
Abstract
Metal orthopedic implants are largely biocompatible and generally achieve long-term structural fixation. However, some orthopedic implants may loosen over time even in the absence of infection. In vivo fixation failure is multifactorial, but the fundamental biological defect is cellular dysfunction at the host–implant interface. Strategies to reduce the risk of short- and long-term loosening include surface modifications, implant metal alloy type, and adjuvant substances such as polymethylmethacrylate cement. Surface modifications (e.g., increased surface rugosity) can increase osseointegration and biological ingrowth of orthopedic implants. However, the localized responses of cells to implant surface modifications need to be better characterized. As an in vitro model for investigating cellular responses to metallic orthopedic implants, we cultured mesenchymal stromal/stem cells on clinical-grade titanium disks (Ti6Al4V) that differed in surface roughness as high (porous structured), medium (grit blasted), and low (bead blasted). Topological characterization of clinically relevant titanium (Ti) materials combined with differential mRNA expression analyses (RNA-seq and real-time quantitative polymerase chain reaction) revealed alterations to the biological phenotype of cells cultured on titanium structures that favor early extracellular matrix production and observable responses to oxidative stress and heavy metal stress. These results provide a descriptive model for the interpretation of cellular responses at the interface between native host tissues and three-dimensionally printed modular orthopedic implants, and will guide future studies aimed at increasing the long-term retention of such materials after total joint arthroplasty.
Impact statement
Using an in vitro model of implant-to-cell interactions by culturing mesenchymal stromal cells (MSCs) on clinically relevant titanium materials of varying topological roughness, we identified mRNA expression patterns consistent with early extracellular matrix (ECM) production and responses to oxidative/heavy metal stress. Implants with high surface roughness may delay the differentiation and ECM formation of MSCs and alter the expression of genes sensitive to reactive oxygen species and protein kinases. In combination with ongoing animal studies, these results will guide future studies aimed at increasing the long-term retention of widely used titanium materials after total joint arthroplasty.
Introduction
Total joint arthroplasty is both clinically efficacious and cost effective.1,2 However, a significant portion of patients can experience long-term complications owing to poor implant fixation.3–7 A common strategy to reduce orthopedic implant loosening is to increase the surface roughness of an implant.8–10 Both clinical experience and in vivo studies have established that osseointegration improves with increased macro-, micro-, and nanoscale surface roughness of titanium (Ti) alloy implants.11–14 Furthermore, in vitro studies have demonstrated that roughened Ti alloy materials can increase the osteogenic potential, differentiation, and extracellular matrix (ECM) mineralization of human mesenchymal stromal cells (MSCs).15–20
Reduced osseointegration may be provoked in part by corrosive processes over time that result in the disintegration of implant material, concomitant with local or systemic release of wear debris, including Ti particles.10,21 These and other potential adverse effects of metal contact on cellular processes have not yet been thoroughly documented, and may, in principle, be magnified with increased contact area.
Cells cultured in contact with Ti alloys exhibit oxidative stress, which results in elevated levels of reactive oxygen species (ROS) within the culture medium.22–25 ROS are known to disrupt cellular functions by binding essential nucleic acids, proteins, and lipids. 26 Specifically in human MSCs, ROS can interfere with Wnt, Hedgehog, and FOXO signaling cascades, 26 potentially altering early stages of osteogenic differentiation. Elevated ROS production, in addition to other limitations of metallic biomaterials (e.g., stress shielding), may contribute to implant loosening. Therefore, it is critical to determine whether implants of increased surface roughness affect cellular oxidative stress. Heavy metal stress is a cellular stress response induced by exposure to a heavy metal (or particles) (e.g., Baranowska-Wojcik et al. 27 ), so we refer to the stress-related response of human MSC exposure to metal orthopedic implant materials as oxidative and/or heavy metal stress because the two processes are often correlated metal exposure to specific cell types (e.g., Paithankar et al. 28 ).
This study investigates the relationship between the surface roughness of Ti orthopedic implants and cellular stress. Our primary goal was to document the ROS-related responses of MSCs to clinically relevant Ti orthopedic substrates with three grades of surface roughness. We also characterized the MSC response (e.g., cell proliferation, ECM production) to Ti orthopedic substrates. Human bone- and adipose-derived MSCs (bone-derived mesenchymal stromal cells [BMSCs] and adipose-derived mesenchymal stromal cells [AMSCs], respectively) were cultured on Ti alloy (Ti6Al4V) substrates of high, medium, and low roughness.
We utilized three groups of clinical-grade Ti6Al4V constructs that differed only in surface roughness (bead blasted = low, grit blasted = medium, and porous structured = high). Each material surface type was characterized using light interferometry to obtain detailed topographic measurements. Whole-genome RNA-seq and real-time quantitative polymerase chain reaction (RT-qPCR) data were collected for select mRNAs that represent biomarkers for cellular stress, differentiation, and proliferation. Our findings indicate that increased surface roughness is associated with increased transcription of ROS-related and metal-related cellular stress genes. The results of this study provide insight into possible mechanisms underlying aseptic orthopedic implant loosening.
Materials and Methods
Fabrication of Ti biomaterials
Porous-structured (ps-Ti), grit-blasted (gb-Ti), and bead-blasted (bb-Ti) Ti disks were provided by Stryker Orthopedics (Mahwah, NJ). Each disk was composed of alloyed Ti, aluminum (6%), and vanadium (4%) (Ti6Al4V) and measured 3 mm in height by 25 mm in diameter. Previous studies describe the precise fabrication methods for the ps-Ti disks.15,29 In short, ps-Ti disks were manufactured using computer-guided laser sintering with Ti6Al4V (grade 5) particles of 25–45 μm in two-dimensional layers. This process was repeated layer-by-layer to construct a three-dimensional (3D) disk that models the porous structure (65% porosity) of human trabecular bone. 29
To manufacture gb-Ti disks, Ti6Al4V wrought bar stock was machined to the desired dimensions and subjected to 80 grit aluminum oxide blast media. Silica glass bead media were used to polish the bb-Ti disks that were machined from the same wrought bar stock. Industrial air pressure blasting cabinets were used with standard air pressure of ∼90 psig. Gb-Ti and bb-Ti disks were then submerged in distilled water in a Branson ultrasonic cleaning unit for 15 min twice to remove any remaining blast media. All disks were dried and packaged for terminal sterilization by 25–40 kGy gamma irradiation dose.
Surface roughness characterization
Surface roughness (Ra) was measured for each Ti biomaterial using a Zygo Newview White Light Interferometer (Zygo Corporation, Middlefield, CT). Representative samples of each Ti biomaterial (ps-Ti, gb-Ti, and bb-Ti) were measured at 10 × magnification. Five locations on each sample were measured for maximum, minimum, and mean (±standard deviation [SD]) Ra to the nearest 0.01 μm, and three samples per material were analyzed for statistical comparison (mean ± SD; Fig. 1 and Supplementary Fig. S1). As a control material, one well from a six-well tissue culture polystyrene plate was also measured at five locations and included for comparison with Ti biomaterials.
