
Abstract
Spinal cord injury (SCI), caused by significant physical trauma, as well as other pathological conditions, results in electrical signaling disruption and loss of bodily functional control below the injury site. Conductive biomaterials have been considered a promising approach for treating SCI, owing to their ability to restore electrical connections between intact spinal cord portions across the injury site. In this study, we evaluated the ability of a conductive hydrogel, poly-3-amino-4-methoxybenzoic acid-gelatin (PAMB-G), to restore electrical signaling and improve neuronal regeneration in a rat SCI model generated using the compression clip method. Gelatin or PAMB-G was injected at the SCI site, yielding three groups: Control (saline), Gelatin, and PAMB-G. During the 8-week study, PAMB-G, compared to Control, had significantly lower proinflammatory factor expression, such as for tumor necrosis factor -α (0.388 ± 0.276 for PAMB-G vs. 1.027 ± 0.431 for Control) and monocyte chemoattractant protein (MCP)-1 (0.443 ± 0.201 for PAMB-G vs. 1.662 ± 0.912 for Control). In addition, PAMB-G had lower astrocyte and microglia numbers (35.75 ± 4.349 and 40.75 ± 7.890, respectively) compared to Control (50.75 ± 6.5 and 64.75 ± 10.72) and Gelatin (48.75 ± 4.787 and 71.75 ± 7.411). PAMB-G-treated rats also had significantly greater preservation and regeneration of remaining intact neuronal tissue (0.523 ± 0.059% mean white matter in PAMB-G vs 0.377 ± 0.044% in Control and 0.385 ± 0.051% in Gelatin) caused by reduced apoptosis and increased neuronal growth-associated gene expression. All these processes stemmed from PAMB-G facilitating increased electrical signaling conduction, leading to locomotive functional improvements, in the form of increased Basso–Beattie–Bresnahan scores and steeper angles in the slope test (76.667 ± 5.164 for PAMB-G, vs. 59.167 ± 4.916 for Control and 58.333 ± 4.082 for Gelatin), as well as reduced gastrocnemius muscle atrophy (0.345 ± 0.085 for PAMB-G, vs. 0.244 ± 0.021 for Control and 0.210 ± 0.058 for Gelatin). In conclusion, PAMB-G injection post-SCI resulted in improved electrical signaling conduction, which contributed to lowered inflammation and apoptosis, increased neuronal growth, and greater bodily functional control, suggesting its potential as a viable treatment for SCI.
Impact Statement
Spinal cord injury (SCI) leads to electrical signaling disruption and loss of bodily functional control below the injury site. Due to their ability to electrically reconnect intact spinal cord portions across the injury site, conductive biomaterials have been considered a promising approach for SCI treatment. We thus developed the conductive hydrogel PAMB-G and found that, compared to untreated and gelatin-injected rats, PAMB-G injection post-SCI resulted in improved electrical signaling conduction, in turn contributing to lowered inflammation and apoptosis, increased neuronal growth, and greater bodily functional control, suggesting its potential as a viable SCI treatment method.
Introduction
Spinal cord injury (SCI) is a severe neurological disorder resulting in the loss of voluntary and involuntary control of bodily functions below the injury site. This often occurs due to physical trauma from various activities, ranging from car accidents to sports injuries. 1 The trauma triggers various secondary pathophysiological processes, such as inflammation, ischemic-associated cell death, demyelination, protein degradation, as well as the production of regeneration-inhibiting molecules, all of which contribute to cavitation and glial scar tissue formation. 2 These processes are responsible for preventing axonal regeneration between intact spinal cord portions, and thus electrical signaling restoration between the central nervous system (CNS) and the peripheral nervous system (PNS), which is crucial for controlling various bodily functions.
Current SCI treatments, ranging from surgery, rehabilitation protocols, and pharmaceutical agents such as the steroid methylprednisolone, have limited effectiveness as they focus on palliation and limiting the spread of further tissue damage rather than restoring bodily functions.3,4 This is due to both the aforementioned secondary pathophysiological processes as well as mammalian CNS having limited postinjury regeneration capacities. 5 In the last decade, a variety of approaches have been developed to bypass this limitation, ranging from using enzymes to remove regeneration-inhibiting molecules, stimulating mitochondrial biogenesis via β2-adrenergic receptor agonists, and transplanting stem cells, such as human pluripotent-derived neuronal or ependymal precursors, at the injury sites.5–8 One such approach is biomaterials, and a number of studies have demonstrated their effectiveness in restoring CNS signaling to the PNS post-SCI, as they are able to reconnect the intact portions of the spinal cord across the lesion site, thereby aiding in restoring proper CNS-PNS electrical conduction. 8 Biomaterials range from biopolymer matrices, such as imidazole-poly(organo phosphazene) or self-assembling peptide-based hydrogels, growth factor-conjugated scaffolds, such as platelet-derived growth factor (PDGF)-A conjugated to a hyaluronic acid-methylcellulose backbone, and those made from conductive materials, such as poly-pyrrole (PPy) conjugated to either poly-lactic (PLA) or tannic acid.2,9–12 All of these biomaterials are able to improve the execution of both involuntary and voluntary bodily functions, in both post-SCI rodent and human subjects, through filling in cavitations, remodeling the extracellular matrix, stimulating neuronal differentiation and growth of implanted or endogenous neuronal stem cells, as well as restoring electrical conduction between intact spinal cord portions.7–12 The last approach, in particular, has led to the development of conductive biomaterials, such as PLA/PPy and tannic acid-PPy, an attractive research field.11,12 This is attributed to the presence of π-bonds within the molecular structures of these materials, which allow the free flow of electrons between different atoms, serving as the basis for electrical conduction. This property facilitates electrical signaling conduction to the intact spinal cord portions post-SCI. These signals, in turn, stimulate neuronal growth factor production from the remaining neurons, thereby activating pathways promoting axonal growth across the injury site and restoring electrical conduction throughout the spinal cord.13,14
In this study, we applied the conductive biomaterial poly-3-amino-4-methoxybenzoic acid-gelatin (PAMB-G) within an SCI model to restore proper CNS electrical signaling in the injured spinal cord. PAMB has been demonstrated to facilitate free movement of electrons between the atoms in the polymer.11–13,15 More importantly, it is a self-doping polymer, resulting in superior conductive capabilities at physiological pH, compared to other materials. Furthermore, conjugation of the conductive biopolymer, PAMB, to gelatin enables its biocompatibility with existing tissue in vivo, owing to the collagen-based composition of gelatin being similar to that of the extracellular matrix. As spinal cord tissues require proper electrical conduction to fulfill their associated tasks of CNS-PNS signal transportation, 15 we postulated that PAMB-G could also aid in restoring spinal cord electrical conduction. Our findings in a rat SCI model validated this hypothesis, in which PAMB-G was able to reduce post-SCI inflammatory responses and better preserve spinal cord neuronal tissues; the latter possibly stems from the conductive properties of PAMB-G enabling the restoration of proper electrical signaling, in turn serving as the basis for motor functional improvements. PAMB-G may thus serve as an effective treatment approach to restore bodily functions negatively affected by SCI.
