
Abstract
For years it has been recognized that engineering of large bone constructs will be feasible only if the hurdle of vascularization is overcome. Attempts to engineer bone tissue have predominantly focused on intramembranous (direct) bone formation. A relatively new and most likely more physiological approach in this line is endochondral bone formation, comprising an intermediate cartilaginous stage. Cartilage in nature is an avascular tissue and its cells are equipped to survive the poor oxygenation and nutritional conditions inherent to implanted tissues. Subsequent terminal differentiation (hypertrophy) of the chondrocytes initiates the formation of a mineralized matrix that will then be converted into bone. Through this mechanism, our long bones grow and most fractures heal through the process of secondary fracture healing. The feasibility of the attractive concept of endochondral bone tissue engineering has already been shown. Most emphasis has gone to the multipotent stromal cells because of their great potential for expansion and differentiation and immunoprivileged nature. This review will focus on the promises and current status of this new field. Further, potent modulators of endochondral bone tissue engineering, including oxygen tension and mechanical stimuli, will be discussed.
Introduction
During intramembranous bone formation, a bone matrix is directly deposited by osteoblasts. It is this process that is commonly encountered by bone tissue engineers,1–3 but it has not yielded optimal results yet.4,5 Such bone substitutes typically consist of cells and a carrier matrix or scaffold. Various cell types have been used in these approaches, including multipotent stromal cells (MSCs), also known as mesenchymal stem cells.6–9 When seeded in a scaffold, MSCs have been shown to enhance in vivo bone formation after implantation.10–16 Several factors such as osteogenic stimulation, duration of osteogenic stimulation, and osteoinductive properties of the scaffold influenced in vivo bone formation.10–16 Additionally, MSCs have been studied in a few clinical cases to a rather limited extent, to assist in filling bone defects when seeded on scaffolds17,18 and to treat genetic disorders.19,20
The alternative route along which bone can be formed is endochondral ossification, which also starts with the condensation of mesenchymal stem cells. Subsequently, in this route, these cells differentiate into the chondrogenic lineage and then go into hypertrophy before finally entering terminal chondrogenic differentiation resulting in a mineralized cartilage matrix. Interestingly, this additional approach for bone formation has been largely overlooked in the field of bone tissue engineering. A possible explanation may be the indirect and apparently time-consuming character of this process and the difficulties associated with chondrogenic differentiation of MSCs. However, the natural process of endochondral bone formation is associated with several advantages when translated to approaches for bone tissue engineering. One benefit over the intramembranous approach is its avascular onset. Clearly, forming bone through the production of a cartilaginous template may circumvent issues with supply of oxygen and nutrients of larger engineered constructs, because cartilage cells are equipped to survive poor environmental conditions, such as low oxygen tensions.21–23 Importantly, promising and accessible sources for cells with significant chondrogenic potential are the populations of MSCs that reside in the bone marrow or nonskeletal tissue, such as adipose tissue. An advantage of using chondrogenic progenitors, such as the MSCs, over using chondrocytes is that they are easier to obtain at high numbers. These benefits demonstrate that endochondral bone formation may be a promising alternative approach for bone tissue engineering.24–28
This review will focus on the promise and limitations of differentiating the MSCs through the endochondral route for bone tissue engineering purposes. The process of endochondral ossification using MSCs will be discussed both in vivo and in vitro, as well as the relevance of several environmental factors that affect their hypertrophy, including growth factors, oxygen levels, and mechanical cues.
Endochondral Ossification
In the embryo, MSCs condense and differentiate into chondrocytes forming the cartilaginous anlage. The anlage forms the cartilage mold for the long bone. Endochondral ossification commences in the primary ossification center in the center of the anlage. Later, secondary ossification centers are formed at the periphery of the long bone structure.29,30 Between the ossified parts, growth plates develop serving the lengthening of the bones, during which several stages characteristic of endochondral ossification can be well observed. The zones can be distinguished morphologically as well as by the production and release of several growth factors and matrix components (Fig. 1).