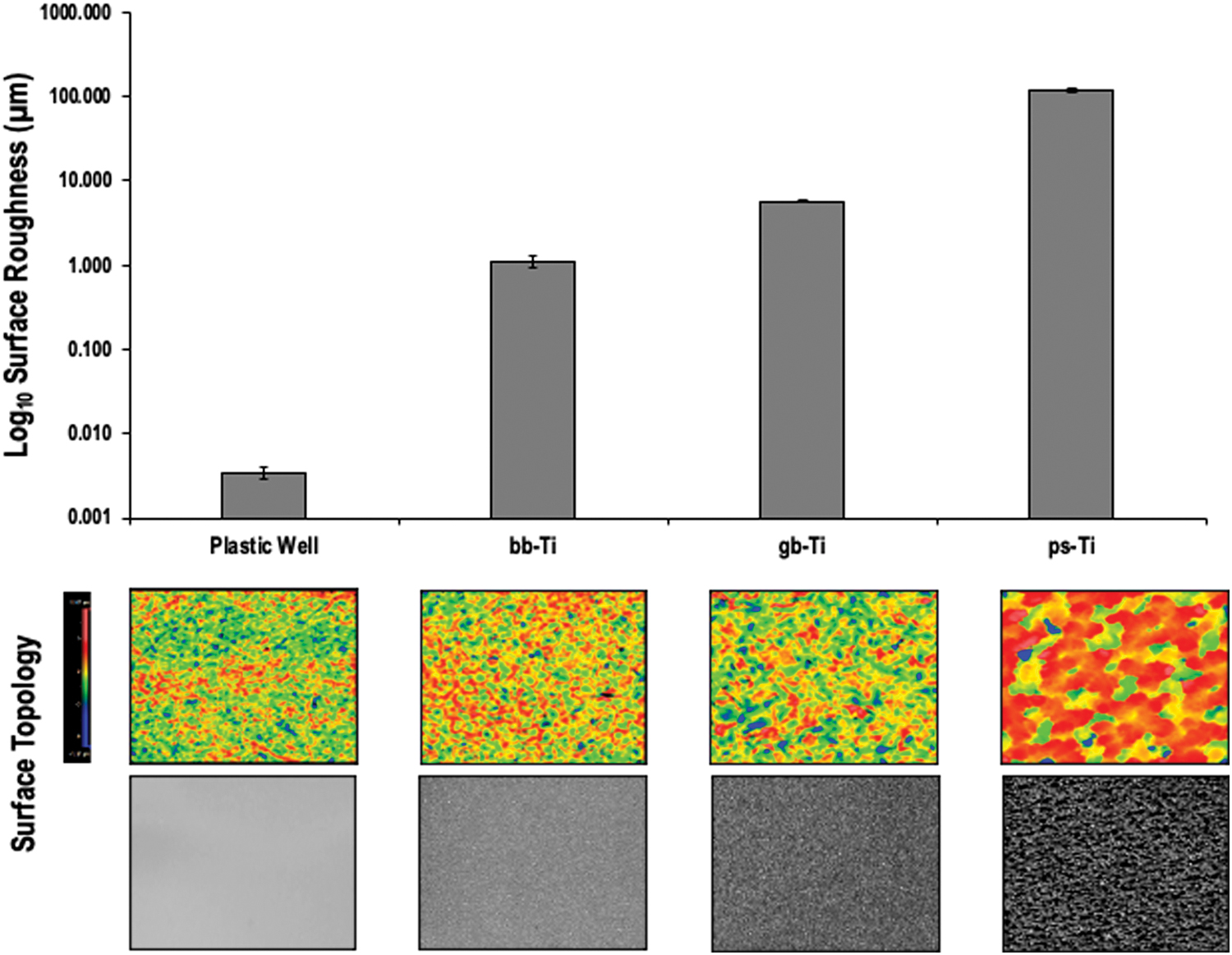
The surface roughness characterization of each material used in cell culture experiments highlights a near 100-fold difference between plastic (low Ra) and ps-Ti (high Ra). Measurements and representative colorimetric and photographic images of the surface topology of each material were obtained by light interferometry. bb-Ti, bead-blasted titanium; gb-Ti; grit-blasted titanium, ps-Ti; porous-structured titanium. Color images are available online.
Scanning electron microscopy
Scanning electron microscopy (SEM) images were obtained for each naked Ti substrate. In addition, SEM images were obtained for cells cultured on each substrate at each time point. Each Ti disk seeded with cells was immersed in Trumps tissue fixative. These samples were then rinsed with solutions of increasing concentrations of ethanol: (1) phosphate-buffered saline (PBS) (2 × ); (2) water (2 × ); (3) 10% ethanol; (4) 30% ethanol; (5) 50% ethanol; (6) 70% ethanol; (7) 90% ethanol; (8) 95% ethanol; and (9) 100% ethanol (2 × ). The 100% ethanol was evaporated using a critical point dryer, and the disks were then fixed to an aluminum plate using double-sided conductive tape. Gold-palladium dust was sputter coated onto the disks for 90 s to improve electroconductivity of the sample. For each sample, images were obtained using a cold-field emission SEM (S-4700; Hitachi, Japan) at low (30 × ), medium (200 × ), and high (1300 × ) magnifications.
Isolation of MSCs
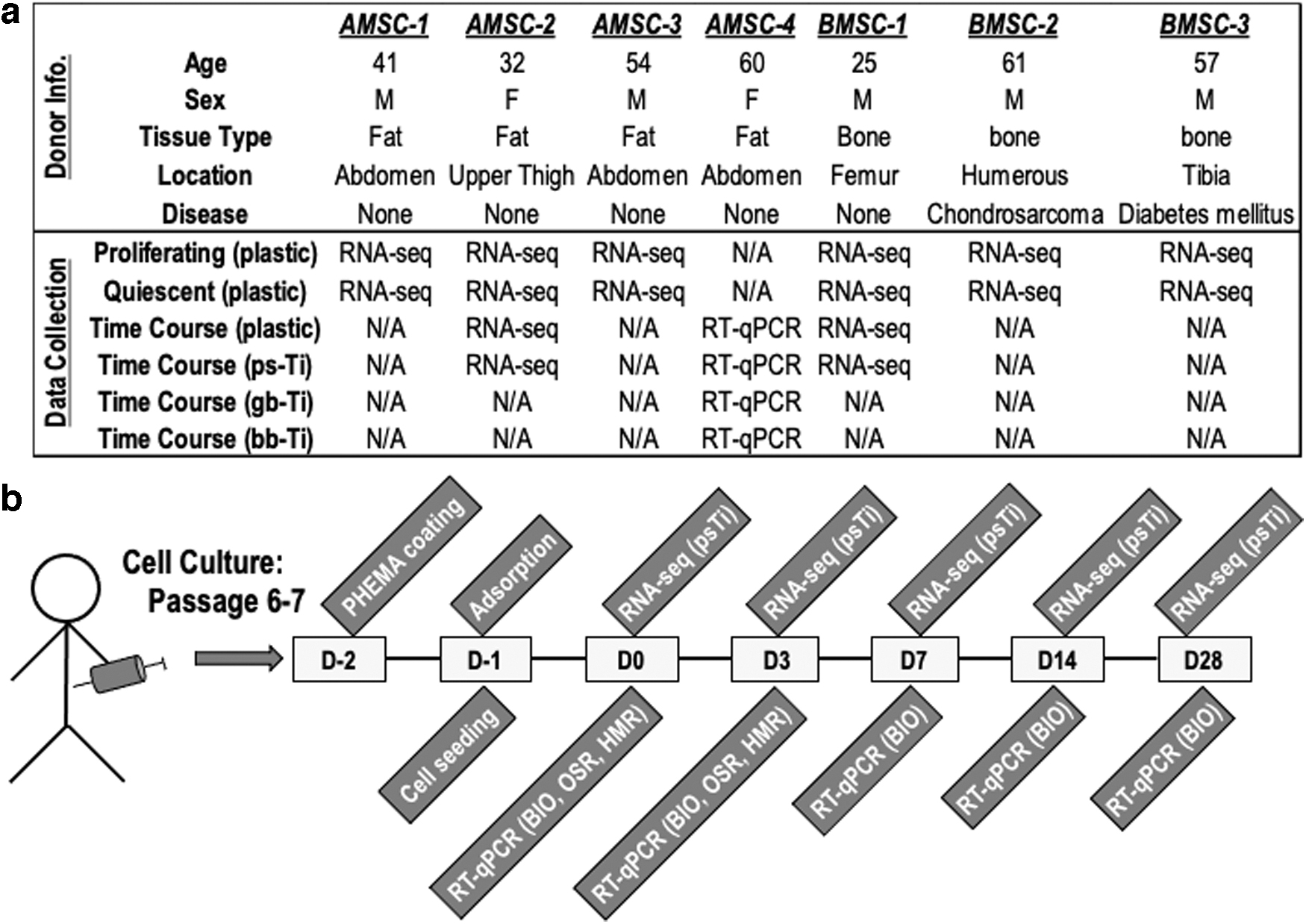
After obtaining institutional approval and written individual consent from each patient (Mayo Clinic Institutional Review Board (IRB-13-005619), Rochester, MN), clinical samples of adipose and bone tissue were opportunistically obtained from both male and female human donors to provide the source materials for all MSCs used in this study. Four adult patients (32–60 years of age) were the donors of adipose tissues that were subjected to a series of enzymatic digestions to isolate AMSCs, and three adult patients (25–61 years of age) were the donors of cancellous bone used for tissue outgrowth culture to isolate BMSCs (Fig. 2a).