Methods
Detailed methods for all biochemical analyses can be found in Supplementary Materials.
Establishing the rat SCI model
Adult Sprague Dawley rats (Charles River) were used. All procedures were approved by the Animal Care Committee of Toronto General Hospital and were performed according to the Guide for the Care and Use of Laboratory Animals (NIH, 8th Edition, 2011). Surgical procedures were conducted as described in previous studies, with minor modifications.16–18 Briefly, rats were anesthetized under ventilation with 3% isoflurane, placed on a 37°C heating pad, and laminectomies performed at the level of T10 to expose the thoracic spinal cord segment, with the dura remaining intact. For SCI rats, a cylinder weighted 20 g, with a 0.05 cm2 cross-sectional area, was dropped freely from 3 cm to compress the spinal cord at the T10 segment. Afterwards, all rats were randomly divided into Control, Gelatin, and PAMB-G groups.
Synthesis and injection of the PAMB-G copolymer
A 1:10 ratio for 3-amino-4-methoxybenzoic acid (AMB) and gelatin, respectively, was used to generate the injectable PAMB-G copolymer as it was previously reported to yield the optimal conductivity. 15 Gelatin powder (2 g) was dissolved in 5 mL deionized (DI) water at 60°C, followed by adding and mixing with 200 mg AMB (Alfa Aesar). AMB monomer polymerization and grafting of the resulting polymer onto the gelatin backbone was carried out by adding 273 mg ammonium persulfate [(NH4)2S2O8] powder, then maintaining the subsequent reaction for 24 h. This resulting mixture was placed into a dialysis bag, with 12–14 kDA molecular weight cut-off, for 10 h in DI water at 50°C, then freeze-dried for 48–72 h. The freeze-dried PAMB-G was redissolved in 5 mL phosphate-buffered saline, adjusted for pH at a physiological range (∼7.4) with 10 M NaOH, followed by the addition of N-(3-dimethylaminopropyl)-N′-ethylcarbodiimide hydrochloride (EDC, 22980, Thermo Fisher) cross-linker and N-hydroxysuccinimide (NHS, 130672, Sigma-Aldrich) to form a hydrogel. Injectable gelatin hydrogel was prepared by crosslinking 20% aqueous gelatin solution with EDC in the presence of NHS.
After hydrogel formation, either gelatin or PAMB-G was injected into rats 1 day after inducing SCI. This injection occurred at four locations within the SCI site: two caudally and two dorsally from the injury site epicenter, and all four sites received, in total, 10 µL of gelatin or PAMB-G; control rats received saline injections instead. The injection rate was 0.5 µL/min, and at the end of injection, the needle was left in the cord for at least 1 min to allow diffusion from the injection site prior to closing the wound. Afterwards, all rats were placed in a bedding-free cage on top of a 37°C heating pad before being returned to their regular cages. Bladders for all rats were manually extruded twice daily until normal bladder function was restored.
Magnetic resonance imaging of the spinal cord
After SCI, the injured spinal cord was observed under magnetic resonance imaging (MRI) for up to 8 weeks, using a 7.0 T MRI scanner (Bruker, PharmaScan), with transmit volume and receive surface radiofrequency coils. The SCI site was scanned transversely, using T2w imaging, at 8 weeks postinjury to observe the gray-to-white matter ratio. Diffusion tensor imaging data were then processed and analyzed with MIPAV software.
Motor evoked potential and multielectrode array measurements
Motor evoked potentials (MEPs) were measured immediately after compression SCI and biomaterial injection, as well as at 2 and 8 weeks post-SCI. Rats were lightly anesthetized using isoflurane (<1%), and recordings were obtained from the hind-limb gastrocnemius muscle by inserting a pair of stainless steel subdermal needle electrodes, serving as the recording electrodes, into the muscle. A ground electrode was placed under the skin between these two electrodes. MEPs were induced using transcranial electrical stimulation, in which two monopolar needle electrodes were inserted subcutaneously over the skull. The tip of the electrode serving as the cathode was placed in such a way that it touched the bone at the motor cortex area, whereas the electrode serving as the anode was placed at the base of the nose. Rectangular pulse stimulation of 4 V, for 0.1 ms, at 0.2 Hz, was applied, and recordings were acquired by Powerlab (AD Instruments, Colorado Springs, Colorado). Amplitude was measured as the difference between positive and negative peaks, whereas latency was calculated as the time from the start of the stimulus to the appearance of the first prominent peak. Multielectrode array (MEA) measurements were then carried out in vitro for all three groups, in which 200 mV of stimulation was provided with a stimulator (STG 4002, Multichannel Systems Reutlingen, Germany), with a 1.5 × 1.5 mm recording area. Regional electrical field potential across the SCI scar area was then evaluated using Cardio 2D software.
Behavioral analyses using Basso–Beattie–Bresnahan and slope test
Bodily functional recovery, in the form of locomotion, among the three rat groups was first assessed by two independent observers using the 21-point Basso, Beattie, and Bresnahan (BBB) locomotor rating scale from 0 to 8 weeks after SCI. In this scale, 0 points represented the lack of any visible movement, and 21 points indicated the presence of a normal gait. 19 BBB was used to assess the extent of post-SCI hind-limb locomotor recovery, including joint movements, stepping ability, coordination, and trunk stability. Testing was conducted every week within a 60 cm × 61.5 cm × 40 cm demarcated space, in which the rats were allowed to freely explore their surroundings for a 4-min period. BBB scores were averaged across both right and left hind limbs to obtain a final motor recovery score for each week of testing.
In addition, the inclined plane test was carried out, in which SCI rats were placed on a gridded slope whose angle could be adjusted in 5° increments, up to a maximum of 90°. Starting from 0°, the angle of the slope was increased every 5°, and rats were observed after each increase to determine whether they could stay on for at least 5 s before slipping off. The maximum angle of the slope the rats were able to stay on before slipping was thus measured.
Statistical analyses
All data were expressed as mean ± standard deviation (SD) and analyzed by GraphPad Prism version 7.0 (San Diego, CA, USA). Comparisons between three or more groups were made using one-way analysis of variance (ANOVA) for single-factor variables or two-way ANOVA for two-factor variables with repeated measurements over time, followed by Tukey’s posthoc tests. p < 0.05 was considered statistically significant.