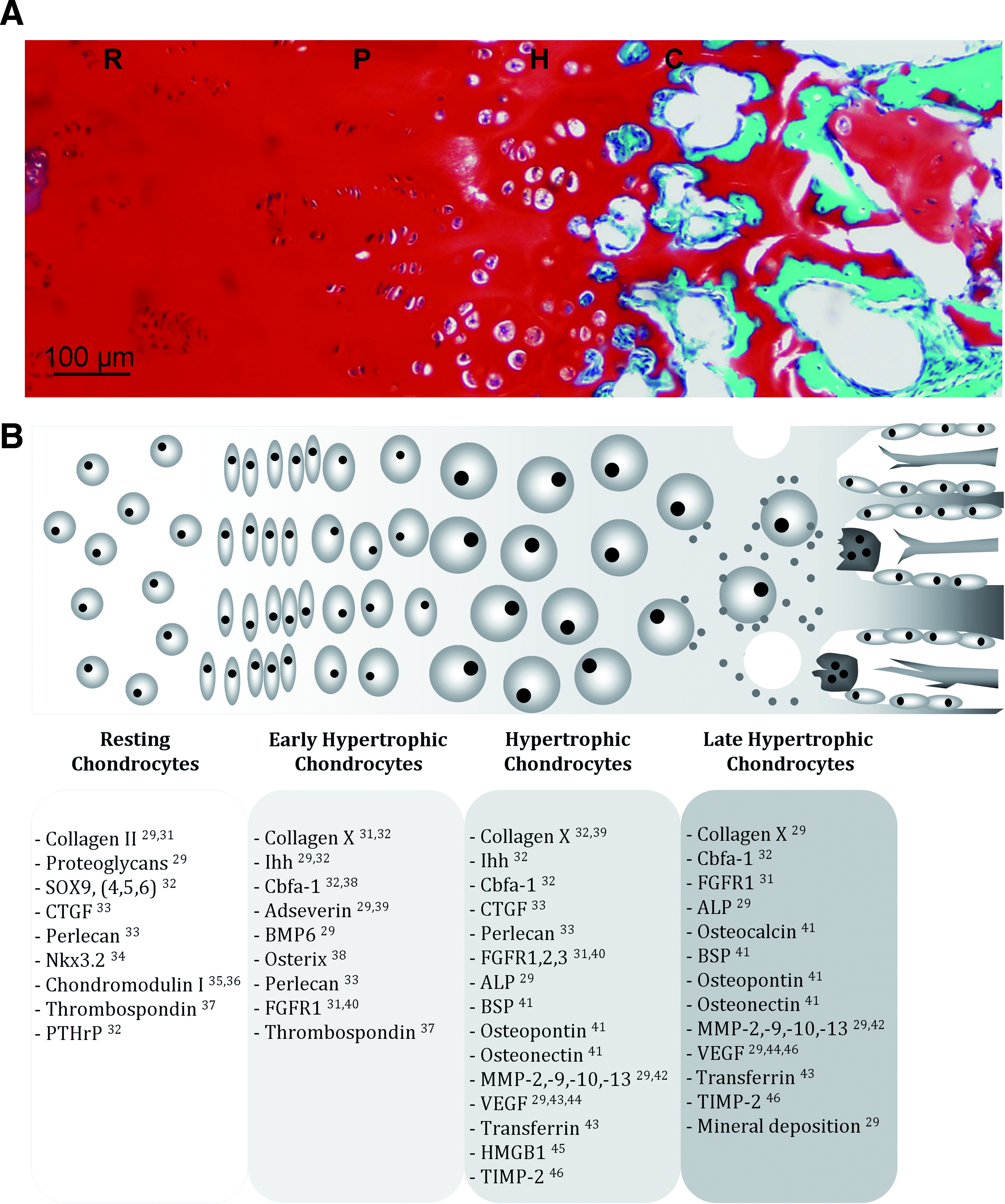
Morphological features of cells during endochondral ossification. (
The first zone consists of resting chondrocytes in an avascular environment. The chondrocytes secrete antiangiogenic factors and synthesize a matrix that mainly consists of collagen II and proteoglycans. In the adjacent zone, flattened chondrocytes proliferate and align in columns. Subsequently, the chondrocytes further differentiate and enter the prehypertrophic stage. Adseverin, bone morphogenetic protein-6 (BMP-6), and Indian hedgehog (Ihh) are produced during this stage, 29 as well as cbfa-1 (or runx-2) and osterix. 38 Expression of cbfa-1 stimulates the upregulation of hypertrophic markers collagen X, Ihh, and vascular endothelial growth factor and is itself controlled by oxygen tension. 29 Adseverin (scinderin) is involved in cytoskeletal rearrangements of maturing chondrocytes and is essential for cell hypertrophy. Further, it stimulates the expression of collagen X and Ihh. 39 In turn, Ihh, expressed by early hypertrophic cells, stimulates proliferation and the expression of parathyroid hormone-related protein, which inhibits transition to hypertrophy, thus together creating a negative feedback loop of chondrocyte hypertrophy. 47 BMP-6 also stimulates the expression of collagen X. Once collagen X is synthesized, the cells are differentiated into the hypertrophic stage, in which the individual cells increase their size. During this terminal stage of differentiation, the hypertrophic cells no longer express chondrogenic genes, such as sox-9. They are now characterized by expression of the osteogenic genes alkaline phosphatase (ALP), cbfa-1, collagen I, osteonectin, and during late hypertrophy, osteopontin (also bone sialoprotein-1 or secreted phosphoprotein-1), bone sialoprotein-2, and osteocalcin.42,48–52 In addition, the matrix around the late hypertrophic chondrocytes is mineralized through deposition of hydroxyapatite.
