cortex, p = 0.037). We retrieved low numbers of follicles when the ovarian cortex was dissected into 1–2 mm3 pieces and incubated in a buffer with 0.5 mg/mL of enzyme for 70 min at 37°C (281 ± 107 for collagenase and 196 ± 72 for Liberase DH).
Stage of isolated follicles
Comparing the developmental stage of isolated follicles between both enzymatic methods showed that collagenase could isolate a significantly higher percentage of primordial follicles than Liberase DH (26.2% vs. 13.1%, p < 0.05, Table 1). However, Liberase DH could isolate significantly more preantral follicles compared to collagenase (25.5% vs. 9.9%, p < 0.05).
Stage of Isolated Intact Follicles Using Collagenase (421 Follicles) and Liberase Dispase High (393 Follicles)
df, degrees of freedom.
Health status of isolated follicles
We evaluated the viability of the follicles after 10 days culturing, and the results are shown in Table 2. Our results demonstrated that more than half of isolated follicles (58.49%) using Liberase DH were almost completely alive (≤1% dead granulosa cells) after 10 days culturing, while the percentage of completely healthy follicles was only 28.57% when collagenase was used. Considering that follicles with <5% dead granulosa cells (GCs) can be considered as alive follicles, we found that the percentage of alive follicles was significantly higher when Liberase DH was used compared to collagenase (82.07% vs. 59.82%, χ 2 = 13.005, p < 0.001).
Viability of Isolated Follicles Using Collagenase (127 Follicles) and Liberase Dispase High (138 Follicles) After 10 Days Culturing
GCs, granulosa cells.
Morphology of isolated follicles
Comparison of follicle morphology isolated by the enzymes revealed that the percentage of good morphology follicles (M1 and M2) after 10 days incubation was statistically higher in the method using Liberase DH compared to collagenase (72.38% vs. 44.44%, p < 0.05, Table 3).
Morphology of Isolated Follicles Using Collagenase (127 Follicles) and Liberase Dispase High (138 Follicles) After 10 Days Culturing
M1, spherical form with complete GC layer; M2, irregular form with complete GC layer; M3, irregular form with <10% GC loss; M4, totally atypical form with 10–50% GC loss.
SEM analysis performed on the porcine ovarian follicles after 10 days of culture on PET substrate extracted with the enzymatic method is reported in Figure 4. SEM analysis shows that with both enzymes a spherical shape of the follicles, with comparable size, is preserved. Moreover, on the follicles extracted with Liberase DH, it was possible to observe theca cells covering part of the follicle surface, as reported in the inset of Figure 4B.
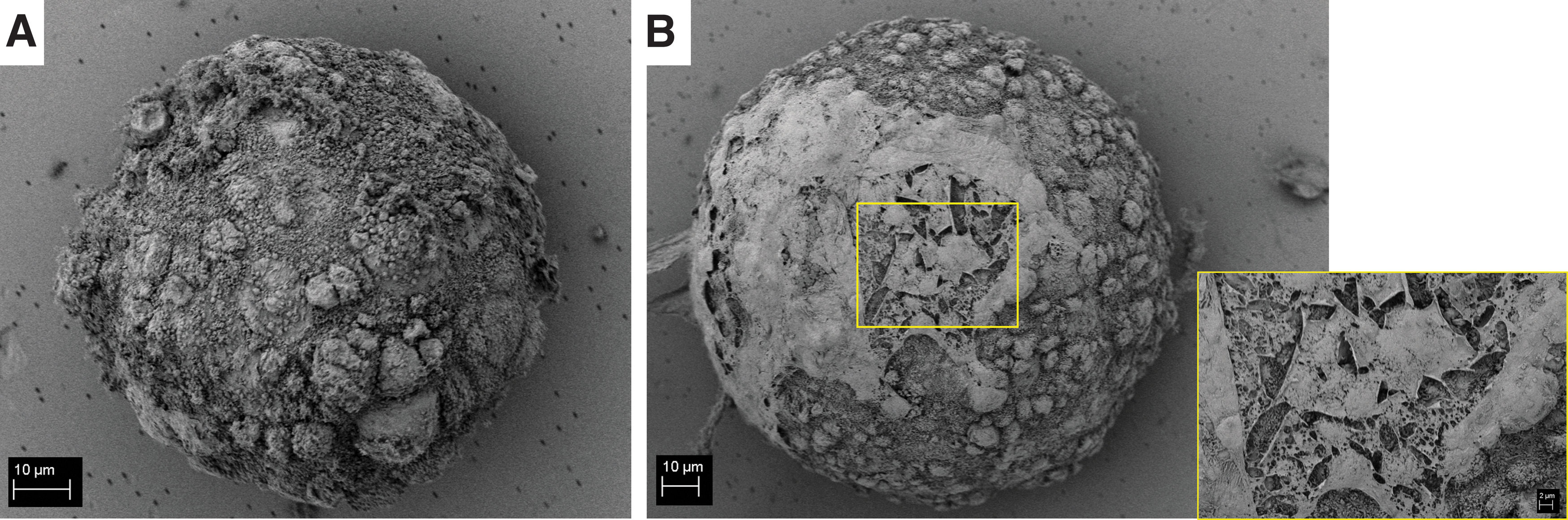
