
Abstract
In the field of regenerative medicine, creating a biomaterial device with the potential alone to affect cellular fate is a desirable translational strategy. Native tissues and growth factors are attractive candidates to provide desired signals in a biomaterial environment. However, these molecules can have translational challenges such as high cost, complicated regulatory pathways, and/or limitations with reproducibility. In regenerative medicine, there is a burgeoning community of investigators who seek to overcome these challenges by introducing synthetic peptides to mimic the desirable signals provided by growth factors and tissue matrices. Since in cartilage tissue, cell-adhesion signaling mediates cell migration, growth, and differentiation, synthetic peptides that mimic a desired cell-adhesion sequence may help to control cellular fate. This review emphasizes the value of the signaling ability of peptides, specifically in the cartilage regeneration field. The primary challenge in cartilage regeneration is to regenerate true hyaline cartilage instead of a fibrous tissue. The vision is to create materials that take advantage of the signaling abilities of peptides and that themselves induce chondrogenesis without the need for tissue-derived matrix or growth factors, which could potentially revolutionize arthritis prevention and treatment.
Introduction
The center of Disease Control and Prediction estimated that 78 million adults aged >18 years in the United States (26% of the population) will be diagnosed by arthritis by year 2040. Despite arthritis being one of the leading causes of disability, it does not yet have an effective treatment and may develop following a cartilage injury. There are surgical treatments that aim to repair cartilage tissue such as autologous chondrocyte implantation, microfracture, osteochondral transplantation (mosaicplasty), and allograft implants. Although these treatments may provide clinical improvement, they are generally not successful in producing a tissue with the same long-term mechanical and chemical properties of native articular cartilage. The deficiency of the treatments to produce effective cartilage tissue has created an impetus for the regenerative medicine community to establish strategies that lead to restoration of fully functional hyaline cartilage. 1
One approach to the design of bioactive material is the incorporation of natural components such as extracellular matrix (ECM) molecules or growth factors in a scaffold structure. Although natural components might be able to mimic the native environment for the cells and enhance cartilage formation, disadvantages such as high cost, limitations of reproducibility, and the risks involved with immunogenicity can be significant limitations for commercialization. Therefore, a synthetic product with the potential of inducing cartilage regeneration without the need for human- or animal-derived elements (if indeed such a material exists) would present a more attractive translational option. Synthetic biomaterials may be utilized for acellular applications where the cartilage regeneration does not rely on cell-based therapies,2–4 although there are applications where a cell-based strategy may be required, for example in regeneration of the entire joint surface,5,6 and a chondroinductive biomaterial would be advantageous in either case. In cartilage regeneration, the goal of designing an all-synthetic biomaterial is to create scaffolds with desired mechanical integrity and the ability to lead endogenous mesenchymal stem cells (MSCs) toward chondrogenic differentiation and prevent chondrocyte dedifferentiation. The properties of synthetic biomaterials may be tailored by changing the polymer composition to achieve desirable failure properties (e.g., maximum stress, strain, and toughness). 7 However, the remaining challenge is incorporating all-synthetic cellular signals to replace ECM molecules and growth factors to lead endogenous stem cells to chondrogenic differentiation and prevent chondrocyte dedifferentiation.
Peptides have the potential to mimic ligands and act through desired cellular signaling pathways. Identifying the bioactive peptides that are chondroinductive and then incorporating them in a biomaterial may lead to 100% synthetic chondroinductive biomaterials, which may potentially revolutionize the field of cartilage injury treatment.
Perhaps the regenerative medicine community may take a cue from the cancer field, as identifying receptor-binding peptides is a crucial research focus for oncological therapy. Delivery of a therapeutic with receptor-binding peptides decreases the side effects of the therapeutic compared to chemotherapeutic methods, in which the drugs are given to the patients at concentrations approaching maximum body tolerance with low efficiency. 1 The regenerative medicine community may likewise design synthetic bioactive materials by incorporating receptor-binding peptides in the material structure to take advantage of their signaling abilities.
