
Abstract
In cartilage, chondrocytes are embedded within an abundant extracellular matrix (ECM). A typical chondron consists of a chondrocyte and the immediate surrounding pericellular matrix (PCM). The PCM has a patent structure, defined molecular composition, and unique physical properties that support the chondrocyte. Given this spatial position, the PCM is pivotal in mediating communication between chondrocytes and the ECM and, thus, plays a critical role in cartilage homeostasis. The biological function and mechanical properties of the PCM have been extensively studied, mostly in the form of chondrons. This review intends to summarize recent progress in chondron and chondrocyte PCM research, with emphasis on the re-establishment of the PCM by isolated chondrocytes or mesenchymal stem cells during chondrogenic differentiation, and the effects of the PCM on cartilage tissue formation.
Introduction
I
In cartilage, the PCM is best studied in the form of chondrons (Fig. 1). A (typical) chondron consists of a chondrocyte and its surrounding PCM. 7 The term “chondron” was coined nearly a century ago, but it was a series of studies accomplished by Poole that introduced chondrons to cartilage biology. Since a comprehensive review of chondrons published in 1997, 4 research on chondrons and chondrocyte PCM has made great advances. For example, the protocol of enzymatic chondron isolation developed by Lee et al. 8 makes large amounts of viable chondrons for biological and biochemical characterization possible. This review summarizes our current understanding of chondron/PCM properties and biology, and their pathological implication in cartilage degradation and potential applications for cartilage tissue engineering.
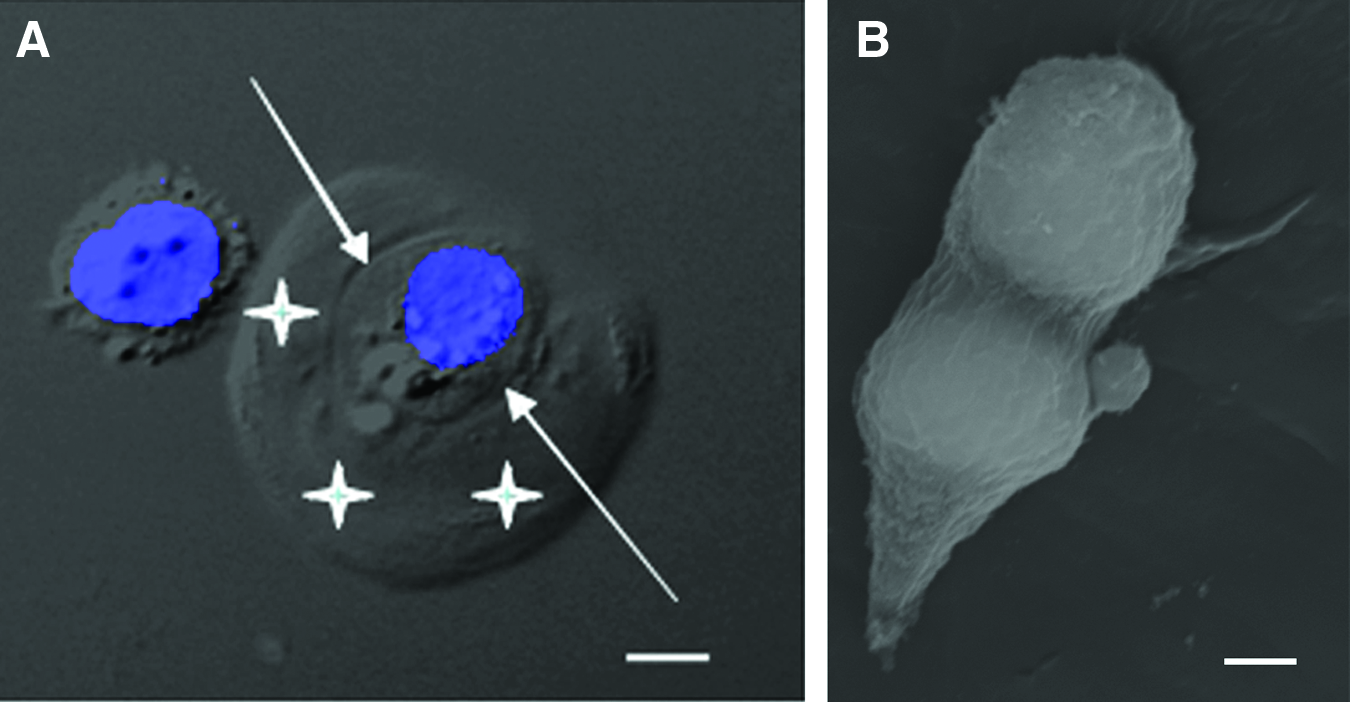
Enzymatically isolated chondrons.
