
Abstract
The physiological role of the actin cytoskeleton is well known: it provides mechanical support and endogenous force generation for formation of a cell shape and for migration. Furthermore, a growing number of studies have demonstrated another significant role of the actin cytoskeleton: it offers dynamic epigenetic memory for guiding cell fate, in particular, proliferation and differentiation. Because instantaneous imbalance in the mechanical homeostasis is adjusted through actin remodeling, a synthetic extracellular matrix (ECM) niche as a source of topographical and mechanical cues is expected to be effective at modulation of the actin cytoskeleton. In this context, the synthetic ECM niche determines cell migration, proliferation, and differentiation, all of which have to be controlled in functional tissue engineering scaffolds to ensure proper regulation of tissue/organ formation, maintenance of tissue integrity and repair, and regeneration. Here, with an emphasis on the epigenetic role of the actin cytoskeletal system, we propose a design concept of micro/nanotopography of a tissue engineering scaffold for control of cell migration, proliferation, and differentiation in a stable and well-defined manner, both in vitro and in vivo.
Introduction
I
A major role of the actin cytoskeleton is to provide mechanical support and a driving force for changes of the cell shape and for cell migration.6–9 In addition, the long-lived actin cytoskeletal structure is thought to serve as an epigenetic determinant of cell fate, 10 particularly, in relation to proliferation and differentiation. The cell modulates the delicate force balance between contractility of the actin cytoskeleton and exogenous mechanical forces transmitted across the cell–ECM adhesion points. 11 Instantaneous imbalance in the mechanical homeostasis is adjusted through reorganization of the actin cytoskeleton and of the transcellular adhesion. In this process, the micro/nanotopography of the ECM niche imposes structural constraints on the cell.12–14 This context gives the impression that the micro/nanotopography of the synthetic ECM niche can direct cell migration, proliferation, and differentiation during developmental processes in tissue- and organ-specific differentiation and morphogenesis through defining the spatiotemporal behavior of the endogenous actin cytoskeleton.
Currently, we are beginning to establish the concept of standard design of topography of a synthetic ECM niche. Although a growing number of studies have characterized the effects of the ECM topography on cell function and fate, the findings remain controversial. Such inconsistency is in part due to the biological complexity: different cell types have been used in different studies, although the response of cells to ECM topography is known to vary depending on a cell type.4,14–25
In this review article, we propose a design concept of the topographical features of the synthetic ECM niche by shedding light on the actin cytoskeleton. First, we present a Biophysical Basis for designing ECM micro/nanotopography by explaining the Mechanism via Which Cells Sense and Respond to ECM Topography. Second, we discuss the essence of Modulation of the Actin Cytoskeleton via Engineered Topographical Constraints. Then, in the “Control of Cell Function and Fate Through Modulation of the Actin Cytoskeleton” section, the studies on control of cell migration, proliferation, and differentiation are reviewed, with an emphasis on the role of the actin cytoskeletal system as an epigenetic determinant. In the last section, these insights are put together into a design concept of the topography of the synthetic ECM niche that gives cues for combinatorial control of cell migration, proliferation, and differentiation within functional tissue engineering scaffolds.
The Biophysical Basis: Mechanism via Which Cells Sense and Respond to ECM Topography
Figure 1 illustrates the major direct mechanical and indirect mechanochemical links between the ECM niche and cytoplasm that are necessary for cells to sense, integrate, transduce, and respond to mechanical perturbations caused by the topographical features of ECM. ECM topography affects the force balance between cells and ECM by constraining the cell, actin cytoskeleton, and integrin molecules at different levels. The mechanical perturbations are transmitted across the plasma membrane through transmembrane receptors directly (the right-hand part of Fig. 1) inside the cytoplasm and change conformation of the actin cytoskeleton, and/or are indirectly converted into biochemically relevant signals (the left-hand part of Fig. 1). This mechanism via which cells sense and respond to the ECM architecture is the basis of the “Modulation of the Actin Cytoskeleton via Engineered Micro/Nanotopographical Constraints” section and “Control of Cell Function and Fate Through Modulation of the Actin Cytoskeleton” section.
