
Abstract
A key challenge in the treatment of large bone defects is the need to provide an adequate and stable vascular supply as new tissue develops. Bone tissue engineering applies selected biomaterials and cell types to create an environment that promotes tissue formation, maturation, and remodeling. Mesenchymal stromal cells (MSCs) have been widely used in these strategies because of their established effects on bone formation, and their ability to act as stabilizing pericytes that support vascular regeneration by endothelial cells (ECs). However, the creation of vascularized bone tissue in vitro requires coupling of osteogenesis and vasculogenesis in a three-dimensional (3D) biomaterial environment. In the present study, 3D fibrin hydrogels containing MSCs and ECs were prevascularized in vitro for 7 days to create an endothelial network in the matrix, and were subsequently cultured for a further 14 days under either continued vasculogenic stimulus, a combination of vasculogenic and osteogenic (hybrid) stimulus, or only osteogenic stimulus. It was found that ECs produced robust vessel networks in 3D fibrin matrices over 7 days of culture, and these networks continued to expand over the 14-day treatment period under vasculogenic conditions. Culture in hybrid medium resulted in maintenance of vessel networks for 14 days, while osteogenic culture abrogated vessel formation. These trends were mirrored in data representing overall cell viability and cell number in the 3D fibrin constructs. MSCs were found to colocalize with EC networks under vasculogenic and hybrid conditions, suggesting pericyte-like function. The bone marker alkaline phosphatase increased over time in hybrid and osteogenic media, but mineral deposition was evident only under purely osteogenic conditions. These results suggest that hybrid media compositions can support some aspects of multiphase tissue formation, but that alternative strategies are needed to obtain robust, concomitant vascularization, and osteogenesis in engineered tissues in vitro.
Impact statement
The combined use of mesenchymal stromal cells (MSCs) and endothelial cells to concomitantly produce mature bone and a nourishing vasculature is a promising tissue engineering approach to treating large bone defects. However, it is challenging to create and maintain vascular networks in the presence of osteogenic cues. This study used a 3D fibrin matrix to demonstrate that prevascularization of the construct can lead to maintenance of vessel structures over time, but that osteogenesis is compromised under these conditions. This work illuminates the capacity of MSCs to serve as both supportive pericytes and as osteoprogenitor cells, and motivates new strategies for coupling osteogenesis and vasculogenesis in engineered bone tissues.
Introduction
A
In the field of tissue engineering, prevascularization of orthopedic tissue constructs can be achieved using either in vivo or in vitro approaches. In vivo strategies typically involve initial implantation of engineered tissue grafts within a well-vascularized site, for example, a muscle pouch, for a period of time to allow ingrowth of a rudimentary vascular pedicle. A second surgery then removes the vascularized construct for implantation into the defect site. This approach allows instantaneous reperfusion by surgical anastomosis to the vascular pedicle; however, it requires multiple surgeries. In vitro approaches harness the vasculogenic capacity of appropriate cell types embedded in the engineered construct. Culture conditions must be optimized to allow tissue maintenance along with the formation of stable vasculature within the engineered tissue over time. This method allows the creation of vascular networks in vitro, which can then inosculate with the host upon implantation. This approach avoids the need for multiple surgeries and has been shown to effectively promote vascularization in vivo.6,7
The choice of cell type(s) is crucial for vascularization strategies in bone tissue engineering. Mesenchymal stromal cells (MSCs) are well established and widely studied as osteoprogenitor cells, and can be readily harvested and expanded from biopsies. MSCs have demonstrated promising results for bone regeneration in various preclinical animal models,8–10 as well as initial clinical efficacy for the treatment of selected orthopedic conditions. 11 Osteogenic differentiation of MSCs can be promoted through in vitro culture with specific inductive factors, 12 which consistently produce expression of osteogenic genes and concomitant tissue mineralization.13,14 In addition, MSCs are known to secrete paracrine factors that promote vascularization and have been used widely as support cells for induction of vascular networks.15,16 Furthermore, MSCs can take on a pericyte-like function upon direct contact with endothelial cells (ECs),17,18 thereby stabilizing and allowing the maturation of blood vessels. 19 Prevascularization strategies that promote the formation of mature, stable vasculature have been shown to promote vessel retention and increase overall vascularization in critical-sized bone defects.6,7
Effective tissue engineering approaches to healing large bone defects require the combination of osteogenesis and vasculogenesis within a single, multiphase construct.20,21 Interestingly, despite the abundance of evidence linking vascular development and osteogenesis in vivo,22,23 it has been decidedly difficult to achieve stable, mature vasculature and appropriate osteogenic differentiation within a unified environment in vitro. 24 The distinct set of biochemical cues necessary for the induction of either vasculogenic or osteogenic function have motivated a variety of approaches using “hybrid” media that promote both functions. However, these methods typically result in only rudimentary vessels,25,26 an effect that is likely attributable to the deleterious effects that osteoinductive factors have shown on EC function and vasculogenic capacity.27–29 These outcomes have motivated biologically inspired strategies that induce vascular development before osteogenesis, in an effort to promote the formation of stable vasculature while mitigating harmful effects associated with osteoinductive factors. This sequential approach has produced promising outcomes, although both vessel formation and subsequent bone development are limited.30,31 The role of supporting pericyte-like cells in enabling vascularization and subsequent osteogenesis is still unclear. Therefore, a key to achieving the full regenerative potential of such coculture systems is to harness both the vasculogenic and osteogenic functions of the supporting stromal cells.
The present study investigates how osteogenesis and vasculogenesis are coupled and can be maintained in a three-dimensional (3D), in vitro EC+MSC coculture system. Engineered constructs were exposed to defined vasculogenic and osteogenic media formulations, and the effects on tissue development and maintenance were examined. In particular, ECs were cocultured with MSCs in 3D fibrin hydrogel constructs under vasculogenic conditions to induce EC network formation, and were then stimulated with osteoinductive factors either alone or in combination with vasculogenic medium. The effects of media composition and culture protocol on EC network formation were measured, and MSC phenotype was assessed by pericyte marker expression, bone cell markers, and tissue mineralization. This study reveals the effects of different culture media conditions on vascularization and osteogenesis, while also examining the corresponding phenotype of MSCs. The findings have utility to tissue engineers working in the creation of vascularized tissues, and in particular vascularized bone.