The Properties of Chondrons/the PCM
The isolated chondrons preserve the integral feature of cartilage: chondrocytes and matrix. Because of the retention of interactions between the PCM and chondrocytes, chondrons have demonstrated functions/properties that are distinct from isolated chondrocytes. Chondrons are ideal for studying the physical and biological contributions of the PCM, without the interference from the greater ECM.
The physical properties of chondrons
Various biomechanical and mathematical models, including direct compression on chondrons in suspension, 9 revealed many of the mechanical properties of chondrons/PCM under specified conditions. For instance, the average compression force required to rupture chondrons and chondrocytes was 3.1 versus 2.1 μN, respectively. 10 The Young's modulus of enzymatically isolated chondrons was averaged at 1.54 kPa, while it was 0.6 kPa for chondrocytes in the same setting. 11 When isolated chondrons and chondrocytes were embedded in agarose gel (4%) and the gel was compressed to 20%, mechanically isolated chondrons demonstrated minimal deformations in the directions either parallel or perpendicular to the force; whereas the control chondrocytes were deformed. 12 This suggests that the PCM of mechanically isolated chondrons is stiffer than the 4% agarose gel. By finite element analyses, collagen fibrils in the PCM were found to be largely responsible for retaining the width and volume of the enclosed chondrocytes during compression. 13 The general consensus is that, because of the addition of PCM, chondrons are stiffer than chondrocytes.
From the joint surface to the junction with subchondral bone, articular cartilage is organized into several zones. In general, the size of chondrons and the volume of PCM increase from the superficial zone to the deep. 14 A finite element analysis revealed that, in different cartilage zones, the stiffness of the PCM collagen fibrils differentially impacts the shape and volume of the enclosed chondrocytes. From zone to zone, however, the fixed charge density in the PCM has little influence on chondrocyte volume. 13 When cartilage explants were compressed 10%, 30%, and 50%, the volume of the PCM and the height of chondrons in each zone were reduced in general but only the PCM in the superficial zone responded uniformly. 15 Finite element analyses suggest that the molecular composition, especially collagen and proteoglycan contents, contributes to the mechanical properties of the PCM. 16 Mathematical models and mechanical testing have shown that, in different cartilage zones, chondrocytes and PCM compensate for each other in stiffness to resist deformation.15,17
Although the isolated chondrons enable precise measurements of the mechanical properties of the PCM, it is noteworthy that a chondron isolation procedure inevitably alters the PCM properties to a certain degree. Atomic force microscopy was used to measured PCM properties in situ on cartilage sections. 18 The Young's modulus of the PCM was about one thirds of that in the interterritorial ECM. Compared with interterritorial ECM, the PCM had reduced contour roughness, indicating a relatively uniform distribution of mechanical properties in the PCM region.
The PCM is a physical barrier/filter for the molecules that enter or exit the chondrocytes. Using scanning microphotolysis, Leddy et al. measured the diffusion coefficients of dextran (molecular weight 70 and 500 kDa) in the PCM and ECM. 19 In the middle zone of the cartilage, the diffusion coefficient of 70 kDa dextran was 19 μm2s−1 in the PCM and 23 μm2s−1 in the ECM, respectively. The diffusivity of 500 kDa dextran was also slower in the PCM than in the ECM. When chondrons were challenged with varied osmolarities, the chondrocyte volumes in the chondrons were much less fluctuated than that of the isolated chondrocytes. 20 Thus, to a certain degree, the PCM acts as a buffer zone for the enclosed chondrocytes.
The molecular property/composition of the PCM
In addition to type II collagen and proteoglycans, type VI collagen is a major structural element of the PCM. 21 Unlike other collagens, type VI collagen is assembled into tetramers intracellularly and has no aldehyde-mediated cross-linking.22,23 The fibrils of type VI collagen form an independent network in most of the connective tissues, but are exclusively located in the PCM in cartilage. 24 Type VI collagen is often used as a marker of chondrons. 4
A complex of biglycan, decorin, and βig-h3 promotes the aggregation of type VI collagen and the formation of a fibril network in the PCM. 25 Biglycan and decorin connect type VI collagen fibrils with matrilins, which, in turn, link to aggrecan and type II collagen, the major structural molecules of cartilage ECM. 26 This molecular complex, including type VI collagen, biglycan, decorin, and matrilins, is the building block of the PCM and also interacts with other interstitial components of the PCM. The PCM is also considered a growth factor repository, where growth factors are activated, degraded, or transported. 27 For example, fibroblast growth factor 2 (FGF2) participates in mechanical signal transduction and is co-localized with perlecan in the chondrocyte PCM. 28
The basement membrane is a two-layer “membrane-like” structure and functions in tissue compartmentalization. 29 During embryonic development, basement membrane proteins are expressed in all three germ layers, including the mesoderm where cartilage arises. 30 A typical basement membrane structure has not been identified in the PCM but individual basement membrane proteins, including laminins, nidogens, collagen type IV, and perlecan, have been identfied.31,32 Although the roles of basement membrane in cartilage development and biology are still to be defined, basement membrane proteins are important in chondrocyte biology and for the PCM mechanical properties. For example, perlecan and laminin are known to facilitate chondrocyte attachment in vitro. 33 Inside the PCM defined by immunohistochemistry of type VI collagen, perlecan is localized in the inner two/thirds. 34 Wilusz et al. found that the elastic modulus in the inner PCM region, where perlecan distributes, was lower than the peripheral PCM region. When cartilage was treated with heparinase III, which specifically degrades perlecan, the elastic modulus of the inner PCM region increased, while it was unchanged in the peripheral PCM region. This detailed molecular dissection demonstrated that perlecan regulates the mechanical properties of the PCM.