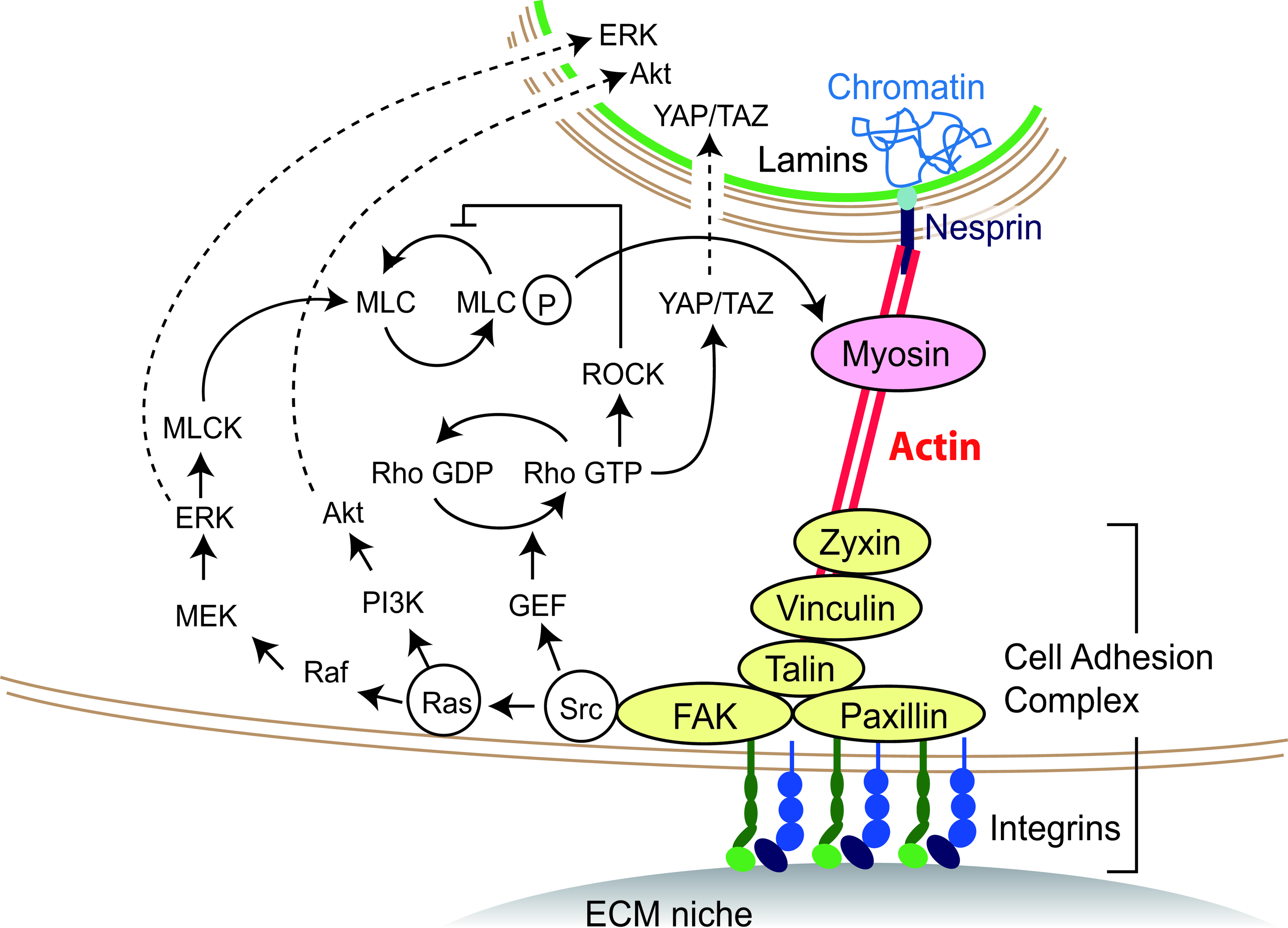
The mechanical and mechanochemical link of the cytoplasm to extracellular matrix (ECM) that contributes for cells to sense, integrate, transduce, and respond to the ECM architecture. The pathways mediated by the actin cytoskeleton are summarized based on the literature.11,44,53,162,163 The actin cytoskeleton transmits mechanical perturbations caused by ECM topography both directly (right-hand colored part) and indirectly (left-hand part) to the cell nucleus. Color images available online at www.liebertpub.com/teb
A direct mechanical link between the cytoplasm and the ECM niche through an integrin cluster and the actin cytoskeleton
At the interface of the cell and its extracellular environment, as illustrated in Figure 1, the cell adheres to ECM through molecular complexes at multiple parts of the cell membrane. Integrins bind to adhesive ligands of ECM via their extracellular domains, and on the intracellular side, their cytoplasmic domains are linked to the actin cytoskeleton via integrin-associated proteins.26,27 The profile of the assembled proteins, such as paxillin, focal adhesion kinase (FAK), talin, vinculin, and zyxin, determines the properties of the adhesive complex as a molecular clutch that links the actin cytoskeleton to ECM-bound integrins, whereby forces of different magnitude are transmitted from the cell to ECM and vice versa.28–30 The protein profile31,32 and the association and dissociation rates 33 are modulated by mechanical tension, which comes either from the myosin activity in the actin cytoskeleton or from external forces applied to the cell. An increase in the mechanical tension facilitates the recruitment and stable association of focal adhesion proteins required for the compositional maturation of integrin clustering, 34 which enhances the formation of well-bundled actin stress fibers and actomyosin contractility.35,36 In contrast, a decrease in the tension leads to the disorganization of the integrin cluster and of the actin cytoskeleton. 37
In the cytoplasm, the actin cytoskeleton is connected to the nucleus through inner nuclear membrane proteins, such as nuclear envelope spectrin repeat (nesprin).38,39 Nesprins interact across the luminal space with other nuclear envelope proteins, such as lamins, which form stable nuclear structures and can bind to chromosomal DNA.40,41 Through this link, the mechanical force that is directly transmitted between ECM and the nuclear interior changes nuclear morphology42,43 and intranuclear architecture, in particular, it can cause chromatin recombination, nuclear matrix distortion, and unzipping of DNA. 44 These nuclear changes alter transcription events and the nuclear transportation of soluble signaling intermediates.45,46
Indirect mechanochemical signal transduction
The cell adhesion complex, in addition to serving as the actin–ECM molecular clutch, works as a node to transduce mechanical forces acting on the cell–ECM adhesion complex into biochemically relevant signals, as shown in the left part of Figure 1. A major pathway in the nuclear relay of mechanical signals is YAP/TAZ signaling 47 for differentiation of human mesenchymal stem cells (MSCs) and for survival of human lung microvascular endothelial cells. 48 In addition, Ras/MAPK (Ras-Raf-MEK-ERK) signaling49,50 is a well-known pathway that controls the proliferation of human MSCs51,52 and cancer cells 53 and participates in keratinocyte stem cell maintenance. 52 Another downstream pathway of Ras is PI3K/Akt signaling, which is involved in self-renewal of mouse embryonic stem (ES) cells.54–58
Cell type-specific sensitivity to topographical features
The sensitivity of cells to topography depends on cell type.14–17,19,21,22,24,25,59,60 By using silicon curved cavities, different types of adhesion have been demonstrated between fibroblasts (Fig. 2A, B) and malignant cancer cells (Fig. 2C, D).16,19 The strength of adhesion and that of mechanical feedback that a cell senses on integrin ligation 61 are believed to determine how much the cell is constrained by ECM; thus, this factor will be a primary endogenous determinant of sensitivity of the cell to micro/nanotopographical features. Generally, transformed cells are insensitive to the topographical features because of a noticeable decline in the mechanical feedback, and they exhibit uncontrolled growth independent of the ECM architecture. 18 Thus, in designing topography of tissue engineering scaffolds, researchers should take into account exogenous parameters to determine how much the cell is constrained by the synthetic ECM.
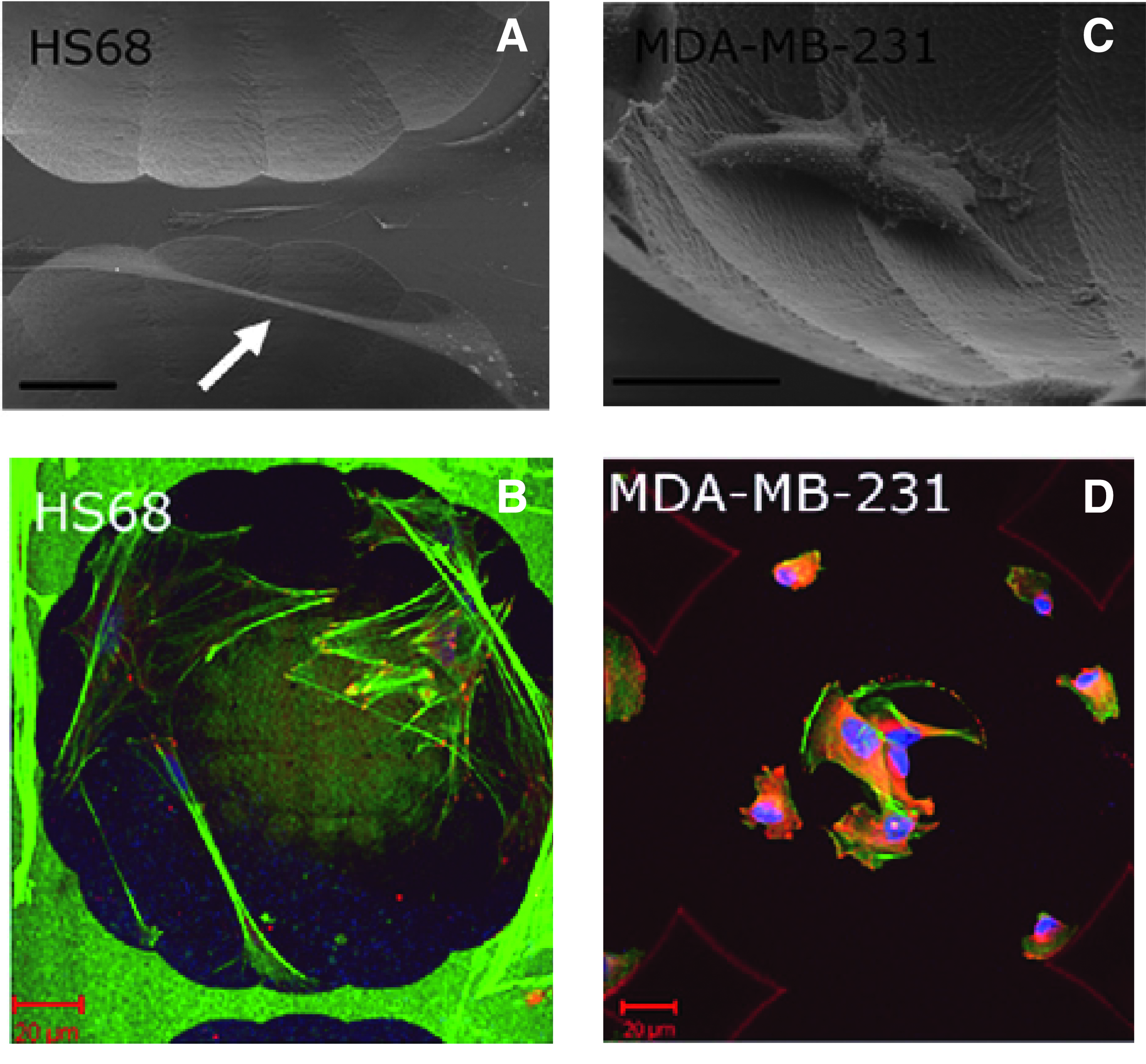
Different adhesion modes of different cell types in a silicon microchamber.