Methods
Cell culture
Human bone marrow-derived mesenchymal stromal cells (MSC; RoosterBio, Inc., Frederick, MD) were expanded following the manufacturer's protocol using a high-performance hMSC basal medium (SU005) and booster supplements (GTF-SE003). Primary human umbilical vein endothelial cells (EC; Lonza, Inc., Walkersville, MD) were expanded in fully supplemented VascuLife endothelial cell culture medium (Lifeline Cell Technology, Frederick, MD). Both cell types were cultured in T-175 flasks to 80% confluency and used in experiments between passages 4 and 6. Cells were maintained at 37°C in standard cell culture incubators with media changes every 2 days.
Cell encapsulation in 3D fibrin hydrogels
Cells were encapsulated within bulk fibrin hydrogels (2.5 mg/mL) at 0.25 mL/construct. All constructs contained 500 K total cells/mL fibrin, and those containing cocultures of MSCs and ECs were cast at a 1:1 ratio. For hydrogel casting, MSCs and ECs were detached using TripLE Express Enzyme (Thermo Fisher), resuspended in VascuLife growth medium, and counted using an automated cell counter (Multisizer 3; Beckman Coulter, Brea, Ca). Fibrinogen (Sigma-Aldrich, St. Louis, MO) was dissolved in phosphate-buffered saline (PBS) at 37°C (4.0 mg/mL active clottable protein), sterile filtered through a 0.22-μm syringe filter (Millipore Sigma) and kept on ice until ready for use. Around 1.0 mL of hydrogel was made by mixing 625 μL of fibrinogen, 100 μL fetal bovine serum (FBS; Gibco), 20 μL thrombin (Sigma), and 255 μL of cell suspension. The well-mixed cell/matrix suspension was quickly dispensed into individual wells of a 48-well plate (0.25 mL/gel). After dispensing the hydrogel solution, the 48-well plates were incubated at 37°C for 30 min to facilitate gelation. Complete media were added to each construct after gelation (0.5 mL/gel).
In vitro cultivation
The timeline and protocol for examining vascularization and osteogenesis in 3D fibrin hydrogels are shown schematically in Figure 1. Formation of vessel networks was first induced through 7 days of culture in vasculogenic growth medium (VascuLife). At this point, one of three experimental treatment conditions was applied to 3D constructs for a further 14-day culture period: (1) continued culture in pure vasculogenic medium (VAS) consisting of VascuLife base medium and defined vasculogenic supplements, (2) transfer to a hybrid medium (VAS+sOST) consisting of VascuLife base medium and both vasculogenic and osteogenic supplements, or (3) transfer to pure osteogenic medium (OST) consisting of DMEM base and defined osteogenic supplements. A list of all supplements and concentrations for each media formulation is found in Table 1. Constructs were cultured in treatment media for 14 days with media changes every 2 days. Samples were taken for full analysis at day 0, 7, and 14.
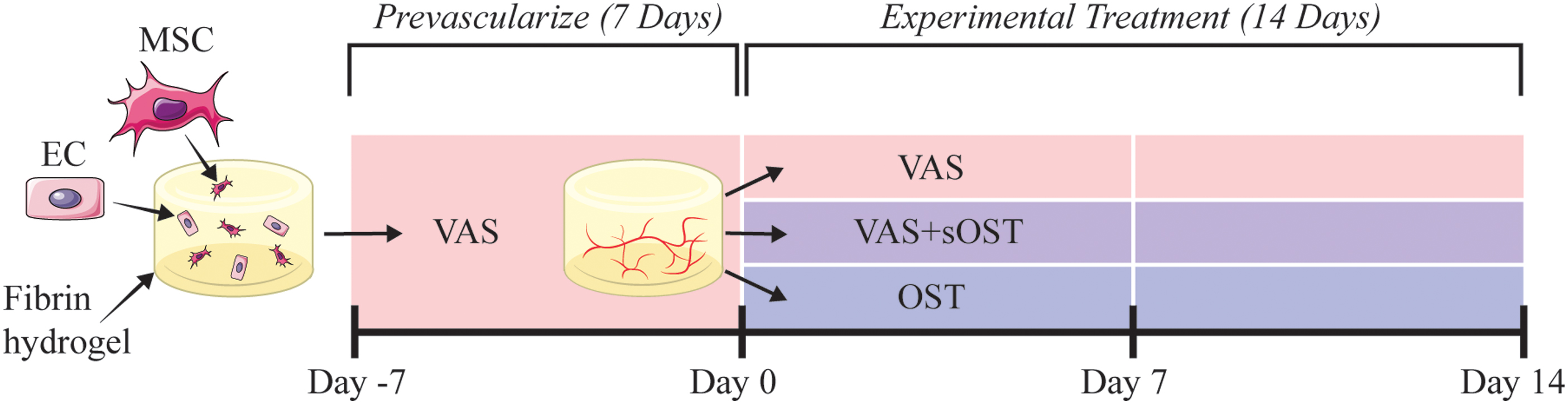
Schematic of Experimental Approach. Fibrin constructs containing ECs and MSCs were prevascularized in VAS medium for 7 days, and were then treated in either VAS+sOST, or OST for an additional 14 days. Vessel formation and osteogenic marker expression were quantified at days 0, 7, and 14. EC, endothelial cell; MSC, mesenchymal stromal cell. Color images are available online.