Results
PAMB-G delivery into injured rat spinal cord
We have previously characterized the rheological behavior of PAMB-G and gelatin and found, under reaction time sweep mode, that both hydrogels have gelation times of ∼5 min, which provides sufficient working time for their injection while preventing premature gelation inside the syringe.15,20 Furthermore, both hydrogels behaved like elastic solids, and their Young’s moduli showed that they had similar compression elasticities, close to that of native tissue (11.9–46.2 kPa). 21 Therefore, PAMB-G was considered to be a suitable biomaterial for tissue engineering applications.
In this study, PAMB-G or gelatin was injected into the injury site 1 day after compression SCI. Biochemical analyses were conducted on days 3 and 7 after injection, and examination of electrical and motor functions was performed weekly for up to 8 weeks (Fig. 1A). Figure 1B depicted, both in the schematic diagram and the actual photographs, that the materials were injected into four locations: two caudally and two dorsally from the injury site epicenter, for a total volume of 10 µL. Photographs of the SCI injury site and the injection of PAMB-G were shown in Figure 1C. Under MRI, it was also found to be visible with respect to SCI effect on the spinal cord white matter (Fig. 1D), as well as under sagittal view (Fig. 1E). This facilitated observations regarding the effects of the conductive biomaterial on spinal cord and subsequent bodily functional restoration.
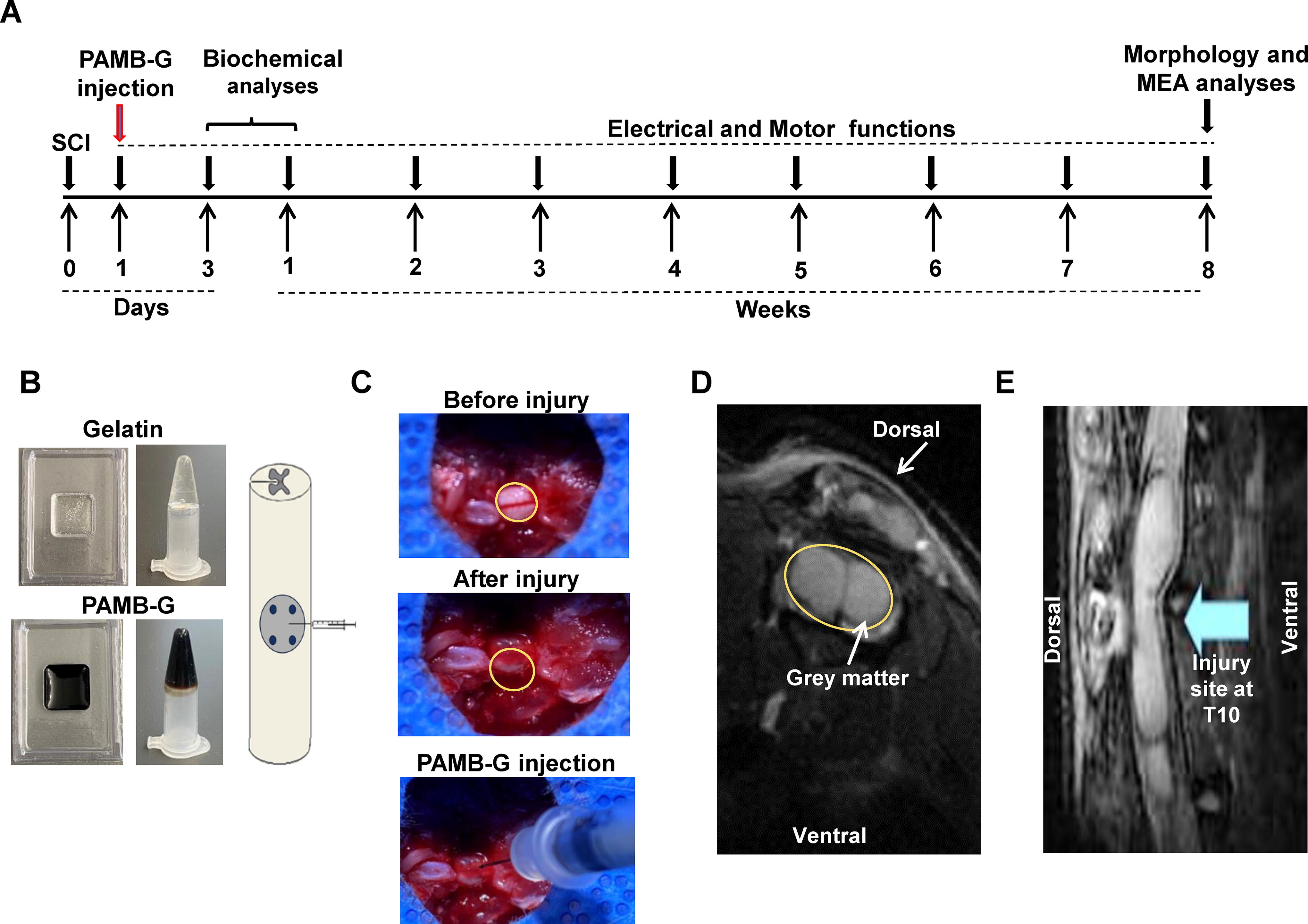
Delivery of poly-3-amino-4-methoxybenzoic acid-gelatin (PAMB-G) into injured rat spinal cord.
PAMB-G reduces SCI-associated astrogliosis and inflammatory reactions
Post-SCI inflammatory reactions and astrogliosis are major contributors to glial scar formation, which is an impediment against neuronal regeneration and thus CNS–PNS signaling restoration. To determine whether PAMB-G had any effect on modulating astrocyte reactivity and inflammatory responses post-SCI, we used real-time quantitative polymerase chain reaction and immunohistochemistry analyses to measure the expression levels of factors associated with inflammatory cells and reactive astrocytes. We found that expression levels for proinflammatory cytokines tumor necrosis factor (TNF)-α and monocyte chemoattractant protein (MCP)-1 were significantly lower in PAMB-G-injected rats compared to those controls at 3 days post-SCI (Fig. 2A–B), thereby indicating a reduction in post-SCI inflammatory responses. This lowered inflammatory response was also associated with lowered inflammatory cells and reactive astrocytes, in that ionized calcium-binding adaptor molecule 1 (IBA-1)+ and glial fibrillary acidic protein (GFAP)+ cell numbers, associated, respectively, with microglia and reactive astrocytes, were significantly fewer at 7 days post-SCI for PAMB-G than for the other two groups (Fig. 2C–F). All these findings suggest that PAMB-G reduced astrogliosis and inflammation, especially around the epicenter of the SCI site.