Modulation of the Actin Cytoskeleton via Engineered Micro/Nanotopographical Constraints
Topographical features of ECM serve as structural constraints at several levels. The features tens of micrometers in size act directly on a single cell or cells in a cell population (Fig. 3A). The features submicrometers to 10 μm in size act on the actin cytoskeleton (Fig. 3B), whereas features ten to hundreds of nanometers in size act on integrin molecules (Fig. 3C). Such varied structural constraints affect the shape of the cell–ECM interface and modulate the mechanical homeostasis between the cell and ECM; 13 this process leads to actin remodeling through downward and upward causation 62 in the mechanochemical interactions between factors of different size levels. Thus, the multiscale perspective on topographical feature is essential in engineering micro/nanotopography for the modulation of the actin cytoskeleton.
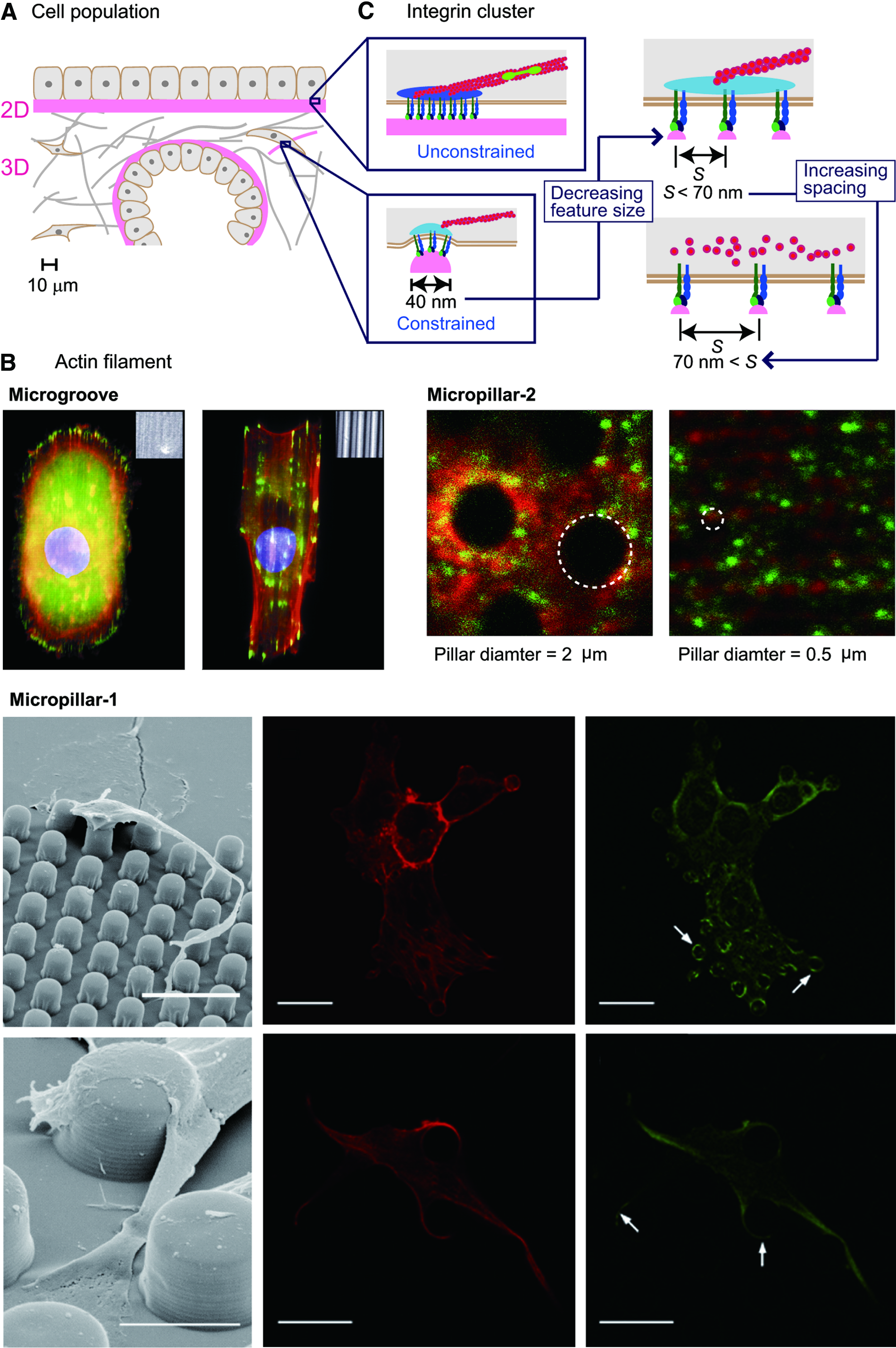
ECM topography as a structural constraint at multiple levels.
Topography on the scale of tens of micrometers: constraints on a single cell and a cell population
ECM topographical features over 10 μm in size correspond to cellular and multicellular levels, as shown in Figure 3A, and are determinants of the shape of a single cell or cells in a cell population.13,63,64 On this scale, the formation of actin stress fibers is promoted or limited for the adjustment of the mechanical homeostasis between the cell and ECM mainly in response to a change in the cell peripheral curvature defined by the topographical features.
Cells on conventional glass or plastic rigid planar surfaces, the so-called two-dimensional (2D) cultures, guide the organization of bundled actin stress fibers that link well-mature focal adhesions at the basal surface. Cells under this constraint experience tractions primarily as shear stresses on their basal surface and are thereby confined to the 2D surface and form a relatively planar interface with ECM. Cells on a dense layer of a 2D basement membrane65,66 and on mineralized bone or teeth are under a similar constraint.
In contrast, three-dimensional (3D) scaffold configuration, such as the loose fibrillar 3D meshwork present in most connective tissues,67,68 allows cells to migrate into the meshwork. Inside cells in the 3D scaffold configuration, actin stress fibers and focal adhesions are dispersed 69 because of the tensional homeostasis in response to the curvature of the plasma membrane. 13
Thus, the 2D scaffold configuration is better for guiding the organization of bundled actin stress fibers that link well-mature focal adhesions on the basal surface. In contrast, the 3D scaffold configuration is better for guiding actin stress fibers and focal adhesions to dispersal. Furthermore, the configuration of a 3D structure is preferable when researchers need additional effects, such as the control of diffusive transport70,71 and interstitial flow, 72 beyond the direct effects of the micro/nanotopography.
Topography on the scale of submicrometers to 10 μm: constraints on the actin cytoskeleton
The size of endogenous actin filaments fits the range of submicrometers to 10 μm. Thus, topographical features submicrometers to 10 μm in size work as direct constraints on actin filaments, the process that has long been recognized as “contact guidance.”4,73–75 Contact guidance explains why the probability of actin filaments' traversing the obstacles is low: due to the mechanical restriction of the cytoskeleton.