Media Types and Components
Cell viability and proliferation
Cell viability and proliferation were evaluated using the Live/Dead™ Cell Imaging Kit (Thermo Fisher) and DNA assay kit (Quant-iT™ PicoGreen dsDNA Kit; Invitrogen), respectively. Live/Dead staining was performed according to the manufacturer's protocol. Live cells (indicated by calcein AM staining) and dead cells (indicated by ethidium homodimer-1 staining) were observed and imaged using a fluorescent microscope. DNA was harvested from constructs by adding 0.5 mL Triton X-100 solution (0.5% in PBS) to each construct. Freeze/thaw cycles were conducted three times with brief vortexing between cycles to facilitate cell membrane disruption. The DNA assay was conducted using the manufacturer's protocol. In brief, standard curves were constructed using serial dilutions of λDNA standard (Invitrogen, 25250) in a 10 mM Tris-HCl solution (pH 7.5). One hundred microliters of 10 mM Tris-HCl was added to each well of a black 96-well plate. Samples/standards were added in triplicate at 10 μL per well. One hundred and ten microliters PicoGreen buffer was added to each well, covered in foil and incubated for 30 min at 37°C. Buffer was made by 200-fold dilution of Quant-iT PicoGreen dsDNA Reagent (Thermo Fisher; P11495) in 10 mM Tris-HCl. Following incubation, the plate was read at an excitation of 498 nm and emission of 518 nm.
Immunofluorescent staining
At each week throughout culture, constructs were imaged using confocal microscopy. Constructs were rinsed 3 × with PBS for 5 min at room temperature and then fixed using 500 μL Z-fix per gel for 20 min at room temp. For assessment of vessel network formation, 250 μL of stain solution comprising PBS, Ulex Europaeus Lectin 1 (UEA, Vector Laboratories, RL-1062, 1:100), and 4′, 6-diamidino-2-phenylindol (DAPI; Sigma-Aldrich, 1:1000) was added to each construct. The entire culture plate was wrapped in foil and stored for overnight incubation at 4°C. The following day gels were rinsed 3 × with PBS for 5 min at room temp. For assessment of pericyte marker expression, gels were first permeabilized with 500 μL of 0.5% Triton X-100 in TBS for 30 min at 4°C. Following a rinse 3 × for 5 min at room temp with 0.1% Triton X-100 in TBS (TBS-T), samples were blocked overnight at 4°C with 2% bovine serum albumin (BSA; Sigma). Two hundred and fifty microliters of primary antibody stain solution consisting of TBS-T containing 2% BSA and anti-smooth muscle actin primary antibody (Abcam; ab7817; 1:200) was added to each construct and stored for overnight incubation at 4°C. The following day gels were rinsed 3 × for 5 min with TBS-T. Two hundred and fifty microliters of secondary antibody stain solution comprising TBS-T containing 2% BSA, Alexa Fluor 488 Goat anti-mouse IgG secondary antibody (LifeTech; A11001; 1:400), UEA (1:100), and DAPI (1:1000) was added to each construct. Entire culture plate was wrapped in foil and stored for overnight incubation at 4°C. Following a 3 × rinse for 5 min at room temperature with TBS-T, gels were incubated with TBS-T overnight at 4°C.
Fluorescent imaging and vessel quantification
Images were acquired using an Olympus IX81 equipped with a Disc Spinning Unit and a 100 W high-pressure mercury burner (Olympus America, Center Valley, PA), a Hamamatsu Orca II CCD camera (Hamamatsu Photonics, K.K., Hamamatsu City, Japan), and Metamorph Premier software (Molecular Devices, Sunnyvale, CA). For vessel network formation, at least four images per construct were taken at random over a 200 μm depth (10 slices at 20 μm thick) at 4 × magnification. Slices were superimposed using maximum intensity z projection for qualitative analysis and network density quantification. Total network density was quantified using the Angiogenesis Tube Formation module in Metamorph Premier (Molecular Devices). For pericyte marker expression, at least six images were taken throughout the volume of the construct to generate a representative image per condition. Images were captured over a 50 μm depth (six slices at 10 μm thick) at 10 × magnification. Slices were superimposed for qualitative analysis.
Intracellular alkaline phosphatase quantification
Intracellular alkaline phosphatase (ALP) activity was quantified using p-nitrophenol phosphate (pNPP; Sigma-Aldrich) assay. Five hundred microliters of 0.5% Triton X-100 solution was added to each sample to permeabilize cell membrane. Three freeze/thaw cycles were performed with brief vortexing in between to facilitate intracellular ALP harvest. Standard curves were constructed using serial dilutions of p-Nitrophenol standard solution (Sigma; N7660) in a 2.0 N NaOH solution. Two hundred microliters of standard solutions were added to a 96-well plate. Ten microliters of cell lysate was added to each well in triplicate along with 100 μL of working solution (composed of ALP substrate solution (Sigma; N2765) and Alkaline Buffer Solution (Sigma). Samples were incubated for 30 min at 37°C, after which reactions were stopped using 90 μL of 2.0 N NaOH and the plate was read at 405 nm.
Mineral deposition
Calcium deposition was quantified using the orthocresolphthalein complex one (OCPC) method as previously described. 32 Briefly, 500 μ of 1.0 N acetic acid was added to each sample and incubated overnight at 37°C. To facilitate digestion, constructs were briefly sonicated for 5 s at 10% magnitude. Standard curves were constructed using serial dilutions of CaCl2 (Sigma) dissolved in 1.0 N acetic acid. Twenty microliters of samples/standards were added to respective wells in triplicate to a clear 96-well plate. Two hundred and fifty microliters of working solution, consisting of 0.05 mg/mL of OCPC in ethanolamine-boric acid-8-hydroxyquinoline buffer, was added to each well. After 10 min of incubation at room temp, absorbance was measured at 575 nm.
Statistical analyses
Data are presented as mean ± SD (n = 3). Statistical significance was determined using 2-way analysis of variance (ANOVA) with a Tukey's multiple comparison post hoc test, and all statistical analyses were performed using Prism 8.3 software (GraphPad, San Diego, CA). A p < 0.05 was considered statistically significant.