After liposuction of adipose tissue from donors, tissues were minced using a scalpel blade and incubated at 37°C for 90 min in 0.075% type I collagenase (≥125 U per mg dry weight; Worthington Biochemicals, Lakewood, NJ). Digested tissues were centrifuged at 400 g for 5 min to establish a pellet that was subsequently washed in PBS, and passed through a 70-μm cell strainer (BD Biosciences, San Jose, CA). Red blood cells were removed by incubating the washed pellet in red blood cell lysis buffer (154 mmol/L NH4Cl, 10 mmol/L KHCO3, 0.1 mmol/L EDTA).
AMSCs were maintained in zoonotic free medium for clinical utility, characterized by RNA-seq for cell surface markers (PHY1, ND5E, ENG, and CD44), and validated for multilineage potential.30–33 AMSCs used for experimentation were cultured in advanced MEM (Life Technologies, Grand Island, NY) with 5% human platelet lysate (PLTMax; Mill Creek Life Sciences, Rochester, MN), 2 mM Glutamax (Life Technologies), 2 U/mL heparin (Fresenius-Kabi, Lake Zurich, IL), 1% Penn-Strep (100 U/mL penicillin; Life Technologies), and 100 μg/mL streptomycin (Cellgro, Corning, NY) to passage six or seven before experimentation. Detailed procedures for isolating and culturing AMSCs have been described previously.30–35
BMSCs were isolated as outgrowth cultures from fresh surgical waste tissue (cancellous bone) and maintained in Dulbecco's modified Eagle medium base media that contained bovine serum albumin (BSA) (instead of human platelet lysate). Similar to AMSCs, cells were maintained in culture until passage of six or seven to obtain sufficient cell numbers for experimentation, characterization by RNA-seq for cell surface markers (CD44, CD73/ND5E, CD90/PHY1, and CD105/ENG), and to allow for increased homogeneity of cultured cell populations.
Culture conditions
MSCs were cultured in standard six-well polystyrene plates (Corning, Inc., Corning, NY). On each six-well plate, three wells contained MSCs seeded onto Ti disks, and three wells contained MSCs seeded onto standard tissue culture polystyrene. Before seeding MSCs on the Ti disks, experimental wells were coated with poly 2-hydroxyethyl methacrylate (PHEMA; Sigma-Aldrich, St. Louis, MO) to prevent cells from attaching to the underlying tissue culture plastic, ensuring that all cell growth was limited to Ti disks. The PHEMA-coated wells were dried and sterilized using UV-light for 24 h. A single Ti disk was placed into each PHEMA-coated well and seeded with 2.5 × 105 cells suspended in 500 μL of 5% human platelet lysate media (BMSCs were treated with BSA instead of human platelet lysate). The same concentration of cells in 500 μL of this media was simultaneously seeded onto the plastic control wells.
To adequately submerge the Ti materials, another 500 μL of this media was then placed in the base of each well. The plates were placed in a standard incubator (37°C and 5% CO2) for 2 h to accomplish full adsorption onto the Ti disks. After 2 h of adsorption, 2 mL of culture media was added to each well. After 24 h, the media was replaced with osteogenic media, which was subsequently changed twice weekly throughout the study period. The following nomenclature was used for defining each time point: day 2 was 48 h before cell seeding, day 1 was the day of seeding, day 0 was 24 h after cell seeding, day 3 was 72 h after cell seeding, day 7 was at 1 week after seeding, day 14 was at 2 weeks after seeding, and day 28 was 4 weeks after seeding (Fig. 2b). RNA-seq data (see hereunder) was used to characterize the MSCs in our study and these cells express mRNAs for classic MSC biomarkers (e.g., CD90/THY1, CD73/NT5E, CD105/ENG, and CD44) 33 (data not shown).
mRNA analysis
Cells cultured on substrates were lysed at each time point using 700 μL of Qiazol (Qiagen, Valencia, CA). The Ti disks were rinsed twice with the same aliquot of Qiazol to ensure full lysis of all cells that adhered to the substrate. Genomic RNA was then extracted with the Qiagen miRNeasy Mini Kit (Qiagen). RNA concentration and purity values were quantified with UV-Vis spectrophotometry (NanoDrop 2000; Thermo Scientific, Wilmington, DE). Complimentary DNA was synthesized from mRNA through reverse-transcriptase PCRs using the Superscript II RT kit (Invitrogen, Waltham, MA). Whole-transcriptome sequencing (RNA-seq) was then performed for MSCs cultured on ps-Ti at all time points. RNA samples were submitted to the Medical Genome Core Facility (Mayo Clinic, Center for Individualized Medicine, Rochester, MN).
Using the RNA-seq data collected for MSCs cultured on ps-Ti and tissue culture plastic (Fig. 2), we prioritized genetic markers related to cellular stress and biological function. RNA-seq cluster analyses included Time Course Network Enricher (TiCoNE), 36 Gene Set Enrichment Analysis (GSEA), 37 and PVCLUST 38 using data filtered for FPKM >0.3, and 1,000,000 bootstrap replicates to calculate approximate unbiased p-values and Bayesian posterior probabilities. Furthermore, RNA-seq data were analyzed by principal component analysis (PCA) using ClustVis, 39 heat map visualization using Morpheus, 40 and functional protein–protein network assembly using STRING v.11. 41 Gene ontology assignments were performed on selected subsets (identified as clades in the heat map analysis) of the RNA-seq data using David v.6.8.42,43
RT-qPCR was used to investigate a subset of genetic markers in cells cultured on all three Ti materials and control tissue culture plastic (Fig. 2). Genetic markers were prioritized based on ongoing RNA-seq data accrual and curation, which is aimed at analyzing cells derived from hundreds of musculoskeletal tissues and cells, including cells from peri-implant tissues recovered from failed joint replacements (unpublished data). Such analyses have allowed formulation of specific hypotheses concerning the potential impact of exposing cells to various titanium biomaterials by focusing on molecular markers that we suspect to be involved in cellular phenotypic changes (e.g., oxidative stress) at the interface between musculoskeletal tissues and metallic implants.
Complementary DNA sequences were amplified using the Superscript III RT kit (Invitrogen) as per the manufacturer's instructions in a total volume of 40 μL. RT-qPCRs were performed in a total volume of 10 μL and included: 2.5 μL of cDNA, 5 μL QuantiTect SYBR Green (Qiagen), and 2.5 μL of sequencing primers. The RT-qPCR measurements (Ct values) were made with a CFX384 Real-time System c1000 Touch Thermal Cycler (BioRad, Hercules, CA). Ct values for each transcript were normalized to GAPDH*100 using the 2−ΔΔCt method. An overview of the experimental design is given in Figure 2.
Results
Physical and ultrastructural characterization of titanium surfaces
To provide a physical framework for understanding the biological properties of human MSCs upon culture on Ti surfaces, we examined the surface roughness using interferometry. All Ra measurements (minimum, maximum, mean and SD to the nearest 0.01 μm) conducted by light interferometry for each substrate are given in Figure 1 and Supplementary Figure S1. ps-Ti had the highest mean Ra values (118.19 ± 9.06 μm), whereas grit-blasted (5.74 ± 0.19 μm) and bead-blasted (1.10 ± 0.18 μm) substrates had lower roughness measurements than ps-Ti. An ∼20-fold to 100-fold increase in the complexity of ps-Ti surfaces compared with polished substrates, substantially increases the surface area available for cell growth and proliferation on this material.
To visualize the surface of Ti surfaces at the ultrastructural level, we performed SEM. Images of each material were obtained at 30 × without cells and with cells cultured for both 7 and 28 days (Fig. 3). Cells covered the ps-Ti material with clear formation of a 3D ECM spanning microscale topological features at both 7 and 28 days after cell seeding (Fig. 3; left column). The gb-Ti material showed clear ECM formation with observable polarization and low-relief structural adhesion to the material (Fig. 3; middle column). In contrast, cells readily detached from the bb-Ti material and partially peeled away from the smooth surface after 7 days; however, cells were able to generate an intact layer on gb-Ti after 28 days (Fig. 3; right column). Differences in cell adhesion per Ti material should, in principle, confer detectable alterations in mRNA profiles.