For instance, in cells on micrometer-sized parallel repeated groove-and-ridge structures, actin filaments align with the longer axis of the grooves (Fig. 3B, Microgroove).76–79 Accordingly, in cells within the regular array of micropillars, the actin filaments and focal adhesions are limited only at the top of the pillars (Fig. 3B, Micropillar-1, top).80–82 On the other hand, if there is enough spacing between the pillars, actin filaments are constrained between the pillars, and the focal adhesions are present only on the side and/or the bottom of the pillars (Fig. 3B, Micropillar-1, bottom). 81 The threshold of spacing that determines whether actin filaments are limited on the top of the pillars or between them is 10 μm in 3T3 fibroblasts. 81 The threshold of spacing is supposed to depend on the actin cytoskeletal flexibility specific to each cell type.22,25 The threshold of pillar diameter that determines whether the actin cytoskeleton is constrained on top of or between the pillars is reported to be 1 μm (Fig. 3B, Micropillar-2), which closely correlates with the minimum size of the actin–myosin contractile complex: a minimum length of ∼1 μm.83,84
Topography on the scale of ten to hundreds of nanometers: constraints on an integrin cluster
Topographical features ten to hundreds of nanometers work, as a direct constraint on integrins, as illustrated in Figure 3C. The critical size of the integrin cluster as a molecular clutch to transmit forces between ECM and the cytoplasm has been measured in an in vitro nanofabricated system.85–87 A cluster consisting of integrins spaced at 70 nm or less (Fig. 3C, right) acts as a molecular link for transmission of a force between ECM and the actin filaments. The smallest number of integrin molecules necessary to form the mechanical link is three 88 or four. 85 The smallest topographical feature that allows for integrin clustering with three or four integrin molecules is a spherical bead 40 nm in diameter. 89 Such a unit can be used as an element to specify the position and strength of a cytoplasm–ECM link. 87
Other representative elements—with effects on integrin clustering and actin cytoskeletal organization—are a nanodot, nanopit, and nanogroove.4,90–92 In cells on an array of these elements, the accessibility of a given region to the cell membrane is defined by the size of features of the element, such as width and height/depth, and by spacing between the elements. In general, more than 40 nm in height/depth and densely packed elements can restrict integrin molecules only on the top of topographical elements.87,90
An array of nanodots smaller than 100 nm in diameter can effectively interfere with integrin clustering and disorganize the actin cytoskeleton. 87 Too small a spacing between the nanodots restores integrin clustering by allowing endogenous integrin-associated proteins to connect to neighboring integrin domains limited on the top of the nanodots. 90 Increasing the spacing between nanodots above 1 μm increases the accessibility of the plasma membrane to the bottom surface and restores integrin clustering. 90 Similarly, an array of nanopits can affect integrins and the actin cytoskeleton. Nanopits ∼100 nm in diameter spaced at hundreds of nanometers effectively interfere with integrin clustering and disorganize the actin cytoskeleton.90,93–95
Cells respond to parallel grooves and ridges in a manner similar to the response to nanodots and nanopits. In a human corneal epithelial cell, lamellipodia at the cell edge perpendicular to the patterns can adhere to the bottom surface of the grooves that are 2100 nm wide but not 330 nm wide, on grooves that are either 150 or 600 nm deep. 77 Integrins in cells on a parallel 330 nm groove pattern are limited on the top of the ridge between the grooves. In contrast to an isotropic pattern of nanodots or nanopits, the anisotropic groove/ridge pattern guides integrins to cluster along the longitudinal direction of the ridges.77,87 Concomitantly, actin filaments align along this longitudinal direction.77,87,96
As outlined in this section, topography on the scale of ten to hundreds of nanometers functions as an effective local cue to regulate the integrin clustering and actin cytoskeletal reorganization in a well-defined manner. Designed for more sophisticated control of cell function and fate, hierarchical scaffolds, in which nanotopographical cues are incorporated into microstructure, benefit from both nanotopography and microtopography. The techniques for fabrication of such hierarchical scaffolds have now been developed.4,97–100 Their effectiveness in enhancing MSC adhesion and proliferation has been shown using a scaffold consisting of microscale strands with deposited micro/nano-sized fibers. 99 Microspheres with nanowires improve adhesion to a wide range of cell types and ensure excellent in vivo biocompatibility while retaining high loading capacity inherent in micron-sized particles. 100
Elasticity: an essential parameter for tuning effectiveness of micro/nanotopography as a structural constraint
As discussed in the “Cell Type-Specific Sensitivity to Topographical Features” section, the effectiveness of micro/nanotopography as a structural constraint varies by cell type because of cell type-specific sensitivity to topographical features. To cope with this cellular property in designing tissue engineering scaffolds, a design variable to tune how much the cell is constrained by micro/nanotopography should be taken into account. Elasticity of the micro/nanostructure on the scaffolds may serve as this sort of a parameter for fine-tuning the effectiveness of the micro/nanotopography as a structural constraint.
Mechanical stimuli have long been recognized as a significant factor in regulating tissue formation, maturation, and functionality. 101 Mechanical loading not only induces hypertrophy and strengthening of skeletal muscles, tendons, ligaments, and bones but also improves functionality of stem cell-derived tissues, for instance, a bioartificial cardiac tissue in vitro102,103 and tendon in vivo 104 . A wide variety of cell types, such as human MSCs,61,105,106 mouse ES cells, 107 fibroblasts,18,108,109 and glioma cells, 110 have been reported to respond to the ECM elasticity within the range of in vivo tissue elastic modulus from 0.1 kPa in the brain to 100 kPa in pre-calcified bone. 111 This property of cells can be used to specify strength of the cytoplasm-ECM link (Fig. 1), namely the effectiveness of the micro/nanotopography as a structural constraint.
