
Abstract
Vascular endothelial cells (ECs), locating at the inner side of vascular lumen, play critical roles in maintaining vascular function and participate in tissue repair and neovascularization. Although increasing studies have shown positive effects of transplantation of vascular ECs or their precursor cells on neovascularization and functional recovery of ischemic tissues, the quantity of in vivo ECs is limited and their quality is affected by age, gender, disease, and others, which hinder their clinical application and further study. Chemical transdifferentiation is a promising approach to generate patient-specific cells. In this process, somatic cells are directly converted into desired cell types without the risk of tumorigenicity by pluripotent cell transplantation and exogenous gene introduction by transgene technology. In the present study, we derived ECs from human cardiac fibroblasts (CFs) through an optimized chemical induction method. The derived ECs expressed endothelial specific markers, took up low-density lipoprotein, secreted angiogenic cytokines under hypoxic condition, and formed microvessels in vitro and in vivo. This CF-EC transition bypassed pluripotency and germ layer differentiation, but underwent a stage of endothelialization. Although p53 maintained the same level during the period of CF-EC transdifferentiation, we could modulate p53 transcriptional activity to further improve cell transition efficiency, which mainly functioned at the later stage of endothelialization. Optimization and exploring the regulatory mechanism of CF-EC transition complement each other, which not only broadens the sources of patient-specific ECs but also provides valuable references for the in vivo direct transdifferentiation study and the elucidation of endothelial development and dysfunction.
Impact statement
This study provides an optimized chemical induction method to derive endothelial cells (ECs) from human cardiac fibroblasts (CFs), which not only broadens the sources of patient-specific ECs but also provides a good research model of mesenchymal–endothelial transition. Studying the molecular process and regulatory mechanism of CF-EC transdifferentiation will provide valuable references for the in vivo direct transdifferentiation for clinical therapy and deepen the understanding of endothelial development and dysfunction.
Introduction
Vascular endothelial cells (ECs), located at the inner side of vascular lumen, play critical roles in maintaining vascular function, such as regulating permeability, secreting cytokines, participating in vascular repair, and neovascularization. 1 Loss or dysfunction of vascular ECs usually results in severe vascular diseases. In 1997, researchers isolated endothelial progenitor cells (EPCs) from human peripheral blood for the first time, and their following studies showed that autologous transplantation of EPCs improved ventricular remodeling after myocardial infarction and reduced amputation rate of patients with critical limb ischemia.2,3 Although increasing studies have suggested positive effects of transplantation of vascular ECs or their precursor cells on neovascularization and functional recovery of ischemic tissues, the quantity of in vivo ECs is limited and their quality is affected by age, gender, disease, and others, which hinder their clinical application and further study. 2
The establishment of induced pluripotent stem cells (iPSCs) provides a novel cell source for autologous transplantation therapy due to its infinite proliferation capacity and multiple differentiation potentials.4,5 Especially in recent years, nonintegrated and small molecule-induced cell reprogramming technology improves the safety of iPSCs for clinical application, which avoids exogenous and even carcinogenic genes being integrated into recipient genome. 6 However, as to pluripotent stem cells, people are still anxious about their tumorigenicity.
Transdifferentiation is another approach to generate patient-specific cell types, and in this process, somatic cells are directly converted into desired somatic cell types without the risk of teratoma formation by iPSCs.7,8 Thus the advantages of small molecule-induced somatic cell transdifferentiation are as follows: (1) no exogenous gene is introduced, ensuring the stability of cell genome; (2) cell transformation bypasses pluripotent state, reducing the tumorigenicity of donor cells; (3) small molecules are easy to synthesize and store, ensuring good experimental repeatability; and (4) cell derivation is a time- and concentration-dependent process, facilitating standardization and large-scale production.
In 2016, Wong and Cooke provided the first report of direct reprogramming of human somatic cells into ECs by forced expression of endothelial specific transcription factor ETV2, Fli1, Gata4, and Klf4, which bypassed an intermediate pluripotent state. 9 In 2015, Sayed et al. demonstrated that functional ECs could also be directly generated from human newborn foreskin fibroblasts only using chemical small molecules. 10 Cognizant of the role of innate immunity in nuclear reprogramming and its directed manipulation to increase epigenetic plasticity, the researchers used PolyI: C, a Toll-like receptor 3 agonist, to activate innate immunity, followed with microenvironment-based induction to endothelial lineage. This is the first trail using chemical reprograming to derive ECs from somatic cells.
Cardiac fibroblasts (CFs) arise from several origins and make up the largest cell population of mammalian heart. Heart after injury heals primarily by CF-mediated fibrosis. CFs proliferate at the site of injury and recruit circulating ECs, which contribute to neovascularization and cardiac repair.11,12 In fact, fibrosis plays a negative role in cardiac function recovery, while the role of neovascularization is just the opposite. Considering the close interaction between CFs and ECs during cardiac repair, researchers are interested in the CF-EC transition, which contributes to the new sources of ECs for rapid neovascularization. 12 Thus, in the present study, we intend to derive ECs from human CFs using a chemical induction method and study the underlying regulatory mechanism, which will provide valuable references for the in vivo direct transdifferentiation for clinical therapy and deepen the understanding of endothelial development and dysfunction.
Materials and Methods
Ethics statement
Animal experiments were approved and conducted according to the guidelines of the Biomedical Ethics Committee of Health Science Center of Xi'an Jiaotong University, P. R. China. The approval number is XJTUAE2023-1426. We performed all surgeries under anesthesia using isoflurane and tried our best to minimize animals suffering.