In regenerative medicine, there are a select group of pioneering studies that have employed peptides as direct signals.8–13 In addition, there are review papers that have eloquently covered receptor-binding peptides.1,14 However, they have not focused on peptide applications for regenerative medicine or the approaches of identifying peptides that may help with cellular differentiation.
The current review provides an overview of cartilage regeneration studies that have taken advantage of bioactive peptides for their potential of affecting cellular fate. Furthermore, cell–matrix adhesion, adhesion receptors, and methods of identifying adhesion receptor-binding peptides are emphasized as potential strategies to affect cellular differentiation and induce chondrogenic differentiation.
Potential Chondroinductive Peptides in Cartilage Regeneration
The most common methods of incorporating receptor-binding peptides in biomaterials are using peptides as a coating for scaffold surfaces and conjugating the peptides to polymers in scaffolds. Incorporating small peptides in biomaterials for regenerative medicine is attractive because short peptide sequences can be synthesized reproducibly in large quantities, their small size reduces the chance of nonspecific binding, and they have the potential to affect cellular fate. Arginine-glycine-aspartate (RGD) peptides are attractive because of their cell-adhesion properties, and as a result, RGD is the most commonly used receptor-binding peptide across a variety of applications. However, there are only a few peptides that have been employed in the cartilage regeneration field for their potential capacity to induce chondrogenic differentiation. Below, a concise overview of each of these studies is given.
Link N peptide
Link protein is a glycoprotein that stabilizes the noncovalent interaction of hyaluronate and aggrecan molecule G1 domain 15 and the Link N sequence (DHLSDNYTLDHDRAIH) is the amino-terminal peptide of the link protein. In 2013, Wang et al. 16 indicated that the Link N sequence acts through the BMP type II receptor. Several studies have investigated the potential of Link N sequence in improving cartilage tissue regeneration both in vitro and in vivo. 17
In 2003, Mwale et al. showed that adding 100 ng/mL Link N to the culture medium of both nucleus pulposus (NP) and annulus fibrous (AF) cells isolated from bovine intervertebral discs (IVDs) every 2 days increased the glycosaminoglycan (GAG) and collagen type II and type IX accumulation after 20 days. Type II collagen and type IX collagen contents were measured by enzyme-linked immunosorbent assay, and GAG content was measured by the dimethylmethylene blue assay. 15 In another study, Gawri et al. reported that injecting 50 μL Link N at a concentration of 20 mg/mL into healthy intact IVDs isolated from 13 adult human donors promoted aggrecan synthesis compared to the control group (50 μL 35 SO4). Aggrecan synthesis was assayed by estimating 35 SO4 incorporation in intact IVDs after 48 h. 18 Injecting Link N at a concentration of 10 mg/mL into 5 mm deep annular punctures in the center of NP of 19 New Zealand White rabbit IVDs significantly increased the aggrecan expression for both NP and AF after 2 weeks. 19
GFOGER peptide
The GFOGERGVEG-POGPA peptide sequence was identified for its ability to bind the α2β1 integrin. The peptide was recognized by comparing the binding abilities of overlapping peptides from collagen I, α1(I)CB3 domain to the α2β1 receptor.20,21 In 2000, Knight et al. 22 reported that removal of the GER sequence from GFOGERGVEG-POGPA sequence stopped the peptide from binding to α2β1. However, the removal of its C-terminal had no evident effect on the level of the peptide adhesion to α2β1 integrin. This study further indicated the ability of the GFOGER peptide to bind to the α2β1 receptor by demonstrating that the adhesion of collagen I to the receptor was inhibited by GFOGER.
In 2010, Liu et al. 23 incorporated GFOGER into polyethylene glycol (PEG) hydrogels through Michael addition chemistry. Human MSCs were encapsulated in PEG hydrogels with and without GFOGER. After 9 days, the relative gene expressions of collagen type II and type X and aggrecan were higher in the groups with the GFOGER peptide.