Matrix-degrading enzymes such as the matrix metalloproteinases MMP-2, MMP-9, MMP-10, and MMP-13 are now also upregulated.29,42 Matrix degradation leads to the formation of (cartilage) canals and cavities inside the cartilaginous matrix. 53 Additionally, hypertrophic chondrocytes produce angiogenic factors such as transferrin, vascular endothelial growth factor, and high mobility group box protein 1, to attract both blood vessels and bone cells.43,45,54–56 Simultaneously with increased production of angiogenic factors, the expression of antiangiogenic factors (chondromodulin I, thrombospondin 1 and 2, and tissue inhibitor of matrix metalloproteinases 2 and 3) is reduced.35,36,57 All together, these factors stimulate blood vessel invasion of the cartilage canals. The newly formed vessels allow for infiltration of osteoblastic and osteoclastic cells, which will convert the (mineralized) matrix remnants into bone. The cavities, on the other hand, will become populated by the bone marrow. Many of the hypertrophic chondrocytes eventually die at the final stage of differentiation. Their death has recently been described as a process morphologically distinct from apoptosis and necrosis.29,58
Chondrogenesis by MSCs In Vitro
For clinical application of in vitro cultured endochondral bone grafts, the development of engineered cartilage as a first template is required. Subsequent conversion of the template into bone may then take place in vivo. Cells from several sources could be directed to engineer such a template. Although chondrocytes from articular cartilage would be a logical choice for cartilage tissue engineering, their clinical application is associated with several drawbacks. Their low numbers in tissue necessitate expansion before application, which in turn leads to their dedifferentiation causing a switch in the synthesis of extracellular matrix molecules toward more fibrous tissue containing collagen I.59–61 Another disadvantage of this cell source is that it is not harvested from a readily renewable tissue, leaving a permanent defect at the donor site. Finally, chondrocytes can only be harvested for endochondral ossification from particular cartilaginous sites. Chondrocytes isolated from permanent articular cartilage cannot differentiate into the hypertrophic stage when using conventional methods,50,62 whereas differentiation can be induced in chondrocytes from transient cartilage.26,50,62 To overcome these drawbacks, MSCs were proposed as an alternative cell source for culturing cartilage tissue equivalents. 63
Besides marrow and fat, MSCs derived from other sources have been shown to possess chondrogenic potential. In rats, a comparison of MSCs from several sources showed that synovium-derived MSCs had higher chondrogenic potential than MSCs obtained from bone marrow, periosteum, adipose tissue, or muscle.64,65 Similar results were reported for synovium-derived MSCs from human origin.64,66 In chondrogenic pellet cultures of human origin, MSCs from bone marrow, synovium, and periosteum exhibited chondrogenesis superior to that of MSCs from adipose tissue,67–70 fetal lung 71 or the placenta, 71 and skeletal muscle.64,65 However, it should be noted that the inferior chondrogenesis of MSCs from these sources may be compensated for by changing in vitro culturing methods according to the specific receptor expression profile on MSCs from these sources. This was, for example, shown for human MSCs from an adipogenic source that had an expression pattern deviating from that of bone-marrow derived MSCs. Correcting the expression profile by stimulation with the growth factor BMP-6 enabled their chondrogenic differentiation. 72 Therefore, the authors opted for a tailor-made chondrogenic induction medium for MSCs from different sources.