Cell culture and CF-EC derivation
Human CFs (CP-H078; Procell) were seeded on 0.1% gelatin-coated plates and maintained in Dulbecco's modified Eagle's medium (DMEM) with 15% fetal bovine serum (FBS). Transdifferentiation of CFs to ECs (CF-ECs) was progressive and according to Sayed et al.'s publication with some modification. 10 First, the cells were treated with 20 ng/mL Poly I: C, and the medium was changed to DMEM with 5% FBS and 10% knockout serum replacement (KOSR). After 4 days, the cells were cultured with DMEM containing 20 ng/mL BMP4, 50 ng/mL VEGF, 20 ng/mL bFGF, and 10% KOSR for another 8 days. Then the medium was changed to Endothelial Cell Growth Medium-2 BulletKit (EGM-2; Lonza) with addition of 50 ng/mL VEGF and 0.1 mM/L 8-bromoadenosine-3′: 5′-cyclic monophosphate sodium salt (8-Br-cAMP; Sigma). Twelve days later, the cells were purified by fluorescence activated cell sorting (FACS) and expended with EGM-2 containing 50 ng/mL VEGF and 10 μM/L SB421542. Throughout the CF-EC derivation, medium was changed every day.
Small molecule treatment
Commercially obtained nutlin-3 (N6782; Sigma) and pifithrin-a (P4359; Sigma) were used for p53 gain- and loss-of function assay. At different stages of CF-EC derivation, the final concentration of 5 nM nutlin-3 or 20 μM pifithrin-a was added into the culture media to activate or reduce p53 transcriptional activity. After three stages of induction, the expression levels of endothelial markers and the percentages of CD31 positive cells were explored to evaluate the effects of p53 activity on CF-EC transdifferentiation.
Immunofluorescence staining
Following our previous publication, 13 the cells were fixed in 4% (w/v) paraformaldehyde (PFA) at room temperature for 30 min, permeabilized in 1% (v/v) Triton X-100 in phosphate-buffered saline (PBS) at 37°C for 1 h, and blocked with 1% bovine serum albumin at 37°C for 1 h. Subsequently, the cells were incubated with primary antibody overnight at 4°C. After three washes with PBS, they were incubated with secondary antibodies for 1 h at 37°C. Finally, nuclei were stained with Hoechst33342 for 5 min. Samples were visualized on Nikon 80i fluorescence microscope. Primary antibodies and their working concentrations/dilution ratios are listed in Supplementary Table S1.
Flow cytometric analysis
Adherent cells were harvested using TrypLE™ Express and resuspended in PBS. The cells were passed through a 50 μm cell strainer and incubated with anti-human CD31 antibody (Alexa Fluor® 488 mouse monoclonal to CD31, ab187594) for 30 min at room temperature. Then the cells were washed thrice in PBS and centrifugated at 300 g for 5 min. After washing, the labeled cells were sorted by flow cytometry on a FACSCalibur and analyzed with FlowJo software.
Uptake of Dil-acetylated low density lipoprotein
According to the manufacturer’ s instruction, the cells cultured on Matrigel were incubated with 20 μg/mL human Dil-acetylated low density lipoprotein (YEASEN) in EGM-2 at 37°C. After 4 h of incubation, the cells were washed thrice with PBS and visualized with a Nikon 80i fluorescence microscope.
In vitro tube formation assay
Following our previous publication, 13 the cells were plated at a density of 1 × 104 cells per well in 96-well plates (Corning) coated with 50 μL growth factor-reduced Matrigel (Corning). The plates were incubated at 37°C for 4–6 h. Then the images were taken from each well using a Nikon 80i microscope.
In vivo Matrigel plug assay
The cells were dissociated using TrypLE Express. Following our previous publication, 13 a total of 1 × 107 cells were resuspended in 200 μL growth factor-reduced Matrigel (Corning) supplemented with 300 ng/mL human FGF-2, and subcutaneously, they were injected into the abdominal flanks of 8-week-old male NOD/SCID mice. Fourteen days later, the Matrigel plugs were removed and fixed with 4% PFA at 4°C for 12 h. After dehydration with ethanol, they were embedded in paraffin.
Histological analysis
Paraffin embedded samples were cut into 7 μm sections using a tissue slicer (Leica). Hematoxylin/eosin (HE) staining was performed with HE Staining Kit (Beyotime, China). Following our previous publication, 13 immunohistochemistry (IHC) staining was performed by the following steps: the sections were permeabilized with 0.1% (v/v) Triton X-100 in PBS for 5 min. After blocking with 10% (v/v) horse serum for 30 min, the sections were incubated with primary antibodies overnight at 4°C, followed by incubation with secondary antibodies and Hoechst33342 staining. Then the samples were visualized by Nikon 80i fluorescence microscope. Primary antibodies and their working concentrations/dilution ratios are listed in Supplementary Table S1.
Western blotting
Following our previous publication, 14 the cells were lysed using lysis buffer radioimmunoprecipitation assay (Beyotime) which contains phenylmethylsulfonyl fluoride (Roche, Switzerland). Approximately 30 μg of total protein was electrophoresed on a 10% sodium dodecyl sulfate–polyacrylamide gel and electro-transferred to a polyvinylidene difluoride membrane (Millipore). The membrane was blocked with 5% FBS for 2 h, immunoblotted with primary antibody overnight, and incubated with secondary antibody for 60 min. The results were visualized by DAB Horseradish Peroxidase Color Development Kit (Beyotime). Primary antibodies and their working concentrations/dilution ratios are listed in Supplementary Table S1.