Results
Formation of vascular structures in 3D fibrin hydrogels
The ability of embedded cells to form EC networks in 3D fibrin constructs was assessed using EC alone and in combination with MSCs as support cells. Cell-seeded hydrogels were cultured in either vasculogenic medium (VAS) or control medium for 7 days. Formation of vessel-like structures was assessed through fluorescent staining and confocal microscopy analysis, and the results are shown in Figure 2. Over the 7-day culture period, both cell content and media formulation were found to affect the extent of vessel formation. Constructs with ECs alone and cultured in control medium displayed individual, rounded cell foci and an absence of multicellular structures (Fig. 2A
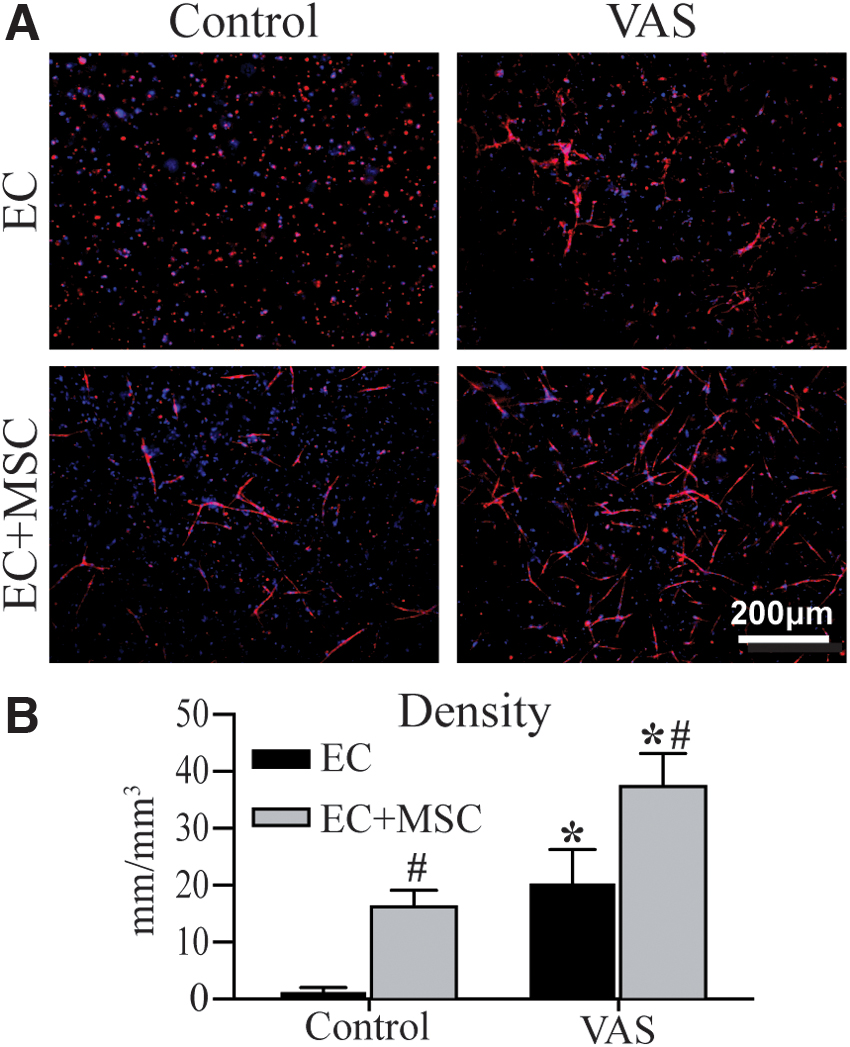
Vessel-like network formation in constructs cultured for 7 days in control or VAS media.
Osteogenic differentiation of MSCs in 3D fibrin hydrogels
The capacity for MSCs to differentiate toward an osteogenic lineage and to mineralize a matrix was investigated by creating 3D fibrin constructs containing either MSCs alone or in combination with ECs. Constructs were cultured in standard osteogenic medium containing β-glycerol phosphate, ascorbic acid, and dexamethasone for 14 days. The phenotype of embedded MSCs was assessed by measuring ALP activity and calcium deposition as early and late-stage markers of osteogenic differentiation, respectively, as shown in Figure 3. At day 14, ALP activity was significantly higher in constructs containing MSCs and cultured in OST, compared with those cultured in control medium (Fig. 3A). This effect of OST was not observed in EC+MSC constructs, which contained only half the number of MSCs. No calcium deposition was measured in control constructs (Fig. 3B); however, OST culture was shown to elicit robust and similar degrees of calcium deposition in both MSCs and EC+MSC constructs, despite the fact that the coculture samples contained fewer MSCs.
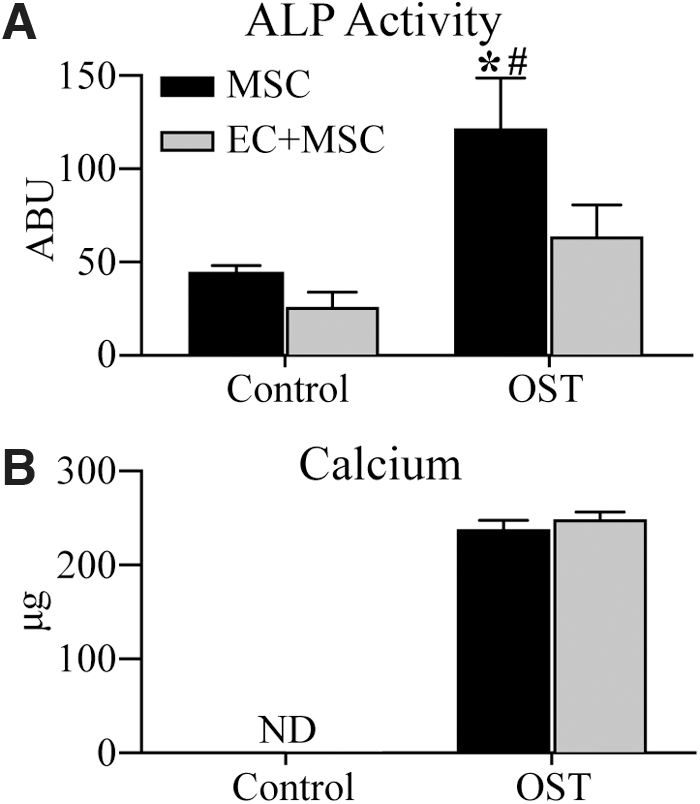
Osteogenic marker expression in constructs after 14 days of culture in either control or OST media. Quantification of
Prevascularized constructs—cell viability and proliferation
The experimental approach shown in Figure 1 was applied to examine the effects of prevascularization of EC+MSC constructs on subsequent osteogenic and vasculogenic potential. Constructs were cultured in VAS for 7 days, followed by 14 days of culture in either VAS, OST, or hybrid VAS+sOST. In this protocol, “Day 0” refers to the point after the initial 7-day prevascularization period. Figure 4 shows the effects of these culture protocols on overall cell viability and proliferation in 3D fibrin constructs. Vital staining revealed high viability and spread cell morphology at the end of the initial prevascularization period in VAS (Day 0, Fig. 4A). Subsequent continued culture in VAS led to higher viability over the additional 14-day treatment period (Fig. 4B). Constructs exposed to either VAS+sOST or OST media showed no statistically significant decrease in viability over time, although viability in the OST group was lower than in VAS at both day 7 and day 14. However, viability was generally high (>75%) and stable in all constructs over time. Quantification of DNA content revealed significant cell proliferation over the prevascularization stage of culture in VAS (Fig. 4C). At day 14 after prevascularization, cell numbers in the VAS and VAS+sOST conditions remained statistically the same as at day 0, whereas those in OST conditions had decreased significantly.