On a substrate with low elastic modulus, cell–ECM adhesion spots, which are linked to an actively contracting actin cytoskeleton, move with the substrate deformation, and the compliant substrate reduces the stress in the adhesion spots to sustain the substrate deformation. 108 The decreased stress reduces the integrin clustering for normal maturation of focal adhesions, and as shown in Figure 4A, the actin cytoskeleton is weakly assembled in the cytoplasm, being concentrated predominantly in the cortex under the cell membrane.106,110,112 On the other hand, in cells on a stiff substrate, mechanical tension is sufficient to facilitate 113 the recruitment and stable association of focal adhesion proteins to mature focal adhesions. This process leads to the formation of large and well-defined actin stress fibers in the cytoplasm,106,110,112,114 as shown in Figure 4B. Within the range of elastic modulus, in order for the cells to sustain constant substrate deformation, the strength of the cytoplasm-ECM link can be modulated to increase with increasing elastic modulus. Under 3D conditions, the effectiveness of a structural constraint can be fine-tuned by changing its elastic modulus, although the tuning efficiency and sensitivity seem to be lower than under 2D conditions. 115
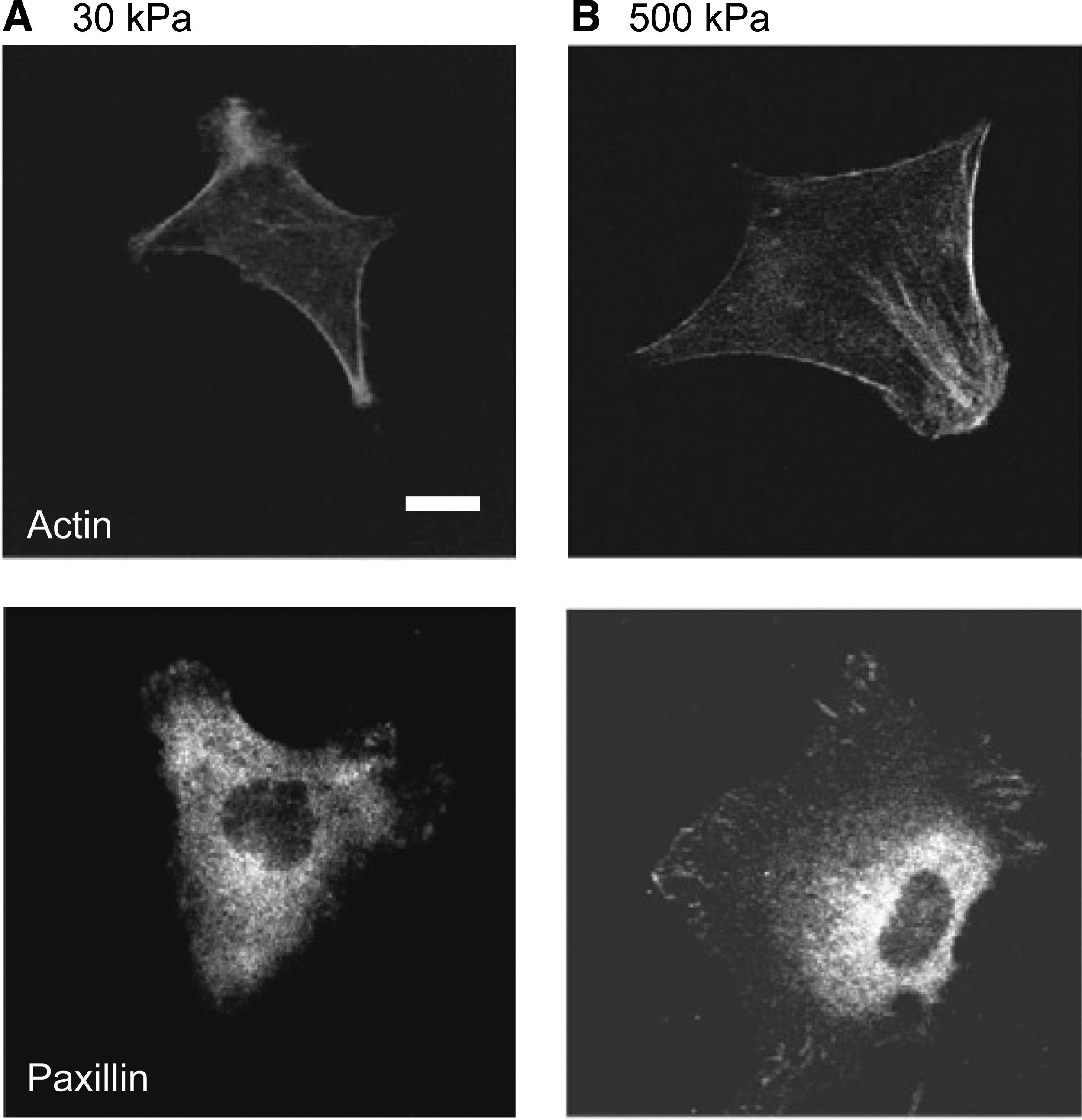
The cellular response to substrate elasticity. Human mesenchymal stem cells (MSCs):
Control of Cell Function and Fate Through Modulation of the Actin Cytoskeleton
Given a platform for modulation of the actin cytoskeleton by means of engineered micro/nanotopography as described in the “Modulation of the Actin Cytoskeleton via Engineered Micro/Nanotopographical Constraints” section, another critical step is to understand the control of cell migration, proliferation, and differentiation through the modulation of actin cytoskeleton. Combinatorial control of these processes potentially patterns complex cell mixtures into functional tissues and organs and also promotes physiological activity of one cell type while suppressing that of another. For instance, a technique that is used to maintain stem cell viability and proliferation, while promoting outward migration at an appropriate stage of differentiation, 111 has a significant therapeutic benefit for tissue regeneration in vivo.
Cell migration
Cell migration is essential for morphogenetic processes in tissue development, homeostasis, and regeneration. 1 Cell migration is driven by the protrusion at the leading edge, adhesion formation, generation of traction stress, and rear-edge retraction. 116 Although the same basic processes are involved, different cell types exhibit different modes of migration related to their specific functions. 21 The mode of cell migration was originally classified based on cell morphology (either rounded or spindle-shaped) and pattern (either individually, loosely connected, or collectively migrating): factors that are defined by actin cytoskeletal stiffness and polarity and by the type of cell–ECM interaction and force generation. 21 Individual cells, with an actin cytoskeleton predominantly in the cortex and migrating while loosely attached to the ECM, exhibit a rounded shape with amoeboid migration. In contrast, cells with well-bundled actin stress fibers—and migrating while firmly attached to the ECM through well mature focal adhesion—develop the spindle-shaped mesenchymal pattern of migration. Individual cells transiently form and resolve cell–cell contacts while moving along a structural frame provided by ECM (Fig. 5A).115,117,118 The maintenance of firm cell–cell adhesion can lead to confined migratory activity in cells inside a population, yet supports cytoskeletal activity at outward edges or at basal cell substrate. The resulting collective migration forms a multicellular spherical acinus or tube inside a structural constraint with a curved surface.63,64,119

The topographical constraint affecting both single-cell and collective cell migration.
Control of cell migration through actin modulation has been well characterized based on contact guidance by means of simple synthetic 2.5D microtopographical features, such as parallel ridges and grooves,73,96,120–124 arrays of micropillars,14,22,80,81 and asymmetric microchannels (Fig. 5B).125,126 In cells on a micro-/nano-grooved structure, actin filaments and vinculin-rich focal adhesions align with the longer axis of the grooves. This effect causes various cells, such as epithelial, endothelial, glial cells, fibroblasts, osteoblasts, and neutrophils, to align73,96,120,121 and migrate4,121,123,124 along the grooves. On an array of micropillars, cellular protrusions are guided preferentially to the pillars.22,80 Accordingly, cells placed on a microchannel with ratchets show directional migration and actin bundles connecting the points of highest curvature and a biased lamellipodial protrusion (Fig. 5B). 126
A density gradient is another anisotropic topographical feature that affects cell migration.79,127 In cells on rectangular lattice arrays with variable local density, actin stress fibers and focal adhesions are preferentially formed or stabilized in the denser local areas along the direction diagonal to the lattice. The cells migrate toward the topographically denser areas away from sparser ones. 127 Because effectiveness of the contact guidance can change with size variation in the topography on the scale of 1 μm, 14 a topographical density gradient can induce biased migration of both a single cell and a cell population (Fig. 5C).79,127
Cell proliferation
Actin cytoskeletal control of cell proliferation has been uncovered in studies of the effect of cell morphology in growth control.128,129 It has been shown in nontransformed endothelial cells and fibroblasts that increased DNA synthesis, and cell growth are tightly coupled to increased cell spreading. 128 Cell shape determines whether individual cells grow or die, regardless of the type of the protein of ECM in contact with the cell. 129 These studies have suggested that the mechanical tension-dependent changes in the cell shape govern cell proliferation. The best-characterized pathway is the one that involves stimulation by actin cytoskeletal tension of the Rho-ROCK (Rho-associated protein kinase) pathway and the resulting integrin clustering, which increases extracellular signal-regulated kinase (ERK) activity, leading to enhanced cell growth through ERK-dependent induction of cyclin D1.50,128,130,131 Thus, cell spreading, with well-bundled actin stress fibers and mature focal adhesions, enhances cell proliferation.
