
Abstract
Recent progress in engineering microvascular networks in vitro and in vivo offers exciting opportunities to create tissue constructs with preformed blood vessels, which are rapidly blood perfused by developing interconnections to the preexisting blood vessels of the host tissue after implantation. This process, termed as inosculation, is well known from the revascularization of various tissue grafts, such as transplanted skin, nerves, or bone. It is characterized by the close interaction of the implant's preformed microvascular network and the host microvasculature. The sprouting angiogenic activity of both counterparts determines whether inosculation takes place internally within the implant or externally within the surrounding host tissue. Successful inosculation involves vascular remodeling as well as infiltration of inflammatory cells and stem cells. With the use of sophisticated in vitro and in vivo models, more detailed analysis of regulatory mechanisms of inosculation will help to develop novel strategies, aiming at further accelerating the establishment of a life-sustaining blood supply to implanted tissue constructs.
Introduction
During the last years, several strategies have been proposed to stimulate the vascularization of implanted tissue substitutes, which are engineered in vitro by seeding cells within three-dimensional scaffolds. 1 First, the chemical composition of biomaterials for scaffold fabrication has been modified.2–4 Second, sophisticated techniques, such as computational topology design and solid free-form fabrication, have been developed to create scaffolds of mechanical stability with precise porous architecture for the optimization of blood vessel ingrowth.5,6 Third, filamentous scaffolds have been designed and bioactive ligands have been micropatterned on the scaffold surface to induce and regulate in vivo–guided angiogenesis.7,8 Fourth, scaffolds have been bioactivated by incorporation of a protein delivery system. This allows for a localized and controlled release of pro-angiogenic growth factors, supporting the initiation of both blood vessel development and maturation.9–12 However, endothelial cell migration and physiological growth of new blood vessels has been demonstrated not to be faster than ∼5 μm/h.13,14 Because the amount of oxygen, which is required for cell survival, is limited to a diffusion distance of only about 150–200 μm from a supplying blood vessel,15,16 all these strategies will therefore not be able to prevent cell death in the center of larger three-dimensional tissue constructs during the very first days after implantation.
Based on these considerations, the strategy of engineering preformed microvascular networks within tissue constructs prior to their implantation is emerging in the field of tissue engineering. 17 This may be achieved in vitro by seeding scaffolds with combinations of endothelial cells, stem cells, and mural cells, including pericytes, smooth muscle cells, and fibroblasts.18–22 On the other hand, the body's capacity to act as a bioreactor may be used to create preformed microvascular networks in vivo by ingrowth of capillaries from well-vascularized sites into implanted scaffolds or by preparation of an arteriovascular loop placed in a protected space.23–25 The approach of prevascularization bears the major advantage that the preformed microvascular network simply has to anastomose to the existing blood vessels of the host tissue after implantation, resulting in a rapid and complete construct vascularization.25–27 This process, which is termed inosculation, is independent of tissue thickness and, thus, may offer the possibility of drastically accelerating and improving vascularization of large three-dimensional tissue constructs, paving the way for a broad applicability of engineered tissues and organs in regenerative medicine.
Revival of a Well-Known Phenomenon
Inosculation is a well-known phenomenon in plastic surgery. More than 100 years ago, clinical reports already suggested that the revascularization of skin grafts occurs due to a direct connection between the host microvasculature and the grafts' blood vessels.28,29 This idea was later on supported by intravenous injection studies with radioisotopes and India ink, demonstrating that blood perfusion is established in the skin grafts' vasculature as early as 2–4 days after transplantation.30,31 In addition, Haller and Billingham 32 showed that the blood vessel pattern in a grafted revascularized hamster cheek pouch is identical to that before the transfer. On the other hand, Zarem et al. 33 proposed that full-thickness skin grafts primarily vascularize by ingrowth of new blood vessels originating from the recipient's microvasculature. By now it is well accepted that skin graft revascularization includes both processes, inosculation of preformed blood vessels and de novo vascular ingrowth, which occur in a time-dependent pattern after transplantation. After an initial imbibition phase, during which the graft survives by diffusion, early revascularization is initiated by anastomosis of graft and host vessels, while after several days newly formed capillaries from the recipient's bed additionally begin to invade the graft.34–38
Besides skin grafts, inosculation has also been described for other transplanted tissue types. Best et al. 39 demonstrated in a 20-mm sciatic nerve graft model in rats that epineural perfusion is consistently established by 48 h and endoneural perfusion by 72 h. Interestingly, the latter was present in an all-or-none-pattern, strongly suggesting that the primary mode of nerve revascularization is longitudinal inosculation. Comparable conclusions were drawn by Chalfoun et al., 40 who directly showed by means of orthogonal polarization spectral imaging that revascularization of primary nerve grafts occurs via bidirectional inosculation. In another study of Albrektsson, 41 analyzing the vascularization of rabbit bone grafts in a titanium chamber, it was found that the grafts' microcirculation was mainly regained via ingrowth of newly formed blood vessels at days 5–8 after transplantation. However, in two grafts, blood vessels observed before grafting were also found perfused afterward, indicating end-to-end anastomoses between host and preexisting vessels. Rothenfluh et al. 42 transplanted the metatarsals of fetal DDY-mice, known to be nonvascularized at day 16 of gestation (M16) but vascularized at day 18 (M18), into the dorsal skinfold chamber of adult DDY-mice. By means of intravital fluorescence microscopy, they could demonstrate that osteochondral grafts of M18 vascularize much faster than those of M16. They concluded that the preexisting blood vessels of M18 osteochondral grafts had inosculated with the recipient vessels.