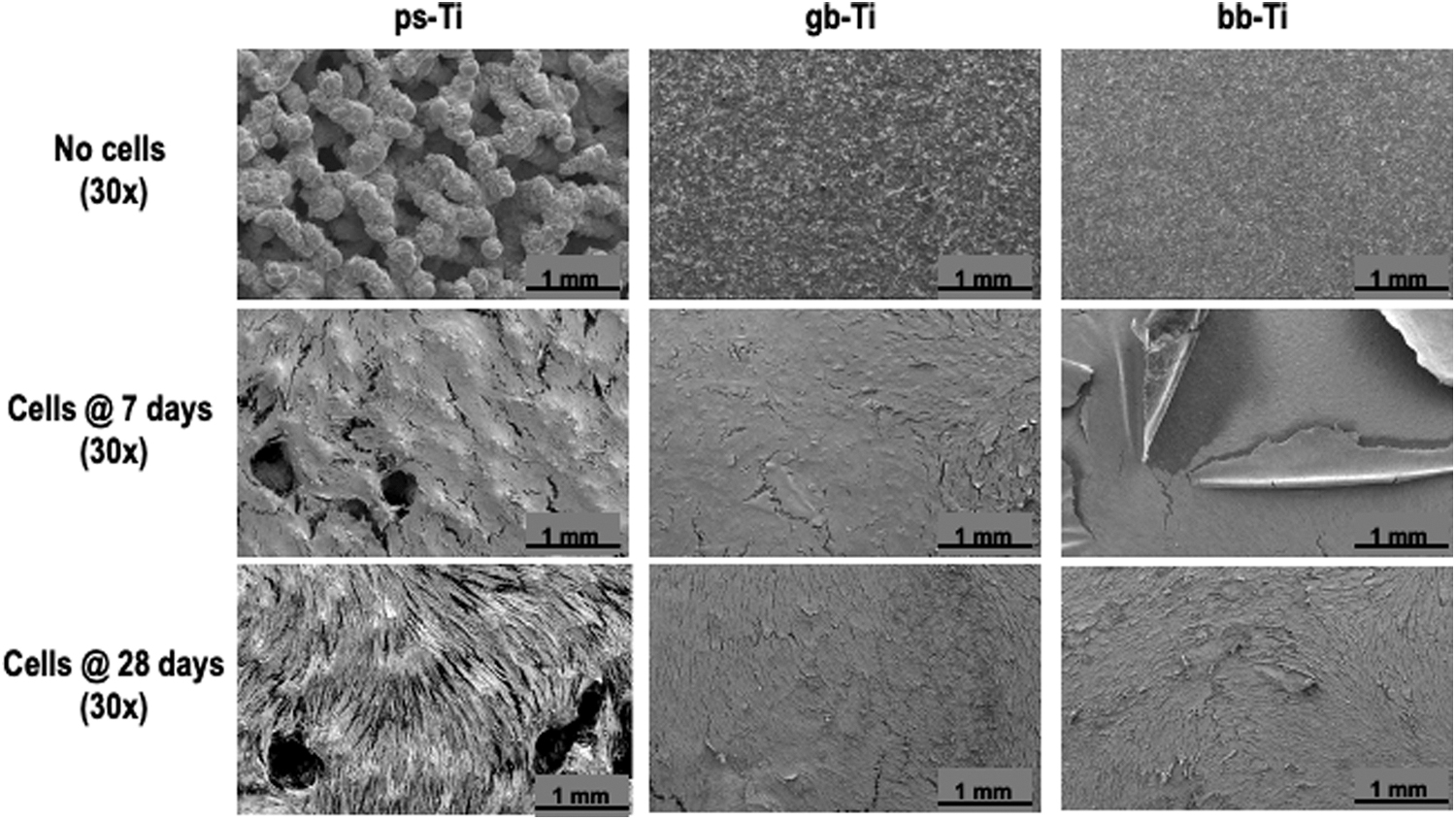
SEM was used to visualize the implant materials with and without cultured cells. Interstitial spaces observed on the ps-Ti surface (e.g., top panel, left) were populated by attached cells (e.g., bottom panel, left), whereas lower Ra materials had “flaked” patterns of reduced cell adhesion (e.g., middle panel, right). Cell populations were confluent when cultured on each material by day 7 (or earlier). SEM, scanning electron microscopy.
Transcriptome characterization and mRNA biomarker analysis of MSCs cultured on Ti substrates
RNA-seq analysis was performed to examine the transcriptomes of MSC growth on ps-Ti versus plastic. Two different types of clustering analyses were performed to establish temporal changes in RNA expression (GSEA) and overall differences (PVCLUST) among sample types. Gene set enrichment analyses revealed 23 total clusters and highlighted 7 clusters of particular interest owing to consistent trends over time (Supplementary Fig. S2).
Clusters were characterized as decreasing (e.g., clusters 3 and 23), increasing (e.g., clusters 7, 19, 22), or altered at day 3 (e.g., cluster 11 up at day 3, and cluster 5 down at day 3). Clusters 3 and 23 encompass various genes related to growth factor signaling and cancer (e.g., oncogenes and tumor suppressors), whereas clusters 7, 11, 19, and 22 contain genes for a variety of glycoproteins and ECM proteins. Of interest, the PVCLUST analysis generated well-supported clusters primarily based on cell type (AMSC vs. BMSC) and time in culture (e.g., proliferating, confluent, and >7 days) (Supplementary Fig. S3). Taken together, this initial bioinformatics analysis revealed stage-specific waves of gene expression in MSCs grown on Ti versus plastic.
Our RNA-seq data encompass AMSCs and BMSCs that are each grown for 4 weeks on ps-Ti disks. For comparison, we examined AMSCs and BMSCs that were grown on plastic tissue culture vessels either as subconfluent or confluent cultures. We filtered our gene set of >20,000 protein-coding genes for a minimal level of expression (FKPM >0.3) and for twofold changes (increased or decreased) between samples grown on plastic versus ps-Ti to obtain a set of ∼4000 genes of interest. PCA for this selected gene set using ClustVis 39 reveals that each biological condition separates well in the first and second principal component, which account for, respectively, 19.5% and 24.0% of total variation in gene expression among the samples (Fig. 4a).
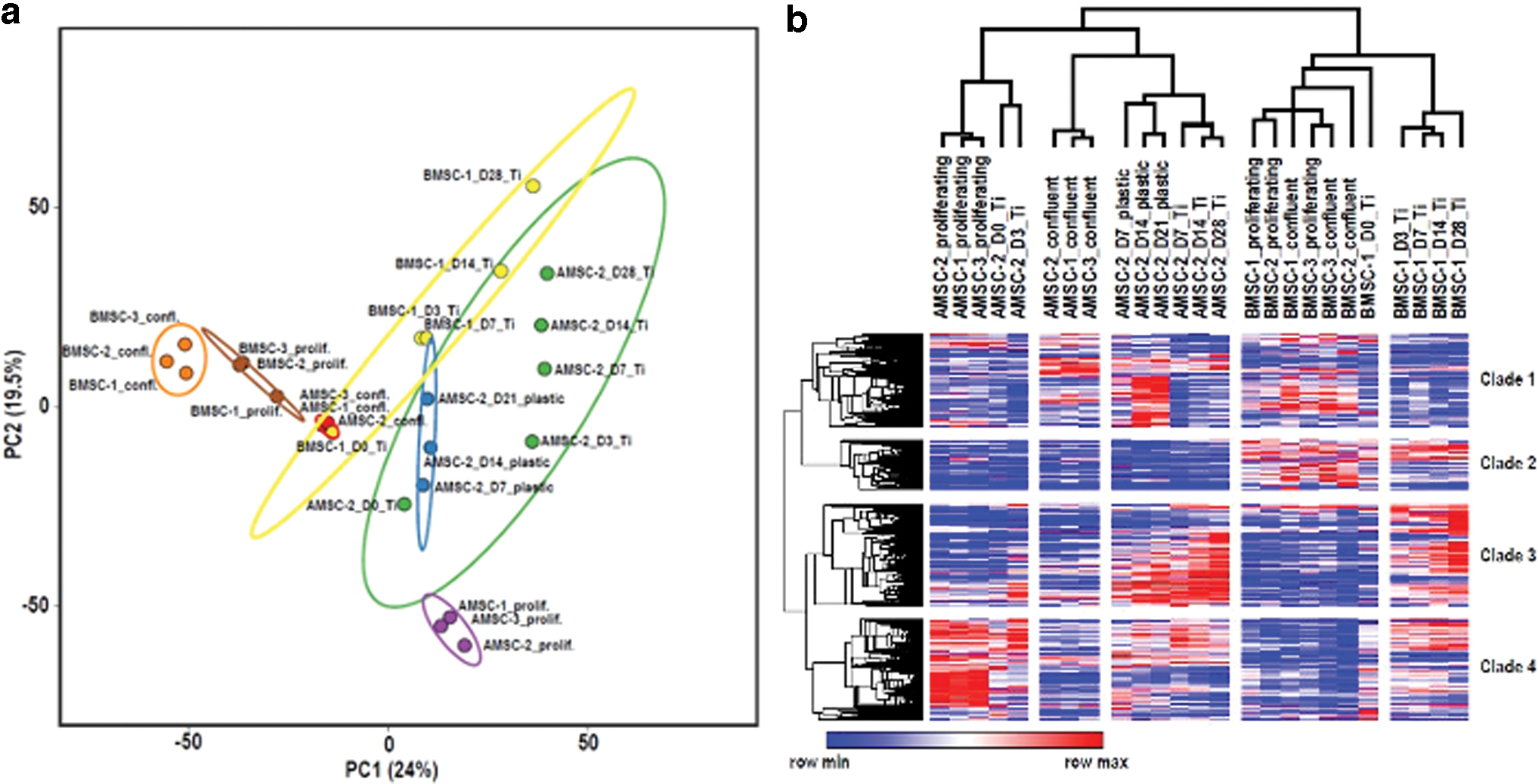
Hierarchical clustering and heat map analysis of these same samples using Morpheus 40 revealed five major clades of biological conditions and four distinct clades of differentially expressed genes (Fig. 4b). The two main biological clades were driven by cell type-specific differences between AMSCs and BMSCs. The second clade subdivides the samples into actively proliferating cells (regardless of cell culture surface) versus confluent and/or differentiated cells. The third subdivision is associated with differences in ps-Ti versus plastic surface. Thus, cell type and cell cycle-related differences are the main drivers of differences between the samples, although that growth on the ps-Ti surface also generates a comparatively modest gene expression difference.