Human synovium-derived MSCs in particular showed promising chondrogenic capacity in vitro in pellet cultures, and additionally, they exhibited a lower tendency to become hypertrophic in vivo (reduced ALP and no mineralization after ectopic implantation) when compared with MSCs from bone marrow and fat tissue. 62 As this is undesirable in endochondral bone tissue engineering, this is a less attractive cell source. Additionally, upon implantation of these pellets in nude mice, the engineered cartilage either dedifferentiated or disintegrated. These results emphasize that the in vitro chondrogenic potential is no guarantee for their successful in vivo chondrogenic performance. Additionally, the opposite case was also reported as it was shown in minipigs that autologous bone marrow-derived MSCs (BM-MSCs) formed more stable cartilage in an in vivo full-thickness defect compared with in vitro culture under chondrogenic conditions. 73 Nevertheless, for chondrogenesis from any cell source it will be desirable to create a stable cartilaginous phenotype. In this line, the in vitro apparent hypertrophic differentiation of human MSCs from bone marrow or fat tissue,50,62,74–76 which appears to lead to the formation of a mineralized matrix in vivo in severe combined immunodeficiency (SCID) mice,50,62 makes MSCs a less-attractive cell source for chondrogenesis under currently accepted culture protocols.
In conclusion, the inclination toward hypertrophic differentiation of MSCs is undesirable for the field of cartilage tissue engineering; however, this offers opportunities in particular for endochondral bone tissue engineering.
Hypertrophy of Chondrogenically Differentiated MSCs
During hypertrophic differentiation of MSCs in vitro, a differentiation pathway is followed, which deviates from the in vivo pathway in terms of upregulation of several genes. Genes normally associated with the hypertrophic stage are observed prior to and during chondrogenesis preceding hypertrophy, whereas this is not the case in chondrocyte cultures. Recently, several studies reported the expression of the hypertrophy-related gene MMP-13 and collagen X protein before collagen II production was evidenced.50,77–79 An elaborate analysis of temporal gene expression during in vitro chondrogenesis (natural hypertrophic tendency was not actively stimulated) of human MSCs was conducted. 78 Several differentiation stages of gene expression were distinguished during a 24-day differentiation period. In the final stage, several osteogenic genes showed increased expression. Additionally, adseverin, gremlin (BMP antagonist), and microfibrillar-associated protein 5 (gene)/microfibril-associated glycoprotein 2 (MFAP5/MAGP-2) (possible role in prehypertrophy to hypertrophy transition) are proposed as putative additional genes for identification of hypertrophy. The roles of these genes may be further elucidated in a model as proposed by Mueller and Tuan, 27 who actively induced hypertrophy in MSC cartilage pellet cultures by withdrawal of transforming growth factor-β and dexamethasone, while adding thyroid hormone (T3).
Endochondral Bone Tissue Engineering In Vivo
Feasibility of endochondral bone tissue engineering in in vivo models is a rather unexplored area. One of the earliest reports of BM-MSCs causing calcification of a chondrogenic culture was in 1981. 80 They showed induction of hydroxyapatite crystals in the matrix surrounding chondrocytes and loss of glycosaminoglycans during an in vitro coculture with bone marrow-derived cells. Ever since, several studies have demonstrated the feasibility of in vivo endochondral ossification of MSC cultures.
Implantation of constructs containing predifferentiated murine embryonic stem cells in mice resulted in hypertrophic chondrocytes lining bone tissue, suggesting the occurrence of endochondral bone formation. When other cells (bovine articular chondrocytes, adult human and goat MSCs) were subjected to the same routine, limited or no endochondral bone formation was found. 24 Possibly, the period of 3 weeks allowed for chondrogenic predifferentiation in this study may not have been sufficiently long for in vivo endochondral bone formation of the (nonembryonic) MSCs. This notion is supported by the observation that chondrogenically primed human BM-MSCs in collagen–glycosaminoglycan scaffolds implanted subcutaneously into nude mice for 4 weeks did show collagen type X expression and invasion from the host vasculature, but no mineralization. 28 Vessel invasion was observed within 4 weeks, and perhaps bone formation would have occurred later. In an additional study, the duration of in vitro culturing was expanded rather than the duration of in vivo implantation. Indeed, prolonging the in vitro preculturing period to 7 weeks did induce matrix calcification in addition to collagen X expression, and vascular ingrowth after subcutaneous implantation of human BM-MSC pellets in immunodeficient mice for 4 weeks. 50 In other studies, the approach consisted of implantation of osteogenically primed MSCs.81,82 Osteogenic differentiation for 4 weeks of bovine MSCs resulted in in vivo evidence of cartilage and bone formation after embedding the cells in 2% alginate in an ectopic murine model for up to 30 weeks. 82 Moreover, implantation of undifferentiated (murine) MSCs in a murine subcutaneous model led to a similar result, with signs of endochondral tissue differentiation in vivo for up to 60 days. 81 In comparison, inclusion of osteoblasts in the latter model gave rise to in vivo bone formation following intramembranous rather than endochondral ossification in vivo. These results may suggest that also osteogenically primed MSCs will undergo endochondral ossification, whereas terminally differentiated osteoblasts are committed to the intramembranous ossification pathway.