The fast kinetics, observed in the vascularization process of different prevascularized tissue grafts, have contributed to the introduction of the inosculation phenomenon in modern tissue engineering as a promising strategy, which may markedly improve the survival rates of implanted tissue constructs. In fact, Tremblay et al. 27 demonstrated for the first time the successful inosculation between normal human capillaries reconstructed in vitro within a tissue-engineered skin graft and the wound bed vasculature in less than 4 days. Shepherd et al. 26 established a technique to generate three-dimensional microvascular beds in vitro by angiogenesis from isolated, intact microvessel fragments that retain endothelial and perivascular cells. After grafting these prevascularized contructs to the epicardial surface in a mouse model of acute myocardial infarction, inosculation of the grafts' preformed microvessels with the host coronary microcirculation occurred 7 days after transplantation. This resulted in a smaller infarct size and an improved left ventricular function when compared to avascular control grafts. 43
Apart from engineering of an artificial microvascular network in vitro, microvascular networks can also be preformed in vivo. For this purpose, we recently implanted poly-lactic-glycolic acid scaffolds into the flank of mice for 20 days. 25 By this, the body itself generated a mature microvascular network within the scaffolds through blood vessel ingrowth from the host site. This network consisted of functional, blood-perfused microvessels. After implantation into the dorsal skinfold of recipient animals, the in situ–preformed blood vessels rapidly reperfused by forming interconnections to the host microvasculature. Importantly, reperfusion could avoid apoptotic cell death within the scaffolds, which further underlines the importance of rapid vascularization for the survival and long-term function of implanted tissue engineering constructs.
Molecular and Cellular Mechanisms of Inosculation
A major precondition for the vascularization of an implanted tissue construct by inosculation is the activation of the host tissue microvasculature to form capillary sprouts, growing toward the preformed microvessels of the construct. This first step in the process of inosculation may be initiated by several stimuli. The implantation procedure may be associated with tissue damage, which induces the classical wound-healing cascade, characterized by the release of numerous pro-angiogenic growth factors, including vascular endothelial growth factor (VEGF), fibroblast growth factor, epidermal growth factor, platelet-derived growth factor, and transforming growth factor-beta.44,45 On the other hand, the implanted tissue construct itself may induce an angiogenic host tissue response. 3 Tissue constructs are created by seeding isolated cells on scaffold biomaterials. These biomaterials can induce a more or less strong foreign body reaction after implantation into the host. This reaction is characterized by the immediate formation of a layer of host proteins on the biomaterial surface. 46 Macrophages interact with these spontaneously adsorbed proteins and respond by liberation of a variety of cytokines, which themselves activate other inflammatory cell types like T cells, mast cells, and polymorphonuclear granulocytes. 46 Upon their activation, these cells release inflammatory mediators such as tumor necrosis factor-alpha (TNF-α), stimulating the ingrowth of new blood vessels from the implant site.47–49 Besides, the cells within the tissue construct directly stimulate the angiogenic process, because they suffer from hypoxia during the initial phase after implantation into the host. This results in upregulation of hypoxia-inducible factor-1α and hypoxia-inducible factor-1α–mediated overexpression of VEGF. 50 In fact, this molecular induction of capillary sprouting and inosculation can additionally be stimulated by bioactivating the tissue constructs in vitro through the incorporation of different growth factor delivery systems. Those systems have been shown capable of enhancing the development of new blood vessels after implantation of the bioengineered construct into the host tissue.9–12
Once the concentration of pro-angiogenic growth factors increases at the implantation site, sprouting angiogenesis occurs. This is characterized by the coordinated sequence of different humoral and cellular interactions.51–53 A major prerequisite for this process is that mature blood vessels first have to become destabilized by loosening interendothelial cell contacts and relieving periendothelial cell support to enable the migration of endothelial cells into the surrounding tissue. This initial step is mediated by angiopoietin-2 (Ang-2), which becomes strongly upregulated in the stimulated endothelial cells. Ang-2 antagonizes the Ang-1–mediated Tie-2 activation, resulting in the switch from the resting quiescent state of the endothelium into the activated responsive state. 54 In consequence, the basement membrane of the blood vessel is degraded by release of matrix metalloproteinases, allowing the endothelial cells to form vascular sprouts. 55