Each of the five biological clades (horizontal) are associated with four distinct gene clades. Each of these gene clades exhibits elevated expression for ∼500 to 1200 distinct genes that in each case match with one or more of the biological samples (Fig. 4b). To understand what types of genes are contained within these four gene clades, we analyzed each for regulatory networks by STRING v.1141 analysis (Fig. 5 and Supplementary Fig. S4a–d) and gene ontology analysis by David v.6.8.42,43

The gene lists within each clade identified using Morpheus were subjected to protein network analysis using STRING. For reference, gene names associated with each string analysis are included in Supplementary Figs. S4a–d, and can be made available upon request. Color images are available online.
The results show that each of these four gene clades is associated with complex gene circuits encompassing multiple cell functions. For example, gene clade 1 contains four subnetworks that are well interconnected with each other. This clade 1 is linked to samples that are nonproliferative and/or have been cultured for multiple weeks. The subclusters within gene clade 1 are generally associated with biochemical and molecular properties characteristic of glycoproteins (including secreted factors), ECM/cell interactions, and ECM remodeling (Supplementary Fig. S4a). Similarly, clade 2 encompasses two subnetworks and is mostly associated with bone-derived MSCs, and contains genes that match gene ontology terms related to membrane-associated glycoproteins (e.g., cell surface receptors) and intracellular signaling (Supplementary Fig. S4b).
Clade 3 not only contains membrane-associated glycoproteins, but also genes involved in fatty acid metabolism, inflammatory responses, redox reactions, and lysosome function. Clade 3 has six interconnected networks that are characteristic of AMSCs and BMSCs grown on ps-Ti (Supplementary Fig. S4c), whereas clade 4 forms a massive network with at least seven subdivisions that directly reflect cell proliferation and contain numerous well-known cell cycle genes (Supplementary Fig. S4d). In addition, clade 4 contains genes linked to metallothioneins and oxidoreductase activity. For each of these gene clades, we have provided representative genes that are preferentially expressed in certain biological conditions (Supplementary Fig. S5).
Collectively, beyond the obvious transcriptome differences related to cell type and cell proliferation, these finding indicate that growth of either AMSCs or BMSCs on ps-Ti may have a modest but measurable effect on global transcriptomes and the expression of select groups of genes.
Differences in the expression of mRNA biomarkers reflect rugosity-dependent differences in MSC proliferation and ECM deposition.
RT-qPCR analysis was performed to examine specific markers of MSCs on Ti with different topologies to understand how Ti topology affects gene expression. The adipogenic potential of cultured AMSCs on differing substrates (e.g., reflected by expression of adiponectin/ADIPOQ mRNA) was similar at day 7 but increased in cells cultured on ps-Ti and plastic. Biomarkers of osteogenic differentiation (alkaline phosphatase/ALPL, bone sialoprotein/IBSP, transcription factor SP1, osteoprotegerin (OPG)/TNFRSF11B, osteocalcin/BGLAP) were consistently elevated in cells cultured on plastic; however, discernible differences among cells cultured on titanium surfaces were not detected. As an exception, ALPL was nearly threefold higher on gb-Ti cells than the lowest values observed within the ps-Ti group (Fig. 6). Cells cultured on low-rugosity substrates (plastic, bb-Ti, and gb-Ti) exhibited elevated expression of the principal collagens COL1A1 and COL3A1 at early time points (Fig. 6).
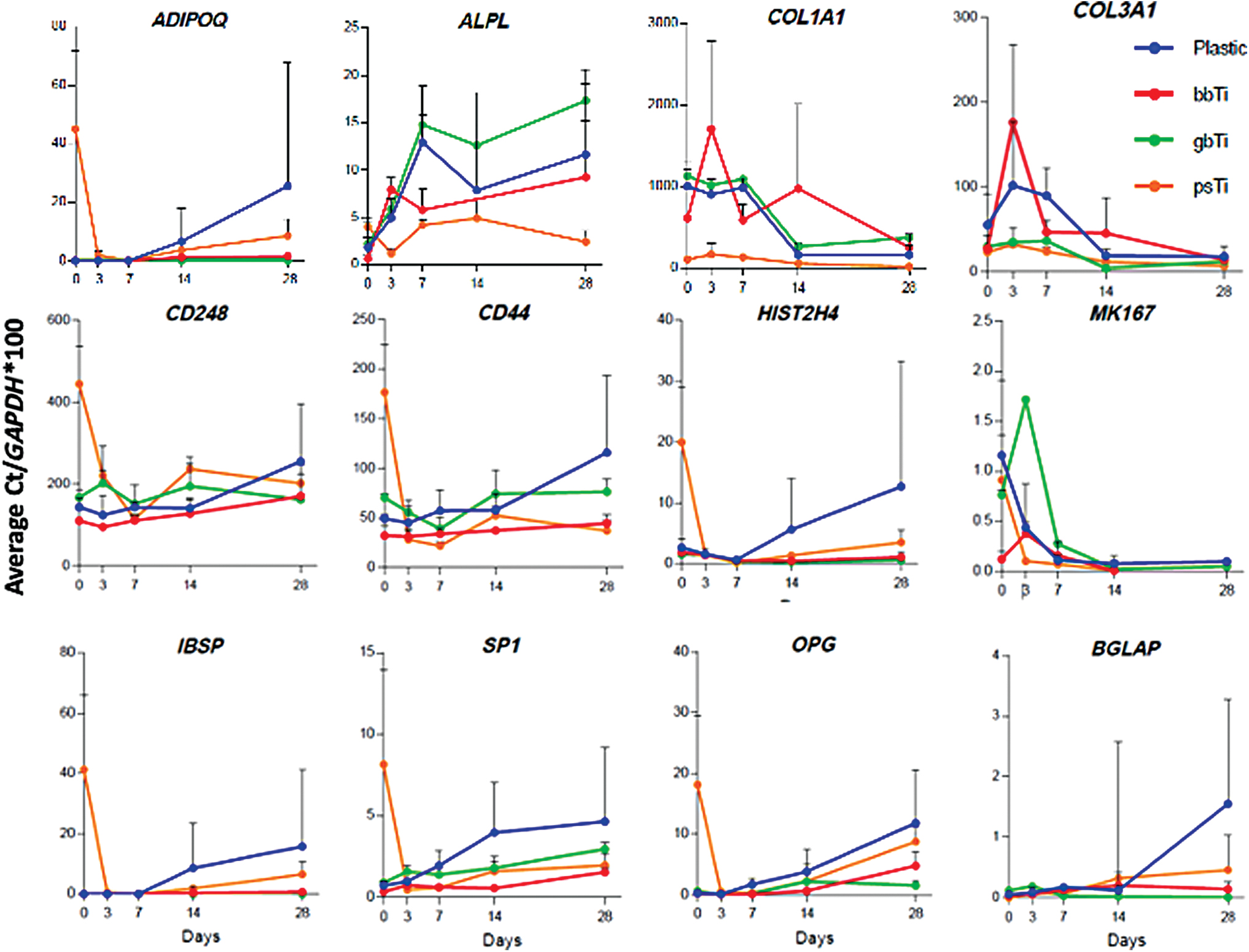
Cells were cultured on plastic, bb-Ti, gb-Ti, and ps-Ti for 28 days and collected at five time points to obtain RT-qPCR data of relative gene expression. Cell culture experiments were performed in triplicate, allowing calculation of mean and SD estimates. Average gene expression values for select biomarkers were normalized using Average Ct/GAPDH*100 to indicate comparative trends in gene expression among populations of cells cultured on each substrate (blue = plastic; red = bb-Ti; green = gb-Ti; orange = ps-Ti). SD, standard deviation. Color images are available online.