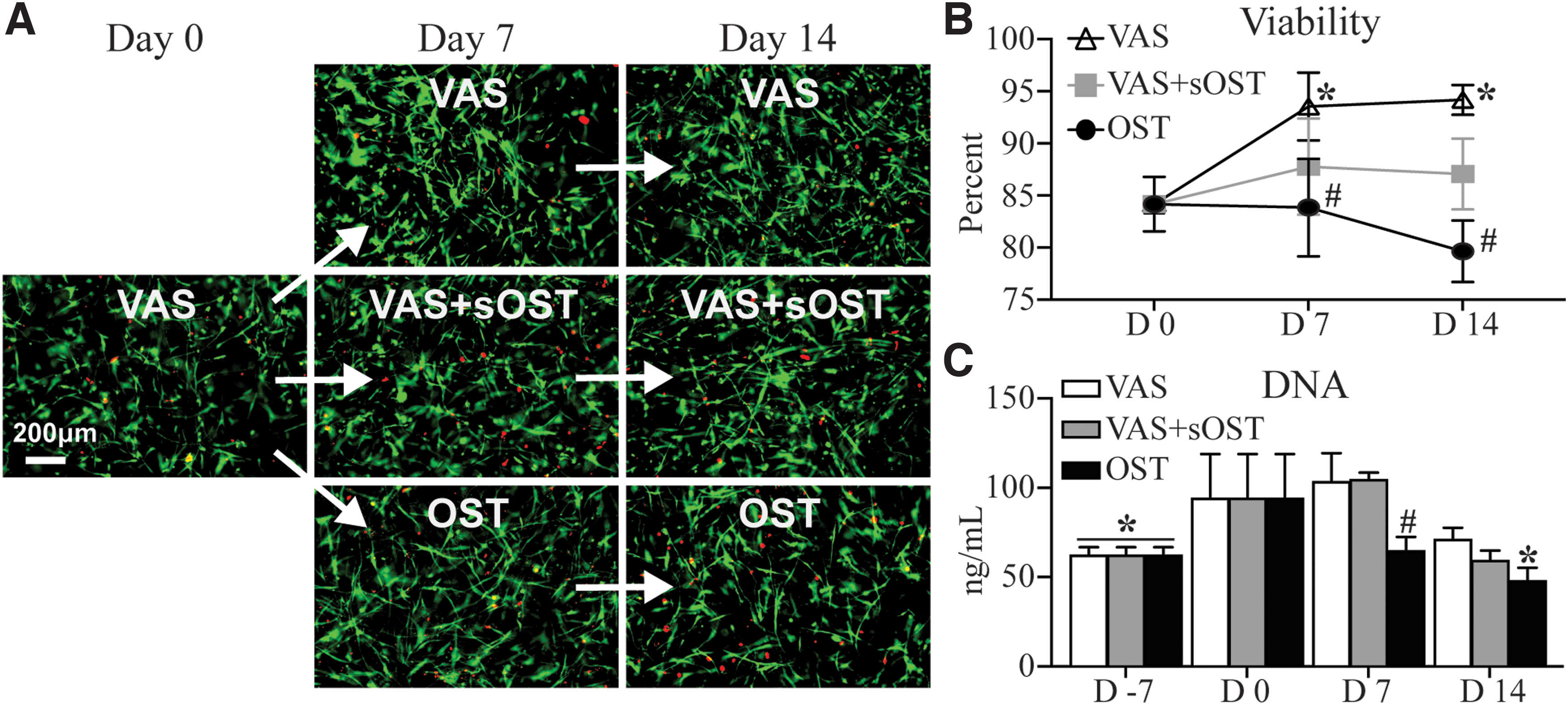
Cell viability and proliferation through the prevascularization and treatment periods.
Prevascularized constructs—vessel network density
The development and maintenance of EC networks in prevascularized 3D hydrogels was monitored over time after these constructs were exposed to different culture conditions, as shown in Figure 5. Staining of EC revealed that the formation of multicellular networks was achieved over the 7-day prevascularization period (Fig. 5A). Constructs subsequently cultured in either VAS or VAS+sOST retained EC networks, whereas those in OST resulted in regression of networks over time, such that by day 14 no multicellular structures were identifiable. Quantification of EC networks revealed that network density in the OST condition had decreased from day 0 throughout the remainder of the culture period (Fig. 5B). Although this regressive behavior was not observed in constructs of the VAS+sOST condition, at day 14 the network density was statistically lower compared with the VAS group.