Consistent with the basic findings, as shown in Figure 6, in rat MSCs, self-assembled vertically oriented TiO2 nanotubes with a 15 nm diameter effectively accelerate integrin clustering and the formation of focal adhesion linked to actin stress fibers; the cell proliferation rate is high. 132 In contrast, tubes 100 nm in diameter disrupt integrin clustering and formation of focal adhesion. Compared to MSCs on tubes of 15 nm diameter, proliferation rates are over fivefold lower in those on the nanotubes of 100 nm diameter. In vascular smooth muscle cells, TiO2 nanotubes with an average pore diameter of 30 nm disrupt actin stress fibers and decrease cell proliferation compared to a flat surface (control). 133

Topography induces actin cytoskeletal changes affecting cell proliferation. Rat bone marrow MSCs on day 1 (middle row) and day 3 (bottom row) after plating on self-aligned TiO2 nanotubes 15 nm in diameter (top row, left) or 100 nm in diameter (top row, right). The cells were stained for actin filaments (green), paxillin (red), and nuclear DNA (blue). One and 3 days after the plating, stress fiber assembly and formation of focal adhesion are extensive on the 15 nm nanotubes (left) but greatly reduced on 100 nm nanotubes (right). Scale bars, 200 nm in the top row, 50 μm in the middle row, and 100 μm in the bottom row. (Adapted with permission from Park et al. 132 Copyright 2007 by American Chemical Society.) Color images available online at www.liebertpub.com/teb
Recent researches indicate more delicate control of cell proliferation can be achieved using mechanical signaling. Densely packed and sharp peaks of a fractal silicon nanostructured substrate promote proliferation of NIH3T3 fibroblasts, whereas more regular ridges favor adhesion but decrease the rate of proliferation. 134 It has been reported for various types of cells, such as NIH3T3 fibroblasts, primary low passage fibroblasts from human lungs, and human MSCs, that proliferation of cells on a soft (<6 kPa) substrate is tightly coupled with cell spreading but is decoupled from myosin light chain phosphorylation, cellular traction, and focal adhesion formation. 43 Actin and focal adhesion signaling with a Rho-ROCK-myosin effector for cell proliferation might not be the case for a synthetic ECM with pronounced nonbiomimetic topography and stiffness.
Stem cell differentiation
Maintenance of a specific stem cell population and guiding their directed differentiation to a desired lineage are necessary for tissue engineering and regenerative medicine. The conventional approach to controlling stem cell differentiation is the discovery and use of soluble biochemical cues and signaling pathways. 135 At the same time, a growing number of researchers have recognized the significant role of the ECM niche in a tissue where stem cells reside.111,136–139
The effect of actin cytoskeletal tension on commitment to a stem cell lineage has been best characterized for MSCs in many studies, 140 including those using the high-content image-based approach for the prediction of a lineage fate of stem cells.141,142 As summarized in Table 1, osteogenic lineage commitment and inhibition of differentiation into other lineage types require actin cytoskeletal tension mediated by RhoA and ROCK in human,61,106,143–145 mouse, 146 and rat 132 MSCs. In line with this observation, human MSCs on grooves or pits 300–500 nm deep and 5–50 μm wide (Fig. 7A) have an increased amount of stress fibers and a greater number of focal adhesions compared to a planar surface (control) and exhibit increased osteoblastic functionality. 147 Nanopits with different randomness (Fig. 7B) change the ability of human MSCs to form mature adhesion spots and stimulate the cells to produce bone mineral components in vitro in the absence of osteogenic supplements. 148 According to Table 1, commitment to the osteogenic lineage corresponds to well-bundled actin stress fibers as well as mature and elongated integrin clusters, but dispersal of these structures leads to lineage types other than osteogenic. Actin stress fibers are likely to be more dispersed (more diffuse) inside the cell during commitment to the following lineages (listed in the order from somewhat bundled to fully dispersed): myogenic, adipogenic, and neurogenic lineage commitment of human MSCs. The same is true of integrin clusters. In the neurogenic lineage, cytoskeletons and nuclei that are aligned and elongated along the nanogratings show significant upregulation of neuronal markers in human MSCs in neuronal induction media. 149
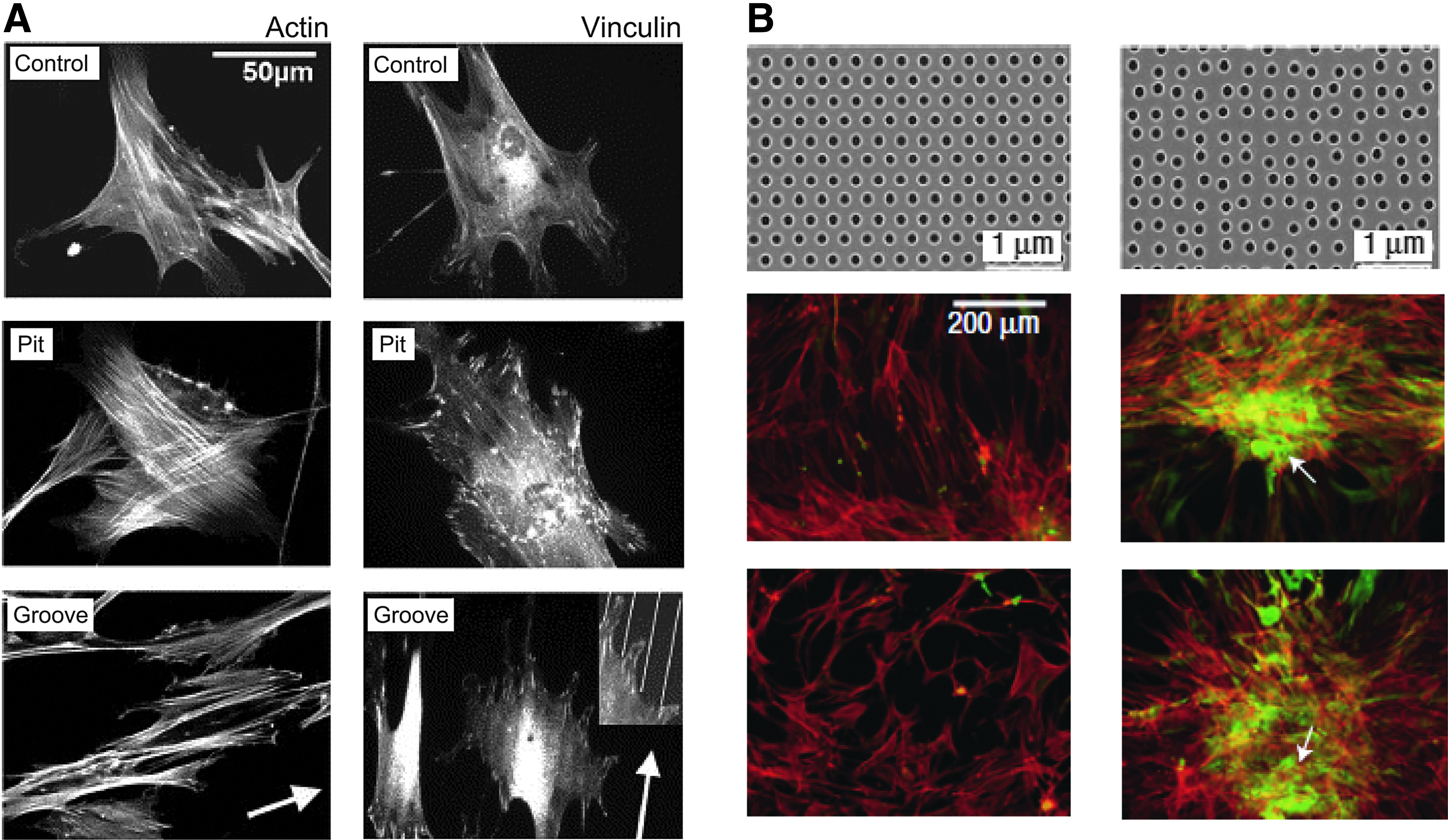
Topography induces actin cytoskeletal changes affecting stem cell differentiation.