Various proteins are continuously being added to the list of PCM components. For example, WARP (von Willebrand factor A domain-related protein) has been recently found to bind to perlecan and type VI collagen in the PCM.35,36 Considering the level of signal exchanges between chondrocytes and the ECM, it is reasonable to speculate that the catalog of PCM molecules is much larger than what is currently known. Proteomics is a powerful tool for studying cartilage biology. The proteomes of “bulk” ECM37,38 or chondrocytes, 39 however, have provided little information about the PCM, largely because PCM proteins could be either “masked” by the abundant ECM proteins or depleted during chondrocyte isolation. By comparing the proteomes of isolated chondrons and chondrocytes, the differential protein profiles comprise mainly PCM proteins. Using this approach, proteins extracted from human chondrons and chondrocytes were used for two-dimensional electrophoresis. 40 Compared with the chondrocyte gels, unique protein spots on the chondron gels were processed for mass spectrometry. This led to the identification of novel PCM proteins, including triosephosphate isomerase, transforming growth factor β (TGFβ)-induced protein, peroxiredoxin-4, ADAM (A disintegrin and metalloproteinases) 28, and latent-TGFβ-binding protein-2. These proteins were localized in the chondrocyte PCM with immunohisto(cyto)chemistry, but their biological roles are still to be determined.
The PCM Biology
The influences of PCM on chondrocytes have been observed in chondrons. Using cDNA microarray, the gene expression profiles of chondrons and chondrocytes were comparatively analyzed. 41 A total of 258 genes involved in chondrocyte proliferation, phenotype and metabolism were differentially expressed in chondrons. According to their expression patterns, those genes formed 10 clusters. For example, the expression of a cluster of 42 genes was similar between chondrons and chondrocytes at day 1 of culture but significantly increased in chondrons at days 3 and 5 (Fig. 2A). In addition, genes clustered together because of a downregulation pattern in chondrons after 3–5 days of culture (Fig. 2B). Clearly, the PCM has a profound impact on the functionality of chondrocytes.
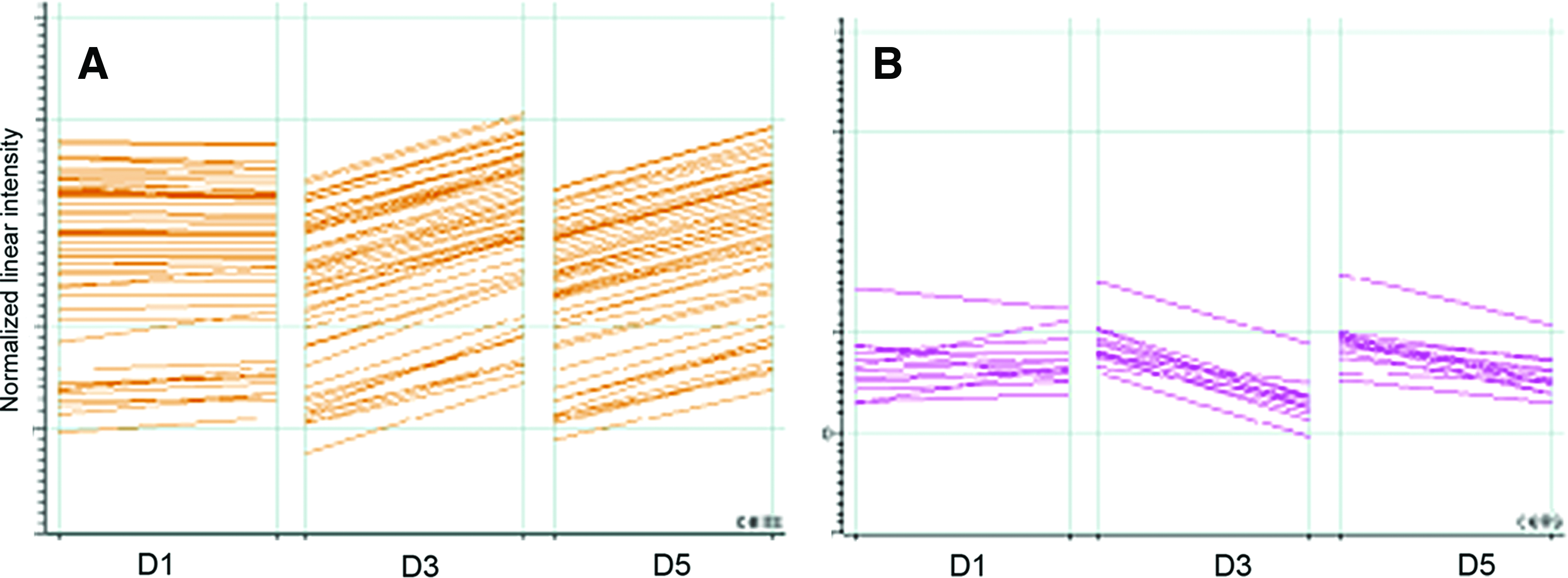
Gene clusters.