Quantitative real time–polymerase chain reaction
We followed our previous publication with some modifications. 13 RNA isolation was performed according to the manufacturer’ s instruction using PureLink RNA Mini Kit (Life). The RNA concentration was measured with NanoDrop 2000 spectrophotometer (Thermo Scientific), and 500 ng RNA from each sample was converted to cDNA using High Capacity cDNA Reverse Transcription Kit (Thermo Scientific) according to the manufacturer's protocol. The cDNA was used for quantitative real time–polymerase chain reaction in the presence of specific primers and SYBR Green PCR Master Mix (Takara). Thermal cycling was conducted on 7900HT sequence detection system. Primers for the detected genes are listed in Supplementary Table S2.
Angiogenesis proteome profiler antibody array assay
Cytokines secreted by CF-ECs under hypoxic condition were assessed using Human Angiogenesis Proteome Profiler™ Array (ARY007; R&D Systems). CF-ECs were grown under 5% oxygen at 37°C for 12 h, and the conditioned media were collected. After centrifugation, the supernates were diluted, mixed, and incubated with Detection Antibody Cocktail at room temperature for 1 h, and then the sample/antibody mixtures were added to the nitrocellulose membrane. Following overnight incubation at 4°C, the membrane was washed thrice to remove unbound material. Streptavidin-horseradish peroxidase and chemiluminescent detection reagents are added sequentially according to the manufacturer's instruction. The array data were quantified by densitometry using ImageJ software.
Statistical analyses
To label CF-ECs and quantify their in vivo angiogenic capacity, histological sections were stained with anti-human CD31 antibody and the capillary-like structures were identified. ImageJ software was used to analyze capillary density, which was presented as percentage of capillary-like structures in a random field. For each treatment, six random fields were observed by two independent observers and then averaged. The averages were used for statistical analysis. Data were obtained from at least three independent experiments. All statistical analyses were performed with GraphPad Prism5. A power of analysis with 95% confidence interval was used to calculate sample size required to obtain significant results (p < 0.05).
Results
Derivation of ECs from human CFs
The transdifferentiation protocol of human CFs into ECs is shown in Figure 1A. In the first stage of induction, the attached CFs were treated with Poly I: C for innate immunity activation to facilitate EC derivation, 10 and after three stages of continuous induction, some clustered cobblestone-like cells emerged, which were CD31 positive (Fig. 1B–D). Flow cytometric sorting was used to obtain the CD31 positive cells, which accounted for 4.58 ± 0.79% of the derived cells (Fig. 1E). After two generations of expansion, the percentage of CD31 positive cells was still above 90% (Supplementary Fig. S1).
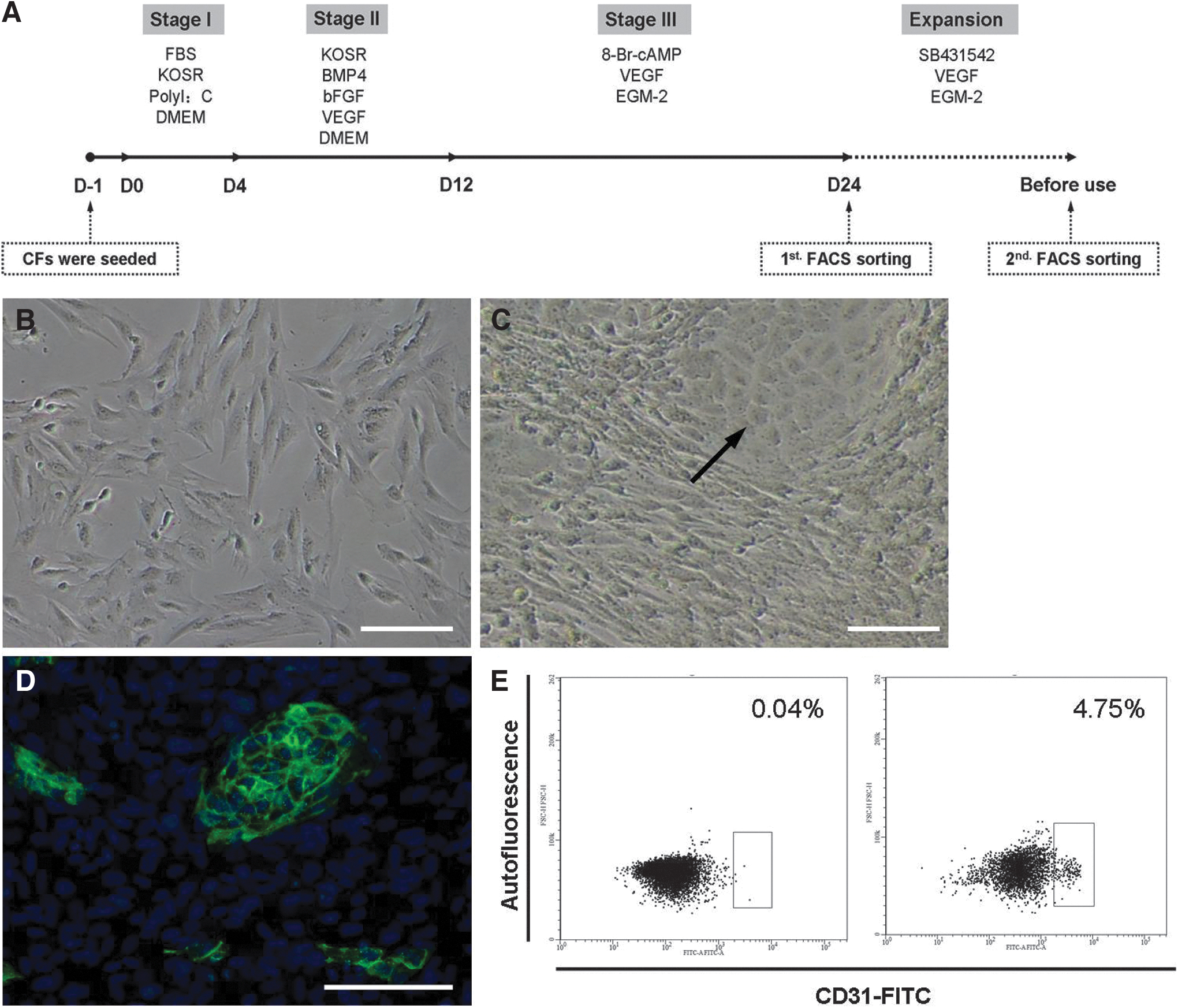
Derivation of ECs from human CFs.