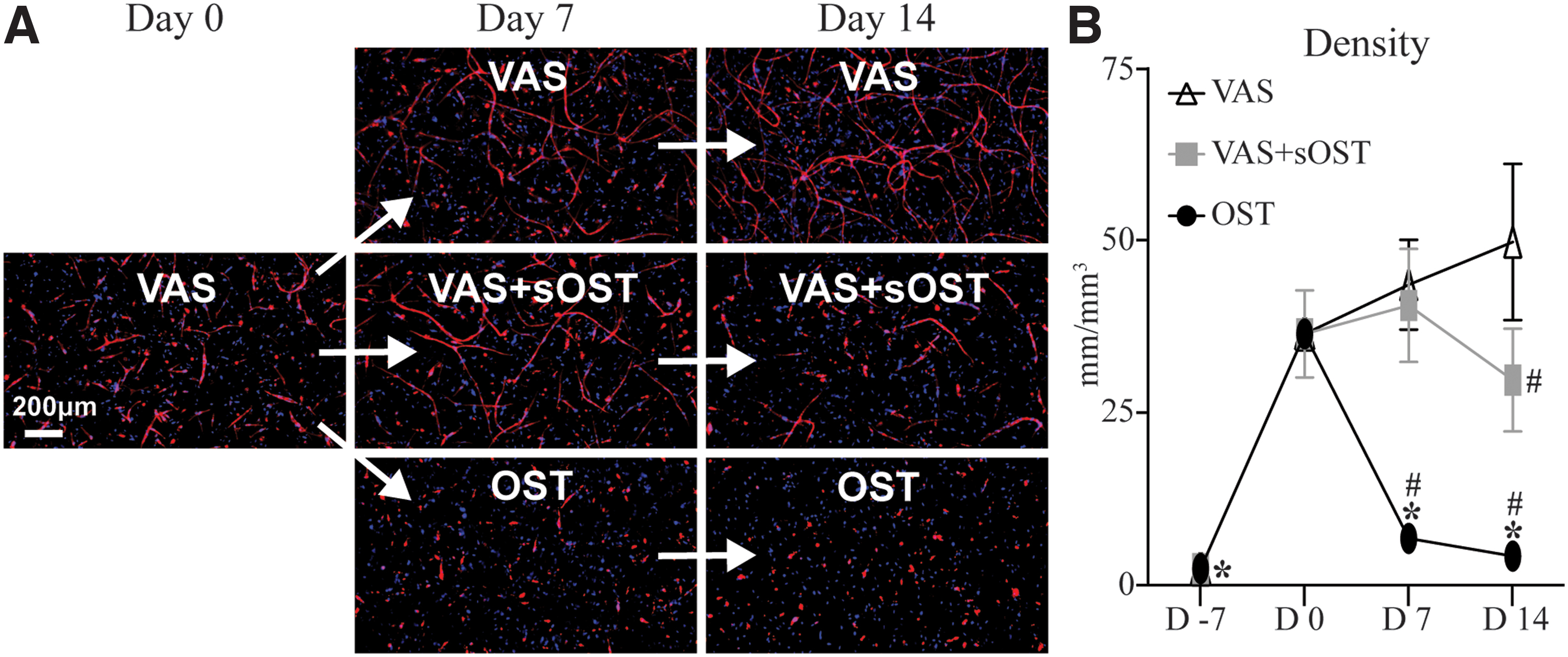
Vessel network formation through the prevascularization and treatment periods.
Prevascularized constructs—pericyte marker expression
The presence and localization of alpha-smooth muscle actin (αSMA), a marker associated with the pericyte phenotype, was examined, as shown in Figure 6. This cytoskeletal protein is increasingly expressed as MSCs take on pericyte-like functions and act to stabilize nascent vascular networks.16,33 Expression of αSMA was apparent at the end of the prevascularization period, indicating that 7 days of culture in VAS was sufficient for inducing a pericyte-like phenotype (data not shown). After a further 14 days of culture past the prevascularization period, constructs continued in VAS medium showed strong αSMA expression, with abundant positively stained cells throughout the matrix volume. In VAS medium, stromal cell morphology was spindle shaped and the αSMA-positive cells tended to be colocalized with EC networks. Constructs cultured in hybrid or OST conditions also exhibited distinct αSMA expression in stromal cells, although the cells in the OST condition were not colocalized with vessel-like structures. Furthermore, αSMA-expressing cells in the OST condition did not exhibit the same spindle-shaped morphology as observed in the VAS or VAS+sOST conditions.
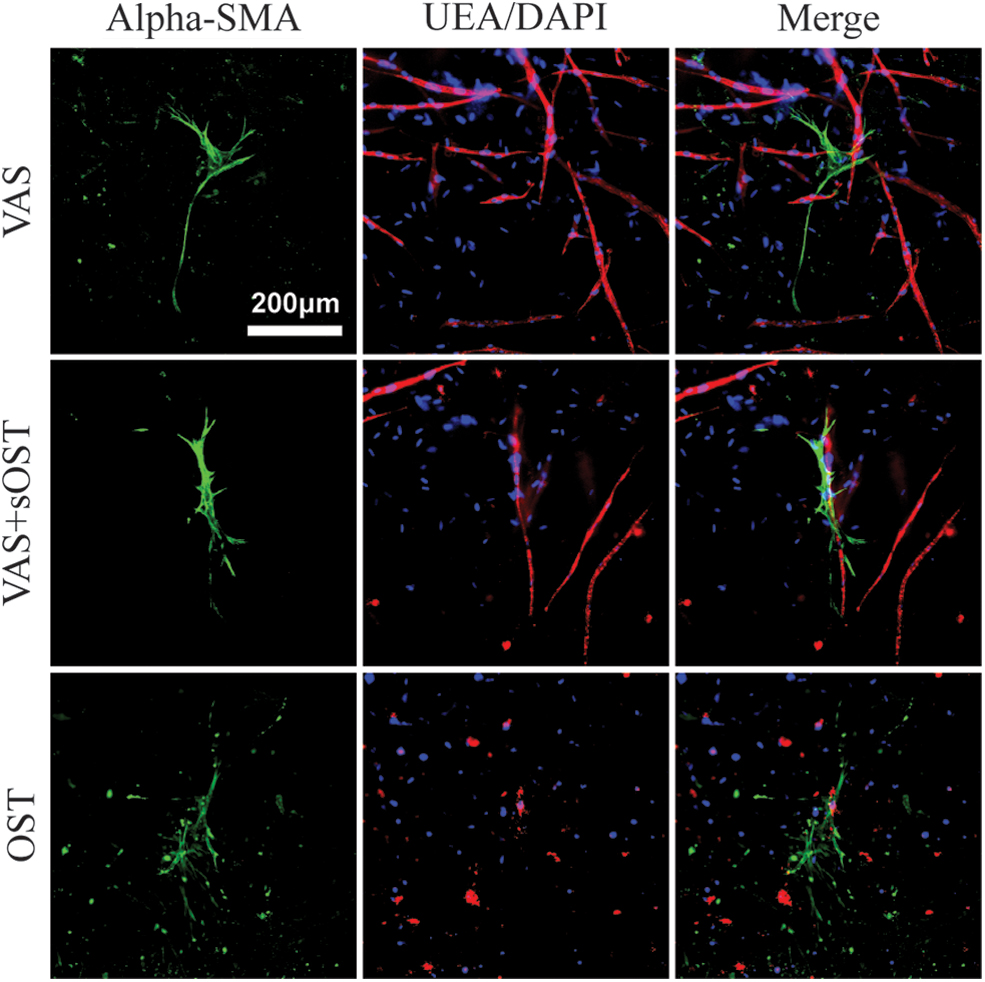
Immunofluorescent staining of constructs for alpha-SMA (green), endothelial cells (UEA, red) and cellular nuclei (DAPI, blue) at day 14 of culture. Best viewed in color. Color images are available online.