Together, these in vivo studies showed that endochondral bone formation from chondrogenically or osteogenically stimulated MSCs is feasible.
Several issues associated with in vivo endochondral bone tissue engineering need to be addressed in future investigations. For example, interspecies differences were reported, 24 which may provide clues to further direct the cells toward the desired tissue. Additionally, growth factor stimulation, for example, by thyroxine, is an interesting means to enhance in vitro hypertrophy of chondrocyte and MSC cultures. However, in vivo persistence of thyroxine-induced hypertrophy has not been studied yet. Additionally, the appropriate time point of implantation needs to be addressed in future studies. Implantation of MSCs during the metabolically less-demanding chondrocyte phase of differentiation may allow for a longer temporal vascularization window after in vivo implantation. Subsequent hypertrophic differentiation will induce the secretion of angiogenic signals that will enable timely vascular invasion of the implant. The increased metabolic demand of the hypertrophic cells may then again shorten the survival period of the implants.
Other cell types than MSCs, such as chondrocytes, may provide additional clues on differentiation processes prolonged or initiated upon in vivo implantation. Recently, the use of chondrocytes for engineering endochondral bone was explored.25,26 In vitro and in vivo endochondral performance was compared for (permanent phenotypic) control chondrocytes and hypertrophic chondrocytes (induced by retinoic acid) from chicken embryo's sternum in chitosan sponges. Only the hypertrophic chondrocytes expressed ALP and collagen X after in vitro culture. Implantation of the constructs into mice resulted within 1 month in cartilaginous tissue formation in the controls and mineralized bone formation with vascular invasion in the constructs containing hypertrophic cells. Thus, formation of mineralized bone from a construct containing chondrogenic cells with hypertrophic characteristics was confirmed with another cell type than MSCs.
Endochondral ossification of both chondrocyte and MSC-based constructs is thus feasible after in vivo implantation. It needs, however, to be stressed that in vivo endochondral progression has only been observed when the implanted cells exhibited a certain degree of hypertrophy prior to implantation.
Modulation of Hypertrophy In Vitro
To further stimulate the hypertrophic nature of potential grafts, several stimulating factors can be employed. Growth factors, oxygen tensions, and mechanical stimuli are known to influence tissue properties and differentiation of cells in general. These stimuli can be applied to control MSC differentiation as well and will be discussed below.
Soluble mediators
Studying the temporal hypertrophic gene expression patterns can be facilitated by biochemical induction of hypertrophy. The most commonly applied way of manipulating cell behavior or differentiation is the addition of growth factors to the culture media. These growth factors are either produced by another cell type or added exogenously at defined concentrations to the media. Several components have been implicated to induce hypertrophy when added to chondrogenic cultures, such as vitamin D3,49,83 retinoic acid, 49 leptin, 84 insulin and thyroxine,27,85,86 and β-glycerophosphate alone, 28 or combined with a concomitant decrease of the concentration of dexamethasone. 27
Oxygen modulation of MSCs
Standard cell culturing techniques have been developed at the ambient oxygen tension of 21%. This oxygen tension is referred to as the normoxic condition. However, oxygen tensions in the bone marrow, where the MSCs reside, range from 1% to 7%.87–90 These low tensions are defined as hypoxic culture conditions, whereas they actually represent the “normal” oxygen levels experienced by cells in the body. Interestingly, these normoxic conditions actually put the MSCs in a hyperoxygenated condition during in vitro expansion and differentiation. These unnatural conditions can cause generation of high concentrations of reactive oxygen species (ROS) (highly reactive oxygen radicals) and concomitant detrimental oxidative stress. The negative effects of normoxic culture conditions have been recognized. 91 The effects of low oxygen tensions have therefore also been studied on MSC expansion and differential capabilities.