The growth of these sprouts toward the connecting ends of preformed microvessels within implanted tissue constructs represents the crucial step in the inosculation process, determining how fast blood perfusion will be established after implantation. During the last years, sophisticated studies have analyzed the mechanisms of guided angiogenic sprouting in the developing mouse central nervous system.56–59 They have shown that in response to VEGF-A, specialized endothelial cells at the tips of vascular sprouts extend multiple filopodia, mediated by VEGF receptor-2. 58 VEGF-A gradients guide these filopodia, leading to directed migration of the tip cells. These are followed by many stalk cells, which proliferate upon stimulation through VEGF-A-VEGF receptor-2-signaling, resulting in the elongation of the vascular sprout. 58 Interestingly, inhibition of delta-like ligand 4 (Dll4)–Notch1 signaling, which is mediated by VEGF-induced upregulation of Dll4, promotes increased numbers of tip cells, while activation of Notch reduces tip cells and vessel branches. Thus, Dll4–Notch1 interaction crucially determines the sprouting and branching patterns during angiogenesis.60–63 In addition, Phng et al. 64 recently demonstrated that the Notch-regulated ankyrin repeat protein acts as a molecular link between Notch- and Lef1-dependent Wnt signaling in endothelial cells to control the stability of new vessel connections. Their results suggest that the balance between Notch and Wnt signaling determines whether to make or break new vessel connections. Accordingly, pharmacological manipulation of these pathways may offer new possibilities in the future to increase the density of developing sprouts at the implantation site of prevascularized tissue constructs. This may enhance the probability for rapidly developing interconnections of the grafts' preformed blood vessels with the preexisting host microvasculature.
Role of Preformed Microvascular Networks in the Vascularization Process
Inosculation is characterized by a coordinated interaction between the implanted preformed microvascular network and the microvasculature of the host site. The fate of graft and host endothelial cells during this process is still a matter of discussion. In an early study, Zarem et al. 33 suggested that the vessels within skin grafts merely provide a conduit for ingrowing wound bed vessels. This hypothesis was based on the observation that graft vessels were empty of cells directly after implantation, but subsequently became infiltrated with leukocytes. The leading front of ingrowing blood vessels from the host site appeared to use these white blood cell–filled channels as conduit. However, at this time, it was not possible to differentiate between graft and host vessels. This could be achieved later on by Demarchez et al. 65 by transplanting human skin on athymic nude mice. Using a crossreacting anti-factor-VIII antigen antibody for the staining of endothelial cells and human-specific monoclonal antibodies directed against vimentin, human leukocyte antigen-DR (HLA-DR) and anti-type-IV-collagen, they could demonstrate by double-labeling immunofluorescence microscopy that after inosculation human graft endothelial cells disappeared and host endothelial cells of mouse origin migrated into the graft over the basement membrane of the preexisting human graft vessels. Comparable results were reported by Young et al., 35 who used the technique of in situ hybridization with fluorescent species-specific DNA probes to distinguish between human and mouse endothelial cells. Capla et al. 66 transferred full-thickness skin grafts between wild-type FVB/N mice and transgenic tie2/lacZ mice, where lacZ expression is controlled by the endothelial-specific tie2 promotor. They found that over the course of 7–14 days, blood vessels from the host site invade the graft periphery along preexisting channels, which resulted from the regression of the native graft vasculature. In more central regions of the graft, however, interconnection between graft and host vessels occurred, finally restoring complete blood flow in the graft vasculature by day 21. Interestingly, reoxygenation of the graft stopped the regression of the donor-derived blood vessels in the graft center, which survived for up to 120 days posttransplantation.