Characterization of the derived CF-ECs
To monitor the transdifferentiation process, expression levels of endothelial and fibroblast specific genes were detected during the derivation period. As shown in Figure 2A, B, the levels of endothelial markers increased with the time, while those of fibroblast markers exhibited just the opposite, indicating continuous endothelialization of the human CFs. FACS purified CF-ECs had typical morphological characteristics of ECs and expressed endothelial markers, such as CD31, CD144, eNOS, and vWF (Fig. 2C–F). When CF-ECs were seeded onto Matrigel, they could uptake low-density lipoprotein and form microvessel-like structures (Fig. 2G, H). To further confirm the function of CF-ECs, we assessed their secretion levels of angiogenic cytokines under hypoxic condition. CF-ECs secreted a set of angiogenic cytokines when subjected to hypoxia, including TSP1, TIMP1, Serpin E1, ANG, ANGPT1, ANGPT2, IGFBP2, IGFBP3, and PTX3, whose levels were comparable to those of HMVECs (Fig. 2I).
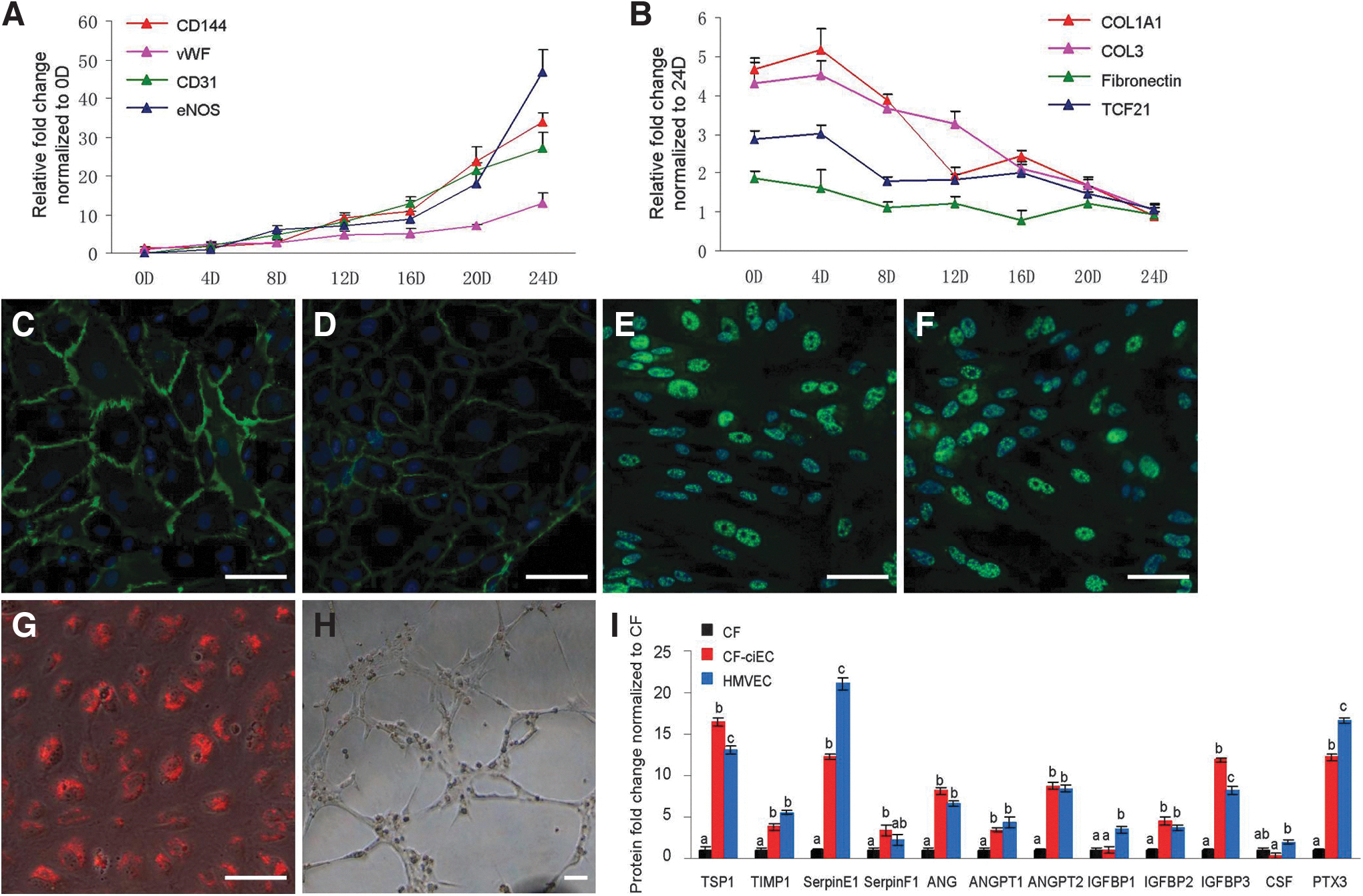
Characterization of the derived CF-ECs.