Isolated chondrons are more active in matrix deposition than chondrocytes alone. When chondrons and chondrocytes were cultured in pellets for 8 weeks, the physical size of the chondron pellets was visibly larger than the chondrocyte pellets. 42 Indeed, the amounts of type II collagen and proteoglycans in the chondron pellets were greater than in chondrocyte pellets. Furthermore, in response to dynamic compression, chondrons upregulated the expression of type II collagen and aggrecan to a greater extent than chondrocytes.43,44 The intact PCM also balances the anabolic and catabolic statuses of chondrocytes. In comparison with chondrocytes, the cultured chondrons expressed much less matrix metalloproteinase (MMP)-2, 9, and 13.45,46
PCM molecules are closely associated with chondrogenic differentiation of stem cells and the differentiation of chondrocytes. For example, domain I of perlecan promoted the chondrogenic differentiation of an embryonic fibroblast cell line in vitro. 47 Perlecan is the binding site of several growth factors, including bone morphogenetic protein 2 (BMP-2). Through BMP-2 signaling, perlecan regulates chondrogenic differentiation and endochondral ossification.32,48 Perlecan promotes the activation of vascular endothelial growth factor receptor (VEGFR) on the endothelial cells and enhances vascular endothelial growth factor (VEGF)/VEGFR signals, which then direct vascular invasion into the cartilage template during endochondral ossification. 49
Type VI collagen, a major component of the PCM, stabilizes chondrocyte phenotypes and is critical for chondrocyte survival. The expression of type VI collagen is diminished after chondrocyte dedifferentiation and restored during chondrocyte redifferentiation. 50 Chondrocyte apoptosis is common in cartilage turnover and aging, and may lead to cartilage degeneration.51,52 Detachment of cells from the matrix disrupts integrin-ligand interactions, which could, in turn, activate cell death pathways.53,54 Unlike the isolated chondrocytes, which are completely detached from cartilage matrix, the isolated chondrons retain attachment of the PCM to chondrocytes. While isolated chondrocytes are vulnerable, chondrons have shown resilience to apoptosis induction. Monoiodoacetate (MIA) treatment induced 1.6% of chondrons to undergo apoptosis, compared with 9% of chondrocytes. 55 Similar results were achieved when staurosporine was used for inducing apoptosis in cultured chondrons and chondrocytes. Furthermore, individual PCM molecules, such as type VI collagen, have been shown to protect cells from apoptosis. 56 In a separate experiment, isolated chondrocytes were treated with soluble type VI collagen before or after induction of apoptosis. 55 Preincubation of type VI collagen effectively prevented chondrocytes from MIA-induced cell death. After apoptosis was induced with MIA, however, treatment with type VI collagen failed to rescue chondrocytes from death.
There may also be other mechanisms supporting chondrocyte survival in chondrons. A group of heat shock protein (Hsp) 70 genes are upregulated in chondrons. 41 Hsp and its co-chaperones promote cell survival by suppression of caspase activation 57 and, thus, inhibit both the mitochondrial (intrinsic) and the death receptor (extrinsic) cell death pathways. 58 Overexpression of Hsp70 alone reduced nitric oxide-induced chondrocyte apoptosis from 29.3% to 0.41%. 59 In fact, chondroprotective reagents, such as glutamine, function through upregulation of Hsp70. 60 The anti-apoptotic effects of Hsp70 on chondrocytes were also demonstrated in vivo. 61 Increased expression of Hsp70 is beneficial for chondrons by conferring resistance toward apoptotic stressors.
The PCM molecules are positioned to translate mechanical signals to biological events. Matrilins connect to the cell membrane, via integrin, and other ECM molecules, such as types II and IX collagen. 26 Matrilin-1 and -3 are mainly found in cartilage. 62 It is well established that mechanical forces regulate chondrocyte differentiation and proliferation. For instance, mechanical stimulation applied to chondrocytes upregulates the expression of Indian hedgehog, which is a key signal in chondrocyte fate determination and proliferation. 63 This upregulation was found to be dependent on domain A of matrilin-3. 64 The study demonstrated a pivotal role of matrilins in translation of mechano-biological signals in the PCM.