B2A2 peptide
The B2A2 peptide consists of a receptor targeting domain, a hydrophobic spacer domain, and a heparin-binding domain. The peptide was designed in 1999 to bind to heparin and endothelial cell proteoglycans. 24 In 2005, the peptide was recognized for its ability to bind to the BMP-2 receptor.25,26 Since BMP receptors are involved in chondrocyte formation, in 2012, Lin et al. 27 hypothesized that the B2A2-K-NS peptide could play a role in cartilage repair. To test their hypothesis, they added 10 μg/mL of the peptide to murine embryonic stem cell medium every 3 days. After 7 days, gene expression of SOX9 and collagen type II increased compared to the untreated group (i.e., medium without the peptide). To identify the effects of the peptide on cartilage regeneration in vivo, osteoarthritis was chemically induced in the knees of six adult rats by injecting monoiodoacetate into the synovial space. On days 7 and 14, 500 ng of the peptide was injected into the knees. After 21 days, the histological analyses indicated that the B2A2 treatment enhanced cartilage repair compared to untreated knees, which received saline instead of the peptide.
KIPKASSVPTELSAISTLYL peptide
The KIPKASSVPTELSAISTLYL sequence, which is known as the BMP2 peptide, corresponds to residues 73–92 of the knuckle epitope of bone morphogenetic protein-2 (BMP-2). This peptide was identified in 2003 as a potential candidate to improve bone formation through raising alkaline phosphatase activity compared to the other overlapping peptides of knuckle epitope of BMP-2.28–30 In 2012, the ability of the peptide to direct chondrogenesis was investigated when it was added to the human MSC medium (100 ng/mL) three times per week. The gene expressions for aggrecan (after 3 days) and SOX9 and collagen type II (both after 1 week) were significantly higher than the negative control (no growth factor in the medium). 31
N-cadherin mimetic peptide
Cell–cell adhesion plays an important role in initiating chondrogenesis. In several studies, peptides that mimic cell–cell interactions have been used as a strategy for designing chondroinductive biomaterials.32,33 In 2012, it was shown that functionalization of hyaluronic acid (HA) hydrogels with N-cadherin mimetic peptides increased GAG and total collagen content of encapsulated MSCs after 28 days compared to MSCs encapsulated in HA hydrogels functionalized with scrambled N-cadherin mimetic peptides.34,35
Self-assembled peptides
Several studies investigated the potential of self-assembled peptide hydrogels in modulating chondrogenesis. Self-assembled peptide hydrogels have been the focus of regenerative medicine applications due to their favorable physical properties. These hydrogels are made from natural building blocks, and they may provide safe degradation products. Their physical properties such as pore size, fiber thickness, and mechanical performance can be tuned, and may improve a material's biological properties if biologically relevant peptides are used.36–44 In the cartilage regeneration field, it was shown that encapsulating MSCs for 21 days in self-assembled peptides resulted in higher GAG content and more spatially uniform proteoglycan and collagen type II deposition compared to MSCs encapsulated in agarose hydrogels.45,46
Summary
The potential for chondroinductivity of bioactive peptides was shown in several studies, which indicates that synthetic peptides may have the potential to duplicate the cell-adhesion site of ECM molecules. The peptides reviewed here have shown promising results for improving chondrogenic differentiation, although more investigation is required to determine peptide specificity for chondroinductivity.
For instance, BMP receptors are not only involved in chondrogenic differentiation, but also naturally play a role in osteogenic differentiation. The B2A2 peptide was designed to bind to BMP II receptor, and likewise the ability of Link N to bind to the BMP II receptor was demonstrated in 2013. Although it was shown that both peptides induce chondrogenic differentiation, evaluation of these peptides for inducing osteogenic differentiation is essential due to the role of the BMP II receptor in bone formation. Regarding GFOGER, it was shown that the peptide inhibits collagen I binding to the α2β1 receptor. This indicates that GFOGER may mimic the binding site of collagen I to the receptor, which is not favorable for chondrogenic differentiation, and thus further investigation is recommended to measure collagen I production in the presence of the peptide. KIPKASSVPTELSAISTLYL's ability to improve bone formation through raising alkaline phosphatase activity was shown in 2003, which would not make the peptide an ideal candidate for designing materials to induce only chondrogenesis.