Scanning electron micrographs of representative porcine ovarian follicles extracted with collagenase
Discussion
The first step for in vitro oogenesis and also the development of an artificial ovary is isolating enough intact and viable follicles with the potential of growth. In this regard, big animal models, as porcine follicles, can be used as suitable reference than rodent model. To find an optimized method for porcine follicle isolation, in the present study we evaluated either mechanical technique alone or in combination with enzymatic method using collagenase or Liberase DH.
Since mechanical isolation of follicles without using enzyme can have less negative effect on the isolated follicles, we first tested different mechanical methods that had been introduced by previous studies. However, we found that the use of hand blender or scalpels could dramatically damage follicles; also changing incubation time or size of the tissue pieces could not result in intact follicle isolation. In contrast to our finding, it has been reported that mechanical isolation technique using a tissue chopper could isolate large numbers of morphologically normal preantral follicles of equine. 32
Moreover, previous studies demonstrated that there was no difference between number of isolated follicles between histological and mechanical isolation methods in goats and sheep.33,34 The underlying reason for failure of the mechanical methods in isolation of porcine ovarian follicles could be due to differences in the structure of porcine ovarian tissue with respect to other animal model. The porcine ovarian tissue is soft and it has been demonstrated that the porcine follicles cluster together in the cortex while in other animals such as bovine and ovine the follicles are unevenly scattered. 35 Moreover, the total number of follicles per cross-section of cortex is higher in porcine compared to other animals, increasing chance of damage by scalpels or blender. 35
The combination of mechanical and enzymatic methods resulted in isolation of a higher number of intact follicles especially when the ovarian cortex tissues with a size of 4–5 mm3 were incubated in a lysis buffer containing 0.5 mg/mL of enzyme for 70 min at 37°C and shaking every 10 min. In supporting our findings, Figueiredo et al. 36 also reported isolation of a higher number of bovine preantral follicles using combined mechanical and enzymatic methods with repeated pipetting. They showed that the combined method could isolate a significantly higher number of follicles compared with mechanical only method. However, Telfer and Watson 37 could just isolate a low number of equine preantral follicles (30–60) per whole ovary after 60–90 min of incubation in a solution containing 1, 3, or 5 mg/mL collagenase (type I-A).
