
Abstract
Immune-related applications of mesenchymal stromal cells (MSCs) in cell therapy seek to exploit immunomodulatory paracrine signaling pathways to reduce inflammation. A key MSC therapeutic challenge is reducing patient outcome variabilities attributed to insufficient engraftment/retention of injected heterogenous MSCs. To address this, we propose directly transplantable human single-cell-derived clonal bone marrow MSC (hcBMSC) sheets. Cell sheet technology is a scaffold-free tissue engineering strategy enabling scalable production of highly engraftable cell constructs retaining endogenous cell–cell and cell–matrix interactions, important to cell function. cBMSCs, as unique MSC subset populations, facilitate rational selection of therapeutically relevant MSC clones from donors. Here, we combine human cBMSCs with cell sheet technology, demonstrating cell sheet fabrication as a method to significantly upregulate expression of immunomodulatory molecules interleukin (IL)-10, indoleamine 2,3-dioxygenase (IDO-1), and prostaglandin E synthase 2 (PTGES2) across GMP-grade hcBMSC lines and whole human bone marrow-derived MSCs compared to respective conventional cell suspensions. When treated with carbenoxolone, a gap junction inhibitor, cell sheets downregulate IL-10 and IDO-1 expression, implicating functional roles for intercellular sheet interactions. Beyond producing directly transferable multicellular hcBMSC constructs, cell sheet technology amplifies hcBMSC expression of immunomodulatory factors important to therapeutic action. In addition, this work demonstrates the importance of cell–cell interactions as a tissue engineering design criterion to enhance consistent MSC functions.
Impact statement
Mesenchymal stem/stromal cell (MSC) production of anti-inflammatory factors has motivated significant clinical interest in their therapeutic utility for immune-related diseases. Strategies that enhance the potency of culture-expanded MSCs and improve MSC engraftment hold promise to advance MSC therapy. Cell sheet technology is a tissue engineering approach enabling direct cell transplantation and localized tissue engraftment. We report the first mechanistic insights into functional roles of gap junctions within MSC sheets and demonstrate that cell sheet fabrication stimulates MSC expression of immunomodulatory factors compared to conventional MSC suspensions. These findings motivate further studies of MSC sheets in diverse immune-related applications and therapies.
Introduction
Mesenchymal stem/stromal cells (MSCs) have attracted significant clinical interest as a therapeutic candidate for treating immune-related diseases1–3 motivated by MSC secretion of soluble anti-inflammatory factors (i.e., cytokines, growth factors, chemokines, exosomes).4,5 For instance, MSC-production of immune-related factors interleukin (IL)-10, prostaglandin E synthase 2 (PGE2), and indoleamine 2,3-dioxygenase (IDO)-1 are reported to modulate T cell proliferation, differentiation, and macrophage polarization toward an anti-inflammatory phenotype.6,7 Growing evidence supports the MSC primary therapeutic mechanism of action driven by this dynamic production of paracrine factors. 7 Therefore, strategies that enhance culture-expanded MSC production of therapeutic immunomodulatory paracrine factors and facilitate MSC engraftment beyond that of conventional MSC suspension injections hold promise to advance MSC therapy for localized immune-related diseases. 4 To this end, promoting three-dimensional culture and tissue-like intercellular interactions is proposed to increase MSC expression of immune-related paracrine factors.8–11
However, these three-dimensional (3D) strategies predominately use directly transplantable biomaterial scaffolds intrinsically limited by irregular cell distribution and cell densities, imposed transport barriers, and spatially restricted cell–cell signaling interactions.8,11 Therefore, to advance MSC therapeutic capacity, we propose cell sheet tissue engineering, 12 a scaffold-free approach using nonenzymatic temperature-mediated cell detachment to harvest MSCs as robust, directly transplantable contiguous 3D sheets. MSC sheets exhibit consistently high cell densities and matrix production, with limited diffusion barriers due to disc-like geometries, and unrestricted cell–cell signaling interactions. 13
Preclinical studies establish MSC sheets as a strong therapeutic candidate in applications of kidney disease,14–16 skin ulcers, 17 uterine fibrosis, 18 and heart disease.19,20 Significantly, direct cell sheet transplantation improves long-term MSC localized engraftment, efficiency, and retention duration compared to conventional cell suspension injections. 15 Our group has recently introduced cell sheet fabrication as a tissue engineering strategy capable of stimulating MSC expression of paracrine factors via a “tissue-effect” attributed to 3D cell sheet dimensionality and multidirectional cell–cell interactions.13,21,22 In contrast to conventional cell suspensions, cell sheets preserve instructive intercellular signaling interactions, such as gap junctions.13,21,22
Intercellular gap junctions are cytoplasmic communication channels controlling bidirectional diffusion of small molecules, such as ions and metabolites, between adjacent cells, thereby regulating cellular physiological activities. 23 Gap junctions regulate many physiological events, including inflammation,24,25 differentiation, 26 and proliferation. 27 MSCs constitutively express gap junction subunits connexin-43 (Cx43), connexin-40 (Cx40), and connexin-45 (Cx45), of which Cx43 exhibits primary expression. 28 During culture expansion, neighboring MSCs form intercellular coupled gap junctions to facilitate molecular transfer by connecting transmembrane homomeric and heteromeric connexin hexamers, called connexons.29–31 MSCs are known to pass mitochondrial information via gap junctions.32,33 Furthermore, Cx43 upregulation is reported to promote the expression of proregenerative angiogenic paracrine factor vascular endothelial growth factor and improve MSC cardioprotective functions.34,35
However, during production of conventional MSC suspensions, enzymatic detachment from culture cleaves cell–cell and cell–matrix interactions disrupt gap junction intercellular signaling pathways. In contrast, nonenzymatic, temperature-mediated cell sheet detachment allows harvest from culture while preserving and promoting intercellular signaling junctions, including gap junctions.13,22 For example, cardiomyocyte sheets retain intercellular gap junctions and uncoupled connexons on both apical and basal sides of the cell sheet, enabling overlaid sheets to rapidly form gap junctions capable of transmitting molecular and electrical signals.36,37 However, the distinct role of gap junctions in stimulating MSC expression of immunomodulatory factors, particularly within cell sheets, remains unknown.