Prevascularized constructs—osteogenesis
To investigate the osteogenic capacity of MSCs in prevascularized constructs under varying culture media conditions, ALP activity and calcium deposition were quantified as shown in Figure 7. ALP activity was not significantly changed over 14 days after prevascularization when constructs were cultured in VAS (Fig. 7A). In VAS+sOST cultures, ALP activity increased significantly over the same period. Constructs cultured in OST showed increased ALP activity at 7 days after prevascularization, which leveled off through 14 days. Calcium deposition was not detected in VAS or VAS+sOST at any time point after prevascularization (Fig. 7B). However, constructs cultured in OST showed robust calcium deposition at both the 7- and 14-day time points.
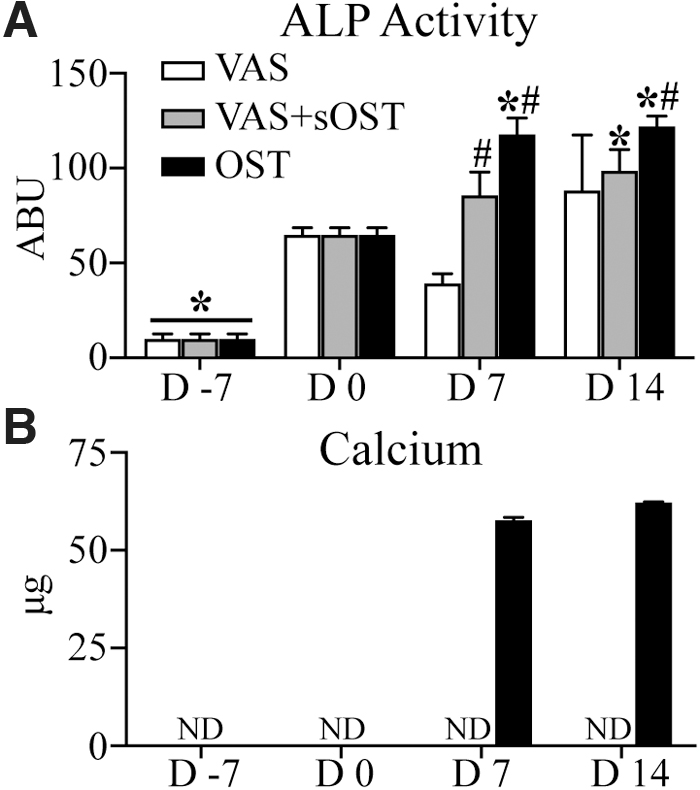
Osteogenic marker expression through the prevascularization and treatment periods. Quantification of average
Discussion
Vascularization of engineered tissues is a broad and continuing challenge in regenerative medicine. 5 This issue is of key importance for tissue engineering approaches to healing large and recalcitrant bone defects, since ischemia has been identified as a major factor in limiting tissue regeneration.34,35 Cell-based approaches to regenerating vascularized bone tissue have focused on the use of MSCs and ECs, since these cell types are known to be involved in the regeneration of both bone and vasculature. It has been demonstrated that preformed vessel networks can persist after transplantation 6 and enhance overall vascularization after transplantation in critical-sized bone defects. 7 Therefore, prevascularization of engineered constructs is a promising strategy to enhance tissue engraftment. Furthermore, inducing osteogenic lineage commitment in MSCs before transplantation has been shown to significantly augment their bone-regenerative capacity.32,36 Therefore, the creation of engineered tissues containing an osteogenic and vascular phase requires specialized culture conditions that concurrently support both phenotypes. Hybrid culture medium containing both proangiogenic and osteoinductive factors has been widely explored as a means to promote both vascular and osteogenic functions.21,26,31,37 Other groups have used an initial period of culture in vasculogenic medium alone to promote prevascularization. 30 However, most bone tissue engineering efforts have been limited to the formation of rudimentary vascular structures and/or hampered osteogenic activity. Therefore, to enable the creation of successful multiphase engineered tissues, there is a need to better understand how the culture protocol influences cellular behavior.
In previous studies, prevascularization of engineered bone constructs has generally been attempted over periods of 3–14 days, although there is no strong consensus on the optimal culture time.21,30,31,38 In the current study, a 7-day prevascularization period was chosen based on our own previous work.39–41 The presence of MSCs as support cells is known to promote EC survival and the formation of vascular networks through the secretion of proangiogenic factors. 17 Specific growth factors can also be provided exogenously in the culture media to induce angiogenic activation of ECs in coculture. 25 Human umbilical vein endothelial cells (HUVECs) were used in the present work because of their widespread application as models for understanding cell behavior and vascularization of engineered tissues.24,42 The clinical utility of HUVECs is limited by their relatively mature phenotype 43 and potential immunogenicity in allotransplantation, 42 although they have been shown to interact with MSCs and to produce robust vessel networks in vitro.40,44 Other potential cell sources include endothelial progenitor cells (EPC) 45 and induced pluripotent stem cell-derived ECs (iPSC-ECs). 46 In the current study, constructs containing only HUVECs did not produce EC networks when cultured in control medium. However, both EC+MSC coculture and exposure to vasculogenic medium produced multicellular and interconnected network structures. Exposure of cocultures to vasculogenic medium further enhanced EC network formation, showing the additive effect of both endogenous and exogenous factors.