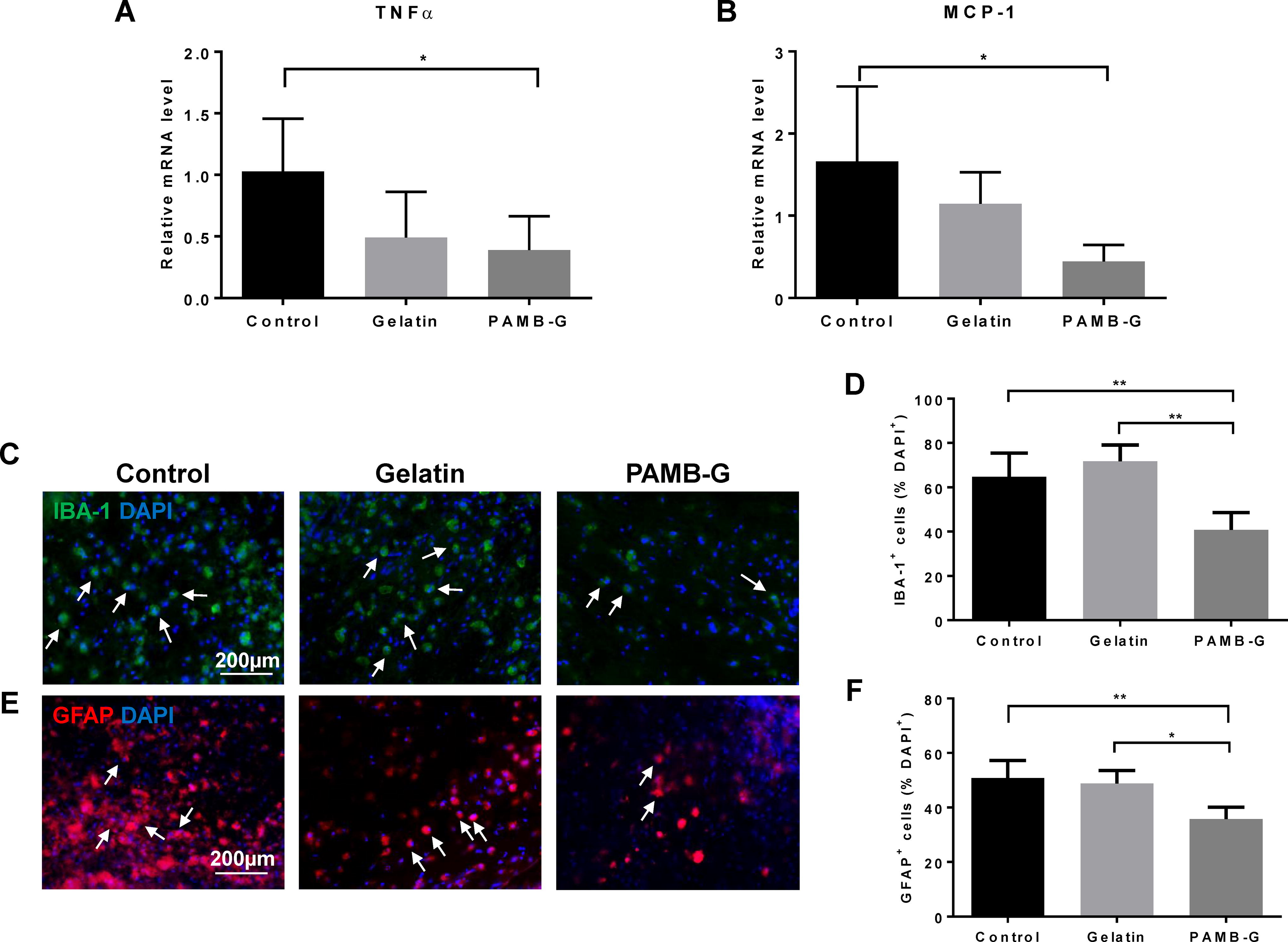
PAMB-G reduces injury-associated astrogliosis and inflammatory reactions. Real-time quantitative polymerase chain reaction measurements of mRNA expression for
PAMB-G reduces post-SCI neuron apoptosis and aids in neuronal tissue preservation
Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining was then used to determine short-term apoptotic occurrence at 1 week post-SCI. We found significantly fewer TUNEL+ cells in PAMB-G-injected rat spinal cords (35 ± 4.082% of DAPI), compared to those injected with either saline control (50.5 ± 4.203) or gelatin (56 ± 3.367; Fig. 3A–B). To determine whether PAMB-G was able to maintain the remaining neuronal tissue postinjury on a macro-scale, as well as for a long term, we performed histological analysis of the spinal cord tissue around the injury site at 8 weeks post-SCI. The cavity resulting from the SCI was the smallest in PAMB-G-injected spinal cords (0.239 ± 0.059%); it was filled with biomaterial and neuronal tissue. By contrast, a larger cavity, in the form of an “empty circle” within the spinal cord tissue section, was present among control (0.546 ± 0.156) and gelatin (0.605 ± 0.089) groups (Fig. 3C–D). This increased neuronal tissue presence in PAMB-G was also supported by a greater percentage of white matter being present in PAMB-G-injected spinal cords (0.523 ± 0.059%), compared to the other two groups under MRI (0.377 ± 0.044% in Control, 0.385 ± 0.051% in Gelatin), indicating the presence of a greater number of both intact and possible newly-grown axons (Fig. 3E–F). These observations thus indicate that PAMB-G reduces apoptosis onset during both short (1 week) and long (8 weeks) terms, enabling neuronal tissue maintenance post-SCI to facilitate possible regeneration and functional restoration.