In this study, we used human bone marrow-derived MSCs (hBMSCs) from four donors, one conventionally derived and, thus, retaining innate BMSC subpopulation heterogeneity (whole hBMSCs), 38 and three clinical-grade clonal (single-cell-colony-derived) MSC populations (clonal hBMSCs). 39 The advent of clonal diversity and profiling of intrinsic MSC heterogeneity within the bounds of the defined MSC phenotype complicate the ability to reliably expand, bank, and produce an MSC end-product with consistent critical quality attributes, defined potency, and reproducible therapeutic efficacy.4,40 Human single-cell-derived clonal bone marrow MSCs (hcBMSCs) used in this study represent clinical-grade homogenous MSC subset populations capable of long-term expansion, banking, and therapeutic effects at late passages (passage 10) with low immunogenicity. 39 To account for MSC heterogeneity, this study uses different subsets of clonal MSCs to determine if observed effects from cell sheet fabrication are uniform or cell line-dependent.
This study aimed to examine the influence of cell sheet fabrication and intercellular gap junctions on hBMSC expression of factors implicated in immunosuppression to ultimately advance hBMSC therapy for localized immune-related diseases. We hypothesized that preservation of cellular interactions facilitated by cell sheet detachment would lead to upregulated expression of immunomodulatory factors in hBMSCs. To address this hypothesis, we first evaluated the expression of immunomodulatory molecules in hBMSC sheets compared to conventional hBMSC suspensions with enzymatically cleaved cell–cell interactions. A follow-up functional blocking experiment revealed a critical role for gap junction intercellular communication (GJIC) within hBMSC sheets paracrine factor expression. Together, experimental results reflect the importance of cell–cell interactions in promoting consistent MSC expression of therapeutic factors.
Materials and Methods
hBMSC isolation and culture
Whole hBMSCs were isolated from a whole-blood bone marrow aspirate sample purchased from Lonza (donor: 30 years, male) by selecting adherent cells. Whole hBMSCs were expanded into a master cell bank at passage 2 and a working cell bank at passage 8 and used for experimentation at passage 10. Human clonal hBMSCs were provided by SCM Lifesciences (Incheon, Republic of Korea) at passages 5 and 6, selected for their known ability to retain expected phenotypic and functional MSC markers, and expand into a large-scale clinical cell bank. A single banked clonal hBMSC, obtained from bone marrow aspirates using subfractionation culture method, patented by SCM Lifesciences, and confirmed to exhibit the expected MSC marker profiles and immunoreactive potencies up to passage 1239,41,42 was used in all studies.
All hBMSCs were confirmed positive for MSC markers CD44, CD73, CD105, and CD90, negative for CD45, CD34, and human leukocyte antigen-DR, using flow cytometry and express multipotent capabilities (Supplementary Fig. S1). All hBMSCs were plated at 1000–2000 cells/cm2 in growth media containing Low Glucose (1 g/L) Dulbecco's modified Eagle's medium (Gibco, Waltham, USA) supplemented with 10% fetal bovine serum (Thermo Fisher Scientific, Waltham), 1% penicillin-streptomycin (P/S) (Gibco), and 0.5 μL/mL MycoZap Prophylactic (Lonza, Morristown) and incubated in a humidified environment (37°C, 5% CO2). Media was changed after 24 h of initiating culture and every 2 days subsequently.
Whole and clonal hBMSC sheet and suspension fabrication
At passage 9, all hBMSCs were passaged using 0.05% Trypsin–EDTA (Gibco), and cell suspensions were counted using a hemocytometer. The resultant passage 10 hBMSCs were seeded onto 35 mm UpCell™ thermoresponsive culture dishes (TRCDs) (Cellseed, Tokyo, Japan) at 4 × 105 (∼41,666 cells/cm2) per cell sheet and cultured in complete growth media supplemented with 50 μg/mL l-ascorbic acid 2-phosphate (Wako, Osaka, Japan). Simultaneously, all hBMSCs were seeded onto tissue culture plastic (TCP) (CellTreat, Pepperell) at a conventional seeding density (5000 cells/cm2) for MSC suspension preparation. 43 In each group, MSCs were cultured for 6 days with one media change on day 4. At 6 days, MSC sheets were moved to 20°C for 20 min, then detached with forceps as 3D cell sheets.
Meanwhile, MSC suspension samples were detached from TCP using 0.05% Trypsin-EDTA (Gibco) for 5 min, followed by neutralization with culture media and cell collection via centrifugation. To inhibit gap junctions within MSC sheets, 100 μM carbenoxolone (CBX) (Sigma-Aldrich, St. Louis) was added to the media on day 0 and 4 of MSC sheet culture. Top-down microscopic images of each group before detachment were obtained using phase contrast microscopy (AX10 microscope; Carl Zeiss Microimaging, Göttingen, Germany). Top-down macroscopic images of MSC sheets were obtained at 0 h, immediately after cultureware detachment.