The regulations of PCM proteins have not been fully understood. Matrilin 1 and 3 are regulated by chondrogenic transcription factor SOX9,65,66 indicating the great value of PCM proteins in defining the chondrocyte phenotype. Similar to most of the matrix proteins in cartilage, the expression of type VI collagen is downregulated by hydrostatic pressure 67 and influenced by the presence of other matrix proteins. 68 The accumulation of perlecan in the PCM of cartilage is believed to be due to increased stability, rather than due to issue-specific upregulation of the expression of perlecan. 32
The PCM in Cartilage Degeneration
The prominent role that the PCM plays in cartilage biology determines its inevitable involvement in cartilage degeneration, such as osteoarthritis. Membrane-type MMPs are proteases that are tethered to the cell membrane and function in the PCM domain for pericellular collagen degradation. 69 MMP-7 is activated by membrane protein CD151 and exhibits increased proteolytic activities in the PCM of osteoarthritic cartilage.70,71
During cartilage degeneration, there are a few notable changes in chondrons. For example, chondrons isolated from osteoarthritic cartilage have a reduced Young's modulus as compared with chondrons from normal cartilage. 11 Further, a relatively low PCM diffusivity demonstrated in normal cartilage is diminished in the cartilage of early osteoarthritis. 19 Thus, the physical properties of the PCM are significantly altered during cartilage degeneration.
Paradoxically, the size of chondrons and PCM density in osteoarthritic cartilage are generally increased, compared with that seen in normal cartilage. 72 In a set of measurements, the average cross-sectional area was 439 μm2 for chondrons isolated from osteoarthritic cartilage and 344 μm2 for chondrons from normal cartilage. 73 In the enlarged chondrons, both keratin sulfate and type VI collagen were increased in the PCM. When treated with TGFβ, the PCM area of chondrons isolated from osteoarthritic cartilage was greater than that of chondrons from normal cartilage. 72 The amount of pericellular perlecan is increased in osteoarthritis, particularly around dedifferentiated chondrocytes. 74
In osteoarthritic cartilage, the amount of type VI collagen is increased23,75 and the collagen is distributed into a broader area—beyond the limits of the PCM.73,76 Furthermore, type VI collagen fibrils are disorganized. 23 The entire type VI collagen network in osteoarthritic cartilage is not as dense as in normal cartilage and is mixed with banded collagen fibrils. 77 A proteomic study on chondrons isolated from osteoarthritic cartilage identified multiple type VI collagen fragments. 40 In early osteoarthritis, “abnormal-looking” chondrocytes with multiple processes and presumably altered cell phenotypes are associated with disrupted pericellular distribution of type VI collagen. 78 Interleukin 1β (IL-1β), a prominent cytokine in cartilage degeneration, downregulates the expression of type VI collagen. 68 Not surprisingly, type VI collagen knock-out mice have increased susceptibility to osteoarthritis. 79
In tissue culture, the addition of matrilin-3 reversed chondrocyte catabolism—with increased production of type II collagen and aggrecan, and suppressed expression of MMP-13, ADAMTS-4 and 5 by chondrocytes. 80 However, the role of matrilins in the pathology of osteoarthritis is still controversial, as increased amounts of matrilin-3 in cartilage were found to positively correlate with the severity of osteoarthritis. 81 In another study, excessive matrilin-3 stimulated the proteolytic phenotype of chondrocytes, with increased expression of MMP-1, -3, and -13, and inflammatory cytokines. 82
The long-term culture of chondrons results in the formation of chondrocyte clusters and PCM remodeling, both of which are hallmarks of osteoarthritic cartilage. 83 Chondrons in long-term culture have been recommended as a model for investigation of the pathology of osteoarthritis in vitro. Using cultured chondrons, it has been found that chondrocytes gradually develop insensitivity or tolerance to IL-1β, 84 which may be significant to the progression of osteoarthritis.
The Role of PCM in Cartilage Engineering
Since articular cartilage cannot self-repair, tissue engineering has been a favorable strategy to restore the damaged or diseased cartilage. A classic approach of cartilage engineering applies isolated chondrocytes on a scaffold for cartilage regeneration. 85 In this process, PCM proteins are among the first to be expressed to stabilize the chondrocyte microenvironment. When chondrocytes were cultured in hydrogel, it was observed that type VI collagen, decorin, and fibronectin appeared as punctate structures on the surface of chondrocytes within 24 h. 86 Under these conditions, isolated chondrocytes required a week to rebuild the PCM.86,87 The expression of type VI collagen by chondrocytes was spiked in the first week of culture, a four-fold increase from normal cartilage. 87 After 1 week, type VI collagen formed a narrow pericellular ring around the chondrocytes. In the next 6–12 weeks of culture, these PCM molecules increased in density and distributed into a pericellular band about 2–4 μm in width (Fig. 3). 86 The major matrix collagens, such as types II, IX, and XI collagen, were only weakly stained intracellularly after 1 day but increased accumulation in the pericellular region as culture continued.