In vivo angiogenesis of the derived CF-ECs
CF-EC contained Matrigel was injected into the abdominal flanks of NOD/SCID mice to identify their in vivo angiogenic ability. After 14 days of development in mice, CF-ECs stretched along each other and formed vessel-like structures, and in the cross-section, we could find aggregated red blood cells, indicating mature blood vessels formed in the CF-EC treated group (Fig. 3A–C). To quantify angiogenic ability of CF-ECs, histological sections were stained with anti-CD31 antibody, which was specific for vascular endothelium (Supplementary Fig. S2). IHC staining result showed that CF treated group was almost negative for CD31, but CF-EC treated group exhibited obvious CD31 expression and vascular structures, which were consistent with the result from morphological assay (Fig. 3D–I). The result of statistical analysis showed that the total capillary density of CF-EC treated group was 10.03 ± 2.00%, which was significantly higher compared with CF treated group (1.63 ± 0.86%; Fig. 3J).
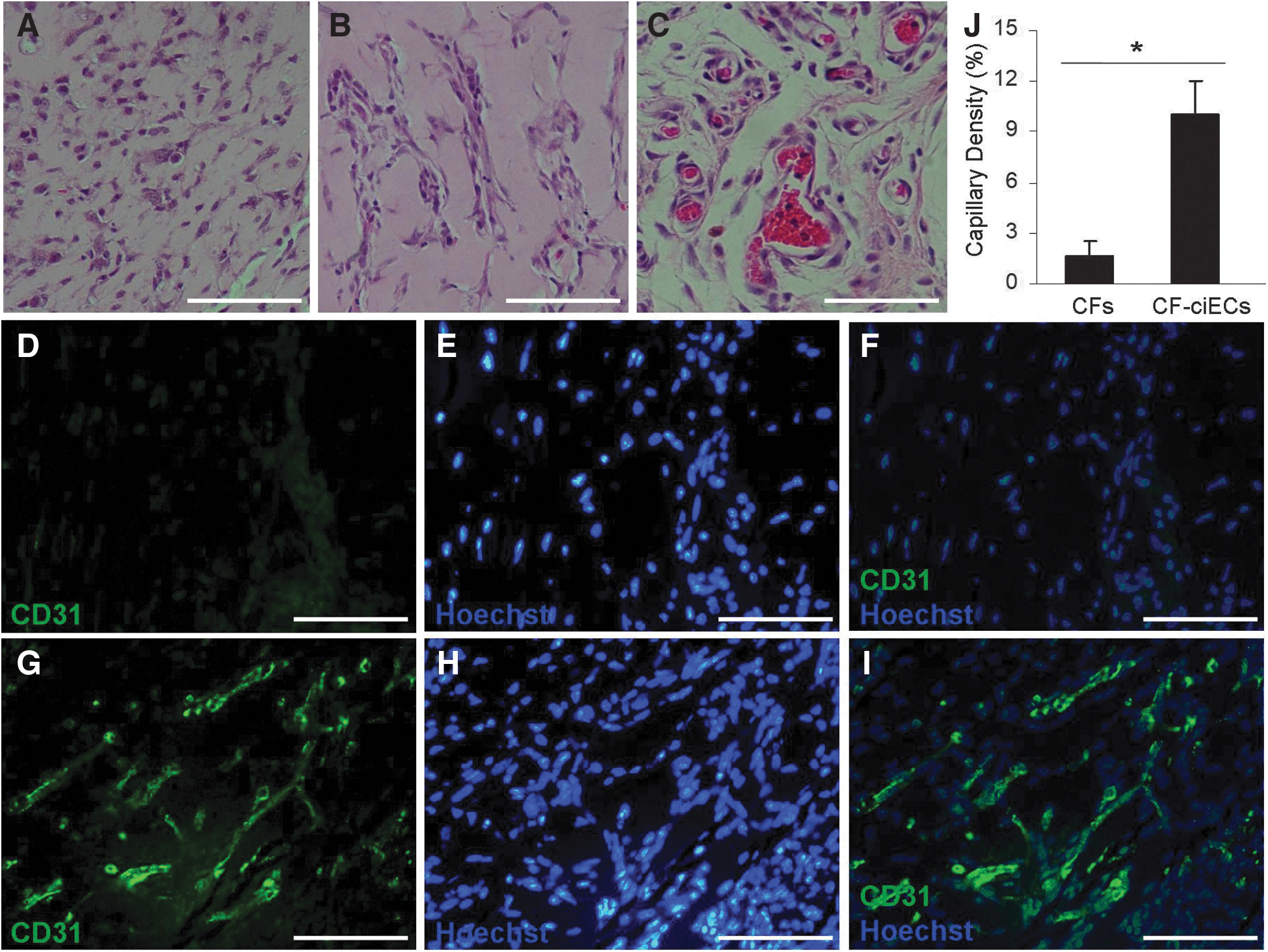
In vivo angiogenesis of the derived CF-ECs.