Histological analysis
Immediately following detachment, MSC sheets were fixed with 4% paraformaldehyde (PFA) (Thermo Scientific) for 20 min and paraffin-embedded (ARUP Laboratories, Salt Lake City). Embedded samples were sectioned at 4 μm. To identify cell sheet structure and cell morphology, Hematoxylin (Sigma-Aldrich) and Eosin (Thermo Scientific) (H&E) staining was conducted. Stained sections were imaged with a BX 41 Widefield Microscope (Olympus, Japan) using AmScope Software (v4.8.15934) using AmScope 18MP Aptina Color CMOS.
Immunofluorescent staining
To visualize connexin-43 in hBMSCs sheets, 0 h cell sheets were fixed with 4.0% PFA for at least 30 min and embedded in paraffin (ARUP Laboratories). Paraffin-embedded cell sheets were sectioned at 4 μm, followed by deparaffinization. Samples were then permeabilized with 0.1–0.3% Triton X-100 (Sigma-Aldrich) in PBS for 10 min and blocked with 5% goat serum in PBST for 1 h, followed by overnight incubation with primary anti-connexin-43 (Ab 11370; Abcam, Cambridge, United Kingdom) at 4°C. Following primary antibody incubation, samples were washed with PBS and incubated with secondary biotin antibody (Abcam) for 1 h, followed by incubation with AF488 Streptavidin (Abcam) for 1 h.
All samples were counterstained with DAPI nuclear stain (Life Technologies, Carlsbad) (2 drops/mL at RT for 5 min) and mounted using Prolong Gold DAPI Mounting Solution (Life Technologies). Microscopic images were obtained using fluorescence microscopy (AX10 microscope; Carl Zeiss Microimaging). Fluorescent area of connexin-43 was determined by normalizing the fluorescence intensity, analyzed via image thresholding (MATLAB) (MathWorks, Natick) by the number of observable cells, analyzed manually by DAPI staining. Ten images per sample were used for quantification.
Quantitative real-time PCR analysis
Total RNA was isolated from hBMSCs using the Qiagen RNeasy Mini Kit (Qiagen, Hilden, Germany) according to manufacturer's instructions immediately following detachment to prepare both MSC suspension and MSC sheet samples. Isolated RNA was quantified with a NanoDrop Spectrophotometer (Thermo Scientific). All cDNA samples were prepared from 1.0 μg of RNA/sample using a high-capacity cDNA Reverse Transcription Kit (Life Technologies). Genes were quantified by real-time PCR using Applied Biosystems primers glyceraldehyde 3-phosphate dehydrogenase [GAPDH, Hs99999905_m1] as a housekeeping gene, interleukin-10 [Il10, Hs00961622_m1], indoleamine 2,3-dioxygenase [IDO1, Hs00984148_m1], prostaglandin E synthase 2 [PTGES2, Hs00228159_m1], N-cadherin [CDH2, Hs00983056_m1], and connexin 43 [GJA1, Hs04259536_g1] performed on Applied Biosystems Step One Plus (Applied Biosystems, Waltham).
Gene expression was normalized to GAPDH expression levels. For MSC sheet to MSC suspension comparisons, relative gene expression was determined using the comparative threshold cycle change algorithm normalized to respective cell suspensions. For gap junction inhibition studies, expression levels are relative to nontreated (control) MSC sheets.
Statistical analysis
All statistical analyses were conducted on n ≥ 3 replicate data sets. Quantitative values are expressed as mean ± SE. Shapiro–Wilk test was used to determine normality of data sets. If data were normally distributed, a one-way ANOVA with Tukey's post hoc test was used. If normality could not be confirmed, the Kruskal–Wallis test was used. Analysis was performed using the Origin(Pro) Version 2022 software (OriginLab Corporation, Northampton). Statistically significant differences were defined as *p < 0.05, **p < 0.01, and ***p < 0.001. No statistical significant difference was defined as p > 0.05.
Experiments
Whole and clonal hBMSCs readily detach as 3D contiguous cell sheets with upregulated cell–cell interactions
To evaluate differences in hBMSC sheets compared to conventional hBMSC suspensions, we simultaneously fabricated MSC sheets and conventional MSC suspensions (Fig. 1). Whole and clonal hBMSCs were prepared as a conventional cell suspensions by seeding hBMSCs onto conventional TCP and culturing for 6 days, followed by enzymatic detachment (0.05% Trypsin-EDTA, 5 min) (Fig. 1). Simultaneously, the same hBMSCs were prepared as cell sheets by seeding hBMSCs onto TRCDs and culturing for the same 6-day period, followed by temperature-mediated sheet detachment by reducing culture temperature from 37°C to 20°C, prompting release of adherent cells under tension (Fig. 1), and resulting in a contracted, detached 3D cell sheet construct (Fig. 2a).
