
Abstract
Research regarding the potency and potential of the fracture hematoma has begun to receive increasing attention. However, currently there is a paucity of relevant literature on the capability and composition of the fracture hematoma. This review briefly summarizes the regenerative fracture healing process and the close interplay between the skeletal and immune systems. The role of immune cells in wound healing is also discussed to clarify their involvement in immunological processes during regeneration. We attempt to describe the current state of knowledge regarding the fracture hematoma as the initial stage of the regenerative process of fracture healing. The review discusses how a better understanding of immune reactions in the hematoma may have implications for bone tissue engineering strategies. We conclude the review by emphasizing how additional investigations of the initial phase of healing will allow us to better differentiate between deleterious and beneficial aspects of inflammation, thereby facilitating improved fracture treatment strategies.
The Bone and Its Regeneration
Bone also has a substantial capacity for repair and regeneration in response to injury or fracture: “The healing of the fracture is one of the most remarkable of all the repair processes in the body since it results, not in a scar, but in the actual reconstruction of the injured tissue in something very like its original form.” 1
Fracture healing involves different spatial areas with their particular contribution to the healing process. These spatial areas are the medullar cavity, the area between the cortices, the periosteal layer, and the surrounding soft tissues. 2 Depending on the stability of fixation of the fractured bone fragments, a primary (direct) or a secondary (indirect) healing pattern may occur. 3 However, in most clinical cases a certain amount of mechanical instability leads to some interfragmentary movements and thus to a secondary bone healing process. Fracture healing is achieved through endochondral bone formation, including the transformation of the fibrocartilaginous callus to mineralized cartilage and finally to bone.4,5
It has become clear that during the early phase of bone healing, the specific nature of fundamental processes such as inflammation and angiogenesis depends largely upon the mechanical conditions under which bone healing takes place.6–8 Initial differences in relative expression of mRNA level (within days) may represent the actual onset of the different healing pathways followed under optimal or less optimal healing conditions. 9 An understanding of the processes involved, their timing, and the molecular basis of these processes may help to avoid delays in healing or help to accelerate regeneration.