These results suggest a more passive role of the preformed microvascular network in the process of revascularization, primarily providing a conduit for the ingrowth of new blood vessels. Accordingly, inosculation takes place inside the prevascularized graft due to the regression of the preformed microvessels and invasion of microvessels of host origin (Fig. 1). In contrast to this mechanism of internal inosculation, recent studies provide evidence that the preformed microvascular network actively contributes to the process of graft revascularization. In fact, by implanting in situ–prevascularized poly-lactic-glycolic acid scaffolds from green fluorescent protein (GFP)–transgenic mice into the dorsal skinfold chambers of wild-type recipient mice, intravital fluorescence microscopy could show that GFP-positive vessels grew out from the scaffolds into the surrounding host tissue. 25 There, “external inosculation,” that is, interconnection between GFP-positive blood vessels with the host's GFP-negative blood vessels, could be observed (Figs. 1 and 2). Moreover, histological analyses revealed that the endothelial cells of preformed blood vessels were not replaced over time by host endothelial cells. Instead, they contributed to sprouting angiogenesis, further increasing the microvessel density also in the center of the implanted scaffolds. 25 In line with these findings, Shepherd et al. 26 reported that after implantation of microvascular constructs, consisting of a network of new microvessels in a collagen I gel, inosculation with the host circulation is restricted to the construct boundaries. Within the constructs, only a few superficially located host vessels were observed. Of interest, α-actin–positive perivascular cell coverage of the preformed blood vessels increased after implantation, indicating that they even were able to differentiate and maturate. This maturation process may be supported by invasion of perivascular cells from the host tissue.67,68
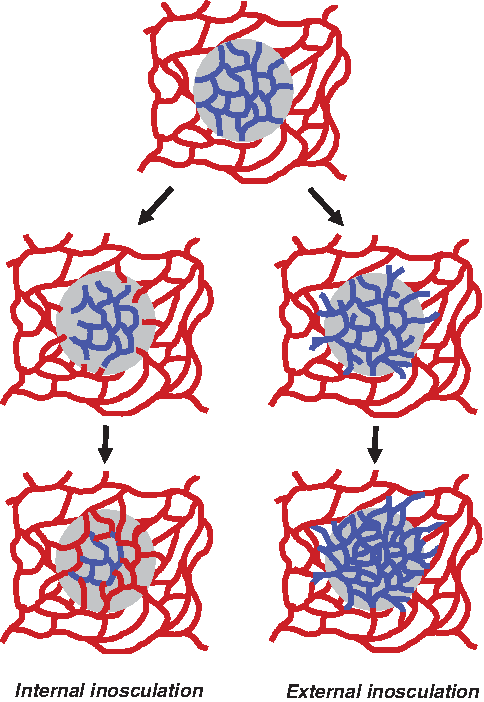
Schematic drawing of the process of internal and external inosculation. After transplantation of a prevascularized (blue network) graft onto the host microvasculature (red network), internal inosculation takes place inside the prevascularized graft due to the regression of the preformed microvessels and invasion of microvessels of host origin along the previously patterned graft vascular channels in the direction of the ischemic stimulus. Successful inosculation results in the establishment of blood perfusion and oxygenation of the graft tissue. In the absence of hypoxia, sprouting angiogenesis ends, and the central portion of the preformed microvascular network persists. On the other hand, high angiogenic activity of the graft's prevascularized network may also result in outgrowth of preformed blood vessels from the graft into the surrounding host tissue, where external inosculation occurs. In parallel, endothelial cells of preformed blood vessels contribute to sprouting angiogenesis in the center of the graft, further increasing the original microvessel density of the preformed microvascular network.
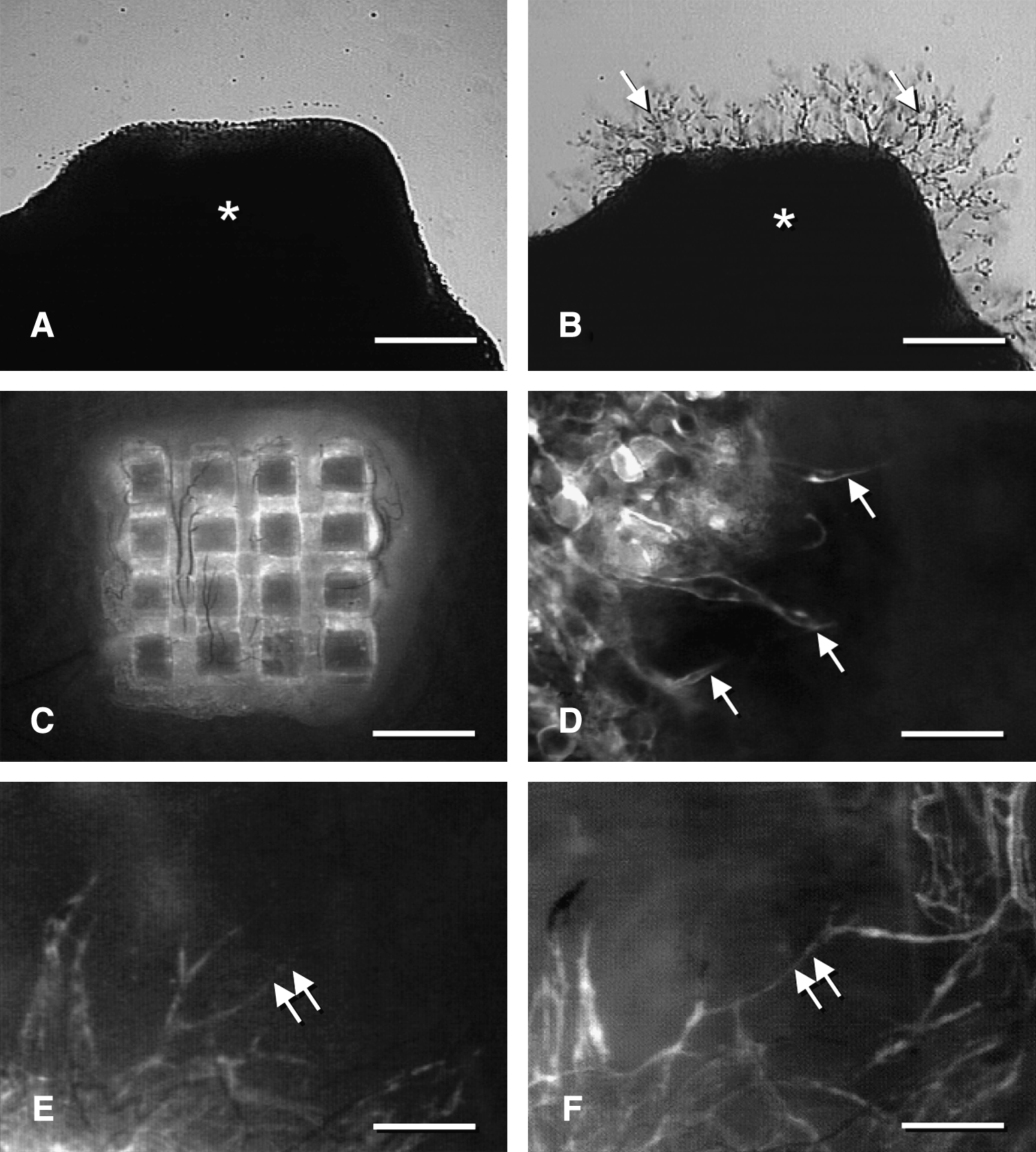
In vitro and in vivo detection of external inosculation according to the study of Laschke et al.
25
(
Taken together, these contradictory findings demonstrate that the role of preformed microvascular networks in the vascularization process of tissue constructs is not yet fully understood. However, there is clear evidence that preformed vessels constitute more than a framework for the ingrowth of new blood vessels. Under certain conditions they may also be the origin for blood vessel outgrowth, promoting external inosculation in the surrounding host tissue. This process may in particular be observed when the angiogenic activity of the preformed microvascular network is high enough to induce the development