In addition, the expression of COL3A1 stabilized by day 28, and was similar among cells cultured on all substrates (Fig. 6). COL1A1 gene expression by MSCs differs by substrate after 3 days in culture, yet remained elevated among cells cultured on smoother substrates, suggesting higher ECM deposition and time-dependent alterations in the formation of ECM (Fig. 6).
MSCs cultured on ps-Ti appear to take longer to reach confluence than on the other surface materials because ps-Ti has a larger surface area where cells can expand and divide. MSC cell proliferation was measured using HIST2H4 and MKI67 mRNA levels as a proxy for proliferating cells. The HIST2H4 gene encodes a classical histone H4 protein that is required for packaging newly replicated DNA into chromatin, which is highly expressed during S phase. The MKI67 gene encodes a proliferation-specific antigen that is used to discriminate between actively dividing and nonproliferating quiescent cells. HIST2H4 expression decreased in all surface roughness groups after 7 days in culture (Fig. 6) but increased in cells grown on plastic. Cells cultured on ps-Ti, when compared with plastic controls, exhibited elevated HIST2H4 expression at earlier time points (days 0 and 3). Similarly, cells cultured on ps-Ti exhibited much higher levels of HIST2H4 at day 0 compared with gb-Ti and bb-Ti. The levels of HIST2H4 expression, however, were similar between substrates at day 3 (Fig. 6). At later time points (days 14 and 28), there was a clear separation in the expression of HIST2H4 in cells cultured on ps-Ti versus tissue culture plastic. MSCs cultured on smooth substrates (bb-Ti and gb-Ti) reached confluence earlier, and showed elevated ECM-related gene expression levels sooner than cells cultured on ps-Ti. At day 3, cells cultured on ps-Ti showed a twofold higher expression of stem-cell related markers compared with cells cultured on tissue culture plastic, suggesting a lack of early lineage commitment.
For comparison, MKI67 gene expression was elevated within the first 3 days of culture for cells in all four groups, and steadily diminished after day 7 (Fig. 6). The differential expression of HIST2H4 and MKI67 in MSCs indicates the respective fractions of proliferating cells that are either progressing through S phase (reflected by HIST2H4) or that are nonquiescent but not necessarily in S phase (reflected by MKI67). In more general terms, our data indicate that MSCs can proliferate on Ti surfaces regardless of topology, but that cells grown on high-rugosity substrates may take longer to reach confluence before differentiation.
At early time-points (days 0 and 3) stem cell markers CD44 and CD248 were elevated among cells cultured on ps-Ti compared with those cultured on smoother substrates. This difference in stem cell marker expression became less pronounced at later time points (Fig. 6). These results suggest that cells cultured on high-rugosity substrates may change the expression of stem cell surface markers. A complete list of biomarkers used for RT-qPCR analysis is given in Supplementary Figure S6.
Expression of mRNA biomarkers for oxidative stress
Relative mRNA expression levels of genes associated with oxidative stress as determined by RT-qPCR are given in Figure 7. Distinct differences were observed among cells cultured on each orthopedic implant substrate type. Of note, GPX1 expression was elevated at early time points (days 0 and 3) in cells cultured on gb-Ti, whereas cells cultured on ps-Ti had higher expression levels of GPX4 at day 0 (Fig. 7; top row).
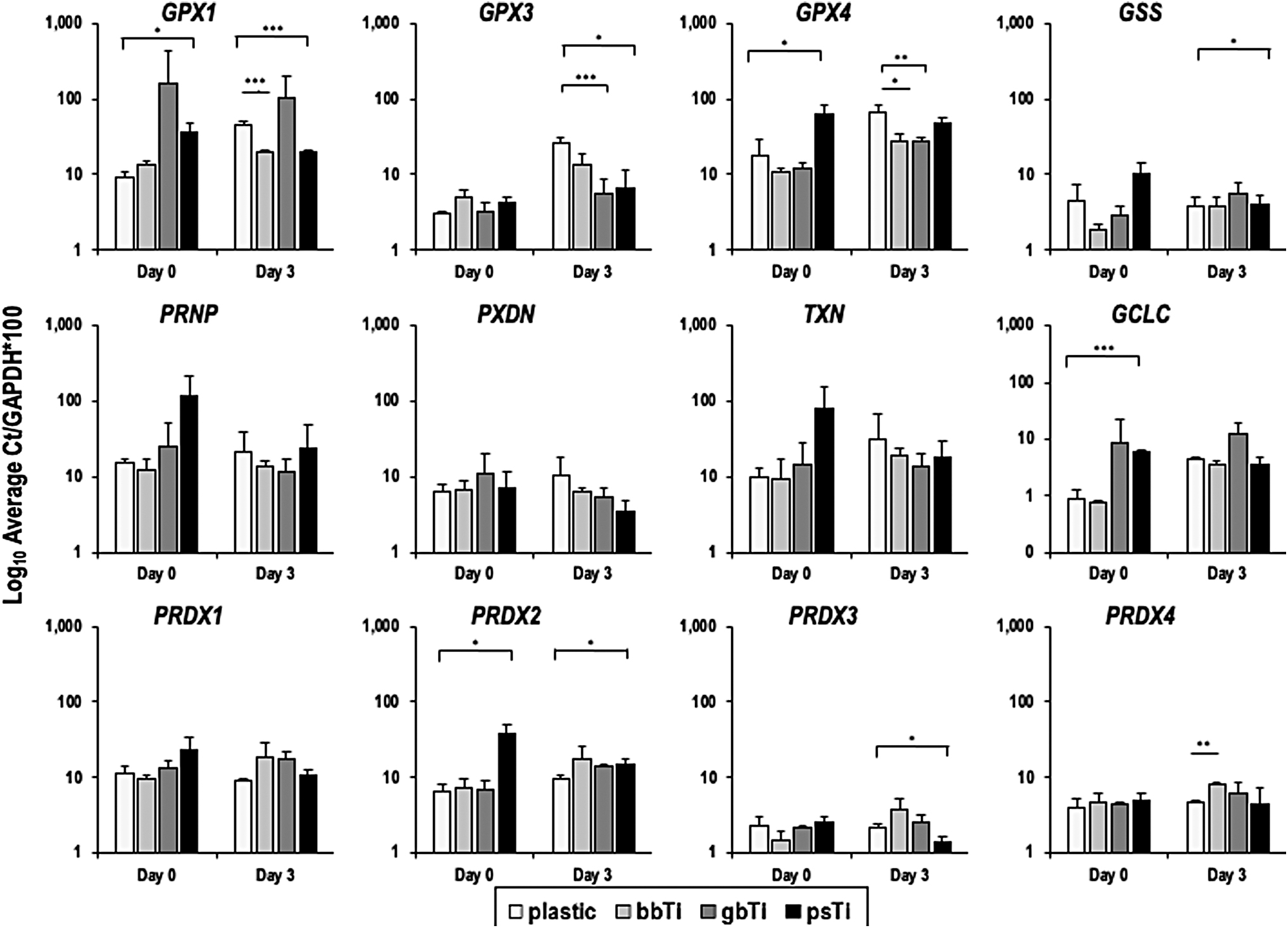
Select indicators of oxidative stress were prioritized for RT-qPCR analysis by substrate type at early time points (days 0 vs. 3) to directly compare early responses of cells with materials with different levels of surface rugosity. Log10 Average Ct/GAPDH*100 calculations were used to normalize data and facilitate comparisons among gene expression values. Cell culture experiments were performed in triplicate, allowing calculation of mean and SD estimates, as presented. Statistically significant differences in expression of genes from cells grown on different substrates were individually analyzed using a Student's t-test using a two-tailed distribution with two sample equal variance (*p < 0.05, **p < 0.01, ***p < 0.001).