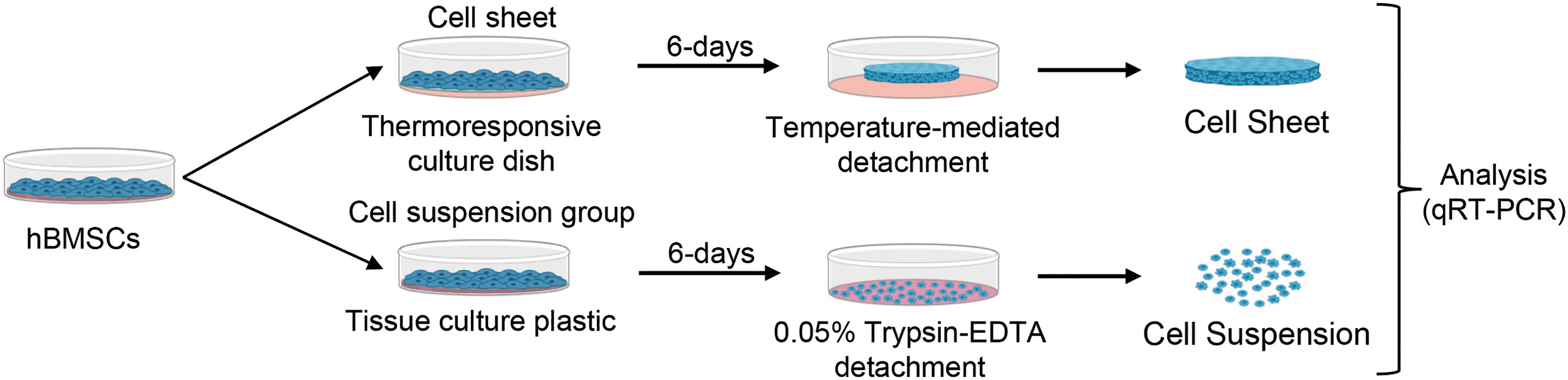
Experimental schematic. hBMSCs from the same passage are either prepared for MSC sheet or suspension cultureware detachment. Immediately following each respective detachment method, samples are prepared for gene expression analysis using qRT-PCR. hBMSCs, human bone marrow-derived MSCs; MSCs, mesenchymal stromal cells; qRT-PCR, quantified by real-time PCR. Color images are available online.
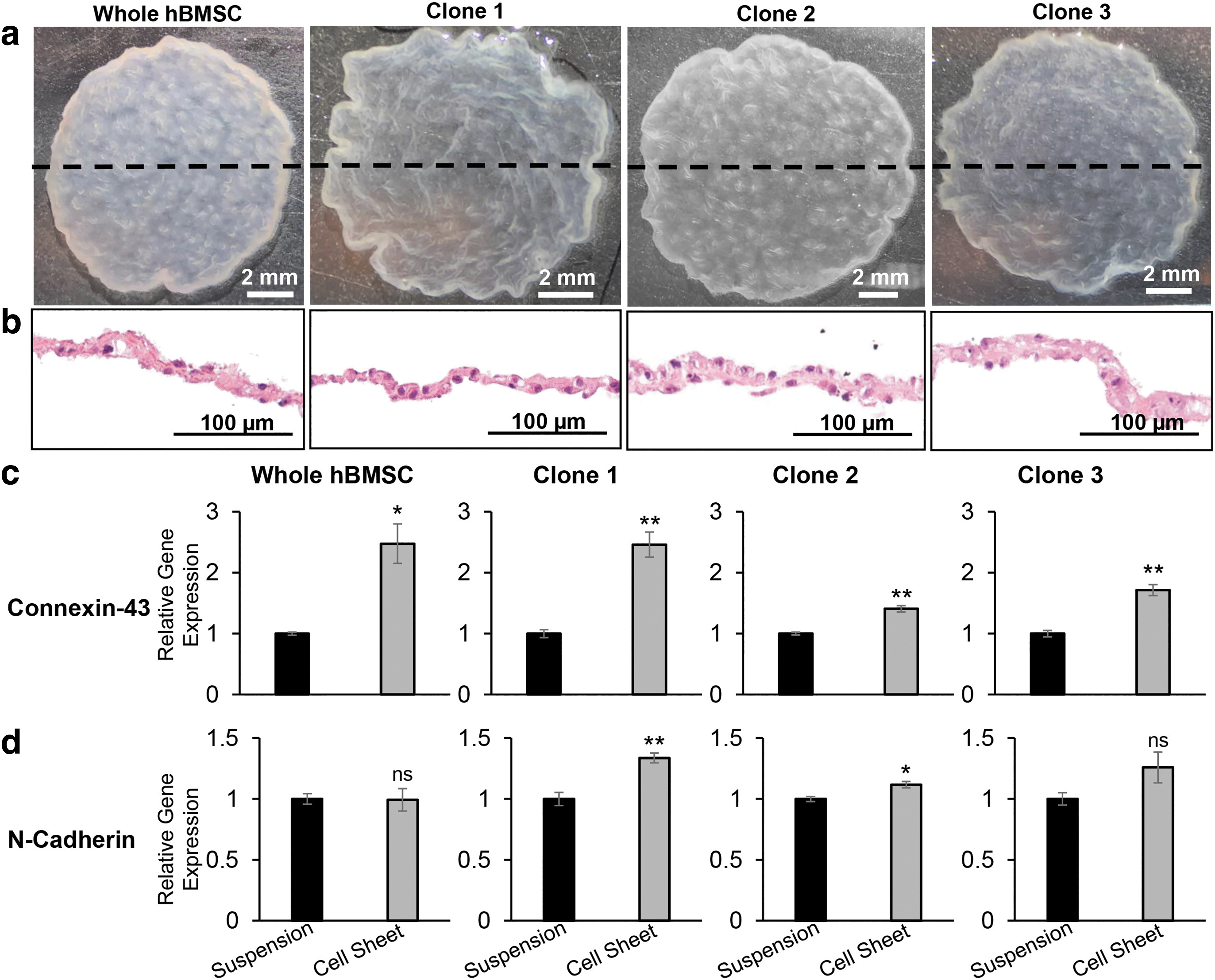
Cell sheets exhibit upregulated expression of genes related to cell–cell interactions.