The fracture healing process has been divided into consecutive but overlapping stages. The most fundamental steps are inflammation, repair, and remodeling. 5 Different classifications have been proposed.2,4,10,11 Taken together, a most elaborate classification of fracture healing processes would be hematoma formation, inflammation, neovascularization and granulation tissue formation, fibrous tissue formation, fibrocartilage, hyaline cartilage (soft callus), cartilage mineralization, woven bone (hard callus), and finally remodeling. It should be stressed that these processes overlap and that some of them occur concomitantly (Fig. 1A).
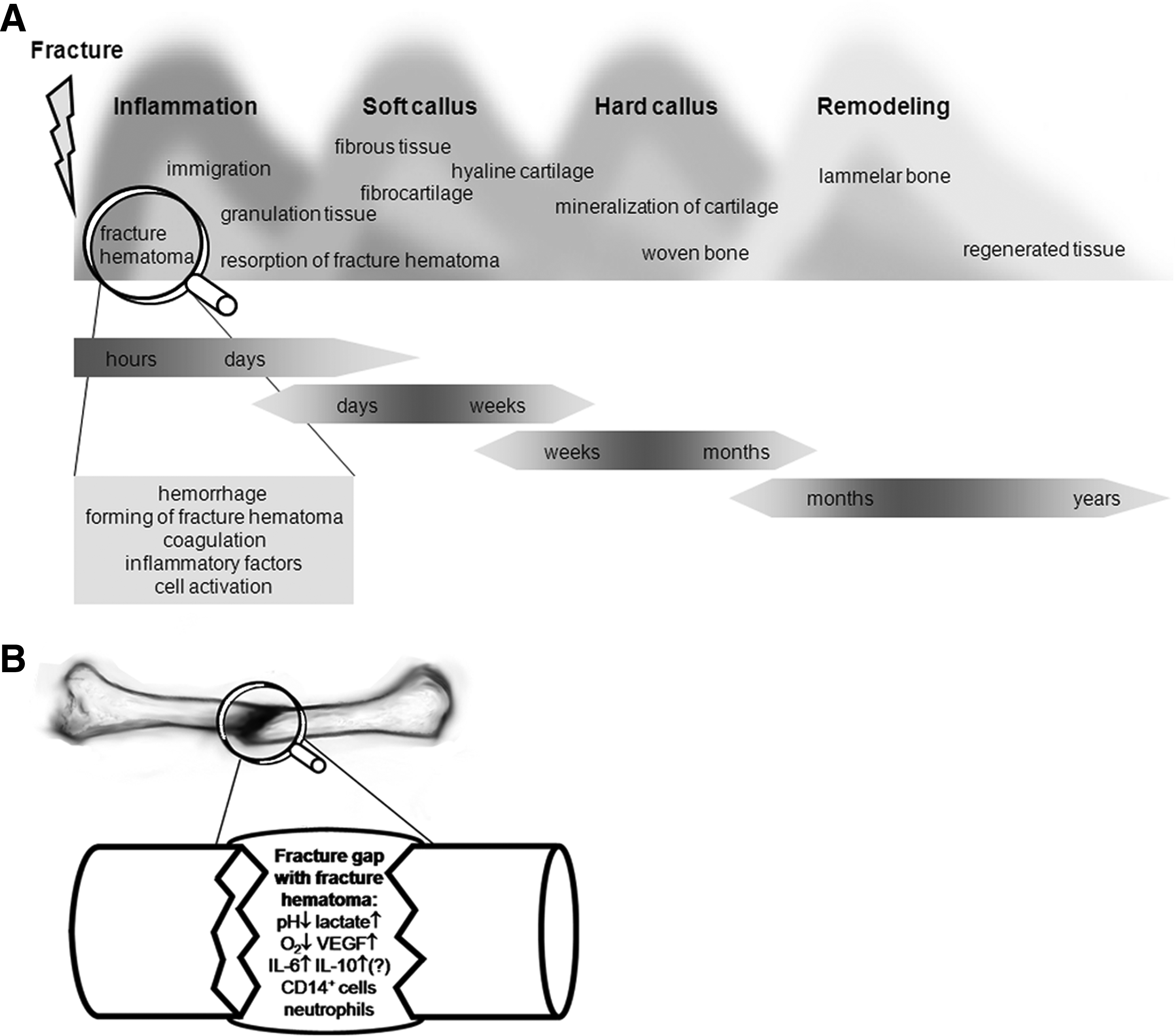
(
The impact causing the fracture not only destroys bone architecture, but also disrupts blood vessels at the site of injury and consequently leads to the formation of a hematoma. Right from the beginning, the hematoma contains immune cells, circulating in the peripheral blood, and bone marrow cells. Interestingly, the inflammation phase is mostly defined as the invasion of inflammatory cells, whereas the immune cells (and their functions) that are already present in the fracture hematoma appear to be ignored in the current literature. Coagulation leading to the fracture hematoma activates the immune cells that are located within the evolving hematoma. The same pro-inflammatory stimuli that activate the human clotting cascade also activate neutrophils, monocytes, and macrophages. 12 A close linkage between clotting and inflammation is supported phylogenetically, and most invertebrates possess a common cellular and humoral pathway of inflammation and clotting. 12 Under stable fixation (3 mm midshaft osteotomy of an ovine tibia) small islands of granulation tissue are visible in an otherwise homogenous hematoma (Fig. 2A), residing in the fracture gap for days after surgery. Around day 7, the main part of the hematoma has developed into granulation tissue (Fig. 2B). After 14 days only a small remnant of the fracture hematoma is present in the osteotomy gap (Fig. 2C). However, with a rotationally instable fixation, the hematoma prevails in the fracture gap for longer than 14 days (Fig. 2D).