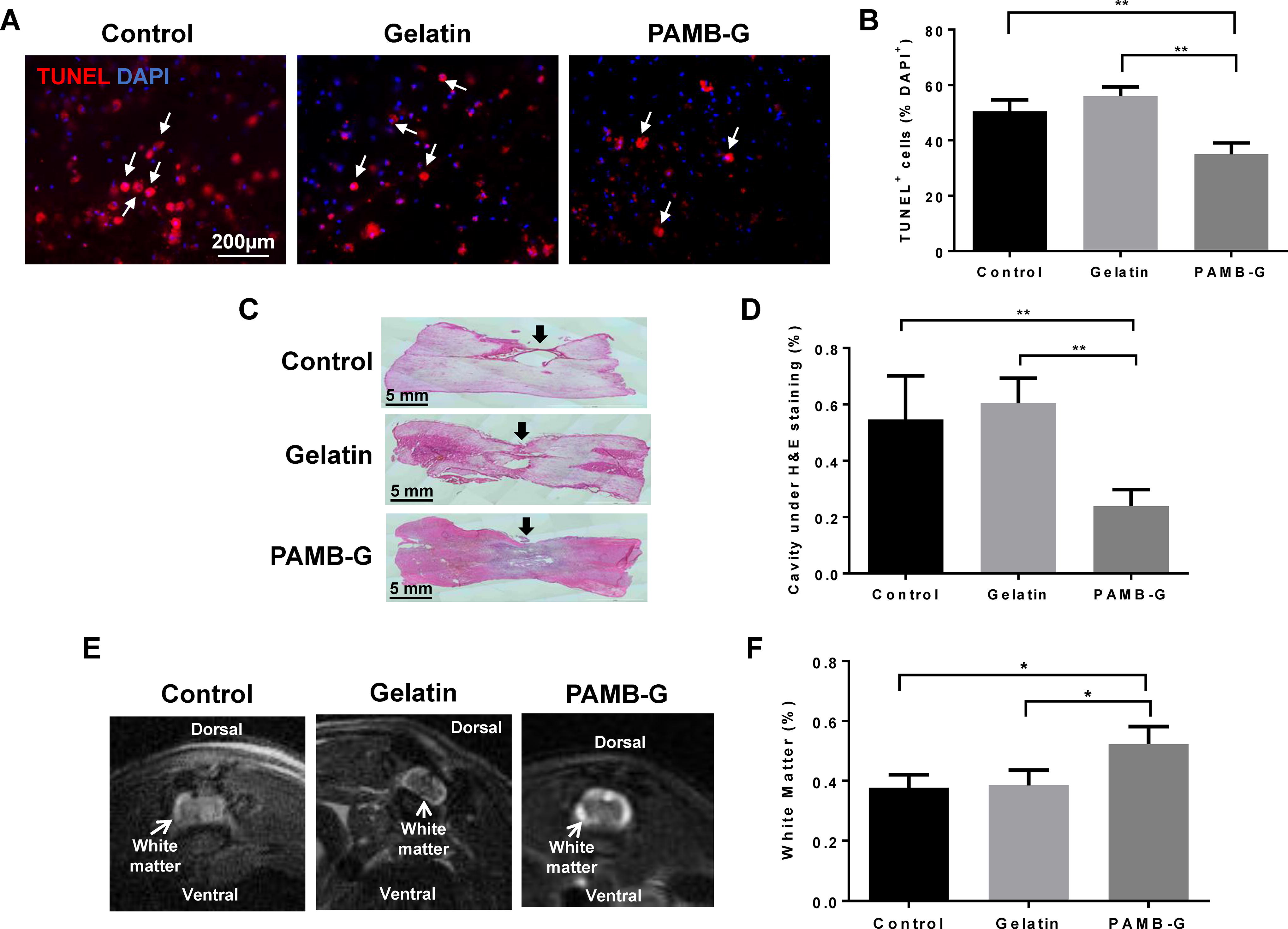
PAMB-G lowers apoptosis and aids in tissue preservation post-SCI.
PAMB-G upregulates neural growth factors and stimulates axonal regeneration
The presence of increased white matter proportions among PAMB-G-injected spinal cords suggested that those rats had increased axonal growth, compared to those without PAMB-G. To further investigate this possibility, we measured the expression levels of growth-associated protein 43 (GAP43) protein, which is expressed by neuronal growth cones during the axonal regeneration process and is associated with neuronal growth and synapse formation. Western blot analysis found that GAP43 levels at 3 days postinjury were significantly higher in the PAMB-G (1.455 ± 0.294, normalized to GAPDH) group, compared to those injected with either saline (0.770 ± 0.130) or gelatin (0.833 ± 0.327), indicating rapid upregulation of factors favoring neuronal regeneration (Fig. 4A–B). The presence of increased neuronal growth itself, on top of increased neuronal growth-associated markers, was also demonstrated by neurofilament 160 (NF160) immunostaining, where NF160+ cell counts across a distal cross-section were significantly higher for PAMB-G-injected spinal cords (1164 ± 143.285), compared to the other two groups (694 ± 167.091 for Control, 679.75 ± 105.427 for Gelatin; Fig. 4C–D). Taken together, all these evidences therefore indicate that PAMB-G fostered neuronal growth and regeneration post-SCI.
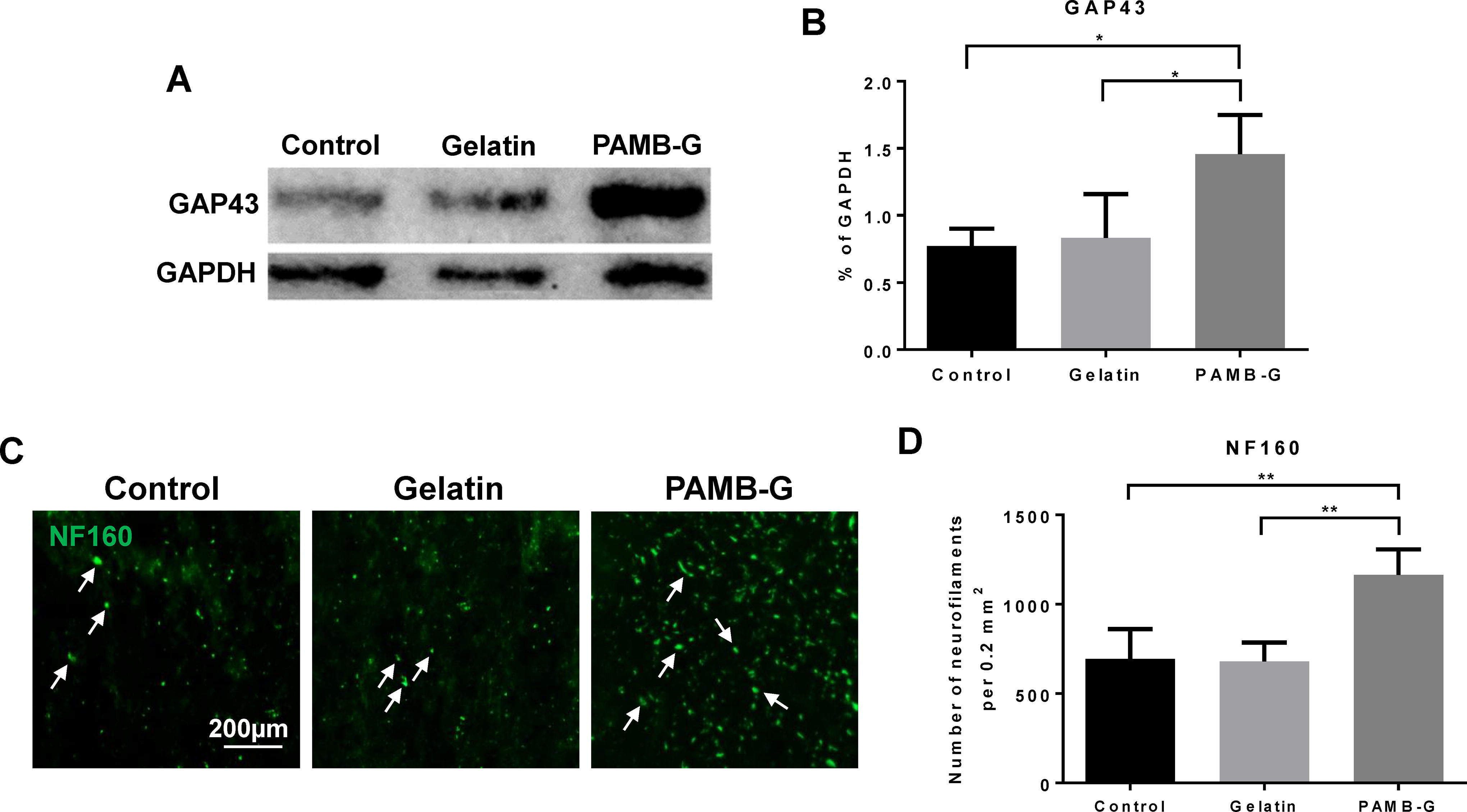
PAMB-G upregulates neural growth factors and stimulates axonal regeneration.