Resultant hBMSC sheets were contiguous 3D constructs retaining cell contact between adjacent cells, as observed macroscopically and microscopically via H&E cross-sectional staining (Fig. 2a, b). Comparisons of cell number revealed a consistent cell density of 2.77 ± 0.44 × 104 cells/cm2 across the final hBMSC sheet constructs and 1.17 ± 0.38 × 104 cells/cm2 across the cell suspensions. To determine the influence of cell sheet fabrication on expression of intercellular signaling molecules, we measured gene expression of gap junction molecule Cx43 and adherens junction molecule N-cadherin (Fig. 2c, d). hBMSC sheets exhibited upregulated expression of Cx43 (1.4–2.5-fold change) and N-cadherin (1–1.3-fold change), compared to respective conventional cell suspensions harvested via enzymatic detachment (Fig. 2c, d). Cx43 expression is significantly upregulated in all hBMSC sheets compared to their respective cell suspensions, attributed to increased gap junction formation.
These data demonstrate robust fabrication of 3D hBMSC sheets across clonal hBMSCs lines and whole hBMSCs, promoting cell–cell interactions and upregulated expression of intercellular gap junction molecule Cx43.
hBMSCs upregulate gene expression of immunomodulatory genes as cell sheets
Previous studies demonstrate promotion of MSC three-dimensionality and cell–cell interactions as a way to stimulate immunomodulatory molecule expression in MSCs.9–11,21 We, therefore, sought to evaluate the influence of cell sheet fabrication on hBMSC expression of IL-10, IDO-1, and PGE2, all immunomodulatory paracrine factors highly implicated in MSC-mediated immunomodulation. 7 We initially validated the expression of these therapeutic factors within MSC sheets (Supplementary Fig. S2), then compared the expression of IL-10, IDO-1, and PTGES2 between hBMSC sheets, with promoted intercellular interactions (Fig. 1), and respective hBMSC conventional cell suspensions. hBMSCs as cell sheets exhibited significant upregulation of IL-10, IDO-1, and PTGES2 compared to conventional cell suspensions (Fig. 3a–c). Clone 1 hBMSCs exhibited the most stimulation by cell sheet fabrication with a 32.4-fold change of IL-10 and 15.4-fold change of IDO-1 expression (Fig. 3b, c).
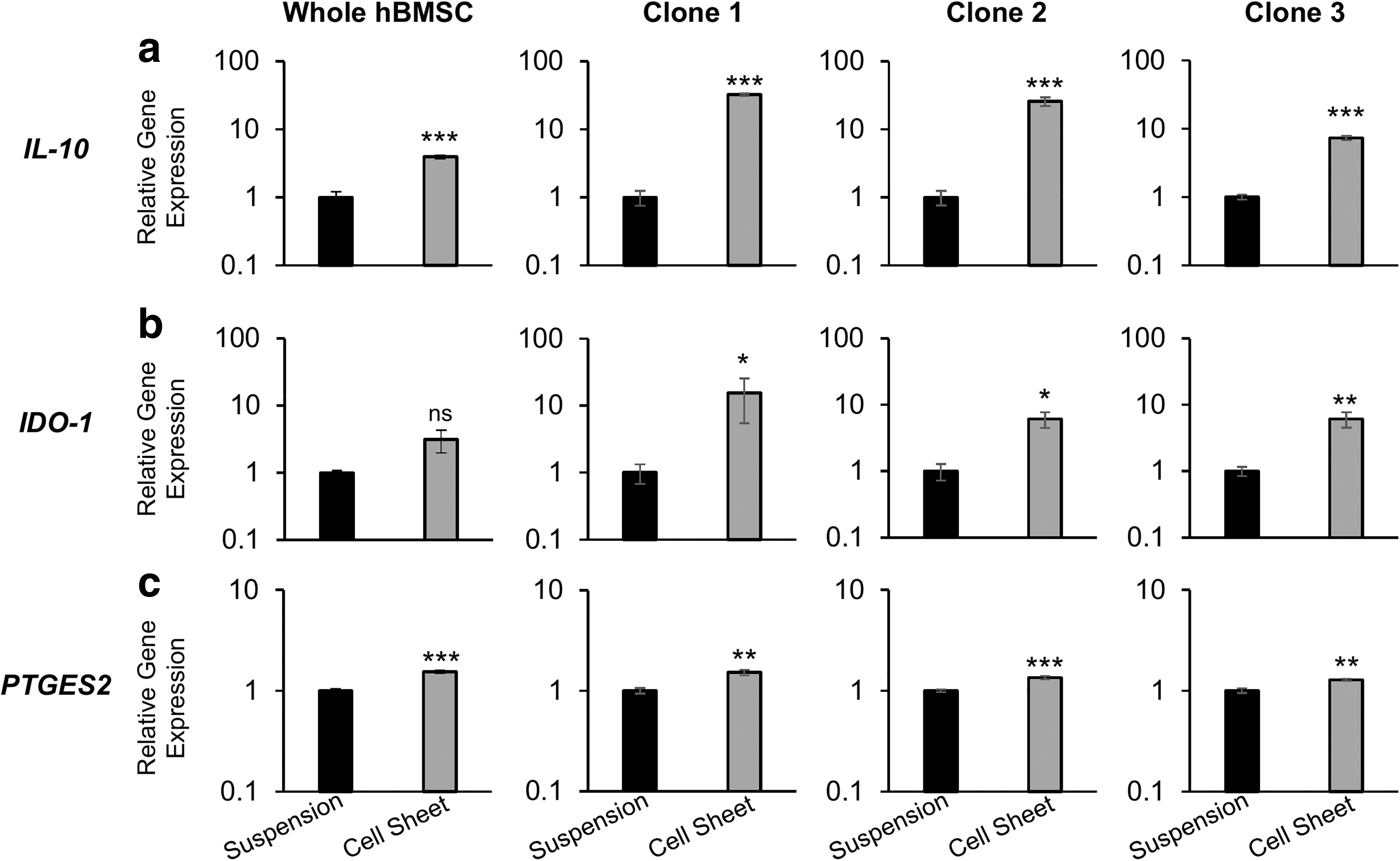
MSC sheet fabrication upregulates gene expression of immunomodulatory molecules in hBMSC sheets compared to respective conventional cell suspensions. qRT-PCR gene expression for immunosuppressive genes
Whole hBMSCs exhibited the least response to cell sheet fabrication with a lesser 3.9-fold change of IL-10 and a 3.1-fold change of IDO-1 (Fig. 3b, c). Fold-changes of PTGES2 expression were similar across all groups ranging from 1.5 to 1.3. Notably, all clonal and whole hBMSCs responded positively to cell sheet fabrication as determined by successful upregulation of immunomodulatory molecules, indicating that the effect is not confined to specific hBMSC subtypes. These data support use of cell sheet technology as a feasible tissue engineering strategy to enhance expression of immunomodulatory factors in culture-expanded MSCs compared to conventional cell suspensions.
Blockade of gap junctions within hBMSC sheets