Similarly, GSS, PRNP, and TXN were elevated in cells cultured on ps-Ti at day 0, but PXDN was reduced in cells cultured on ps-Ti after 3 days (Fig. 7; middle row). Glutamate-cysteine ligase (GLCL), a key enzyme in the synthesis of glutathione and a marker of oxidative stress, was highly expressed in cells cultured on rougher substrates (gb-Ti and ps-Ti) at day 0, but normalized as cells reached confluence (Fig. 7; middle row). PRDX1 was constitutively expressed among all treatment groups, yet peroxiredoxin 2 (PRDX2) was elevated in cells cultured on ps-Ti compared with cells cultured on all other substrates at day 0: a pattern that normalized by day 3 (Fig. 7; bottom row). Future experiments aimed at quantifying the amounts and types of ROS present in each treatment group will be informative toward interpreting the oxidative response of cells to differing levels of ROS.
Expression of enzymes that support ECM remodeling
Members of the ADAM family of secreted metalloproteinases, which support ECM remodeling but are also indicative of cellular responses to heavy metal stress, were expressed similarly among cells on all surfaces at all time points (Fig. 8; top row). Matrix metalloproteinase 1 (MMP1) showed elevated gene expression at day 0 on ps-Ti and among all surfaces at day 3 in comparison with plastic-cultured cells (Fig. 8; middle row).
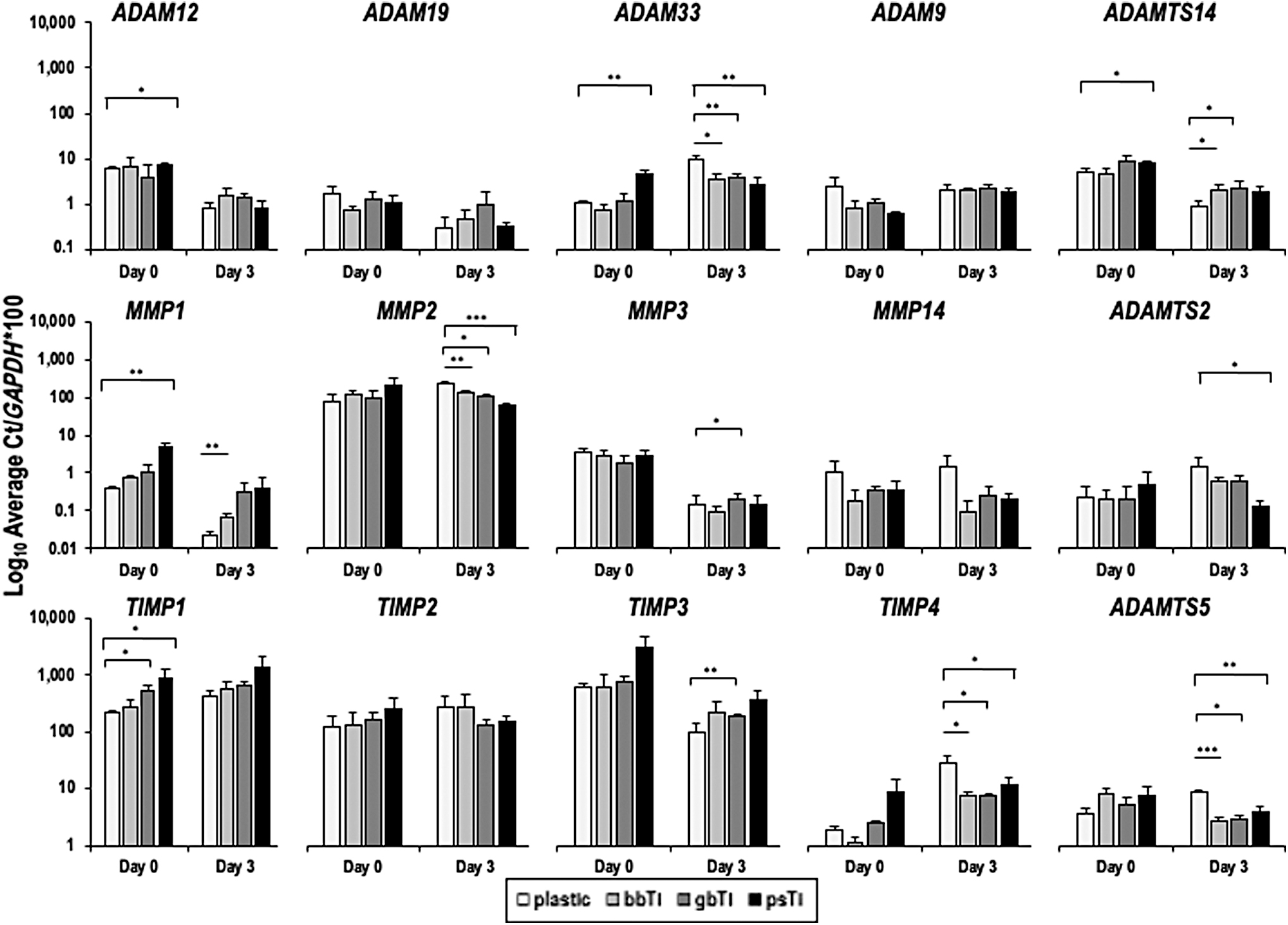
RT-qPCR data (Log10 Average Ct/GAPDH*100) pertaining expression levels of HMR genes (e.g., ADAMs, MMPs, and TIMPs) were compared among cells cultured on each substrate type at early time points (days 0 vs. 3). Cell culture experiments were performed in triplicate, and mean (±SD) data are presented. Statistically significant differences in expression of genes from cells grown on different substrates were individually analyzed using a Student's t-test with two-tailed distribution and two-sample equal variance (*p < 0.05, **p < 0.01, ***p < 0.001).
In contrast, MMP14 was marginally reduced (low expression values) in cells cultured on all three Ti substrates. MMP2 and MMP3 were not differentially expressed among the experimental groups. Representatives of the metalloproteinase inhibitors (e.g., TIMP1, TIMP3, and TIMP4) showed variation in gene expression among substrates, with the elevated gene expression on ps-Ti at day 0 (Fig. 8; bottom row). Differences in the expression of key metalloproteinases between cells cultured on each substrate type suggest that changes in the local biochemistry of each titanium material may elicit an important ECM remodeling response, whereby the additional surface area available in ps-Ti substrates may be detrimental to long-term ECM homeostasis within attached cells.
Discussion
Metal orthopedic implant materials can be 3D printed to customize musculoskeletal joint performance. Changes to the material properties of orthopedic implants, including surface roughness characteristics, may modulate the behavior of cells at the bone-implant interface. At present, modular orthopedic devices use interconnected implant materials that often differ in surface roughness. However, the biological consequences of this strategy on endogenous musculoskeletal repair cells are unknown. Characterization of the biological response of cells and tissues around these orthopedic implants may help our understanding of the mechanisms behind common and debilitating postoperative complications, specifically aseptic implant loosening.
This study focused on the biological responses of two types of human MSCs (adipose and bone derived) to three different types of orthopedic implant materials. The bb-Ti, gb-Ti, and ps-Ti disks were first characterized for surface roughness and used as in vitro models of the bone-implant interface. MSCs derived from the vascular fraction of adipose tissues and fibroblastic cells derived from bone tissues were used to represent resident peri-implant cells that respond to the implantation of metal into bone. We examined the ultrastructural properties of Ti surfaces using SEM, whereas cultured cells in contact with Ti surfaces were interrogated for differential gene expression by RT-qPCR and RNA-seq.
Well-characterized biomarkers of ECM formation (e.g., COL1A1), cell proliferation (e.g., HIST2H4), and cell surface signaling (e.g., CD44) were analyzed. Transcription levels of genes that are sensitive to ROS (e.g., glutathione peroxidases) and heavy metal accumulation (e.g., metallothioneins) were specifically investigated. High-throughput RT-qPCR methods were used to identify and highlight differential gene expression differences among cells cultured on the three substrate types analyzed in this study. Our results showed key differences in ECM deposition, cell proliferation, and lineage commitment, consistent with our previous observations using cultured human cells and surgical joint capsular tissue specimens obtained during revision surgery.25,44,45 Cells cultured on the roughest substrates showed reduced mRNA expression of key ECM markers, specifically COL1A1 and COL3A1, compared with cells cultured on smoother substrates. This difference was most pronounced at earlier time points, suggesting that cells exposed to titanium materials had delayed production of collagenous and noncollagenous proteins required for ECM formation, which may be a key process in orthopedic implant fixation. Tissue culture plastic is specifically designed to favor the adhesion of cells in culture, so elevated mRNA expression of ECM markers may be owing to inherent engineering differences between plastic and titanium (e.g., 3D printing strategies).