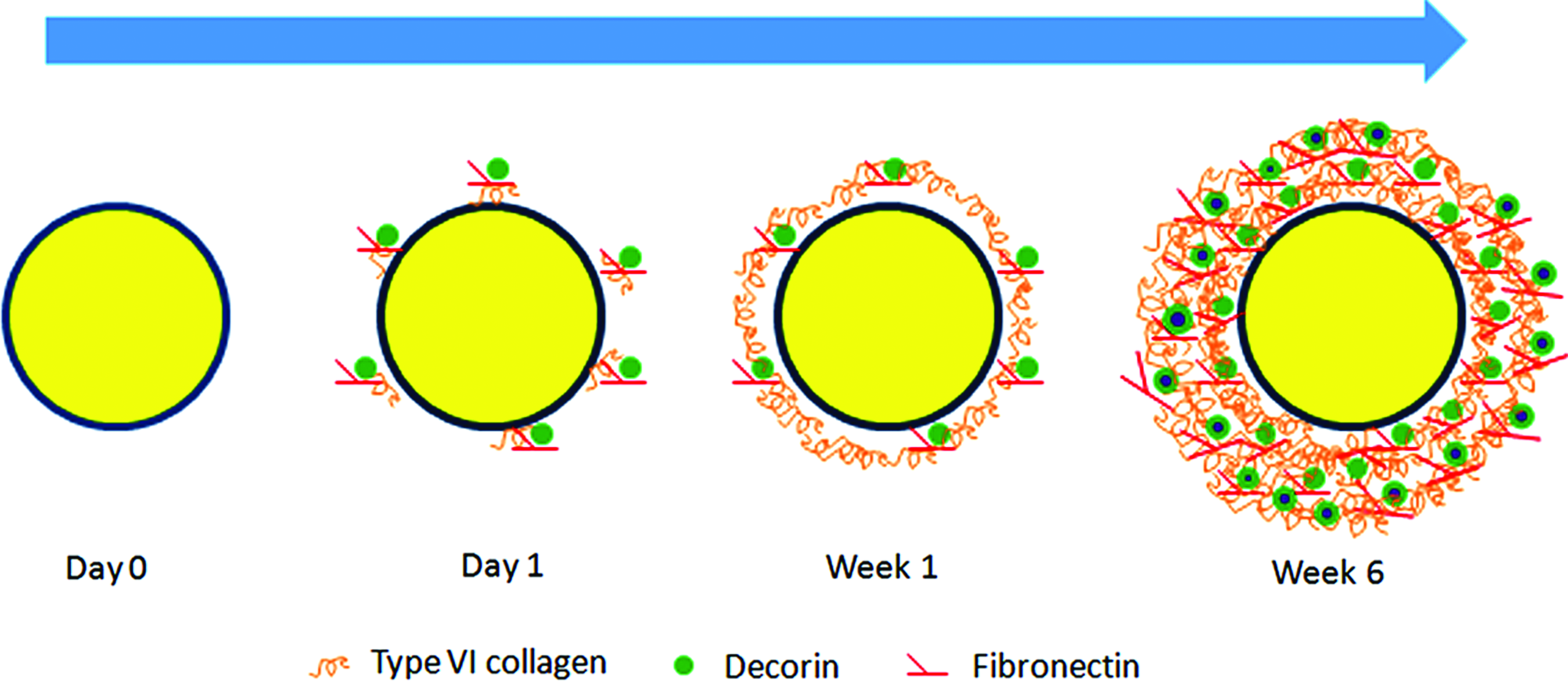
Rebuilding of the PCM of an isolated chondrocyte in tissue culture. The featured PCM components, such as type VI collagen and decorin, appear around the chondrocytes at day 1 of culture. Type VI collagen forms a narrow pericellular band at about a week. A much denser, complete PCM is rebuilt in 6–12 weeks. Color images available online at www.liebertpub.com/teb
By 12 weeks, type II collagen was deposited beyond the range of the PCM. Antibodies against keratin sulfate-rich region (5D4), chondroitin sulfate-rich region (2B6), core protein and hyaluronan-binding region (EFG-4) detected varied proteoglycan distribution patterns around the cultured chondrocytes. 86 After 12 weeks of culture, the deposition of aggrecan around the chondrocytes was divided into three discrete layers, which were about 2–4, 5–10, and >10 μm from the cell surface, by the antibody 5D4. While proteoglycans detected by the antibody 2B6 were mainly in the middle region, antibody EFG-4 detected proteoglycans in the two regions close to the chondrocytes. The dynamic processing and constant remodeling of the PCM, ultimately, result in this stratified proteoglycan distribution around the chondrocytes. In the rebuilt PCM, type VI collagen was co-localized with antibodies against keratin sulfate-rich region and hyaluronan-binding region of aggrecan.