Cell–Matrix Adhesion and Chondrogenic Differentiation
It is widely known that adhesion molecules are crucial in mediating multiple cellular signaling pathways,47,48 and have a high impact on critical cellular processes such as gene expression, cell cycle, and programmed cell death. 49
Key components of cell adhesion can be divided into three main groups: adhesion receptors, ECM proteins, and cytoplasmic plaque membrane proteins. The adhesion receptors have a critical role in mediating cell–cell and cell–ECM signaling, and they are typically classified using four groups: integrins, cadherins, immunoglobulins, and selectins. These receptors have been the subject of significant interest, as reflected by the thousands of publications that describe them in detail.49,50 This review provides only a brief overview, given that more extensive descriptions are available elsewhere.
Adhesion receptors
Integrins are known as cell receptors for ECM proteins and mediate cell–ECM adhesion. The structure of each integrin consists of an α and a β subunit, with both subunits composed of three main domains: an extracellular domain, a cytoplasmic region, and a single membrane-spanning domain. There are at least 16 α and 8 β subunits known for integrins, and the specific subunit pairing determines explicit ligand–integrin binding properties.55–53 Cadherins mediate cell–cell adherence junctions and communicate with a group of linking proteins called catenins, which are crucial for cadherin function. Cadherins have different types of N-, P-, R-, B-, and E-cadherins, all with essential roles in both tissue formation and signaling cascade regulation.54–58 Immunoglobulin cell-adhesion molecules are a member of the immunoglobulin superfamily, and studies show that some are critically engaged in T- and B-cell activation.59,60 Selectins are smaller families of adhesion receptors that mediate rolling interactions of leukocytes on vascular endothelial cells, and therefore play a critical role in controlling inflammatory diseases.61–64
Adhesion receptors and cartilage regeneration
Cartilage has an ECM-rich environment with a limited number of chondrocytes for which the value of ECM as a survival factor has been demonstrated.65,66 Integrins therefore play a critical role in cartilage regeneration because integrins mediate cell–ECM signaling pathways. To design chondroinductive biomaterials, it is valuable to identify the integrins expressed by chondrocytes and the changes of integrin expression during chondrogenic differentiation. Incorporating integrin-specific sequences in biomaterials may help to control the cellular differentiation better through activating desired adhesion signaling pathways. This review covers the studies that focused on identifying chondrocyte-specific integrins and the studies that aimed to monitor the integrin expression modifications during chondrogenic differentiation of stem cells.
Integrin expression of chondrocytes
In 1994, Woods et al. 67 investigated the integrins expressed by chondrocytes of human healthy knee cartilage. Cartilage tissue was harvested with biopsies from adult donors, and the chondrocytes were isolated from the tissue. The integrin expression of the cells was analyzed by immunohistochemistry using monoclonal antibodies against different integrins, which indicated that normal human articular chondrocytes display a substantial quantity of α1β1, α5β1, and αvβ5 integrins, as well as limited quantities of α3β1 and αvβ3 integrins. The expressions of α1β1, α3β1, and α5β1 were highlighted in 1995 by Loeser et al. 68 by immunostaining chondrocytes harvested from bovine articular cartilage (Fig. 1).