of new blood vessels before angiogenesis is initialized at the host site. Thus, it is critically dependent on the exact in vivo environment of the graft–host combination, which is determined by many factors, such as the extent of tissue hypoxia and the maturation stage of preformed blood vessels within the tissue construct as well as the tissue type and degree of the inflammatory reaction to the implant at the host site. Accordingly, it is not surprising that the aforementioned studies suggest different roles of preformed microvascular networks in the vascularization process of engrafted tissue-engineered substitutes. Future studies have to clarify now whether the stimulation of external inosculation may represent an additional strategy to further accelerate the restoration of blood perfusion in prevascularized implants.
Consequences of Successful Vascular Inosculation
Successful inosculation to the host circulation has distinct consequences for the preformed microvascular network within an implanted tissue construct. The restoration of blood perfusion induces vascular remodeling, that is, reorganization of the preformed microvascular network to achieve tissue homeostasis and to maintain a fully functional construct. 17 This may be associated with the development of new blood vessels in regions, where blood vessels are exposed to high shear stress.69,70 Angiogenesis is additionally induced by hypoxia of individual cells.71–74 On the other hand, preformed vessels may regress after inosculation due to the adjustment of vascular density to meet the nutritional requirements of the cells within the construct. The mechanisms, which induce regression, are still under investigation. Presumably, upon the onset of blood flow in unnecessary preformed microvessels, hyperoxia will suppress VEGF production, which has been shown to be critical for the survival of blood vessels by inhibition of endothelial cell apoptosis.75–77 Besides, immature preformed blood vessels, which lack vessel wall–stabilizing pericytes, may regress due to endothelial hyperplasia with vascular dysfunction and subsequent obliteration.78–80
Inosculation will not only abruptly change microhemodynamics and oxygen distribution within a prevascularized tissue construct but also will open the door for infiltrating leukocytes and other inflammatory cell types. The recruitment of circulating leukocytes into the tissue is crucially dependent on their interaction with the microvascular endothelium. This leukocyte–endothelial cell interaction is a multistep process, comprising initial leukocyte rolling followed by subsequent firm leukocyte adhesion and transendothelial migration.81,82 Depending on the activation of the endothelium, this process is mediated by the upregulation of specific endothelial surface receptors, such as P-selectin, intercellular adhesion molecule-1, and vascular cell adhesion molecule-1. 83 Accordingly, the expression profile of these receptors on preformed blood vessels will determine the initial leukocytic inflammatory host response to the implanted tissue construct. Rothenfluh et al. 42 reported that inosculated preformed microvessels in fetal osteochondral grafts of DDY-mice exhibit significantly more adherent leukocytes than newly formed blood vessels ingrowing into avascular grafts. They interpreted their findings by the different maturation states of the inosculated versus the newly developing microvascular network. However, because experimental studies indicate that newly formed blood vessels exhibit a pattern of basal and cytokine-induced expression of adhesion molecules that is similar to nonproliferating vessels, 84 there may be other mechanisms causing these differences in leukocyte–endothelial cell interaction. For instance, Ang-2 expression has been shown to be upregulated in endothelial cells under hypoxic conditions, as they may occur in implanted prevascularized tissue constructs. 85 In vitro and in vivo experiments demonstrate now that Ang-2 promotes leukocyte adhesion by sensitizing endothelial cells toward TNF-α and modulating TNF-α–induced expression of intercellular adhesion molecule-1 and vascular cell adhesion molecule-1. 86