ERK, extracellular signal-regulated kinase; ES, embryonic stem; FAK, focal adhesion kinase; iPS, induced pluripotent stem; JNK, c-Jun N-terminal kinase; MAPK, mitogen-activated protein kinase; MSC, mesenchymal stem cell; ROCK, Rho-associated protein kinase.
Differentiation of ES cells in relation to actin cytoskeletal organization and integrin clustering has also been characterized (Table 1). In human ES cells, nanopatterned substrates that downregulate cell adhesion upregulate ES self-renewal properties (Tables 1 and 2). 150 Similarly, in mouse ES cells, nanopatterned substrates that downregulate cell adhesion and spreading upregulate ES self-renewal properties; these effects have been shown to result from integrin-mediated FAK signaling to suppress the ERK/JNK activity (Tables 1 and 2). In contrast, mouse ES cells with upregulated stress fiber formation and cell adhesion exhibit enhanced mesendoderm differentiation (Table 1). 107 The integrin/FAK-mediated commitment of ES cells (both human and murine) seems to follow logically from the data presented in Tables 1 and 2 (also in the “Topography on the Scale of Ten to Hundreds of Nanometers: Constraints on an Integrin Cluster” section) because there are three lines of evidence: First, topographical features tens to hundreds of nanometers in size (nanotopography) are thought to work effectively as constraints on integrin clustering (thus, FAK activity; “Topography on the Scale of Ten to Hundreds of Nanometers: Constraints on an Integrin Cluster” section). Second, integrin/FAK signaling affects the differentiation of ES cells (Table 1). Third, the fate of ES cells is affected by the nanotopography of ECM in both mouse and human ES cells (Table 2). Concerning the neuronal lineage, it was demonstrated in human ES cells that the elongation of the cytoskeleton during changes in the cell morphology guided by ridge/groove patterns results in a transfer of the tensional force to the nucleus, influencing gene expression and signal transduction in the absence of a differentiation-inducing reagent. 151
PDMS, polydimethylsiloxane.
More sophisticated control of cell fate is achieved by fine-tuning of the balance between proliferation and differentiation.132,152,153 A higher survival rate of human ES cells and human induced pluripotent stem (iPS) cells in dissociated culture is the result of inhibition of ROCK-dependent hyperactivation of actomyosin.154,155 Protocols involving ROCK inhibitors for dissociated human ES cells and human iPS cells have already improved a number of practical procedures.154,155 Furthermore, the degree of actin cytoskeletal tension mediated by ROCK plays a major role not only in directing stem cell differentiation to a specific cell type but also in retention of multipotency by stem cells (Table 1). 152 Some micro/nanotopographical features can increase cell proliferation concomitantly with the enhancement of osteogenic differentiation,152,153 and others have decoupled effects, 153 although the detailed mechanism remains to be clarified. Further studies of pathways and molecules involved in actin cytoskeleton-mediated control of stem cell fate will open up new ways of optimizing the design of synthetic ECM micro/nanotopography, which might be more effective than the natural one for differentiation control and stemness maintenance.
Summary, Discussion, and Future Perspectives on Topography Design Concept of Tissue Engineering Scaffolds
By reviewing and putting together previous insights, Figure 8 summarizes the deduced design concept of different types of micro/nanotopography of a synthetic ECM niche on functional tissue engineering scaffolds that are intended to provide cues for controlling targeted cell function and fate. In the “Control of Cell Function and Fate Through Modulation of the Actin Cytoskeleton” section, our overview of the endogenous and exogenous regulation mechanisms of cell migration, proliferation, and differentiation—with an emphasis on the actin cytoskeletal system—shed light on the role of the actin cytoskeleton as dynamic epigenetic memory. 7 According to this notion, we believe that control of cell function and fate can be achieved by separating the design procedure into two steps mediated by the modulation of the actin cytoskeletal system.
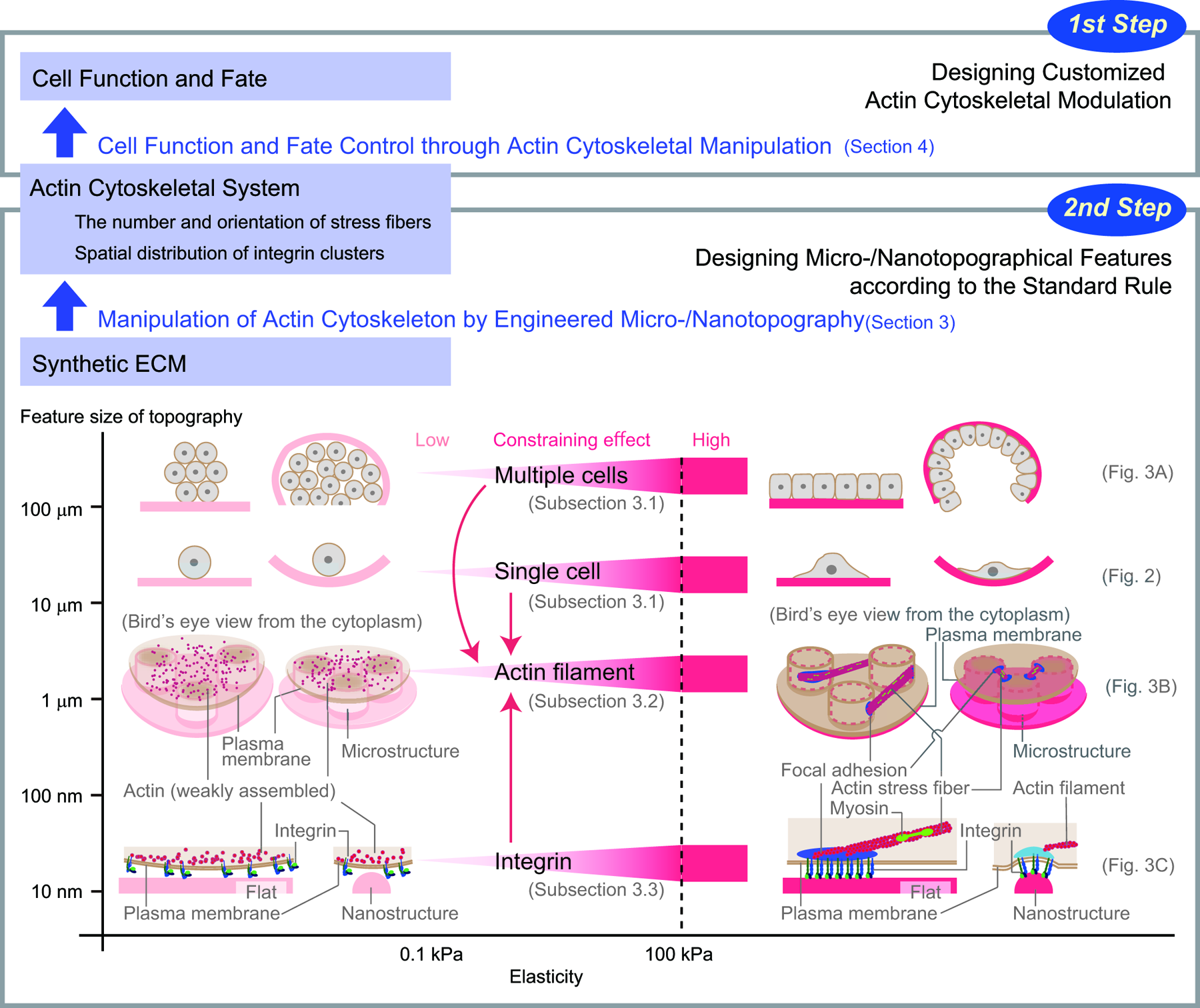
Topography design concept of a tissue engineering scaffold. Micro/nanotopography controlling cell function and fate is designed in two steps. The standard rule for the topographic design deduced in the “Modulation of the Actin Cytoskeleton via Engineered Micro/Nanotopographical Constraints” section is summarized in the lower boxed area. The number and orientation of the actin stress fibers and spatial distribution of the integrin clusters can be manipulated precisely by micro/nanotopography designed based on the standard rule. Elastic modulus (horizontal axis) is a fundamental parameter that affects the effectiveness of topographical features as constraints. By fine-tuning the elasticity, we can deal with the cell type-specific sensitivity to topographical features and achieve a desired effect on each cell type. The size of topographical features (vertical axis) determines the size of a substance affected. The red arrows indicate downward and upward causation for actin remodeling between different levels in the interactions between size levels. In designing the topographical features, the effect of such interactions should be carefully considered. Color images available online at www.liebertpub.com/teb
Summary of the topography design concept: a two-step design procedure based on modulation of the actin cytoskeletal system
As indicated in Figure 8, the first step is to implement customized modulation of the actin cytoskeleton to control cell migration, cell proliferation, and stem cell differentiation, which are critical processes in tissue engineering, for each medical application (the upper boxed area). The design strategy for modulation of the actin cytoskeleton involves reproducing the actin cytoskeletal behavior that occurs in the course of a desired cell function and fate based on the analysis of how the actin cytoskeletal system defines the desired cell function and fate. As summarized in the “Control of Cell Function and Fate Through Modulation of the Actin Cytoskeleton” section, the common feature of the actin cytoskeletal control of cell migration (Fig. 5), proliferation (Fig. 6), and differentiation (Fig. 7 and Tables 1 and 2) is the modulation of the number and orientation of actin stress fibers and of the spatial distribution of integrin clusters at the sites of stress fibers' anchoring to ECM.