Unique upregulation of intercellular gap junctions in hBMSCs sheets allows for information sharing between adjacent cells throughout the cell sheet construct. 33 To evaluate the functional role of gap junctions within hBMSC sheets, we performed a functional blocking experiment using small molecule CBX, a general gap junction inhibitor, demonstrated to inhibit gap junctional communication via reduced dye transfer between cells.28,33,35,44,45 For these experiments, we used clone 1 hcBMSCs. Clone 1 hcBMSCs were seeded for cell sheet fabrication with a gap junction decoupler CBX (100 μM) for the duration of the 6-day cell sheet culture period. The gap junction blockade did not affect cell sheet fabrication, as indicated by similar microscopic cell morphology before cell sheet detachment (Fig. 4a, c) and macroscopic images of the resultant intact cell sheet (Fig. 4b, c).
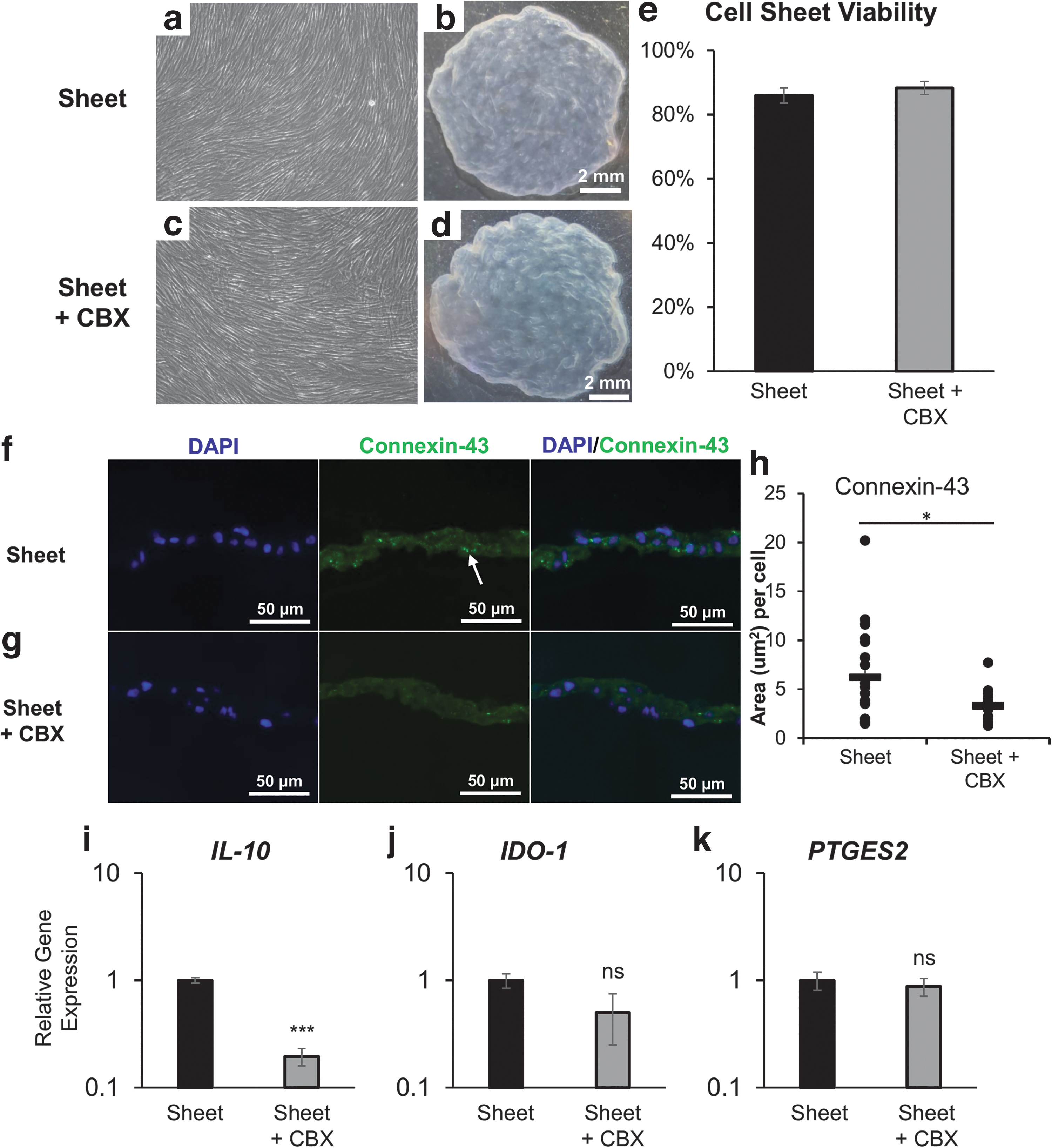
MSC sheets reduce gene expression of immunosuppressive molecules in response to gap junction blockade. Representative
In addition, no toxicity (i.e., reduction in cell viability) was observed following CBX treatment (Fig. 4e). To assess gap junctions within cell sheets, we performed immunohistochemical staining to visualize Cx43 in hcBMSC sheet cross-sections (Fig. 4f, g). Decreased gap junction presence, measured by fluorescence area per cell, was observed in the CBX-treated group (3.31 ± 1.5) compared to hcBMSC sheets without CBX treatment (control) (6.2 ± 4.8) indicating reduction of gap junctions (Fig. 4h).
Retention of functional gap junctions within hBMSC sheets correlates to increased gene expression of immunosuppressive genes
To evaluate the importance of retained functional cell–cell gap junctions within hBMSC sheets, we compared expression of immunomodulatory genes IL-10, IDO-1, and PTGES2 in hcBMSC sheets ± CBX. hcBMSC sheets+CBX downregulated gene expression of IL-10, IDO-1, and PTGES2 to 0.20-fold change, 0.50-fold change, and 0.88-fold change, respectively (Fig. 4i–k). These data indicate that loss of functional gap junction interactions contributes to decreased expression of immunomodulatory molecules within MSCs, most significantly IL-10, and thus, the promotion of cell–cell interactions within cell sheets is important to improve hcBMSC immunomodulatory function. To our knowledge, this is the first report of a direct mechanistic relationship between functional gap junctions within hcBMSC sheets and stimulation of immunomodulatory molecule production.
Discussion
Toward improving MSC-based therapy for immune-related diseases, cell sheets are a promising strategy to enhance culture-expanded MSC expression of immunomodulatory paracrine factors and improve engraftment beyond the capacity of conventional cell suspension injections. 12 Before cell sheet detachment, MSC sheets attached in two-dimensional (2D) cultureware exhibit normal stromal tensions, high cell strain, and forced basal polarization.8,11 During temperature-mediated detachment, MSC sheets lose their points of cultureware contact, actively detach and spontaneously contract to produce a 3D sheet with significantly increased thickness and preserved intercellular and matrix interactions.21,46 This dynamic sheet contraction from 2D to 3D upon detachment (so-called “tissue effect”)21,22,46 promotes multidirectional cell–cell and cell–matrix interactions proposed to stimulate MSC paracrine expression. 21