Photomicrographs of histological sections depicting the ovine fracture hematoma in a stable (
From an immunological point of view, the fracture hematoma represents the starting point of inflammatory processes, thus initiating the healing cascade (Fig. 1A). These events suggest that the initial inflammatory phase may be crucial in determining the fracture healing outcome.13–15 This would be of special interest to fracture patients suffering from challenging immune conditions.
Crosstalk of Immune and Bone Cells
Even though the close relationship between the skeletal and immune system has been known since the early 1970s, 16 the osteoimmunological research field is still described as being heuristic. 17 The complex regulatory interactions between bone-remodeling cells and immune cells led to the term “osteoimmunology.” 18 Immune cells and skeletal cells share various regulatory molecules, receptors, signaling molecules, and transcription factors, accentuating their connection.17,19,20 This is even more prominent, taking into account that macrophages and osteoclasts develop from the same progenitor cell. Immune cells do influence skeletal cells: activated T cells expressing RANKL can enhance osteoclastogenesis.17,21,22 Additionally, THelper17 (TH17) cells exhibit an osteoclastogenic effect mediated via interleukin (IL)-17 secretion. 17 On the other hand, osteoclast formation can be suppressed by CD4+CD25+Foxp3+ regulatory T cells either via cell–cell contact or the secretion of transforming growth factor β (TGF-β), IL-4, and IL-10. 23 Different cytokines produced by T cells inhibit RANKL signaling and thus osteoclastogenesis, for example, interferon γ (IFNγ), as a typical TH1 cytokine and IL-4 (TH2 cytokine).18,21 In turn, T cell cytokines also affect differentiated osteoblasts, and among them is IFNγ, which inhibits the collagen synthesis 24 and IL-4, which is a chemoattractant for osteoblasts. 25 Hence, bone homeostasis is greatly influenced by immune cells.
Immune Cells in Wound Healing: Can These Findings be Transferred to Fracture Regeneration?
The current literature does not give thorough information on the cells that are active in the initial fracture hematoma, and thus the beginning of the pathway that ends in a regenerative bone repair is today unknown. Therefore, the findings on immune cells in wound healing are recapitulated and the possibility to transfer this knowledge to the fracture regeneration is considered in the following.
Unlike wound healing, at the end of the remodeling phase in bone no inferior scar tissue persists, but rather a tissue with completely regenerated capacities. To this extent, research findings concerning immune responses and their participation in wound repair may not be directly transferred to the processes of bone regeneration. However, wound healing may contribute to the understanding of the immunological response in fracture healing.
Within 24 h after injury, neutrophils migrate to the site of wound repair. Due to their phagocytotic capacities, they decrease the occurrence of wound infection. Neutrophils do not seem to be essential for tissue repair, 26 although a beneficial role has been reported for neutrophilic growth factors. 27 A second wave of migration toward the site of tissue injury is carried out by macrophages after 48–96 h. Macrophages play an important role in successful wound healing 28 as they are crucial for both the inflammatory and debridement process. Cytokines and growth factors produced by macrophages influence cellular migration toward the site of injury chemotactically, and they affect cell proliferation as well as collagen synthesis and angiogenesis. 29 Cytokines and growth factors secreted by macrophages include tumor necrosis factor-α, IL-1, IL-6, IL-8, IL-12, TGF-β, platelet-derived growth factor, and insulin-like growth factor-1. Macrophages also express inducible nitric oxide synthase (iNOS) and produce reactive radicals, such as NO.28,29 Wound healing is less effective in animals depleted of macrophages, 30 whereas supplementation with macrophages increases wound-breaking strength and collagen synthesis. 31
Similar to macrophages, depletion of all T lymphocytes resulted in less effective wound healing, 32 whereas combined depletion of CD8+ and CD4+ T cells as well as CD8+ T cells alone resulted in enhanced wound healing,28,29 while CD4+ T cell depletion alone has no effect on wound breaking strength. These data suggest a downregulating role in wound repair for CD8+ T cells, while CD4+ TH cells seem to have no effect on wound healing. 33 However, CD4+ TH cells are composed of subpopulations with differing cytokine expression patterns. TH1 cells produce IFNγ, a cytokine that induces expression of iNOS in macrophages. 34 Through the positive effect of NO on wound healing, these TH1 cells could, nevertheless, be beneficial for the healing process. 29
Although similarities between the role immune cells play in wound healing and fracture repair could be anticipated, the initial presence of bone marrow cells in the fracture hematoma could make a crucial difference. Further, the migration of immune cells into the fracture site could be initiated from the peripheral blood, but also to a large extend from bone marrow. Thus, similarities as well as differences are to be expected when comparing wound and fracture healing.