CF-EC derivation bypasses pluripotency and germ layer differentiation
Direct cell reprogramming, also known as cell transdifferentiation, refers to transformation from a somatic cell type into another without undergoing an intermediate state. Thus we detected the expression profiles of pluripotency, germ layer differentiation, and endothelialization related genes to explore CF-EC transdifferentiation pathway during the derivation period.
As shown in Figure 4A, B, there was no obvious change in the expression of pluripotency markers Nanog, Oct4, or Sox2 nor was the expression of most germ layer marker genes. Only the mesoderm marker PDGFR2 decreased more than 90% during the 24 days of transdifferentiation (Fig. 4B). Among the endothelial progenitor markers, Flk1 and CD34 kept increasing, while ETV2 and Scl increased from days 0 to 12 and maintained approximately constant levels from then on (Fig. 4C). Consistent with the transcription levels, the protein levels of Flk1 and CD34 increased throughout the derivation period (Fig. 4D). These results suggested that CF-EC transdifferentiation bypassed pluripotency and germ layer differentiation, but it underwent an endothelial progenitor stage.
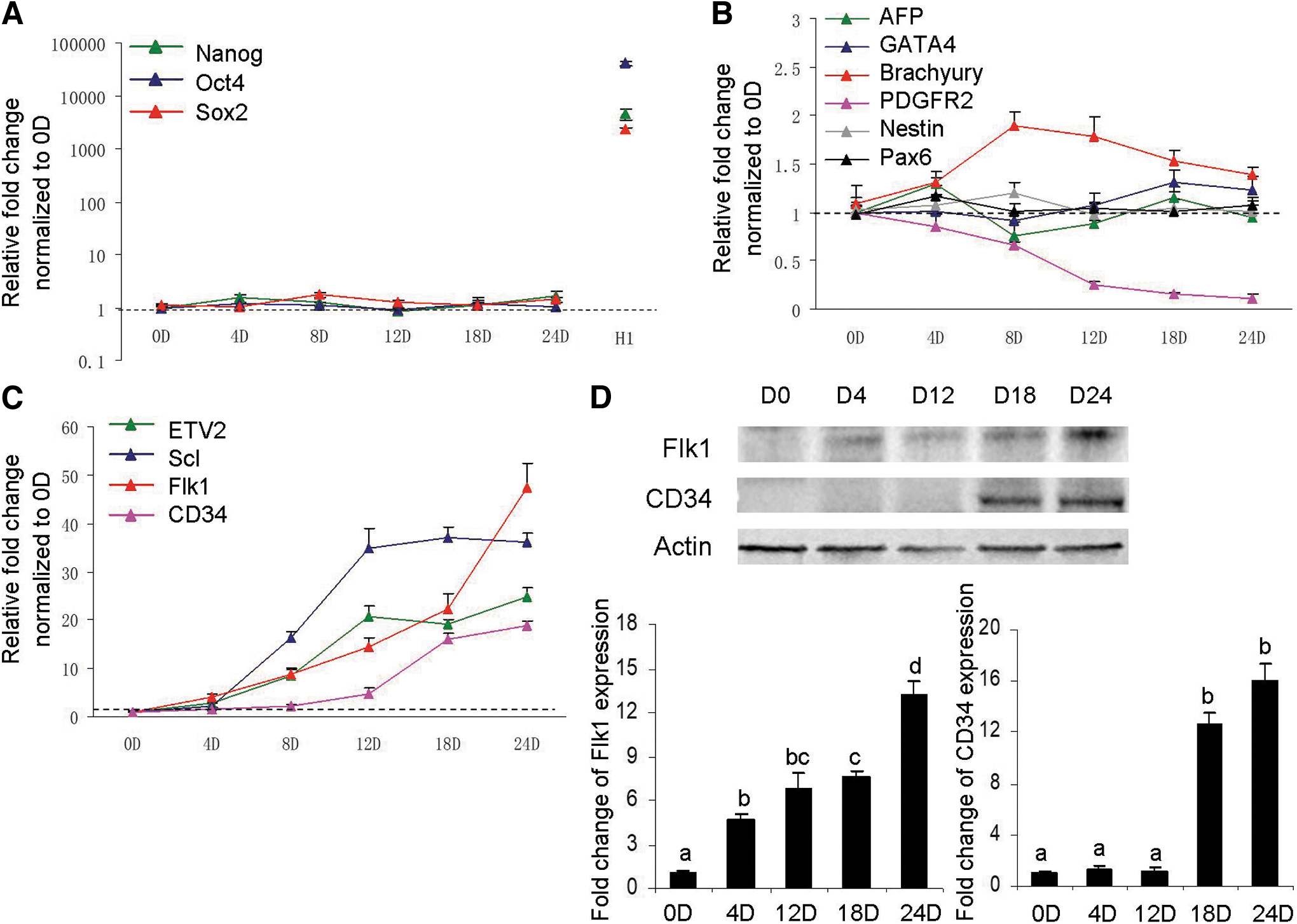
CF-EC derivation bypasses pluripotency and germ layer differentiation.
p53 activation promotes transdifferentiation of CF-ECs
Former studies showed that p53 plays critical roles on mesenchymal–endothelial transition after ischemic cardiac injury and astrocyte-endothelial transition under cellular stress.12,15 However, different from Ubil et al.'s report that p53 protein levels increased upon myocardial ischemia, 12 we did not observe p53 activation throughout the CF-EC transdifferentiation process (Supplementary Fig. S3). To determine whether p53 participates in our endothelial induction process, Nutlin-3, a compound which stabilizes p53 protein by inhibiting p53 from interacting with its negative regulator MDM2, and pifithrin-a, a widely used p53 inhibitor which blocks p53 transcriptional activity by modulating p53 nuclear import and/or export, were used for p53 gain- and loss-of function assay.16,17 As shown in Figure 5A, B, p53 protein levels were upregulated along with the increasing of Nutlin-3 dosages, and p53 target genes were activated accordingly. Upon pifithrin-a treatment, the levels of p53 target genes decreased, especially with the concentrations of 20 and 50 μM (Fig. 5C). Thus we adopted 5 nM of Nutlin-3 and 20 uM of pifithrin-a for the subsequent experiments.