Changes to cell dimensionality, polarization, and intercellular signaling interactions maintained both during cell sheet fabrication and sheet release prompt this “tissue-effect” to upregulate MSC expression of therapeutic cytokines and growth factors.21,22,46 Therefore, in this study we sought to investigate gene expression changes after cell detachment equivalent to their delivery state, in contrast to assessing 2D adherent cell monolayers on plastic (standard culture method) that do not represent the actual MSC suspension delivery state in vivo.
In accordance with previously reported studies, we report consistent fabrication of 3D hBMSC sheets across all whole and clonal hBMSCs (Fig. 2a).14,46 In addition, all hBMSC sheets upregulate expression of instructive intercellular gap junction (Cx43) and adherens junction (N-cadherin) molecules following temperature-mediated detachment compared to conventional cell suspensions (Fig. 2b, c). Our central finding was that cell–cell interactions, namely GJIC, promoted by cell sheet fabrication contribute to upregulated hcBMSC expression of immunomodulatory factors (Fig. 3). These findings, in combination with previous reports demonstrating rapid gap junction coupling between cell sheets, support a pathway for transplanted MSC sheets to regulate inflammation via gap junctions coupling with the host cells.36,37,47 This study expands current understanding of the cell sheet tissue-effect by reporting direct influence of GJIC on clinically relevant hcBMSC 3D sheet expression of immunomodulatory paracrine factors.21,46
Anti-inflammatory factors IL-10, IDO-1, and PGE2 are commonly implicated in MSC-mediated T cell suppression and regulation,6,48–51 and we have previously demonstrated their role in hcBMSC sheet-mediated suppression of T cells in vitro. 52 Here, we report that hBMSCs as sheets upregulate the expression of IL-10, IDO-1, and PTGES2 (PGE2 synthase) beyond their production in conventional cell suspensions (Fig. 3). Umbilical cord MSC sheets have similarly been reported to upregulate IL-10 expression in relationship to upregulated intercellular signaling molecules connexin-43 and β-catenin.21,22 Qazi et al. 10 also reported higher secretion of paracrine factors, including IL-10, in MSCs encapsulated in biomaterials that promoted a higher degree of cell–cell interactions. Beyond MSCs, T lymphocyte abilities to produce IL-10 are similarly linked to cell–cell interactions; inhibition of gap junction signaling using a synthetic derivative of CBX results in decreased T cell production of IL-10. 53
T cells and MSCs both express gap junction subunits Cx43 and Cx40. 54 Further, Cx43 degradation is proposed to reduce levels of IL-10, and inhibition of Cx43 degradation increases levels of IL-10 in astrocytes. 55 Here, we demonstrate that treating hBMSC sheets with GJIC inhibitor CBX, proven to reduce gap junction signaling in MSCs,28,33,35,44 downregulated immunomodulatory molecules in hBMSCs (Fig. 4), most significantly IL-10, supporting the importance of intercellular gap junction interactions in stimulating IL-10 expression.