Fracture Hematoma as the Onset of Fracture Regeneration
Histological studies carried out as early as the first half of the 20th century already suggested that the periosteum and the fracture hematoma play an important role in fracture healing. 35 Meanwhile, the sequence of processes during fracture healing has been well characterized2,4,5,10,11 (Fig. 1A). Although the importance of the initial fracture hematoma for effective healing is apparent,36,37 its exact regulatory role in the initiation of regeneration still remains vague. Little is known about the initial cellular and humoral composition of the fracture hematoma.
The importance of the initial fracture hematoma for optimal fracture healing has been unambiguously shown in animal models. Removal of the fracture hematoma between days 2 and 4 as well as repeated hematoma debridement on days 1 and 2 resulted in delayed or nonunion.36,37 This is a clear indicator that the fracture initiates healing—processes in the fracture hematoma that are so far not understood start a cascade that results in regenerated bone if undisturbed. In contrast to these findings, open fracture treatment frequently employs irrigation and debridement to minimize the risk of infection. Even in closed fractures, the initial hematoma is seldom considered as the onset of bone healing and removed if indicated.
Abilities of the fracture hematoma such as induction of angiogenesis or periosteal cell proliferation and subsequently initiation of intramembranous bone formation develop over time. A 2-day-old rat fracture hematoma produces new bone only in cooperation with neighboring periosteum, whereas a 4-day-old fracture hematoma forms ectopical bone when implanted in a muscle.38,39 An even more matured fracture hematoma harbors mesenchymal stem cells able to differentiate into a chondrogenic or osteogenic cell type.40,41 Secreted factors also play an important role for the development of osteogenic abilities of the fracture hematoma. This becomes evident through the osteoinductive characteristics of the fracture supernatant.39,41 These studies underline the fact that in a mature fracture hematoma the initial steps of fracture healing have already taken place.
Direct Fracture Hematoma Analyses
Knowledge about the fracture hematoma has been gained indirectly through analysis of the fracture hematoma supernatant (Table 1): low pH and increased phosphate, alkaline phosphatase, and lactate values 42 could be interpreted as first hints of the low O2 tension later proven in the fracture hematoma.43,44 An increased β- and γ-globulin fraction has been detected in fracture hematoma supernatant via electrophoresis. The β-globulin rise could be due to hemoglobin and fibrogen, 42 whereas the elevated γ-globulin fraction could indicate the involvement of immune cells. 45 The fracture hematoma supernatant containing the angiogenic factor vascular endothelial growth factor has been found to be toxic to endothelial cells in vitro,46,47 but subcutaneous hematoma transplantation led to new blood vessel formation in vivo. 47 This implies that the fracture hematoma is not toxic per se, but exhibits a complex microenvironment with different effects on different cell populations, which could be of great importance for migratory cell populations.