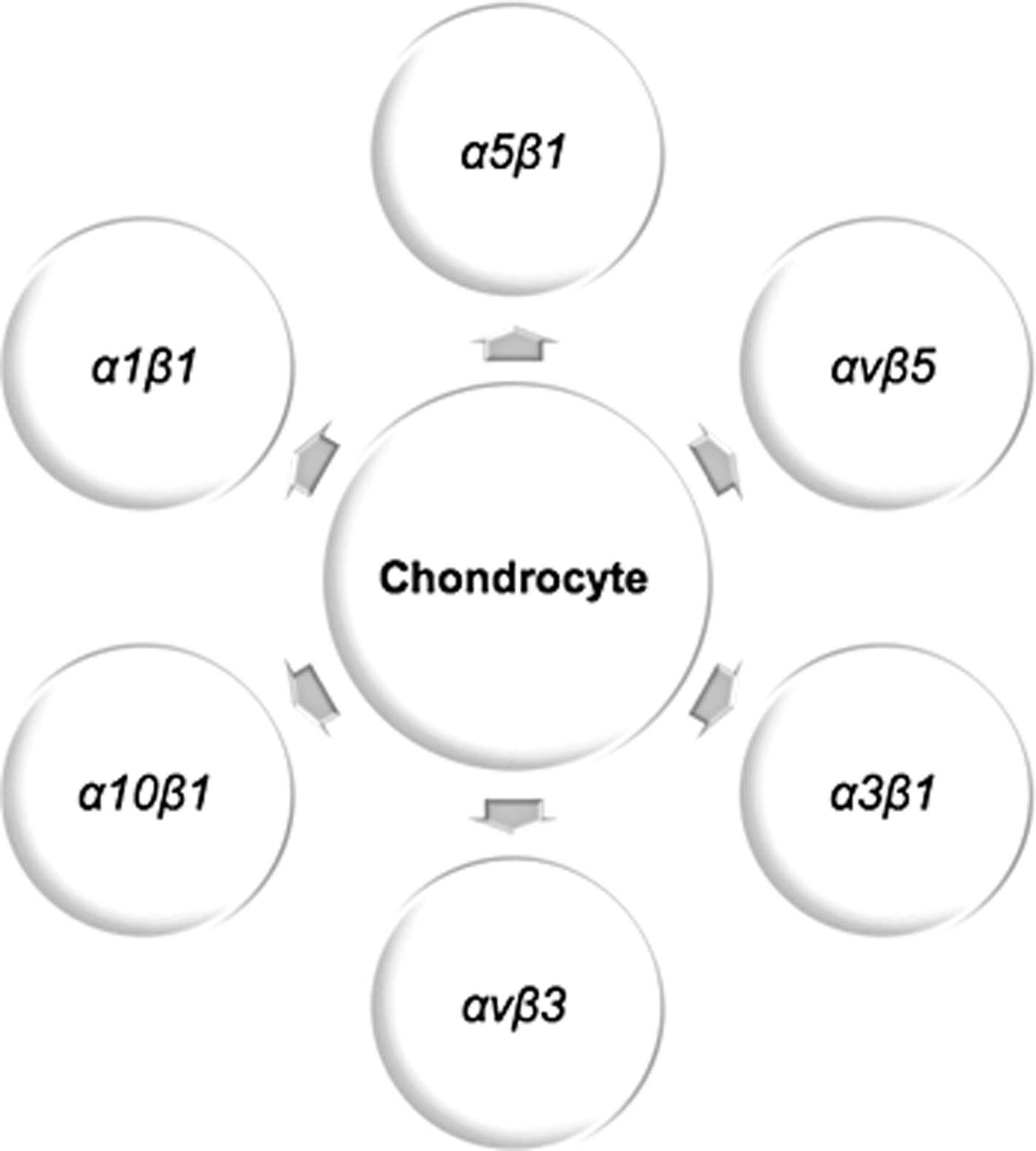
Chondrocyte integrin expression.
In 1998, Camper et al. 69 described α10β1 expression by human chondrocytes via immunostaining of chondrocytes in tissue sections of human cartilage tissue. In 1995, it was shown that the integrin expression of fetal chondrocytes is different from that of adult chondrocytes. 70 The immunostaining of fetal healthy knee sections demonstrated that fetal chondrocytes expressed α6 and α2, which are not expressed by adult chondrocytes. In addition, the fetal chondrocytes did not express β3 and α3, which were expressed by adult chondrocytes in previous studies.