The bone-forming capacity of MSCs is well established, and culture media have been developed for directing these progenitor cells toward the osteogenic lineage.12,47 In the present work, a standard osteogenic medium had a clear effect in upregulating the early marker ALP, as well as in robustly enhancing mineral deposition over 14 days of culture. The interactions between ECs and MSCs in the context of osteogenesis are not well understood, and a variety of effects have been reported in the literature. Some studies have indicated a positive effect on mineralization,48–50 while others have described molecular pathways through which ECs can inhibit osteogenesis.51,52 Interestingly, in the current study the presence of ECs did not have a potentiating effect on ALP activity, both in the presence and absence of osteogenic medium. However, EC+MSC cocultures achieved similar levels of mineral deposition compared with MSC monocultures, in spite of the fact that the cocultures contained half the initial number of MSCs. This is in agreement with previous work exploring adipose-derived MSCs in coculture with ECs, 53 and suggests that the presence of ECs may potentiate the effect of osteogenic medium on MSC mineral deposition. These results reflect the interplay between ECs and MSCs under different media conditions, and indicate that ECs can support osteogenesis.
Maintenance of EC viability and the ability to form vascular structures in the presence of osteoinductive factors is a challenge when creating engineered bone tissues.30,48 While osteogenic medium induces proliferation and differentiation of MSC, 12 it has also been shown to influence EC behavior in vitro.28,54,55 Specific media supplements, such as ascorbic acid and dexamethasone, have been found to induce EC apoptosis,28,29 and to reduce expression of cell adhesion molecules necessary for the development of multicellular networks. 27 In turn, reduced EC viability and proliferation are known to reduce vessel formation. 56 In the present study, the use of osteogenic medium resulted in reduced overall cell viability in cocultures over time, compared with pure vasculogenic medium. Similarly, EC network maintenance and progression were most evident in pure vasculogenic medium, while pure osteogenic medium caused complete regression of multicellular structures over time. Interestingly, the use of a hybrid (VAS+sOST) medium resulted in maintenance of EC network structures, although overall density was lower compared with the VAS condition at day 14. These findings are in agreement with previous work showing that the addition of osteogenic medium to cocultures of ECs and MSCs was detrimental to vascular formation, 30 yet a hybrid medium formulation was able to maintain the presence of networks within a hydrogel system. This work motivated our investigation of the role of MSCs in this multicellular system, with a focus on whether hybrid medium influences their role as supportive pericytes. The present study investigated the expression of αSMA, a marker of pericyte cells that play a role in the development and stabilization of neovessels. 57 αSMA-positive MSCs in both VAS and hybrid conditions exhibited a spindle-shaped morphology and colocalization to EC networks, thus suggesting a pericyte-like function. However, αSMA-positive MSCs in the osteogenic condition had a more diffuse morphology and did not colocalize with ECs. Altogether, these results suggest that pure osteogenic medium compromises preformed EC networks while influencing pericyte behavior.
Previous work has highlighted difficulties in achieving osteogenesis in environments permissive to vasculogenesis. In the present study, constructs cultured in hybrid or pure osteogenic media displayed an increase in ALP activity over time. In addition, only constructs in osteogenic medium exhibited detectable levels of mineral deposition, which is a more functional measure of bone formation in 3D matrices. These results are in agreement with previous work in two-dimensional culture systems, which found that osteogenically supplemented vasculogenic medium does not support osteogenic differentiation of MSCs.25,48 It has been suggested that specific vasculogenic medium components, such as FGF, 58 IGF-1, 59 EGF, 60 and VEGF 61 reduce the osteogenic potential of MSCs. Altogether, these findings confirm the ability to induce osteogenic activity within prevascularized constructs, although the extension of EC networks is compromised. The altered morphology of αSMA-positive MSCs and decreased colocalization with ECs in pure osteogenic medium suggests that this culture condition influences their pericyte-like behavior and ability to support EC network formation. However, it has been suggested that vascular pericytes can maintain a degree of osteogenic potential both in vitro and in vivo, 62 which motivates future work to explore the fate of MSCs in coculture systems. Understanding the phenotype shifts induced in coculture is a key to designing engineered constructs that support both vasculogenic and osteogenic functions.
Conclusion
The rational and controlled combination of ECs and MSCs in 3D constructs is a promising approach to achieving concomitant vascularization and bone formation in engineered bone tissues. Prevascularization in vitro may be a particularly effective means to enhance construct engraftment and survival after transplantation. The current study highlights the importance and influence of culture medium composition and culture protocol on supporting and maintaining EC network formation and mineral deposition. The results show that while vessel networks can be maintained over time in 3D fibrin constructs using a hybrid culture medium, it is challenging to achieve robust osteogenesis under conditions, which also support vessel maintenance. An advantage of 3D tissue engineering approaches is that the scaffold or matrix can be tailored to provide biomimetic cues to promote and guide bone regeneration. Therefore, more advanced tissue engineering approaches may be needed to create stable multiphase tissues, either by mimicking key aspects of natural bone development, or by developing tissue assembly systems in which osteogenesis and vasculogenesis can be promoted independently.
Footnotes
Acknowledgment
The authors gratefully acknowledge the assistance and guidance of their colleagues, and in particular Dr. Ben Juliar and Dr. Andy Putnam (U. of Michigan).
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported in part by the National Institute of Arthritis and Musculoskeletal and Skin Diseases (R01-AR062636, to JPS). NGS is partially supported by the Cellular Biotechnology Training Program (T32-GM008353) at the University of Michigan. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. Some of the figures in this article were created using Servier Medical Art templates, and are licensed under a Creative Commons Attribution 3.0 Unported License (smart.servier.com).