PAMB-G facilitates spinal cord electrical signaling restoration postinjury
Increased axonal regeneration observed for PAMB-G-injected rat spinal cords suggested that PAMB-G may also increase electrical signaling conduction post-SCI. To investigate this possibility, we first measured MEPs among all three groups. All groups demonstrated a biphasic deflection pattern, and PAMB-G-injected rats had the highest peak amplitude compared to those injected with saline control or gelatin from 2 to 8 weeks postinjury (Fig. 5A–B). More specifically, mean MEP amplitudes for PAMB-G were >3 times greater than Control or Gelatin at 2 weeks and ∼1.7–2.8 times greater than the other two groups at 8 weeks postinjury (Fig. 5B). Therefore, PAMB-G had the highest conduction capability, which was coupled with those rats having the shortest latency period, compared to control or gelatin (Fig. 5C). The mean MEP latency period decreased by ∼25–30% compared to Control and Gelatin groups at 2 and 8 weeks postinjury (Fig. 5C). Latency period length has been found to be inversely correlated with conduction velocity, where shorter lengths are associated with higher velocities, meaning that the PAMB-G group yielded the highest conduction capabilities and velocities post-SCI at the end of the 8-week period (Fig. 5C).
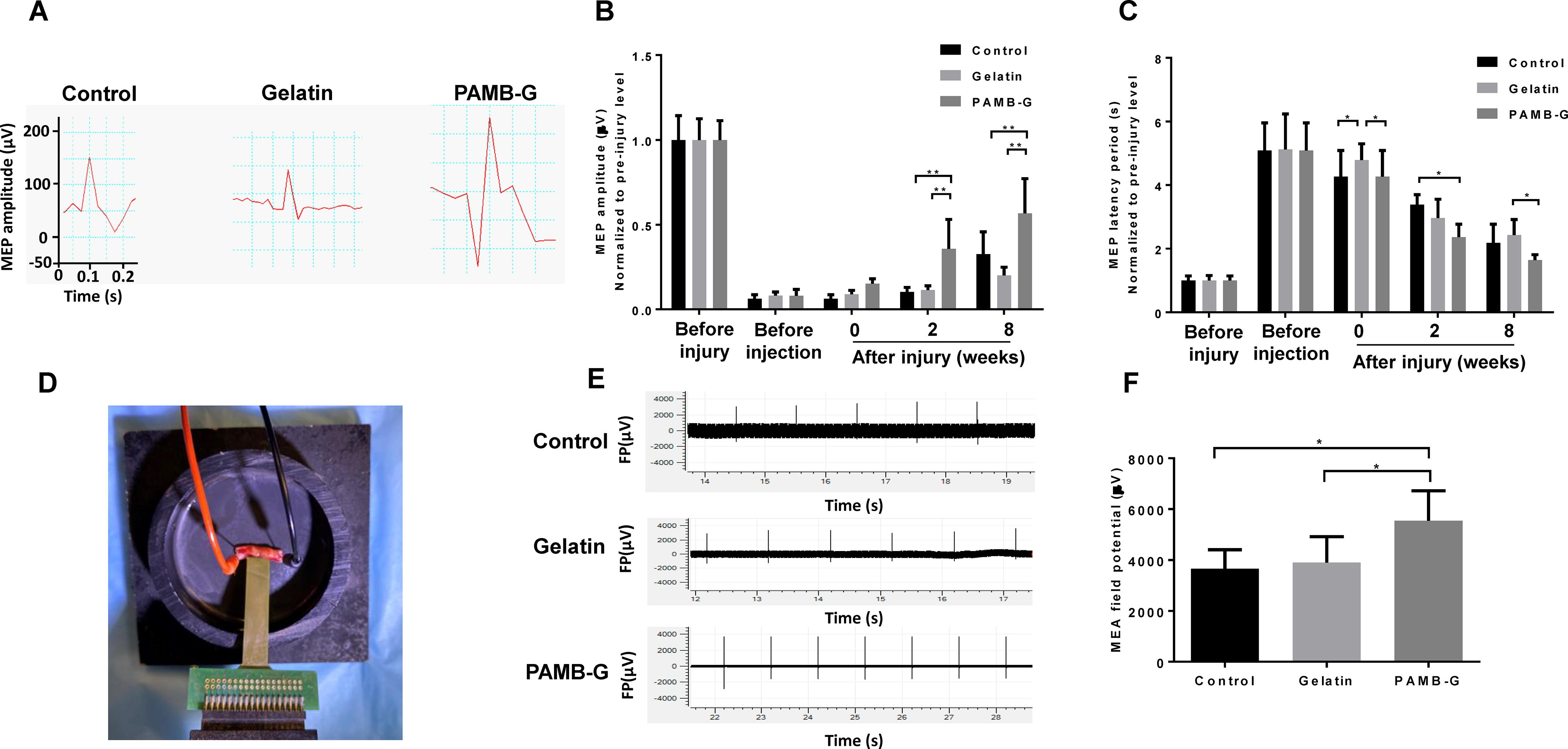
PAMB-G facilitates the restoration of spinal cord electrical signaling.