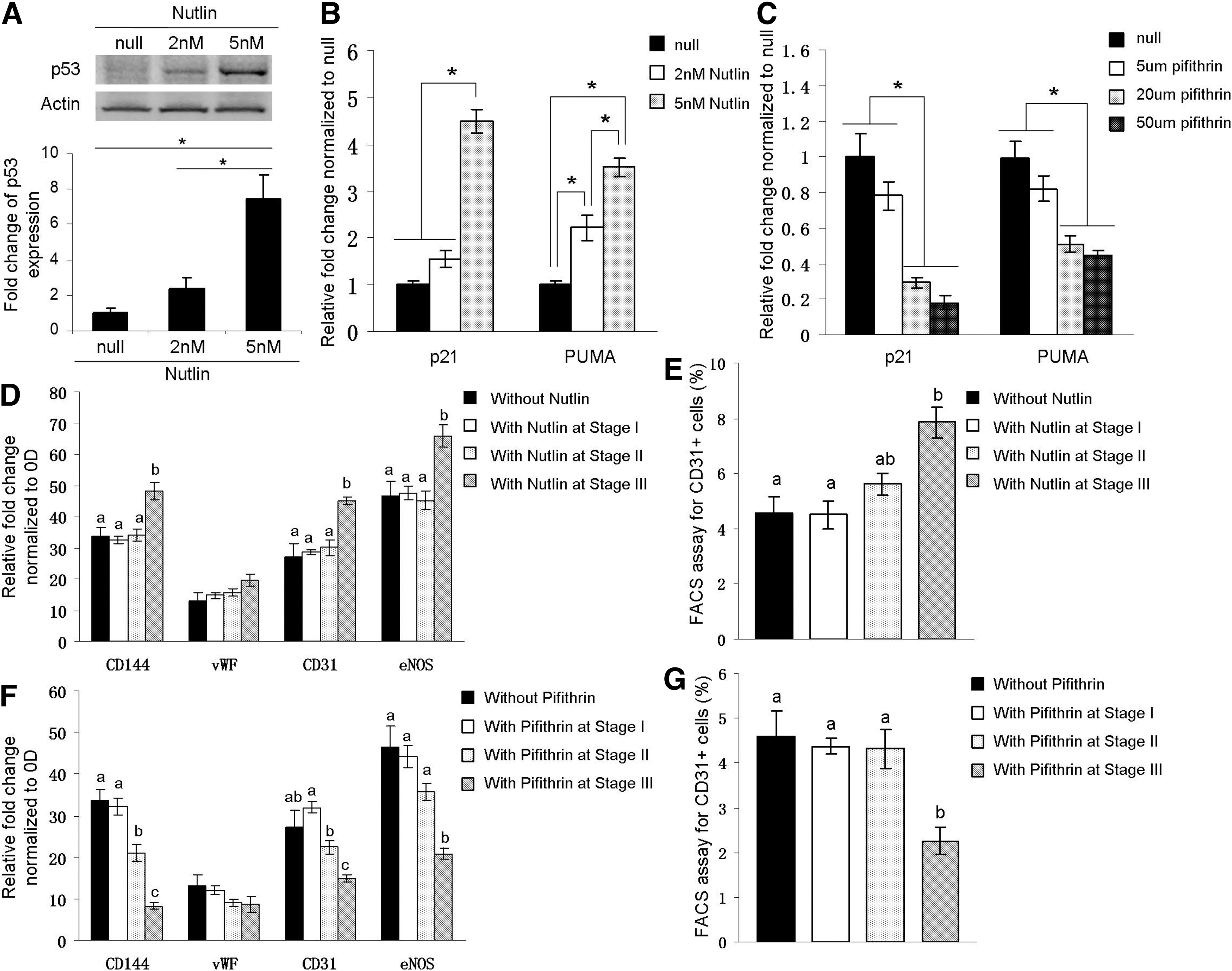
p53 activation enhances transdifferentiation of CF-ECs.
As shown in Figure 5D, E, when Nutlin-3 was added in stage I or stage II of CF-EC derivation, there was no obvious change of expression levels of endothelial markers or percentages of CD31 positive cells, but when it was added in stage III, it significantly promoted the expression levels of endothelial marker CD144, CD31, and eNOS and the percentages of CD31 positive cells. The results regarding pifithrin-a were relatively complex. When it was added in stage II, the expression levels of a part of endothelial markers decreased, but the percentages of CD31 positive cells were maintained (Fig. 5F). When pifithrin-a was added in stage III, it significantly reduced the expression levels of endothelial marker CD144, CD31, and eNOS and the percentages of CD31 positive cells (Fig. 5G). The above results suggested that p53 activity modulated the CF-EC transdifferentiation process, which mainly functioned in the later stage of endothelialization.