Cells grown on rougher substrates also took longer to reach confluence than cells grown on smooth substrates. The common biomarker for cell proliferation, HIST2H4, was elevated early on ps-Ti, suggesting these cells reached confluence later than cells grown on smoother substrates. This could be because of the increased surface area, allowing the cells more surface area to proliferate before becoming contact inhibited when cells form a contiguous layer over the Ti substrate (i.e., cell confluence).
Alternatively, the reduced rate of cell proliferation on rougher substrates could result from an adverse biological reaction to the increased exposure to titanium. Ongoing studies have shown that titanium nanoparticles induce adverse reactions in vitro, which could account for delayed cell proliferation (unpublished data). If the increased exposure to titanium slows cell proliferation and delays confluence, this could have a deleterious effect on the ability of host tissues to rapidly incorporate orthopedic components into bone.
A delayed lineage commitment of MSCs was observed on rougher titanium substrates. Cells grown on ps-Ti showed higher levels of CD248 and CD44 expression at early time points. These are key stem cell biomarkers, and their increased expression at early time points suggests that cells grown on rough titanium make take longer to differentiate. A delay in stem cell differentiation in vivo could delay cell proliferation and ECM deposition, ultimately causing a delay in orthopedic implant osseointegration.
However, it is important to note that concurrent alterations in cell cycle and stem cell markers may obfuscate attempts to identify the principle component responding to surface rugosity (e.g., the expression levels of cell cycle markers may need to reach a biological threshold before modifying stem cell markers). Additional experiments aimed at identifying the interplay between classes of biomarkers would be necessary to better identify the specific effects of metal exposure, ROS exposure, and/or differences in rugosity.
Distinct differences were observed in the expression of markers for oxidative and heavy metal stress between cells grown on rough and smooth substrates. Common markers for oxidative stress, including mRNAs encoding enzymes like glutathione peroxidases, thioredoxins, and GLCLs, were generally upregulated at early time points on rougher titanium substrates. These findings are consistent with our earlier studies that showed exposure to titanium induces oxidative stress in cultured cells. 25 Of interest, in vivo experiments using antioxidant-treated Ti implants have shown improved levels of osseointegration and clinical performance.22,46–48 Those findings may be owing to the biological response observed in this study, supporting that titanium induces an oxidative stress response in peri-implant tissue, potentially impairing the osseointegration of titanium orthopedic implants. Of interest, titanium biomaterials are known to produce a thin layer of TiO2 when exposed to air because of its high affinity for oxygen, which can affect cellular processes for a variety of cell types (e.g., Ziental et al., 49 Setyawati et al., 50 and Setyawati et al. 51 ).
Identifying nanostructural differences in the formation of TiO2 among biomaterials considered here (e.g., ps-Ti should produce more because of greater surface areas exposed to air) would aid in the development of future strategies to limit the potential consequences (e.g., oxidative stress and limited cellular adhesion) of cellular exposure to this layer. Additional data detailing the early adhesion and proliferation of multipotent progenitor cells to Ti biomaterials in vivo will aid in determining whether bb-Ti, gb-Ti, and ps-Ti or other surface modifications will increase implant fixation.
An understanding of specific cellular recruitment processes and selective microenvironmental suitability of different cell types in response to each implant material (using animal models) would also inform translational efforts to bolster implant fixation and assist in identifying candidate cell populations that best facilitate osseointegration.
Markers for heavy metal stress were elevated in MSCs grown on rougher substrates. Specifically, mRNAs for secreted metalloproteinases (e.g., ADAM33, MMP1, and TIMP3) showed elevated expression on cells grown on ps-Ti at early time points. These findings are somewhat intuitive, as titanium is a heavy metal. Yet, the prevailing orthopedic concept holds that titanium is metabolically inert and nontoxic to surrounding cells in vivo.7,9,10 The findings in this study challenge this concept and raise the possibility that the increased surface area of rough titanium implants induces an oxidative and heavy metal stress response in surrounding tissues, thus potentially negatively impacting osseointegration and long-term implant survival.
There are several limitations of this study that restrict a broader interpretation of our data at present. First, adipose-derived stem cells from only two human patients and bones samples from three patients were used in these experiments. Hence, the potential for small sample size effects reduces the extrapolation potential of these results. Using a standardized human stem cell line or increasing the number of patients sampled would reduce this variation. Yet, the stem cells used in these experiments have well-characterized properties previously described.
A second limitation of this study is the absence of measured ROS in the culture media. A quantitative analysis of ROS within the media would help confirm the observed responses of MSCs to rough titanium substrates. Similarly, quantifying trace amounts of heavy metals that erode from the materials and diffuse into the media would be important to analyze, although measurement protocols rarely include trace amounts of titanium in culture media. 21 Finally, including positive control substrates that are known to cause toxic responses to cells, such as cobalt–chrome alloys, would contextualize the potential stress response of cells to titanium.
In conclusion, the surface roughness of metal orthopedic implants can change the phenotype of MSCs. Increased surface roughness of orthopedic implants may delay the differentiation and ECM formation of MSCs and alter the expression of genes sensitive to representative ROS and protein kinases. These findings challenge the prevailing orthopedic dogma that titanium alloys are biologically inert in patients.7,9,10 Furthermore, the effect of titanium on peri-implant tissues may account at least in part for poor implant osseointegration, and impaired long-term implant survival. Future work should aim to directly measure ROS in the media after cells have been cultured on each substrate type. In addition, by expanding the scope of this study to include additional cell types (e.g., osteoclasts and chondrocytes) and patient donors (e.g., patients with diabetes), we will examine patient-specific differences in the response of cells to different material types. To facilitate a clinically translatable system for testing our in vitro observations, it will be interesting to use radioisotope- and fluorescent-labeled cells to track the location and behavior of cells in contact with the surface interface of each material in vivo.
Footnotes
Acknowledgments
The authors thank the members of our laboratories and affiliated investigators, including Christopher Salib, Christopher Paradise, Endre Søreide, Hilal Kremers, Matthew Getzlaf, Dakota Jones, and Darcie Radel for stimulating discussions, as well as sharing their expertise in research design, experimental execution, and data interpretation. The authors are also grateful to Matthew Hainy (Engineering Department) for assistance with surface roughness measurements of each material, and Scott Gamb (Electron Microscopy Core Facility) for assistance with electron microscopy. For production of the biomaterials, the authors thank Brent Mitchell, Dale Swarts, and Mark Gruczynski from StrykerMako.
Disclosure Statement
D.J.B. reports personal fees from Journal of Bone and Joint Surgery, DePuy, Wolter Kluwer, and Elsevier. In addition, D.J.B. is a President-Elect on the International Hip Society, and a member on the International Society of Arthroplasty Registries. D.J.B. also reports Consulting for Bodycad, and is a Member on the Board of Trustees for the Orthopaedic Research and Education Foundation, and is a Senior Director for Current Concepts in Joint Replacement. Finally, D.J.B. has a patent DePuy issued. R.C.C. serves as the President and General Manager of Reconstructive Research and Development at Stryker Orthopedics. D.G.L. reports personal fees and other from Zimmer Biomet, and Mako/Stryker, Corin USA, Acuitive Technologies, and Ketai Medical Devices. In addition, D.G.L. has patents on selected hip and knee implants with royalties paid by Zimmer, and is employed part time as the Medical Director for The American Joint Replacement Registry and Orthopaedic Research and Education Foundation. M.P.A. reports personal fees from Stryker, and M.P.A. is on the American Academy of Orthopedic Surgeons Board of Directors. No other authors have conflicts of interest to disclose.
Funding Information
Financial support was provided by the National Institute of Arthritis and Musculoskeletal and Skin Diseases (R01 AR049069 to A.J.v.W; T32 AR056950 and F32 AR068154 to E.A.L., and F32 AR066508 to A.D.), as well as generous philanthropic support from William H. and Karen J. Eby. In addition, this study received support from the Mayo Clinic Center for Regenerative Medicine (to R.T.).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.