Increased inflammatory cell infiltration into prevascularized implants may also be observed when the microvascular cells of the preformed blood vessels do not originate from the recipient of the tissue construct. In this case, a typical allograft rejection with T-cell–mediated injury of the donor microvascular cells is likely to occur, which can currently be treated by immunosuppressive therapies. 87 In the future, cultured endothelial cells may also be genetically modified before their incorporation into tissue constructs to prevent allogeneic reactions. Zheng et al. 87 demonstrated that B-cell lymphoma 2 (Bcl-2) transduction protects human endothelial cell synthetic microvessel grafts from allogeneic T-cell–mediated injury in vivo. However, to avoid allograft rejection, those endothelial cells may be used, which are isolated from that patient, who finally will receive the prevascularized tissue construct. Alternatively, a tissue construct may be directly prevascularized in the patient at a well-vascularized site by extrinsic vascularization before its final transfer into a tissue defect site. 17
Finally, rapid establishment of blood perfusion by inosculation of preformed microvessels may allow the release of ischemic signals, such as stromal cell–derived factor-1, 88 resulting in systemic mobilization and homing of bone marrow–derived endothelial progenitor cells in the implanted tissue constructs. These cells have been suggested to contribute to the development of new blood vessels via the process of vasculogenesis at sites of ischemia.89,90 Recently, Capla et al. 66 could show that invasion of bone marrow–derived endothelial progenitor cells into prevascularized skin grafts indeed coincided with the beginning of inosculation. Of interest, they further found that their contribution to the overall process of skin revascularization was up to a considerable rate of 15–20%.
In summary, these findings indicate that inosculation constitutes a key event in the vascularization process of prevascularized grafts, which does not only ensure a rapid life-sustaining supply of oxygen and nutrients but also initiates systemic molecular and cellular interactions between the graft and the host. Consequently, inosculation is associated with remodeling, involving both the microvasculature and the grafted tissue cellular composition.
Future Challenges and Conclusions
Recent progress in generating microvascular networks in vitro and in vivo offers now the exciting opportunity to create tissue constructs that can rapidly vascularize after implantation into the host by the process of inosculation. 17 In fact, in many cases inosculation could be observed within the first 4 days after implantation of prevascularized tissue substitutes, while ingrowth of new blood vessels into large three-dimensional tissue constructs via the process of angiogenesis can take several weeks. However, experimental studies indicate that even this approach does not result in adequate blood perfusion of grafted tissue constructs during the very first days after implantation.25,27 This is most probably due to the fact that inosculation can only occur when the preformed blood vessels of a tissue construct and the preexisting blood vessels of the host tissue grow toward each other to finally form interconnections. Accordingly, time-consuming angiogenic processes may first have to be initiated. Therefore, strategies have to be developed that will increase a directed sprouting angiogenic activity in both counterparts, namely, the preformed microvascular network and the host microvasculature.