Further studies utilized a mixture of fracture hematoma and the surrounding soft tissue hematoma to gain further knowledge on the fracture hematoma composition (Table 1). Studies investigated the assumption that immunosuppressive factors were present in the fracture hematoma as it is known that clinical trauma is able to suppress cell-mediated immunity resulting in systemic immune suppression.48,49 Mononuclear cells and cell-free fluids from experimental wounds were shown to have immune suppressive properties.45,50 In an animal model, closed fractures, not open fractures, were shown to be immunosuppressive, suggesting the fracture/wound microenvironment as a possible source for immune modulatory factors. 51 Thus, Hauser et al. investigated the presence of immunosuppressive cytokines in fracture/soft tissue hematoma in humans. 48 No significant alteration of IL-4 and TGF-β could be detected, but high amounts of IL-10 were present in the early (<24 h) fracture/soft tissue hematoma. However, the material examined was a mixture of fracture and soft tissue hematoma so that no firm statement about the pure fracture hematoma is possible. In a subsequent study, more detailed analysis of cytokines was performed using fracture/soft tissue hematoma. 52 This study has to be considered carefully, since the hematoma were treated with urokinase, which enables matrix-bound (and therefore in vivo inactive) cytokines to be released. However, IL-6, IL-8, IL-12, and, as shown previously, IL-10 were time dependently upregulated in the fracture/soft tissue hematoma: IL-10 peaked before 24 h postfracture, IL-6 before 48 h, IL-8 after 48 h, and IL-12 on days 4–7. Elevated IL-6, IL-8, and IL-12 protein levels could even be detected in the plasma of the injured patients as well. 52 Thus, at least for the mixture of fracture and soft tissue hematoma expression of immunosuppressive cytokines has been shown.
Even though the fracture healing outcome has been investigated in several animal studies when the fracture hematoma was removed at different time points,36,37 no further analysis of the removed fracture hematoma was done. The identification of cellular composition, activity of these cells regarding factor synthesis, and release has so far not been done. In our group, a sheep osteotomy model provided a fracture hematoma with only minimal soft tissue damage and was therefore considered as pure fracture hematoma. We found that the cellular components, especially the immune cell populations of the fracture hematoma, change very quickly in the initial phase (within hours after fracture) and differ from those of a parallel investigated, corresponding muscle hematoma. 53 Further, our preliminary unpublished data show differences between the cellular composition of fracture hematoma and peripheral blood: in the initial human fracture hematoma (6–72 h), the ratio of granulocytes/monocytes to lymphocytes is doubled in comparison with the ratio concerning the peripheral blood. Additionally, within the lymphocyte population, the percentages of CD3+CD4+ TH cells and CD3+CD8+ cytotoxic T cells are decreased within the fracture hematoma when matched to corresponding blood samples.
There is plenty of evidence that the immune system influences fracture healing and vice versa, that fractures influence the systemic immunity, and that fracture healing can be delayed in immune suppressed patients.54,55 As elderly patients exhibit a higher risk of fracture,56,57 the limited immune function in the elderly has to be considered.58–62 Trauma is known to suppress the immune functions,48,63 and it was shown that the human leukocyte antigen-isotype DR (HLA-DR) expression on circulating monocytes as an immunocompetence marker was downregulated in patients with fractures. 52 The close relationship between bone and the immune system is obvious.
Bone Tissue Engineering and Immunological Features
The treatment of large bone defects represents a challenging task. Gold standard so far is transplantation of autologous bone material (autografts) with limitations like donor-site morbidity, material availability, and difficult tissue in-growth. A tissue engineering approach might offer a solution for these restrictions by modifying the immune reaction in the wound area, being a source for pluripotent cells, and aiming to stimulate regeneration ex vivo or in vivo. Therefore, cells are being seeded in a three-dimensional construct to create a biocompatible, osteoconductive, as well as osteoinductive scaffold with good tissue-bonding capabilities. For tissue-engineered bone the clinical application is the ultimate test. So far only a few approaches are being investigated in clinical trials. The most important are Quarto et al., with the first report of a cell-based tissue engineering approach for a 4–8 cm bony defect; Hernigou et al., who optimized the cell number necessary to achieve positive results; and Marcacci et al., with an application of hydroxyapatite scaffolds seeded with bone marrow cells.64–66 Cells applied so far in bone tissue engineering are mesenchymal stem cells and stromal cells from bone marrow,65,66 periosteal cells67,68 and osteoblasts. 