MSC expansion under hypoxic conditions has resulted in maintenance of MSC stemness, extension of their in vitro life span, and inhibition of adipogenic and osteogenic differentiation.89–96 The inhibition of osteogenic differentiation by low oxygen tensions could be reversed by subsequent differentiation at atmospheric oxygen tension.89,90 Contradictorily, also stimulation of osteogenesis in rat MSCs under low (5%) oxygen conditions compared with normoxic culture was observed. 97
MSC expansion under low oxygen tensions was also suggested to select for a population of MSCs with chondrogenic potential. 95 Murine fat-derived MSCs were expanded at 2% oxygen and early subsequent chondrogenesis was subsequently observed when cultures were transferred to ambient oxygen tensions. The early onset of chondrogenesis was even maintained when the final stage of expansion was conducted at 21% oxygen, indicating chondrogenic selection had taken place at an early time point during expansion under hypoxia. Also during MSC chondrogenesis, low oxygen levels (2%) were demonstrated to increase collagen II and proteoglycan production. 98 Hypoxia-inducible factor 1α was identified as a mediator, as inhibition of hypoxia-inducible factor 1α by siRNA diminished the effect. Additionally, ROS have been shown to arrest proliferation and stimulate hypertrophic differentiation of chondrocytes. 99
Low oxygen conditions thus favor the selection of a chondrogenically potent subpopulation and stimulate chondrogenesis when culturing in chondrogenic medium. An increase in the oxygen level after chondrogenic differentiation may then stimulate ROS formation and commitment to the endochondral route.99,100 Indeed, late hypertrophic cells in the growth plate do not appear to be hypoxic as reflected by the absence of EF-5 binding (a marker for bioreductive activity) in this part of murine growth plates.22,23 Complementary to this observation, in vitro culture of a murine mesenchymal cell line under hypoxia suppressed terminal differentiation of chondrocytes. 101 This finding was also supported by the observation that angiogenesis and associated increased oxygen availability is required for endochondral ossification.43,54,102
All together, these data indicate that oxygen is an important tool for MSC expansion, cell cycle arrest, and differentiation pathways, including regulation of endochondral ossification of MSCs in vitro. Hypoxia may be imposed to stimulate maintenance of the chondrogenic phenotype and production of a cartilaginous matrix, followed by restoration of normoxia to allow regular transition to hypertrophy.
Mechanical modulation of MSC differentiation
Another key parameter that influences the differentiation of developing tissue and thus could be used for controlling construct differentiation in vitro is mechanics. Several modes of mechanical loading have been shown to affect cell proliferation, metabolism, and differentiation. As already known the mechanical properties of the surface to which cells attach determine the differentiation patterns.103,104 In the mechanical microenvironment, an interplay between the degree of attachment to a substrate and its stiffness orchestrates differentiation.104,105 In addition to the local mechanics, externally exerted forces are also transferred to the cells and affect their behavior. Compressive, tensile, shear, torsional, and hydrostatic forces are known to be able to change tissue differentiation.105–108 For example, the respective magnitudes of combined shear strain and fluid flow predict the final differentiation status of mesenchymal stem cells. If both are low, formation of mature woven bone is predicted, whereas more intense loading conditions favor cartilaginous or fibrous tissue formation.109,110
Mechanical stimulation of chondrogenically differentiated MSCs is typically performed after encapsulation of the cells in a gel. The gel provides increased mechanical integrity to the constructs, which enables mechanical stimulation of the cells. A wide range of gels are applied for chondrogenic 3D cultures, such as alginate, agarose, fibrin, and polyethylene glycol gels. In general, gel-loading protocols consist of 10–15% unconfined compression at 1 Hz in a preferably intermittent fashion.111–114 Intermittent compression of MSCs in a gel construct has been shown to enhance chondrogenic gene expression when compared with continuous dynamic compression. In addition, mechanical stimulation was shown to improve chondrogenic expression profiles even in the absence of transforming growth factor-β.112,113 This demonstrates that mechanical stimulation is a strong differentiation stimulus in 3D chondrogenic cultures. Nevertheless, contradictory results showing inhibition of chondrogenesis were also found with MSCs in a 2% agarose gel, subjected to 10% compression at 0.5 Hz for 1 h/day (5 days/week). 115 This long-term culture for 42 days indicates that small variations in construct preparation, cell source, and loading protocols can lead to large differences in outcome.