Once the actin cytoskeletal modulation is implemented, the second step is to design micro/nanotopographical features according to the standard rule (the lower boxed area in Fig. 8) deduced from the discussion in the “Modulation of the Actin Cytoskeleton via Engineered Micro/Nanotopographical Constraints” section. As reviewed in the “Topography on the Scale of Tens of Micrometers: Constraints on a Single Cell and a Cell Population” section, “Topography on the Scale of Submicrometers to 10 μm: Constraints on the Actin Cytoskeleton” section, and “Topography on the Scale of Ten to Hundreds of Nanometers: Constraints on an Integrin Cluster” section, in the standard rule, micro/nanotopographical features are utilized as a structural constraint at multiple levels (based on the size of topographical features) for the modulation of the actin cytoskeleton. In addition, as suggested in the “Elasticity: An Essential Parameter for Tuning Effectiveness of Micro/Nanotopography as a Structural Constraint” section, the effectiveness of a structural constraint can be tuned by its elasticity. A greater effect of the structural constraint could be achieved by increasing the elastic modulus within the range of soft tissues: between 0.1 and 100 kPa. 111
This two-step methodology reduces the problem of diversity, which results from a broad range of medical requirements, to the subject of modulating the actin cytoskeletal system. This approach is expected to produce micro/nanotopography of a tissue engineering scaffold without comprehensive screening but after clarification of the role of the actin cytoskeleton in the regulation of the desired cell function and fate.
The custom-designed micro/nanotopography has an advantage as a cue for stable control at high spatial resolution. 91 In tissue engineering, it is necessary to control cell migration, the balance between proliferation and apoptosis, and stem cell differentiation. These characteristics of cell behavior are controlled not only by the presence or absence of guiding cues but also by their absolute and relative amounts and by spatial arrangement at the subcellular resolution. Different types of micro/nanotopography effectively supply local quantitative cues to cells in a stable and well-defined manner.
Discussion of application of the design concept to engineering of an implantable scaffold
Living organisms have developed mechanisms to control cell function and fate through the establishment of complex spatial and temporal distribution of diffusible factors during tissue development and regeneration.156–158 Insoluble ECM proteins,1,138 such as collagens, noncollagenous glycoproteins (elastin, fibronectin, nephronectin, or laminin), and hydrophilic proteoglycans with large glycosaminoglycan side chains serve as a local cue in synergy with soluble factors. Similarly, micro/nanotopography supplies local cues for stable control at high spatial resolution. 91 In vivo effectiveness of micro/nanotopography has been shown, for instance, in the regulation of phenotypic behavior of endothelial cells by means of topographical replicas of the basement membrane 159 and in cancer cell invasion, metastasis, and progression by means of 3D connective-tissue models.3,67,116
As expected, an implantable scaffold and devices with well-defined micro/nanotopography can provide local cues effectively. Micro/nanotopography of osteoconductivity of a porous titanium implant affects efficiency of bone formation in vivo. 160 Aligned electrospun nanofibers work as an instructive microenvironment for tendon stem/progenitor cells for tendon differentiation in vivo. 104
Our two-step design procedure based on the modulation of the actin cytoskeletal system can be applied to engineering of an implantable scaffold for use in vivo. To design customized modulation of the actin cytoskeleton at the first step, researchers can analyze in vivo behavior of the actin cytoskeleton using techniques, such as in vivo imaging 161 and fabrication of an in vivo topographical replica. 159 At the second step, the standard rule, which is for the design of micro/nanotopographical features to achieve the customized modulation of the actin cytoskeleton, can be applied in the same way regardless of whether one is dealing with in vivo or in vitro cues. The custom-designed micro/nanotopography effectively provides local quantitative cues to cells in a stable and well-defined manner in vivo and in vitro.
Future perspectives: discovery of ultimate effects of topography of a tissue engineering scaffold
An expected outcome of the micro/nanotopography developed based on the design concept presented here is that it might involve external nonbiomimetic cues, which possibly give excellent control of cell function and fate beyond the effect of the topography naturally provided by living organisms. It is now clear that this ultimate effect of the synthetic micro/nanotopography will be the result of putting together the cellular biophysical insights (collected at the first step in the concept shown in Fig. 8) and the result of refinement of the engineering standards of topographic design at the second step.
Current significant challenges for the acceleration of the discovery of the ultimate effect of the micro/nanotopography are a fundamental understanding of the role of the actin cytoskeletal system as an epigenetic determinant, and mathematical formulation of the qualitative standard rule that is shown in Figure 8 for topographical design. Further challenge to fully elicit the potential of the designed micro/nanotopography of a tissue engineering scaffold is to understand how to utilize the local microenvironmental effect of the topography in synergy with the soluble and highly diffusible factors that are most effective for global and systemic control.
Footnotes
Acknowledgments
This work was partially supported by the Japan Society for the Promotion of Science (JSPS) through the Funding Program for Next Generation World-Leading Researchers (LR017) and by Shiseido Female Researcher Science Grant.
Disclosure Statement
No competing financial interests exist.