69 One important result of these studies is a positive correlation between applied cell numbers and healing result. Directly after implanting the tissue-engineered bone is surrounded by a developing hematoma. A hematoma also develops between the refreshed bone fragments and the scaffold at the contact sites. Hematoma itself harbors conditions toxic for some cells (e.g., endothelial cells) such as a low pH, high potassium, and sodium concentrations. 47 These conditions are probably also harmful for the cells seeded on scaffolds, at least in the contact areas. Immune cells are able to withstand conditions in fresh hematoma 53 and are able to increase fracture healing through specific factor release. For example, vascular endothelial growth factor and platelet-derived growth factor are angiogenic factors released by immune cells, and early revascularization is one of the major prerequisites for fracture healing. Specific immune cell subpopulations (regulatory TH cells) are able to reduce the inflammatory reaction induced by scaffold implantation as well as coagulation. Seeding scaffolds with immune cell subpopulations or coating them with factors inducing/chemoattracting specific subpopulations could enhance/optimize tissue in-growth abilities and revascularization as well. Additionally, the buffering of, for example, high concentrations of potassium could enhance survival of seeded cells. Thus, deeper knowledge of the processes occurring in the fracture hematoma initiating fracture healing could also benefit bone tissue engineering. Favorable processes induced by immune cells in the fracture hematoma could be used to elevate tissue-engineered bone in-growth, whereas negative influences of distinct immune cell populations could be inhibited by specific depression via drugs. One could imagine to, for example, enhance the inflammatory reaction time dependently through modified release of inflammatory factors but also to terminate (using antiinflammatory drugs such as glucocorticoids or nonsteroidal anti-inflammatory drugs [NSAIDs]) the inflammatory phase in case of prolonged inflammation as it might occur in patients with autoimmune diseases. Also, inhibitors of tumor necrosis factor-α or IL-6 could be used locally to dampen inflammatory reaction. To influence costimulation of T cells via cytotoxic-T-lymphocyte-associated antigen-4-Ig could also be advantageous. Immunomodula tory drugs are already used successfully in the clinic and the growing knowledge in the field of osteoimmunology could establish new application possibilities.
Further Questions and Experimental Needs in the Future
It is now accepted that the inflammatory phase in fracture healing is important. However, the inflammatory phase has been described as the phase when inflammatory cells are recruited to the fracture site, for example, 24 h after injury, but immune cells are present in the fracture hematoma immediately after fracture, since bleeding into the fracture gap forms the fracture hematoma. The role of these immune cells and factors in the hematoma is not yet fully understood.
Prospectively, it will be important to characterize the initial fracture hematoma more thoroughly and to define generally accepted terms. This, however, can be problematic as the sample quantity of pure fracture hematoma, without soft tissue hematoma, can be very small, depending on the type of fracture. This fact might explain the difficulties in characterizing the material immunologically in more detail on a cellular and on a humoral level as seen in the past. Figure 1B gives an overview about the known components of the initial fracture hematoma. Due to the development of new technologies, it is now possible to perform more detailed and necessary examinations in this field. Clarifying the role of the immune system at the beginning of the fracture healing could lead to the development of new and specialized treatment strategies in fracture healing. Further, it is not only positive consequences that may derive from the inflammatory phase: a prolonged inflammatory phase might be detrimental to the bone healing process. A better understanding of the cascades present within the inflammatory phase—as based on an understanding of the cells and humoral factors—will potentially allow us to differentiate between those aspects that are essential for a fast and effective bone healing, and those factors that delay healing. As life expectancy and numbers of immune-deficient patients increase, incidences of fractures and consequently nonunions increase.56,57 Thus, faster mobilization and reduced hospitalization duration is of great ethical and economic importance, not only for the patients, but also for the community. In light of these aspects, a detailed investigation of the initial fracture hematoma would be of great interest to ensure a more effective fracture healing treatment in the future, especially in patients at risk for delayed-healing or nonunion.
Footnotes
Acknowledgments
Reported studies were supported by a grant from the German Research Foundation (DFG SFB 760) and partially by the Berlin-Brandenburg Center for Regenerative Therapies. The authors would like to thank Dr. Bettina Willie for editing this article.
Disclosure Statement
No competing financial interests exist. All authors have no conflicts of interest.