Also hypertrophic differentiation of chondrocytes has been shown to be affected by biomechanical cues. Cyclic tension, for example, was shown to upregulate expression levels of hypertrophic chondrocyte markers (mRNA) in chondrocytes in an alginate gel. 116 It was suggested that the stimulatory effects of loading on endochondral ossification are controlled by cbfa-1. Also the expression of collagen type X mRNA levels in hypertrophic chondrocytes seeded in sponges was enhanced in regions of high local tensile strains (9%) within 48 h. 117 Apart from cyclic tension, mild cyclic shear can accelerate endochondral ossification, whereas cyclic hydrostatic pressure increases chondrogenic expression.118–120 Besides these in vitro studies, the importance of mechanical loading during the ossification of long bones was demonstrated in explanted femoral condyles. Herein, no formation of a structure resembling the secondary ossification center was observed when no mechanical stimulation was exerted. In contrast, only 12 h of dynamic compression did induce formation of such a structure containing hypertrophic and mineralized regions. 121 The roles of mechanical cues have not been explored as yet in inducing and maintaining hypertrophic differentiation by MSCs. However, combined fluid shear and mechanical stimulation significantly enhanced osteogenic differentiation 122 of MSCs compared with purely perfused constructs.122–126 This may suggest that it would also enhance hypertrophic differentiation of MSCs. Finally, the effects of in vitro mechanical loading of constructs on in vivo performance after implantation have not been studied before.
Summarizing, it can be suggested that during the chondrogenic stage of MSCs, intermittent, cyclic compression will be most stimulating for matrix formation and differentiation, whereas upon endochondral progression, decreased loading magnitudes and increased fluid shear will stimulate the formation of a mineralized matrix.
Conclusion
Differentiation of MSCs through the endochondral route holds promise for the creation of larger osseous constructs by tissue engineering. The in vivo and in vitro hypertrophic tendency of chondrogenically stimulated MSCs is beneficial in the making of these constructs. More insight into the modulatory potential of the environmental cues, such as mechanical stimuli and oxygen tensions, on tissue development from adult mesenchymal stem cells and their maturation will result in culture conditions that provide better control of endochondral bone tissue engineering.
Several challenges remain to be addressed before endochondral bone tissue engineering is a reality. Still, the fate of the implanted cells has not been established. Although the containment of undifferentiated MSCs within the implant has only been observed for a period up to a month after implantation, the fate of predifferentiated MSCs has not been described.81,127 Another milestone will encompass the elucidation of the timely correct window of implantation. On the one hand, hypertrophic cells have increased metabolic demands compared with chondrocytes, which shortens their in vivo survival. But, on the other hand, they secrete angiogenic signals to enhance vascularization. Moreover, based on the few results from in vivo studies currently available, a certain degree of maturation of the cells beyond the chondrocytic stage can enhance in vivo endochondral bone formation. It is therefore expected that hypertrophy should be initiated in these cells prior to implantation to ensure progression of endochondral differentiation and secretion of appropriate angiogenic and osteogenic signals. Once functional vascularization is established, more oxygen will be available to finalize hypertrophic differentiation and to induce the transition of the cartilaginous anlage to a mineralized matrix and the conversion to bone. It is evident that both effects (cell survival and progression of endochondral bone formation) need to be balanced to establish the optimal window for implantation. Eventually, when these hurdles are overcome the feasibility of endochondral bone tissue engineering has to be established in a large animal model before clinical trials can be conducted.
Footnotes
Acknowledgments
This work is part of the University Medical Center Utrecht Strategic Program on Regenerative Medicine. The authors gratefully acknowledge the support of the Smart Mix Program of the Netherlands Ministry of Economic Affairs and the Netherlands Ministry of Education, Culture, and Science. Dr. L.B. Creemers was financially supported by the Dutch Arthritis Association. Dr. J. Malda was supported by a Veni Fellowship from the Dutch Technology Foundation (STW), Applied Science Division of Netherlands Organization for Scientific Research (NWO), and the Technology Program of the Ministry of Economic Affairs.
Disclosure Statement
No competing financial interests exist.