The number and density of sprouts at the implantation site may be increased by topical treatment with injections of pro-angiogenic growth factors or addition of growth factor containing gels, such as activated platelet-rich plasma. 91 Alternatively, drugs like erythropoietin that have been shown to induce angiogenesis at ischemic sites92,93 may be administered systemically. On the other hand, the maturation state and angiogenic activity of preformed blood vessels within tissue constructs may be modified to optimize the conditions for inosculation. For this purpose, stem cells or endothelial progenitor cells can be additionally seeded onto scaffolds to capitalize on their vessel-forming capacity.21,94 Moreover, Shepherd et al. 26 reported that during culture of microvessel fragments, perivascular cell coverage decreased to almost half of that observed in freshly isolated ones. Considering the fact that perivascular cell withdrawal from vessel segments relaxes control of the endothelial cell tube and permits rapid sprouting and vessel elongation during angiogenesis, 95 this approach may be useful to modify the maturation state of preformed microvascular networks in vitro, resulting in external inosculation and accelerated reperfusion after implantation into the host. However, these strategies will accelerate the inosculation process only to a certain degree. To reduce the loss of functional tissue during the very initial period after implantation of the tissue construct, it may also be considered to increase the ischemic tolerance of cells inside the construct through distinct methods of preconditioning.96,97
To test the efficacy of all these approaches, sophisticated in vitro and in vivo models are needed in tissue engineering research, allowing a detailed analysis of the molecular and cellular mechanisms that are involved in the process of inosculation. For this purpose, Nicosia et al. 98 established a new ex vivo model to study angiogenic sprouting from explants of the rat inferior vena cava. Combined with the well-established aortic ring assay, this easy model enables for the first time the study of arterio-venous anastomosis formation in vitro (Fig. 3). For the in vivo analysis of inosculation, the dorsal skinfold chamber model was recently introduced.25,37 By means of intravital fluorescence microscopy, this model allows the direct and repetitive in vivo visualization of the interconnection of preformed microvascular networks to the blood vessels of the host tissue and the quantification of microhemodynamic parameters, including volumetric blood flow and wall shear rate (Fig. 4). Moreover, novel transgenic mouse models, such as the tie2/lacZ or tie2/GFP mouse,66,99,100 offer the opportunity to precisely delineate preformed microvessels in a tissue construct from the host microvasculature using immunohistochemical methods. This makes it possible to study the extent of blood vessel outgrowth and ingrowth after implantation of prevascularized tissue constructs. Taken together, these models may help to gain new insights in the regulatory mechanisms of inosculation and to develop new strategies to further accelerate the establishment of a life-sustaining blood supply to implanted tissue constructs.
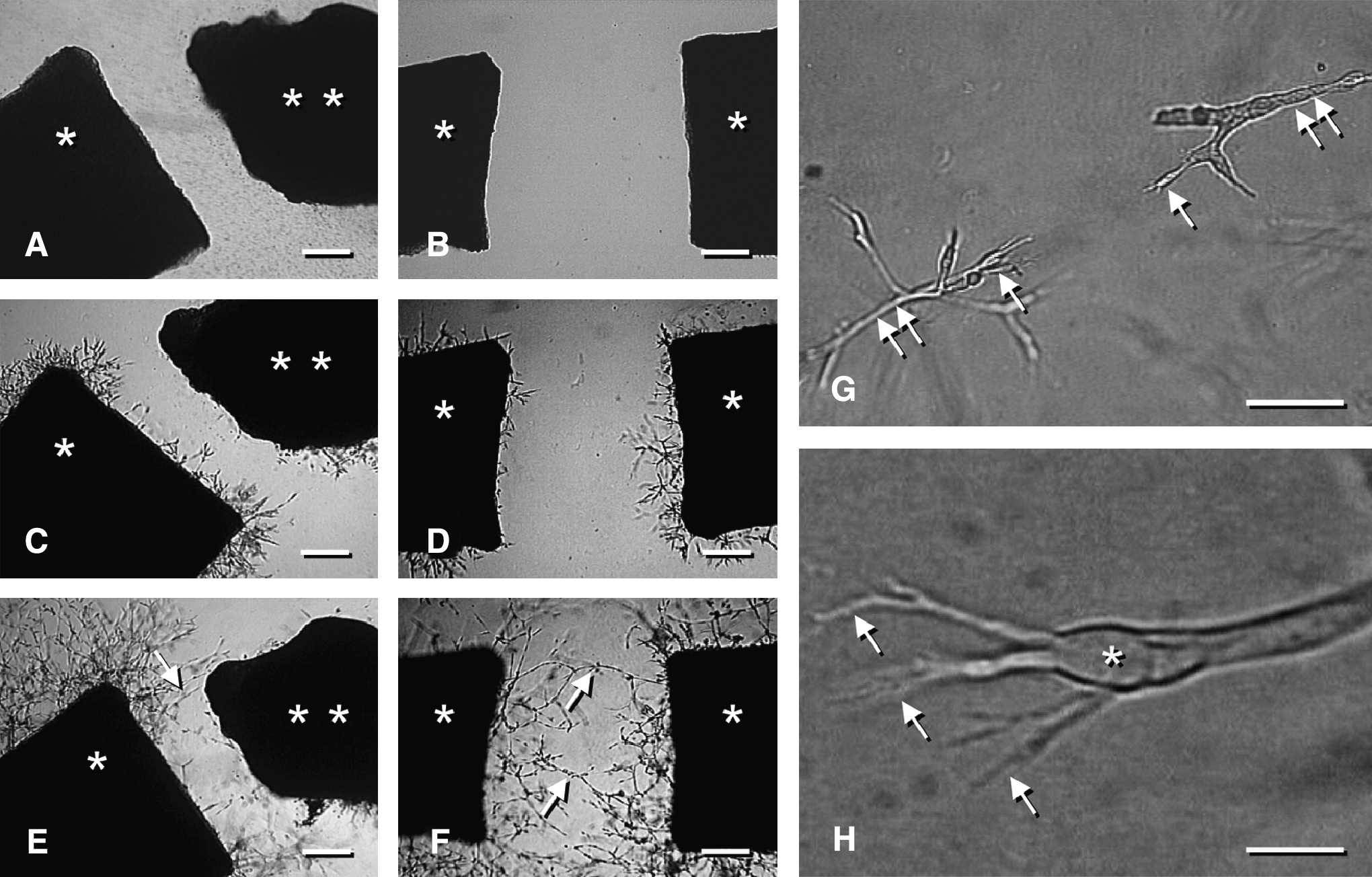
In vitro analysis of inosculation according to the model of Nicosia et al.