The biology of cells, such as donor age, pathology and tissue type, and the properties of the biomaterials affect the formation of PCM during chondrogenesis and tissue engineering. For example, the PCM may be in different forms in different types of cartilage. Chondrocytes isolated from elastic auricular cartilage produced little type VI collagen, after being cultured in the form of aggregates for 3 weeks. 88 When articular and nasal chondrocytes were cultured on hyaluronic acid polymer, chondron structures exhibited in the areas of neo-cartilage mostly (>65%) made up of type II collagen and aggrecan, but not in the areas where there was predominantly type I collagen. 89 This suggests that a fully developed PCM is required for engineering hyaline/articular cartilage. Articular chondrocytes isolated from aged and osteoarthritic joints generally produce less type II collagen and aggrecan than chondrocytes from normal cartilage. They also tend to form flawed PCM with type VI collagen appearing both intracellularly and pericellularly. 89 In contrast, chondrocytes isolated from normal human and bovine articular cartilage form a typical chondron structure in tissue culture, as indicated by type VI collagen distribution. In tissue engineering, the deposition and distribution of type VI collagen in the PCM are also influenced by the types of scaffolds. The isolated articular chondrocytes formed chondron-like structures on nonwoven fleece made of esterified hyaluronic acid polymer (HYAFF 11) in tissue culture, but only spottily deposited type VI collagen on HYAFF 11 sponge. 89
Manipulating type VI collagen production toward rebuilding PCM could be a means of improving the quality of engineered cartilage. As shown in chondrons, the rebuilt PCM can stabilize the chondrocyte microenvironment, and a stable chondrocyte microenvironment likely contributes to the retention of the chondrocyte phenotype and production of cartilaginous matrix. Encapsulation of mesenchymal stem cells (MSCs) or chondrocytes with alginate adds an artificial microenvironment to the cells and stiffens the cell-gel structure. 90 However, when dynamic compression was applied, the encapsulated chondrocytes failed to increase matrix production in a manner comparative to isolated chondrons. 44 Neverthless, with composition optimization and material modification of the hydrogels91,92 a chondrocytic microenvironment that is comparable with the native PCM may be engineered.
The instability of chondrocyte phenotype in tissue culture is an obstacle faced by cartilage engineering.93–95 Dedifferentiated chondrocytes produce fibrocartilage, which prevents durable cartilage repair. 96 The chondrocyte phenotype is more complex than what was initially described. 97 At least 36 genes are differentially expressed by dedifferentiated chondrocytes. 98 Besides the traditional markers of types I and II collagen, types IX and XI collagen, several other matrix proteins, proteases, growth factors and cytokines also define, or are critically related to, the chondrocyte phenotype.99,100 While redifferentiation of chondrocytes has become a large undertaking of tissue engineering, 101 retention or minimizing the loss of the original chondrocyte phenotype may be another equally important approach to engineer hyaline cartilage. Compared with chondrocytes, chondrons have a distinct gene expression profile 41 and this unique phenotype translates to increased matrix production in culture. 42 After 1 week of culture, chondrons accumulated more type II collagen than chondrocytes. The cultured chondrons were able to retain more proteoglycans in the ECM than cultured chondrocytes. 42 Retention of matrix proteins, particularly proteoglycans, is essential for cartilage growth and the functionality of engineered cartilage. Furthermore, it seems that the presence of PCM influences the post-transcriptional modification of matrix molecules in the chondrons. Unlike chondrocytes, chondrons in tissue culture produce a collagen fibril network that resembles the ECM of native cartilage. 45 Probably owing to better retention and organization of matrix proteins, chondron pellets demonstrated greater mechanical properties than chondrocyte pellets. 102
Tissue engineering exposes cells to a foreign environment. Chondrons are more robust to withstand the environmental challenges than isolated chondrocytes. For cartilage engineering, chondrocytes are often embedded in poly(ethylene glycol) (PEG), with gelation induced by photopolymerization. 103 The procedure increases reactive oxygen species and membrane lipid peroxidation of the embedded chondrocytes. Using the same protocol, when chondrons were embedded in PEG hydrogel, oxidative stress was significantly reduced. 104 Manipulation of tissue culture (engineering) conditions, such as buffer systems, 105 serum, and osmolarity, 106 is a common approach to improve chondrocyte survival and functionality. However, the re-establishment of the PCM appears to best promote chondrocyte survival and matrix production in cartilage engineering applications. 106
MSCs are capable of chondrogenic differentiation and are a promising cell resource for cartilage tissue engineering or cartilage repair. 107 During chondrogenic differentiation, MSCs build a complete chondrogenic PCM that includes type VI collagen and decorin within 2 weeks.108,109 It has been found that the PCM, but not cytoskeleton, supported MSC-derived chondrocytes against mechanical stress which causes deformation. 109 Interestingly, basement membrane proteins such as laminin and type IV collagen, which have been found in the chondrocyte PCM, 31 go through a series of spatiotemporal redistributions during MSC chondrogenic differentiation and, eventually, are located pericellularly. 110 The biological significance of the pericellular distribution of basement membrane proteins during chondrogenic differentiation is still unclear. A useful note for cartilage engineering is that the formation of PCM during MSC chondrogenesis can be adjusted by the properties of the applied scaffold and the density of the seeded MSCs. A high polymer cross-linking density increases the pericellular distribution of cartilaginous matrix. A study has found that, at a seeding density of 1×107/mL, MSCs produced limited collagen (type II) fibrogenesis in the pericellular region. 111 Fibril formation of type II collagen was extended into a broad area of ECM, when MSCs were seeded at a density of 5×107/mL.