Integrin expression during chondrogenic differentiation
The variation of integrin expression during chondrogenic differentiation was investigated for the first time in 2005 by Goessler et al. 71 In this study, the integrin expression of human bone marrow MSCs was analyzed with microarray hybridization. During the 21 days of chondrogenic differentiation, the expression of integrin α5β1 was downregulated, while the expression of other integrins remained constant. In 2006, the important role of the α5 integrin subunit was further emphasized by Connelly et al. 72 when anti-α5 antibody was added to the medium of bone marrow MSCs harvested from the tibiae and femora of an immature calf, producing a reduction in GAG accumulation after 6 days relative to the untreated control group. In 2007, Goessler et al. 73 studied the integrin expression of both bone marrow and adipose-derived human MSCs before and 20 days after chondrogenic differentiation. Microarrays and immunohistochemistry indicated that the integrin α5β1 was expressed by undifferentiated MSCs, and its expression rose during chondrogenic differentiation of both types of MSCs. A 2013 study indicated the importance of the integrin β8 subunit during chondrogenic differentiation: β8 expression was upregulated after 21 days of chondrogenic differentiation of human MCSs when measured by quantitative polymerase chain reaction. The knockdown of the β8 receptor via a shRNA lentiviral construct resulted in no collagen type II expression, which further supported the importance of the β8 receptor in chondrogenic differentiation. 74
In summary, the important role of adhesion molecules in regulating cellular signaling is widely recognized. The roles of integrins in cartilage tissue and chondrocyte signaling are of special importance, given the ECM-rich nature of the tissue and the role of integrins in mediating cell–ECM signaling, as cartilage has an ECM-rich environment with low cell density. The integrins that affect cartilage regeneration are typically categorized into two main groups; the integrins expressed by differentiated chondrocytes, and the integrins expressed during chondrogenic differentiation.
For the biomaterials that target chondrocytes, utilizing ligands specific to integrins of chondrocytes may possibly prevent chondrocyte dedifferentiation and contribute in mediating the integrity of cartilage tissue. For the biomaterials that target MSCs for chondroinductivity, identifying and incorporating ligands specific to the integrins that change expression during chondrogenic differentiation may further improve cartilage tissue development. This approach can therefore produce biomaterials with the capability to activate cartilage regeneration.
The current studies in the field of cartilage biology may reveal valuable information about the chondrogenic role of integrins, but more extensive studies are required to elucidate the contributions of a wider range of integrins and to identify the specific role of each integrin during chondrogenic differentiation.
Adhesion Receptor-Binding Peptides
In 1984, Pierschbacher and Ruoslahti 75 indicated that the cell-binding potential of fibronectin can be duplicated with the cell attachment domain of fibronectin, a peptide sequence with three amino acids (Arg-Gly-Asp, i.e., RGD). RGD was subsequently recognized for the ability to bind to 12 integrins.76–80 The fact that RGD is the cell attachment site to many other adhesion proteins has given this peptide a distinguished position in cell-adhesion biology, and numerous regenerative medicine and pharmaceutical applications are taking advantage of this sequence to design cell-adhesion materials. 81 The RGD example underscores the notion that the binding site of a receptor can indeed be recapitulated by a relatively short peptide sequence, which can be used instead of growth factors, antibodies, and ECM proteins.82–85 Replacing entire adhesion molecules with a short adhesion peptides is attractive because such peptides are easy to synthesize in large quantities. Moreover, their small size reduces the chance of any nonspecific binding, and they can be designed to obtain a desired cell response. It is important to keep in mind that several distinct proteins can bind to an individual receptor. 86 For instance, the αvβ3 integrin has been reported to bind to collagen VI, laminin, fibronectin, vitronectin, thrombospondin, von Willebrand factor, and fibrinogen. 87
In addition, a peptide that binds to a desired integrin may be unable to target a specific pathway and even inhibit the binding of the other suitable sequences. In 2006, Connelly et al. 72 demonstrated the inhibition of chondrogenesis in RGD-modified alginate gels. In that study, bovine bone marrow–derived MSCs were encapsulated in either RGD- or RGE-modified alginate gels. After 7 days, the chondrogenic medium significantly stimulated chondrocytic gene expression (i.e., collagen II, aggrecan, and SOX9) in the RGE- but not in the RGD-modified gels. The results indicated that RGD interactions significantly inhibit the chondrogenic response in terms of gene expression for this particular system.