98
(modified). (
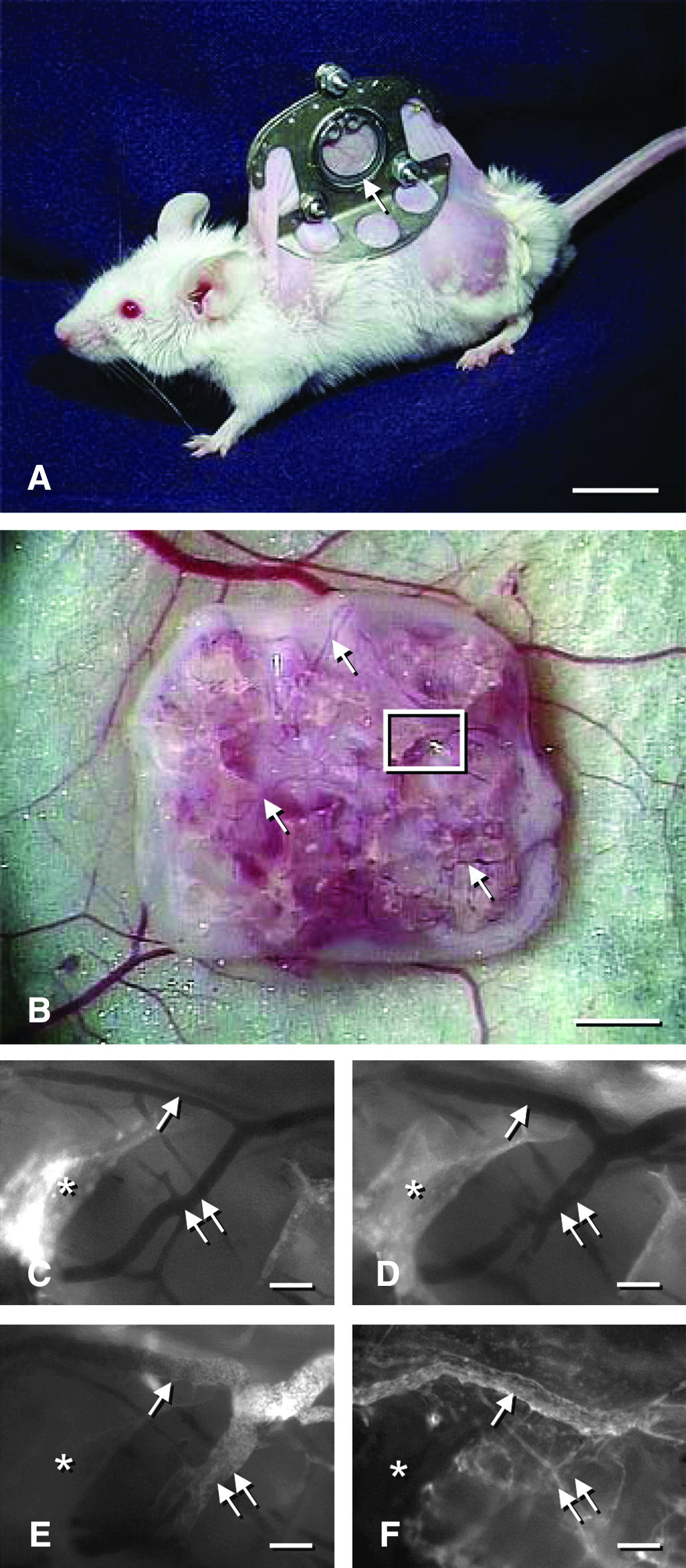
Dorsal skinfold chamber model for the in vivo study of inosculation, as previously described by Laschke et al.
25
(
Footnotes
Acknowledgments
We are grateful for the excellent assistance of Janine Becker, Gunnar Jensen, and Haider Mussawy while creating the figures.
Disclosure Statement
No competing financial interests exist.