Among many challenges to use MSCs for cartilage repair is the efficiency of their chondrogenic differentiation. As in cartilage biology, the PCM is at the center of signal exchanges between MSCs and the environment during MSC differentiation. When MSCs and chondrocytes were embedded in agarose gel and cultured in regular medium used for chondrocyte culture, the PCM around MSCs was poorly formed. 112 This PCM lacked type VI collagen, but contained more decorin, biglycan, and fibronectin than the typical chondrocyte PCM. For the purpose of cartilage engineering, the conditions for MSC chondrogenic differentiation can be fabricated in many ways. It has been known for a while that the mechanical property of the matrix alone can guide MSC differentiation: A softer matrix promotes adipogenic, neurogenic, and chondrogenic differentiation and a stiffer matrix supports myogenic and osteogenic differentiation.113,114 MSCs form a denser/stiffer PCM in the presence of a denser/stiffer matrix. 114 The exact mechanism behind this observation is not entirely known. It is, however, known that the integrin-cytoskeleton complex formed in the PCM influences MSC phenotypes. In one chondrogenic differentiation system, blocking the interaction between integrin and PCM effectively interrupted cytoskeleton and was able to cancel the effects of hydrostatic pressure and matrix stiffness on MSC chondrogenic differentiation. 114
The chondrocyte PCM is itself an inducer of chondrogenic differentiation. When MSCs and chondrons were cultured together at a ratio of 9:1, the amount of glycosaminoglycans in the matrix was twice that of MSCs co-cultured with chondrocytes. 115 A similar result was observed when MSCs and chondrons were mixed and implanted subcutaneously in vivo. After 6 months of implantation into chondral defects in goat knees, the mixed MSCs and chondrons produced better cartilage repair both macroscopically and microscopically than the control group, for which microfracture procedures were performed to introduce bone marrow MSCs into the chondral defects.
Not only does the entire PCM but also individual PCM molecules influence the chondrogenic differentiation of MSCs. For example, the overexpression of matrilin-1 and -3 enhanced chondrogenesis of MSCs derived from synovium, as assessed by gene expression of type II collagen and aggrecan, and the production of proteoglycans. 116 However, matrilins alone were unable to induce or initiate chondrogenic differentiation of MSCs. The effects of matrilins on the chondrogenesis of MSCs require the presence of TGFβ. It is most likely that matrilins facilitate cell adhesion, via an α5β1 integrin-dependent mechanism and Akt phosphorylation, and this improves ECM synthesis and accumulation. 117 In support of a role for the TGFβ family in chondrogenesis, the binding between perlecan and BMP-2 was utilized in a study for the design of a cartilage engineering scaffold. 48 Incorporated with perlecan domain I, the scaffold released BMP-2 in a linear kinetics, leading to improved chondrogenic differentiation of MSCs and abundant matrix production.
In summary, the use of isolated chondrons as a model for studying the PCM and chondrocyte microenvironment has enriched our understanding of cartilage biology. Implementation and refinement of cartilage engineering protocols with emphases on the microenvironment of chondrocytes/MSCs and the regulation of PCM could lead to the production of more robust and functional cartilage.
Footnotes
Acknowledgments
This review is supported in part by a grant provided by Saint Louis University and Dr. Lew Schon Innovation Fund. The author thanks Dr. Jan Ryerse, the Research Microscopy & Histology Core, Saint Louis University, for assistance in imaging and Dr. Joseph Tran, Department of Embryology—Carnegie Institution for Science, for assistance in preparation of this article.
Disclosure Statement
No competing financial interests exist.