In cancer therapy, the limitations of current chemotherapeutics make identification of adhesion receptor-binding peptides an attractive research topic. In chemotherapeutic procedures, the drugs are typically not of maximal efficiency and are given to the patients at the highest level of body tolerance. Identifying peptides that bind to adhesion receptors of cells, and incorporating them in cancer therapy drugs, may reduce side effects, as the therapeutic would only target the tumor cells. Table 1 shows a selected group of adhesion receptor-binding peptides that have been discovered for cancer therapy applications. 1
Integrin-Binding Peptides Identified for Cancer Therapy Applications
OBOC, one-bead one-compound.
In summary, the success of the cancer therapy field in identifying peptides that bind to specific adhesion receptors indicates opportunities for the regenerative medicine community to discover peptides that induce or facilitate chondroinductivity by targeting integrins. In addition, the extensive research surrounding the RGD sequence indicates the potential of peptides to affect cellular fate. Although adhesion receptor-binding peptides have the potential to change cellular fate, it is important to keep in mind that the ability of a peptide to bind to the target integrin may not always lead to activating the desired signaling pathways and may even inhibit the preferred cellular activities. However, an adhesion peptide sequence may hold tremendous value for retaining endogenous cells in the construct in vivo, and may play a crucial role in a combined strategy with a differentiation peptide.
Methods of Identifying Adhesion Receptor-Binding Peptides
If the regenerative medicine community is to pursue the recognition of novel peptide sequences to create desirable cell responses, then eventually the methods of identifying them must be understood. Investigators have applied many different methods for discovering adhesion receptor-binding peptides, from synthesizing overlapping or random sequences of an adhesion protein to using complex peptide libraries.
Peptide libraries provide the opportunity to screen the binding ability of a given protein in a high-throughput manner. Peptide libraries are categorized based on their method of display, and can be categorized into two main groups: biological and nonbiological. Biological libraries use DNA, or genotype, to encode peptide sequences, 1 whereas nonbiological libraries use peptides synthesized in vitro. To screen the binding ability of a desired protein or cell line, it is incubated along with the peptide library, and unbound peptides are subsequently washed away. The remaining peptides are then screened according to their design method. Biological libraries include bacterial, bacteriophage, ribosome, mRNA, yeast, cDNA, retrovirus, baculovirus, and mammalian cell display. Bacterial and phage libraries are applied for adhesion receptor-binding peptide isolation. In the bacterial libraries, peptides are genetically connected to the membrane flagella and fimbriae proteins and are displayed on the surface of bacteria. The DNA of the isolated bacterial clone is sequenced to reveal the identity of the adhesion receptor-binding peptide.88,89 In the phage display libraries, the DNA sequence of a desired peptide is embedded into the DNA of a phage coat protein to encode the peptide. After the incubation process, the DNA of the resulting phage is sequenced to discover its peptide content.90,91 Positional scanning synthetic peptide combinatorial libraries (PS-SPCLs) and one-bead one-compound (OBOC) libraries are the popular nonbiological libraries for recognition of adhesion receptor-binding peptides. In OBOC libraries, the peptides are synthesized on beads that are approximately 80–100 μm in diameter; the split mix synthesis method generates the library, and after incubation of the protein of interest and washing away of the unbound proteins, mass spectrometry or Edman's sequencing determines the peptide content of adhesion receptor-binding beads.92,93 In PS-SPCLs, the peptides are synthesized individually. To synthesize individual peptides, one amino acid is held fixed while the remaining amino acids of the peptide vary. Holding a different amino acid constant and alternating others creates combinational libraries. There are various screening methods for PS-SPCLs such as using specific cellular effects as the readout or fluorescent tagging. 94 The detailed explanation of each peptide library could be the focus of an entire review, and indeed there are many excellent reviews about them that we recommend.88,89,92–100 These methods have been utilized to identify adhesion receptor-binding peptides for other applications than regenerative medicine. For example, Table 1 shows a selected group of peptides from the field of cancer therapy, identified by panning in vivo, OBOC, and phage display libraries, and indicates the feasibility of the proposed methods for identifying receptor-binding peptides.