The flexible MEA apparatus was also used for field potential measurement across the spinal cord (Fig. 5D), in which PAMB-G has significantly higher post-SCI field potentials compared to the other 2 groups (∼1.4–1.5-fold increase compared to Control/Gelatin; Fig. 5E–F). All these evidences support the idea that PAMB-G conductive properties, along with its stimulation of axonal regeneration from remaining intact neurons, facilitates electrical reconnection across the SCI-caused lesion, allowing conduction to be restored throughout the spinal cord and between CNS and PNS, for possibly regaining bodily functional control.
PAMB-G improves the execution of postinjury motor functions
To examine PAMB-G effects on rat motor functions, a number of behavioral assays were utilized. The extent of rat hind-limb locomotive behavior was judged according to the 21-point BBB open-field locomotor scoring system, and average scores for the three groups were taken weekly over the course of 8 weeks postinjury. Under BBB, PAMB-G had the highest degree of hind-limb locomotive behavioral recovery over the 8-week period, reaching a score of ∼15/21 points, compared to ∼10/21 for the other two groups (Fig. 6A). Furthermore, the inclined plane test showed that at the end of the 8-week period, PAMB-G-injected rats were able to stay on the plane at significantly steeper angles (∼77°), compared to control (∼59°) and gelatin-injected (∼58°) rats (Fig. 6B). This CNS-PNS connectivity restoration, via PAMB-G serving as a spinal cord reconnector, was further confirmed by gastrocnemius muscle wet weight measurements. It has been previously noted that the gastrocnemius muscle loses weight after SCI, owing to atrophy associated with the loss of electrical signaling from the CNS, and subsequently, the PNS. Based on these findings, we observed that PAMB-G had significantly higher gastrocnemius muscle-to-tibia length ratios (∼1.4–1.6-fold increase), as well as larger cross-sectional areas under hematoxylin & eosin staining, compared to the other two groups (Fig. 6C–D), indicating that PAMB-G aids in alleviating post-SCI muscle atrophy by restoring electrical signaling from the CNS to the PNS, and in turn, PNS-controlled muscles like the gastrocnemius.
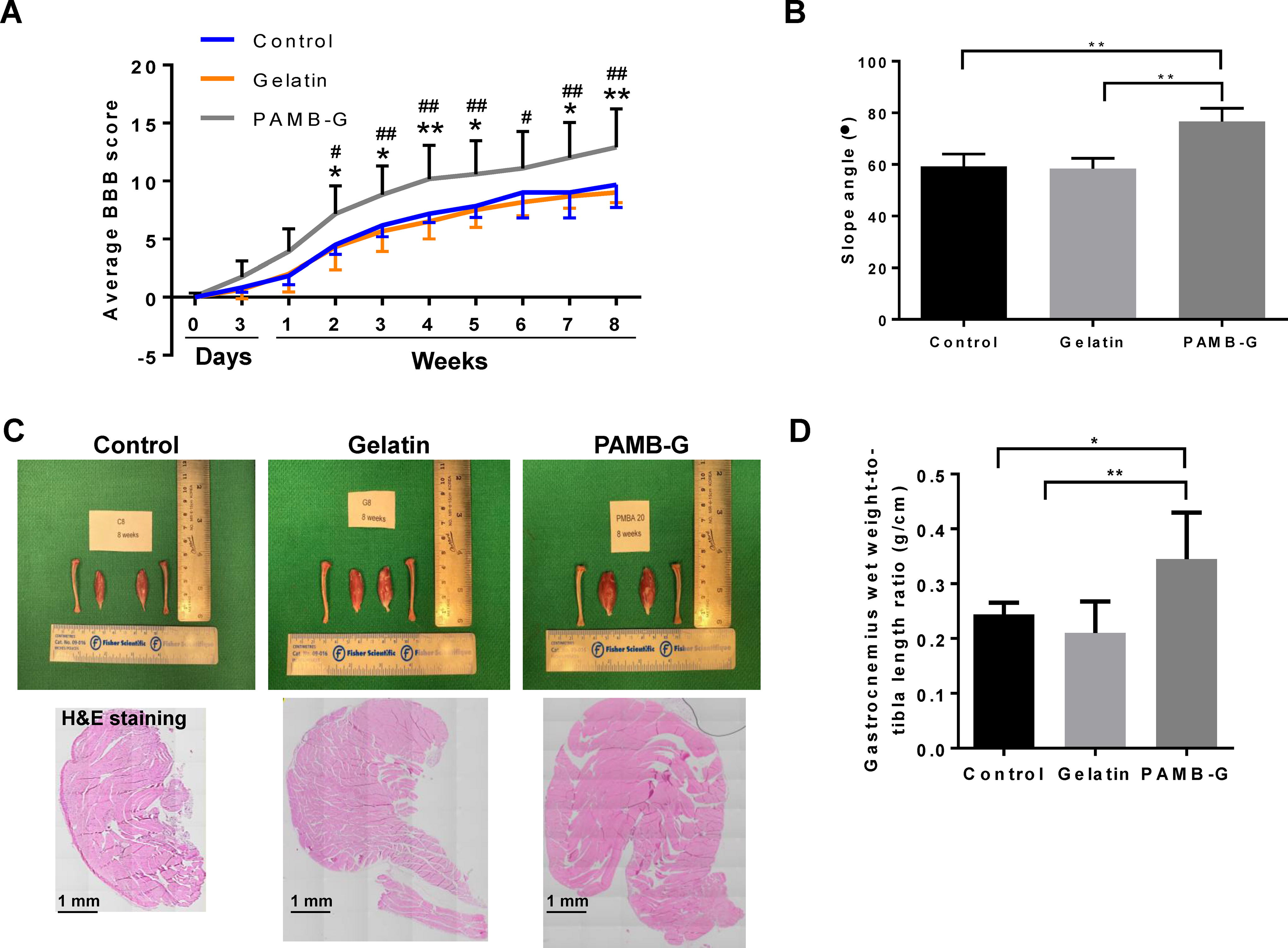
PAMB-G improves motor functions post-SCI.