Our results also indicated that collagenase could isolate a higher number of follicles than Liberase DH at a similar concentration. Such a high number of isolated follicles following collagenase treatment could be due to strong digestive activity of the enzyme. The collagenase used in the present study was type I-A which has a very strong activity, as it digests collagen from both ends. Moreover, collagenase type I-A has caseinase, clostripain, and tryptic enzymes that give additional digestive activities. 38
In addition to the rate of isolation, there was a significant difference in the developmental stage of isolated porcine follicles between the enzymes as collagenase caused more isolation of primordial follicles, while Liberase DH resulted in isolation of more preantral follicles. It is possible that collagenase damaged preantral follicles more than Liberase DH, and therefore, it increased the percentage of intact primordial follicles, which are small in size, have no cortical granule and no zona pellucida, 39 and are therefore maybe less sensitive. In support of this hypothesis, Lierman et al. 31 reported that different enzymatic isolation methods can differently affect the health of follicles at different maturation stages. However, they found that the enzyme protocol with Liberase DH isolated a higher percentage of normal shaped primordial follicles in comparison with the protocol based on collagenase IV.
Moreover, preantral follicles have theca cells, as well as multiple layers of GCs, 40 and collagenase type I-A with a very strong activity can more effectively dissociate the cell–cell connection, destroying the follicle integrity, than Liberase DH with mild activity. In contrast, the primordial follicles are surrounded by a single layer of flattened GCs, 40 and therefore, their cells are less exposed to the digestive enzyme.
Ten-day culture of isolated follicles revealed that follicles isolated by Liberase DH had higher developmental potential as more than half of the follicles were completely alive after 10 days, while around 75% of follicles isolated by collagenase could not completely survive after the culturing. Moreover, the morphology of follicles was better in the group isolated by Liberase than in the group isolated by collagenase. In accordance with our findings, Dolmans et al. 41 indicated that Liberase DH could isolate more vital follicles with good morphology from human ovary in comparison with collagenase I-A.
It seems that high endotoxin levels of collagenase, due to a crude preparation of C. histolyticum, could negatively influence follicular cells. 24 In this regard, a higher DNA fragmentation level has been observed in human granulosa cells isolated by collagenase IV compared to Liberase DH. 31 In contrast, high DNA fragmentation in GCs could induce apoptosis in these cells, and studies demonstrated that apoptosis of GCs could induce atresia in porcine follicles. 42 Since early apoptosis of follicular cells can negatively affect the developmental competence of the whole follicle, an isolation protocol which can isolate healthy follicles would be necessary for in vitro folliculogenesis, as well as artificial ovary development.
Further studies are required to investigate the association between DNA fragmentation of GCs and developmental competence of follicles. Moreover, the mechanism of collagenase in negatively affecting DNA of GCs should be determined to pave the way for developing modified follicle isolation methods without the destructive effect; for example, using antioxidants.
SEM analysis is a characterization technique, usually used in material science and tissue engineering applications, to investigate the microstructure of the samples. In fact, in particular for tissue engineering applications, the morphology of the scaffolds must be characterized before putting them in contact with any cells, to investigate cell–biomaterial interactions. Those kinds of interactions are based on chemical, physical, mechanical, and morphological signals provided by the biomaterials, which can affect the biological response.43–45
In this framework of interdisciplinary fields, as REPROTEN, it is fundamental to expand the characterization techniques, applying them also in fields which were previously unexplored. In this case, we used SEM analysis to investigate the porcine ovarian follicle morphology, confirming the spherical three dimensional shape, and we could also acquire some information about the presence of some theca cells on the follicle surface, which were not visible with the other analysis performed to investigate their morphology and viability. These results confirmed the relevance of a multidisciplinary approach in which novel characterization techniques could be introduced and standardized for further studies.
In conclusion, the mechanical method alone could isolate just a few numbers of porcine follicles with a high rate of granulosa cell loss, and a combination of enzymatic and mechanical methods is required to isolate intact follicles. Furthermore, although using collagenase could isolate more follicles than Liberase DH, collagenase-based follicle isolation could negatively affect morphology and developmental potential of the follicles. Therefore, it can be suggested that incubation of ovarian cortex tissues with size of 4–5 mm3 in a 0.5 mg/mL Liberase DH solution is an optimized protocol for porcine ovarian follicle isolation, and the protocol can isolate follicles with developmental competence.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
No specific funding was received for the study. The research was supported financially by institutional funding from Erlangen University Hospital.