In summary, the methods of identifying adhesion receptor-binding peptides may be an excellent start to recognize bioactive sequences, although further investigation is required to confirm the ability of the sequence to mimic the desired binding and to activate the target signaling pathways.
Discussion
Chondroinductive biomaterials are categorized into two broad groups: natural and synthetic. Natural materials are attractive candidates to be incorporated in scaffold structure because they have the potential to communicate with cells not only through chemical signaling but additionally through mechanical signaling, which is beneficial to tissue-dependent cells. The regenerative properties of natural materials may be attractive for biomaterial design, but their disadvantages, such as reproducibility and the risks involved with immunogenicity, may complicate regulatory pathways. As a result, natural materials may be less attractive for commercialization if 100% synthetic materials are capable of eliciting a comparable biological response, which indeed makes designing all-synthetic biomaterials a more appealing translational strategy.
The main challenge in designing an all-synthetic biomaterial is to identify chemical signals that mimic the adhesion receptor-binding site of ECM or growth factors and lead the stem cells to the desired lineage. In the case of cartilage regeneration, cell–ECM signaling is crucial for both chondrocyte survival and chondrogenic differentiation. Cell–ECM signaling is mediated by ECM interactions with adhesion receptors, namely integrins. Therefore, the integrins expressed by chondrocytes and the integrins that change expression during chondrogenic differentiation may play a role in mediating cartilage tissue development through interaction with ECM. As a result, the ligands specific to these integrins may have the potential to provide chemical signals for the cells and to produce chondroinductivity. Bioactive peptides are attractive candidates to mimic integrin–ligand binding because of favorable properties such as reproducibility and low cost.
The integrin-binding peptides, such as RGD, are mainly employed in biomaterials for their cell-adhesion properties. In the field of cartilage regeneration, a few studies have employed bioactive peptides for their signaling abilities, although none have employed strategies to identify these peptides based on their ability to bind to chondrocyte integrins or integrins expressed during chondrogenic differentiation.
The cartilage regeneration community may take advantage of the literature regarding integrins that influence this process, and identify peptides that have the potential to bind to these receptors. The adhesion receptor-binding peptides described in the literature are likewise attractive candidates. Fortunately, the methods of identifying adhesion receptor-binding peptides are well described in the literature. Although further studies would be required to investigate cell–cell and cell–ECM interactions during chondrogenic differentiation, and the widespread effects of adhesion receptor-binding peptides on cellular fate, the latter may unlock the potential to activate desired signaling pathways and induce chondrogenic differentiation. A future strategy in vivo may be to combine both adhesion sequences to retain endogenous stem cells in the construct, with chondroinductive sequences, to drive differentiation to lead to hyaline cartilage regeneration.
In addition to adhesion receptor-binding peptides, identifying additional signaling pathways that contribute to chondrogenic differentiation and incorporating peptides that mimic key interactions would be necessary to design a biomaterial tailored to known receptor–peptide interactions. This approach could yield the next generation of bioactive biomaterials for cartilage regeneration. Identifying signaling pathways that contribute to chondrogenic differentiation of MSCs is a growing subject of research in cartilage biology. Although there are many ongoing studies on this subject, more investigation is required to understand fully the mechanisms involved in chondrogenic differentiation. In addition to the adhesion receptors that represent the focus of this review, the regenerative medicine community may focus on other known signaling pathways for chondrogenic differentiation and design peptides that activate them. Signaling processes such as the trophic effects of MSCs or canonical Wnt pathways have been shown to increase chondrocyte proliferation and matrix formation.101,102 A powerful future strategy may be to combine sequences from various signaling pathways to retain endogenous stem cells in the construct, along with chondroinductive sequences to drive differentiation and regenerate hyaline cartilage.
Footnotes
Disclosure Statement
No competing financial interests exist.