Discussion
Using an optimized small molecule-induced method, ECs could be derived from human CFs directly. The derived ECs expressed endothelial specific markers CD31, CD144, vWF, and eNOS, took up low-density lipoprotein, secreted angiogenic cytokines under hypoxic condition, and formed microvessels in vitro and in vivo. Further study showed that the CF-EC transition bypassed pluripotency and germ layer differentiation, and p53 activity regulated the transdifferentiation process, which mainly functioned in the later stage of endothelialization. Thus modulating p53 and its downstream effectors may be an effective way to optimize the current EC inducing protocol.
From the viewpoint of developmental biology, ECs have two distinct cell origins, namely hemangioblasts and angioblasts. 18 Both of them generate from Flk1 positive mesodermal cells, so endothelial development can be generally divided into mesoderm differentiation and endothelial specification. To derive ECs, manipulating the signaling pathways that participate in the two phases is the main strategy.19,20
In previous study, Sayed et al. demonstrated the necessity of innate immune activation for endothelial induction from fibroblasts, which permitted them to respond to microenvironmental cues with a phenotypic plasticity. 10 Different from adult CFs we used in the study, the cell that Sayed et al. used was newborn foreskin fibroblasts, which had better proliferating ability and less heterogeneity.21,22 CFs arise from several origins and make up the largest cell population of heart, 23 while our study showed that Poly I: C, even at a lower concentration, played an effective role on innate immune activation. Growth factors BMP4 and bFGF are commonly used for EC induction, especially BMP4, which functions not only on mesoderm differentiation but also on endothelial progenitor generation. 24
VEGF plays a critical role in endothelial specification by binding to its receptor tyrosine kinase Flk1, and once withdrawn, the Flk1 positive mesodermal cells tend to become hematopoietic cells.25,26 In our study, this is the reason VEGF was applied throughout the endothelial inducing period even during EC expansion.
The effect of TGF-β on angiogenesis is relatively complex, depending on species, developmental stage, and microenvironment. Bai et al. found that upon mesoderm differentiation from human embryonic stem cells, addition of TGF-β type I receptor inhibitor SB431542 reduced the expression levels of mesodermal markers and the number of endothelial progenitors. 27 While when Aoki et al. derived ECs from human iPSCs, they found that combination of three small molecules, namely Y-27632 (a selective inhibitor of ROCK), A83-01 (a receptor-like kinase inhibitor of TGF-β), and CHIR-99021 (a selective inhibitor of GSK3b), extremely improved the expansion of ECs. 28 In a previous study, we systematically explored the effect of TGF-β throughout the EC derivation from porcine iPSCs and found that TGF-β signaling played positive roles in mesoderm formation and negative roles in its further endothelialization; thus, inhibiting TGF-β signaling upon endothelial induction might be a good strategy to promote endothelial specification. 14
During CF-EC transdifferentiation, there were no obvious changes in the expressions of pluripotency and most germ layer marker genes, except for mesoderm marker PDGFR2. We speculate that although PDGFR2 is a mesoderm gene, it is continuously expressed by most fibroblast lineage, thus along with the transdifferentiation process, its expression decreased, just like other fibroblast markers. In our culture system, CF-EC transition bypassed pluripotency and germ layer differentiation, but it underwent an endothelial progenitor stage, which was different from Sayed et al.'s result that no c-kit or CD34 positive cells were detected. 10
In recent years, great progress has been made to delineate cellular sequence leading to hemangioblast, hematopoietic, and endothelial development and tracking cells expressing specific genes facilitates our understanding of the mesodermal commitment to hematopoietic and ECs. 18 Brachyury is a member of T-box gene family and its expression in the primitive streak persists up to E9.5 of mice, but becomes undetectable in paraxial-lateral mesoderm.29,30 Flk1 is first detected in the extraembryonic and paraxial-lateral mesoderm, which is destined to become heart, and subsequently, Flk1 is confined to ECs of the yolk sac blood islands and the developing endocardial tube. 18 In 2003, Fehling et al. examined Brachyury+ embryonic stem cells and demonstrated that Flk1 was turned on within a subset of Brachyury+ cells. 31 Then a subpopulation of Flk1+ cells began to express Scl, giving rise to Flk1+Scl+ cells. The cells present in the early stage of embryoid bodies were enriched for blast colony-forming cells, suggesting that the Flk1+Scl+ cells can be identified as hemangioblasts. 32 These studies indicated that there are two populations of endothelial progenitors, Scl dependent and Scl independent, and the Scl-dependent ECs develop from hemangioblasts. 18 Thus the differences of cell transdifferentiation pathways caused by different culture systems or induction protocols may explain the discrepancy in expression patterns of endothelial precursor markers between our study and previous report. 10
In summary, this study provides an optimized chemical induction method to derive ECs from human CFs, which not only broadens the sources of patient-specific ECs but also provides a good research model of mesenchymal–endothelial transition. Studying the molecular process and regulatory mechanism of CF-EC transdifferentiation will provide valuable references for the in vivo direct transdifferentiation for clinical therapy and deepen the understanding of endothelial development and dysfunction.
Footnotes
Authors' Contributions
Y.Z. and X.L. conceived the study, carried out the experiments, analyzed the data, and drafted the article; H.T. and M.X. participated in the experiments and the data analysis; Y.Z. drafted and revised the article; J.Z. and H.L. supervised the study and supplied the funding. All authors have read, discussed, and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (Grant No.: 32370854) and the Natural Science Foundation of Heilongjiang Province of China (Grant No.: JQ2021C006).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.