IDO-1 expression was similarly downregulated by CBX GJIC inhibition, and while not statistically significant, suggests that preservation of gap junctions in cell sheets regulates IDO-1 expression. PTGES2 showed the smallest change in response to CBX treatment, indicating that its expression is likely not regulated by gap junction metabolic pathways. This finding was unexpected since prior reports consistently show upregulated PGE2 in MSC 3D spheroid culture, where intercellular interactions are also promoted.8,56,57 Instead, other mechanotransduction pathways may be at play, such as the YAP/TAZ pathway, adherens junction molecules, N-cadherin, or other signaling molecules, including β-catenin that has been shown to be upregulated in cell sheets.9,21,46,56 Consistently, N-cadherin inhibition has been shown to disrupt MSC expression of paracrine factors. 10 Hence, N-cadherin interactions within the cell sheet may also play an important role in regulating expression of IL-10, IDO-1, and PTGES2.
Further investigation into these other key signaling pathways may reveal unique links between specific communication pathways and MSC expression of paracrine factors. Use of global analysis techniques such as RNA-seq and broad cytokine panels would best facilitate this further investigation. 58 In addition, beyond the investigation of more general GJIC inhibition, as reported here, more specific RNA-mediated GJIC inhibition approaches could reveal differential roles for MSC-expressed connexins 43, 40, and 45 in cell sheets and their dynamic regulation of downstream therapeutically relevant processes. Ultimately, in vivo studies are required for establishing clinical relevance. In this regard, our group has recently demonstrated therapeutic utility of these hcBMSC sheets in mitigating both acute and chronic fibrotic responses in a nude rat ischemia reperfusion model of renal fibrosis. 16 This model provides a research platform to validate possible therapeutic cell sheet benefits compared to injected single cells, and specific roles of gap junctions related to MSC therapeutic potency.
Finally, while this study focuses on the use thermoresponsive cultureware to fabricate the cell sheets using temperature-mediated detachment, several other methods of cell sheet fabrication are reported using various surface modification and nonsurface modification technologies. 59 Future studies investigating the consistent translation of these findings in other cell sheet detachment systems would prove interesting for the field.
Conclusions
We report the first mechanistic insights into roles of functional gap junctions within MSC sheets in paracrine signaling relevant to immunomodulatory therapeutic functions (Fig. 4). We demonstrate that cell sheet fabrication stimulates important MSC immunomodulatory factors related to intact intercellular interactions compared to respective conventional MSC suspensions. These findings highlight the advantages of MSC sheet technology not only as a tool to develop directly transplantable tissue-like constructs, but also as a strategy to amplify MSC anti-inflammatory properties for therapeutic purposes. In addition, this work contributes to increasing evidence demonstrating the importance of preserving cell–cell interactions as design criteria in tissue engineering to improve MSC functions within implanted regenerative medicine constructs. Finally, the practical clinical inference from this study is that robust MSC sheets that reliably retain essential therapeutic and engraftment capabilities hold potential to advance MSC therapy for localized immune-related diseases compared to currently reported use of MSC injections.
Footnotes
Acknowledgments
SCM Lifescience (Korea) has provided financial support, key reagents, and expertise for this research. We thank S. Bou-Ghannam and K. Kim for scientific comments and N. Metzler for technical edits. The University of Utah HSC Core Flow Cytometry Facility is acknowledged as a valued technical resource. Figures were created with BioRender.com.
Authors' Contributions
C.M.D., S.K., D.W.G., and T.O. conceptualized the study concepts. C.M.D. and S.K. designed and interpreted all experiments and analysis. C.M.D. performed all experiments, data collection, and wrote the article. S.K. performed technical support for cell banking and characterization of hBMSCs and hPBMCs. T.P. contributed to isolation and banking of whole hBMSCs and phenotypic characterization. Y.-K.C. and S.U.S. performed isolation, selection, and technical support for clonal hBMSCs. T.O. and D.W.G. reviewed and edited the article. All authors read and approved the final article.
Disclosure Statement
S.K. and T.O. received compensation from an SCM research contract. Y.-K.C. and S.U.S. hold equity in SCM Lifescience (Korea) and are developers/inventors designated on intellectual property for clonal MSCs. T.O. holds equity in CellSeed, Inc. (Japan). No other competing financial interest exists, and all authors declare that they have no other competing interest.
Funding Information
This research was funded by SCM Lifescience Co Ltd, Incheon, Republic of Korea. Role of funding source: SCM Lifescience (Korea) performed isolation, selection, and technical support for the use of their patented clinical grade clonal BMSCs.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.