Discussion
SCI is currently an incurable, permanent condition, owing to the resulting inability of the patient to control bodily functions associated with regions below the injury site. This lack of control is due to the initial physical force, such as shearing, compression, and laceration, damaging the spinal cord neural tissue, as well as associated blood vessels, at the injury site. 22 This primary damage is then followed by ischemia, along with inflammatory and proapoptotic processes, such as inflammatory cytokine and reactive oxygen species production, as well as loss of calcium homeostasis, all of which contribute to astrocyte hypertrophy and neuronal cell death. 22 As a result of the hypertrophic astrocytes and neuron loss, glial scar tissue forms at the injury site, blocking electrical conduction between the remaining intact portions of the spinal cord and leading to the lack of bodily function control below the scar.22,23 In this study, we have developed a method to counteract against these damages, via the usage of a biomaterial, PAMB-G. Its injection into the SCI site reduced post-SCI inflammatory reactions, in the form of lowered astrogliosis, microgliosis, and proinflammatory cytokine levels, such as TNF-α. This lowered inflammation was coupled with reduced neuronal apoptosis, both of which yielded increased preservation of remaining intact spinal cord tissue. In addition, PAMB-G stimulated greater axonal and synapse regeneration. These regenerative processes most likely stem from the conductive properties of PAMB, allowing it to serve as a reconnector across the SCI site, in turn enabling PAMB-G to restore electrical signaling conduction throughout the spinal cord and subsequently the execution of bodily functions, particularly with respect to locomotion.
This “bridging” of the glial scar is predominantly the focus for biomaterial-based approaches, both with and without associated stem cells meant to replace the dead ones within the glial scar site. Biomaterials consist of a 3D polymer “scaffold,” with materials ranging from conductive electro-spun nanofibers like polylactic acid, or hydrogels comprised from the likes of hydroxyethyl methacrylate [2-(methacryloyloxy)ethyl] trimethylammonium chloride and hyaluronic acid-methylcellulose.3,24 Often, these “scaffolds” have additional molecules, such as growth factors or molecules, to impart specific properties of interest for aiding in neuronal regeneration. Growth factors, such as basic fibroblast growth factor or PDGF, encourage the growth of neurons via their activation of pathways related to axonal growth postinjury or directing the differentiation of associated stem cells on the biomaterials. The latter, in particular, has been extensively examined in various studies, such as one involving the incorporation of small molecules within a collagen scaffold that encouraged differentiation of endogenous parenchymal neural stem cells into new neurons.25,26 An alternative approach is the application of conductive biomaterials, which not only aids in restoring proper electrical conduction between the CNS and PNS via providing a channel for communication between the remaining intact spinal cord portions but also stimulates axonal growth.11,27–30 The ability of electrical signaling to stimulate such growth has been demonstrated in which the implantation of a PLA/PPy conductive biomaterial scaffold was associated with bodily function recovery and axonal regeneration. 11 In line with those findings, we have developed a conductive biomaterial, PAMB-G, that is also able to stimulate neural growth, reduce the occurrence of inflammation-associated tissue damage, and restore proper electrical conduction post-SCI.
We have shown in a previous study that PAMB-G is a biocompatible conductive biopolymer, able to restore proper electrical signal conduction, and therefore synchronized contraction, within infarcted cardiac tissue in a rat model. 15 These properties are owed to the AMB monomeric units consisting of alternating π and σ bonds, which allow free flow of electrons between the individual atoms within those molecules. Such an arrangement has also been found for other conductive biomaterials, such as PEDOT and polyaniline.31,32 PAMB-G, though, is notable for its self-doping properties to ensure electrical conduction under physiological pH levels, making it highly suited to serve as a channel for electrical signaling conduction in vivo, unlike other biomaterials that are otherwise nonconductive within physiological pH. 15 We also showed that PAMB-G is highly durable, lasting up to a year postimplantation in vivo. 33 Based on those properties and findings, we examined its ability in restoring electrical conduction between intact spinal cord portions, the CNS and PNS, and subsequently bodily functional control. Our current findings have shown that PAMB-G indeed served as a connector between the intact spinal cord sections, conducting signals throughout the organ and facilitating communication between the CNS and PNS. This restoration of signal conduction was reflected by improvements in bodily function control below the SCI site, particularly in regard to lower body locomotion. These improvements are owed to both the biomaterial scaffolding serving as “artificial axons” between the remaining living neurons on both sides of the injury site, as well as being able to conduct electrical signals to stimulate new axonal growth postinjury.
The lowered occurrence of inflammation post-PAMB-G injection could also be attributed to the combination of PAMB and gelatin, as studies have shown that fish gelatin hydrolysates, such as from milkfish and skipjack tuna, are able to quench free radicals because of the presence of unique Gly-Pro-Y and Gly-X-Hyp motifs.34,35 Despite the uniqueness of those motifs in fish gelatin hydrolysates, a study has demonstrated that bovine collagen peptides, which are able to be derived from gelatin, itself utilized as part of the PAMB-G hydrogel, also possess the ability to quench free radicals, 36 further supporting the possibility that the lowered neural tissue inflammation post-PAMB-G injection at the injury site could be owed to gelatin degradation by-products. However, the specific mechanism is less well-understood, though that study indicated that the quenching ability is dependent on temperature, the type of enzyme digesting the gelatin, and the origin of the bovine gelatin, such as tibia versus femur. 37 It is worth noting, though, that our results showed the combination of gelatin with PAMB was more effective in lowering inflammation, compared to gelatin alone. This could be owed to some sort of synergistic effect. Therefore, future studies will account for these factors, such as obtaining in vivo cerebrospinal and body fluid samples from the rat model to investigate the presence and composition of gelatin hydrolysates, as well as gelatin-digesting enzymes and PAMB chains.
Overall, this study demonstrated that PAMB-G can be used to aid in restoring spinal cord electrical conduction, and thus bodily function control, post-SCI. This restoration is via its conductive properties providing channels allowing electrical signals to be transported between remaining intact neurons, as well as stimulating new axonal growth from those neurons upon electrical stimulation, which is demonstrated by a ∼1.4-fold increase in mean white matter percentage among PAMB-G, compared to both Control and Gelatin groups. In particular, PAMB is responsible for the electrical conduction, and the conjugation with gelatin also provides anti-inflammatory attributes against post-SCI secondary inflammatory cascades. These anti-inflammatory activities result in lowered astrocyte (by ∼30%) and microglial activation (by ∼40–45%), as well as apoptosis (by ∼30–40%), among the remaining neuronal tissue at the SCI site. The anti-inflammatory, proregenerative, and electrical conduction capabilities of PAMB-G thus facilitate its usage as a possible treatment approach for counteracting against SCI-related bodily functional deficits.
Footnotes
Authors’ Contributions
YZ: investigation, data curation, methodology, formal analysis, writing-original draft; AY: visualization, validation, resources, writing-original draft; JW: investigation, data curation, methodology, formal analysis; SL: methodology, visualization, validation, writing-review & editing; MY and ZP: investigation, data curation, methodology; HS: conceptualization, methodology, writing-review & editing; BJ: conceptualization, funding acquisition, supervision, writing-review & editing; RL: conceptualization, project administration, funding acquisition, supervision, writing-review & editing.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Heart and Stroke Research Foundation of Canada (
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.